
Biorefinery-Based Approach to Exploit Mixed Cultures of Lipomyces starkeyi and Chloroidium saccharophilum for Single Cell Oil Production

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Pre-Culture Media
2.2. Culture Media and Operating Conditions
2.3. Analytical Methods
2.4. Parameter Analysis
3. Results and Discussion
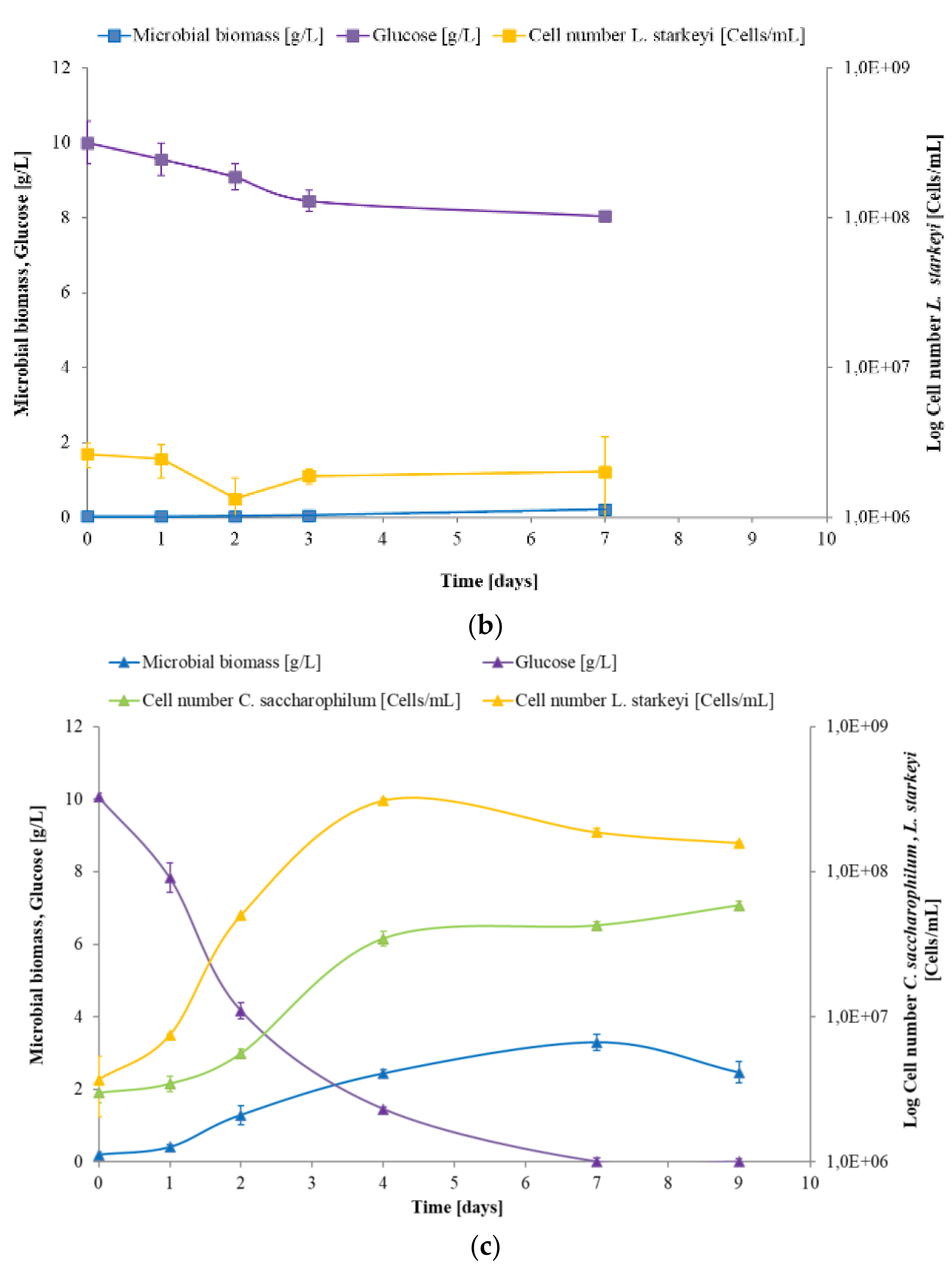
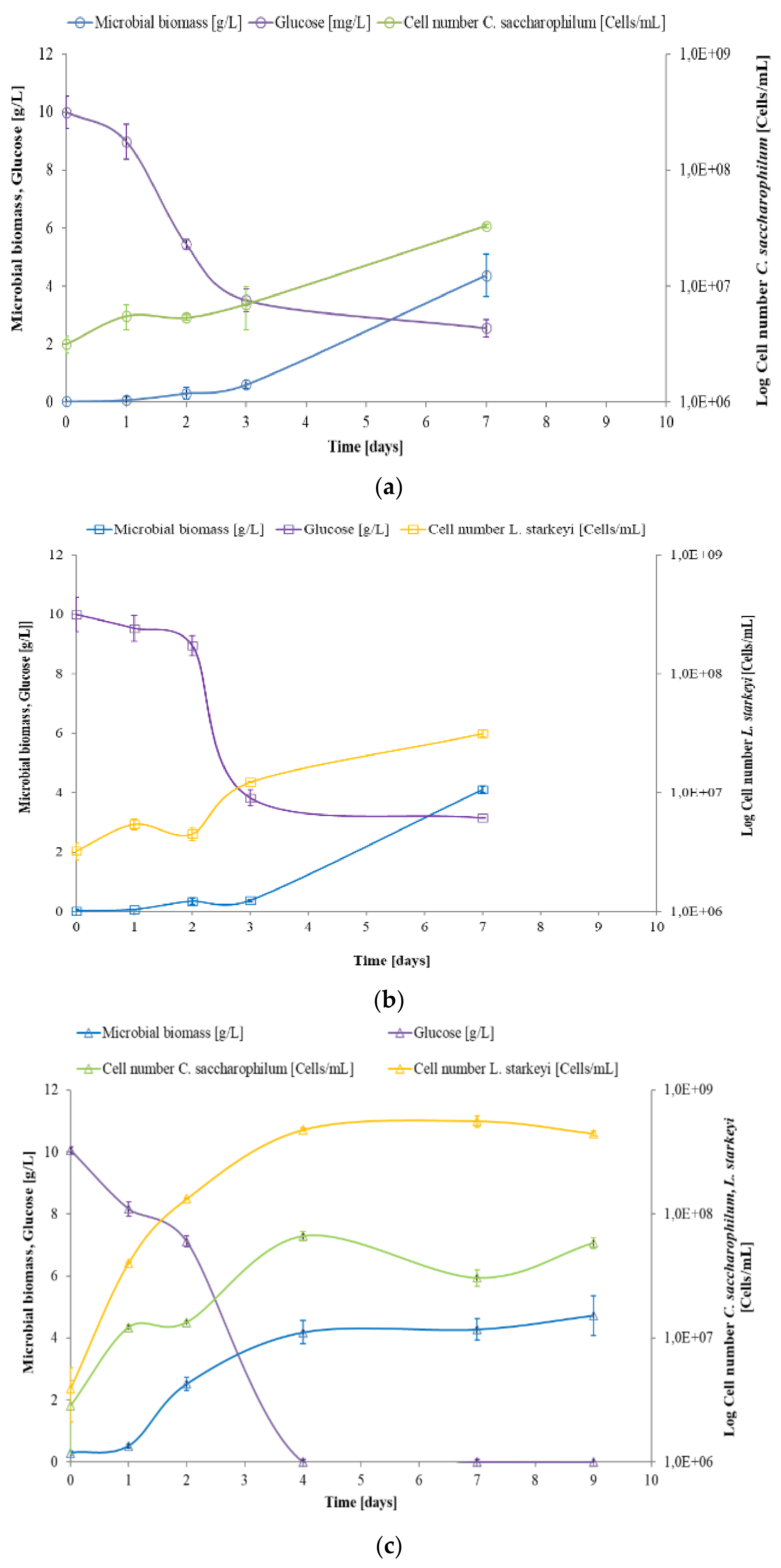
3.1. Effects of Synthetic Media on C. saccharophilum and L. starkeyi Growth Performances in Mono and Mixed Culture
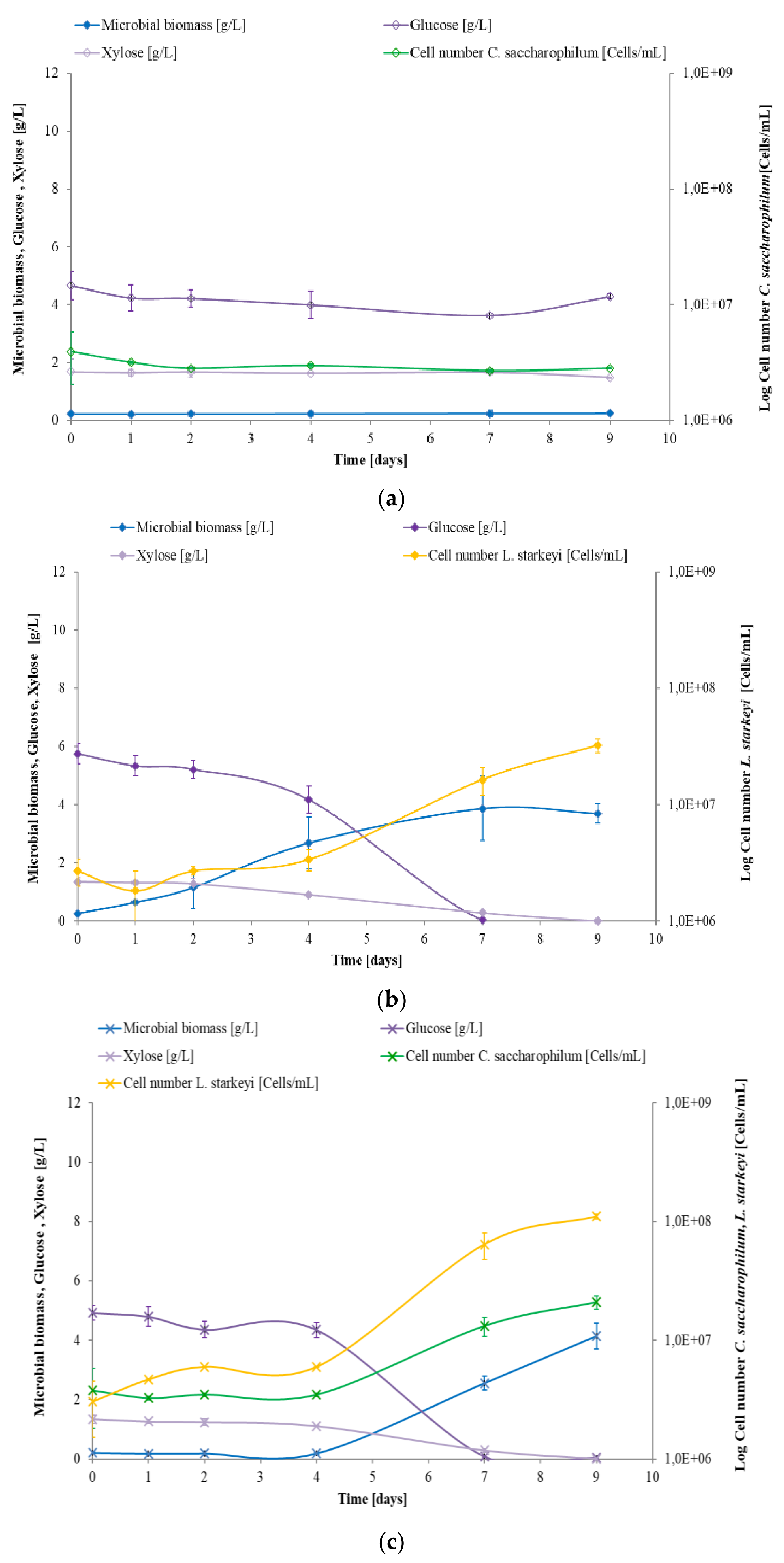
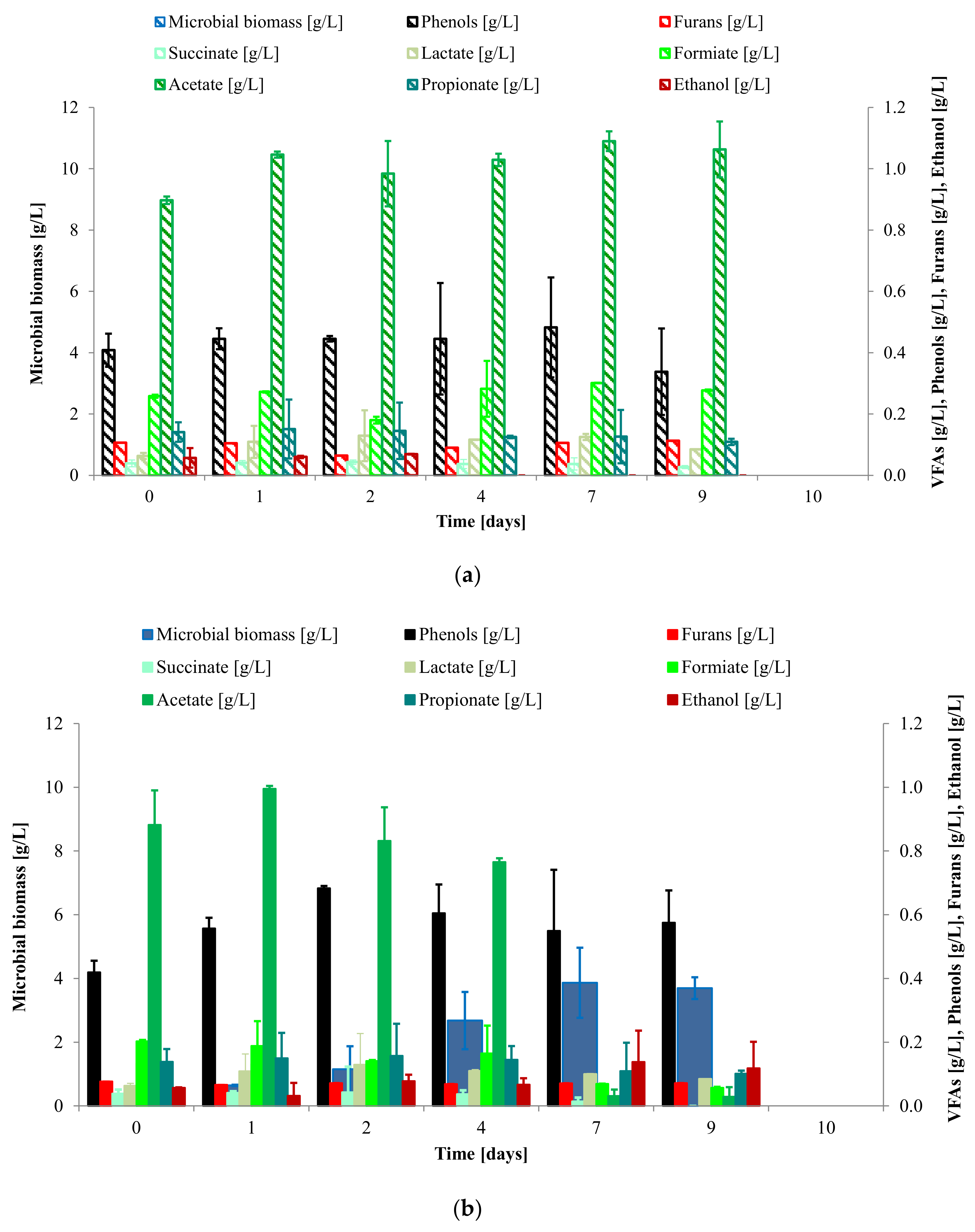
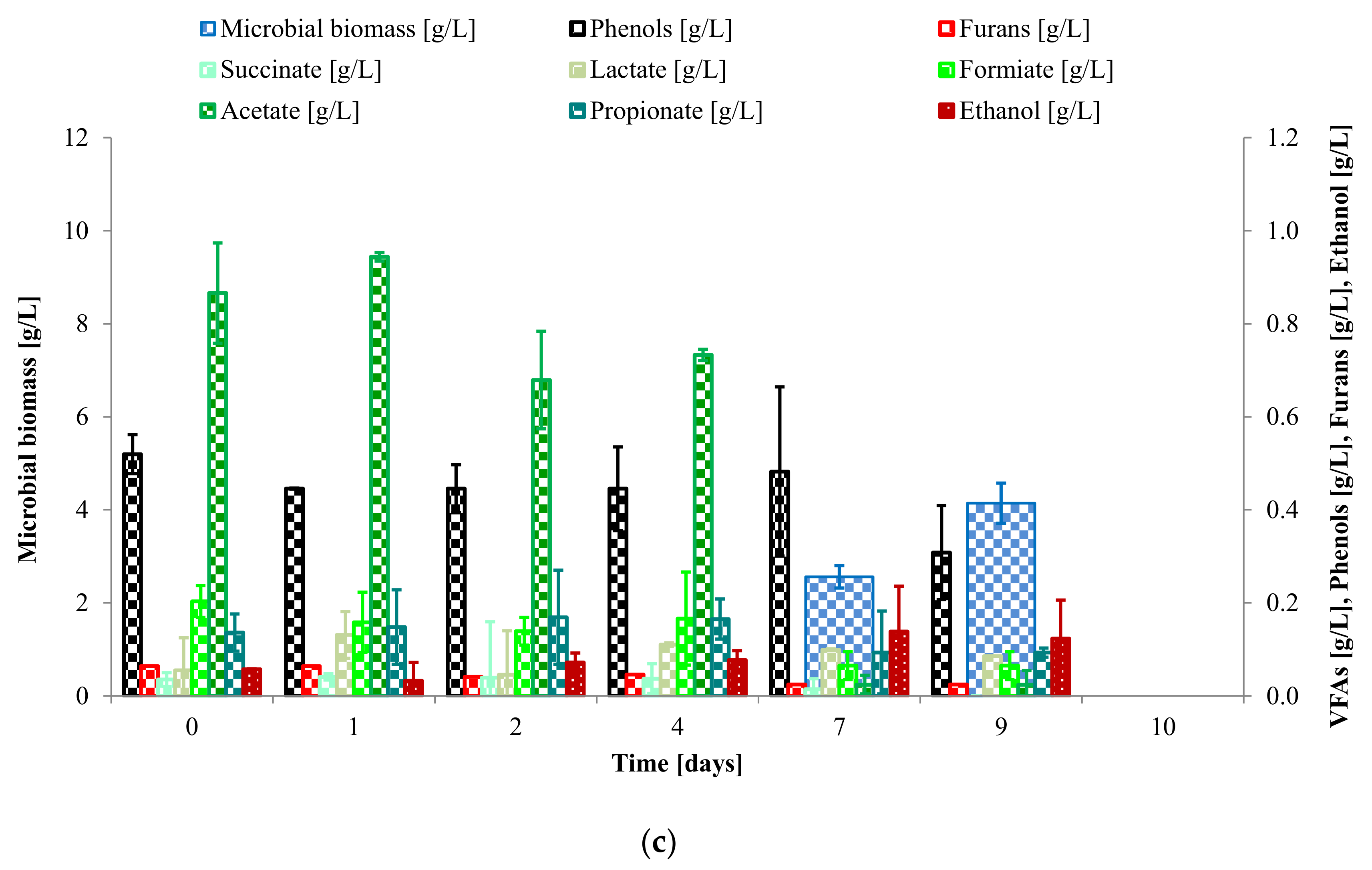
3.2. Study of C. saccharophilum and L. starkeyi Single and Mixed Cultures in Arundo donax Hydrolysate
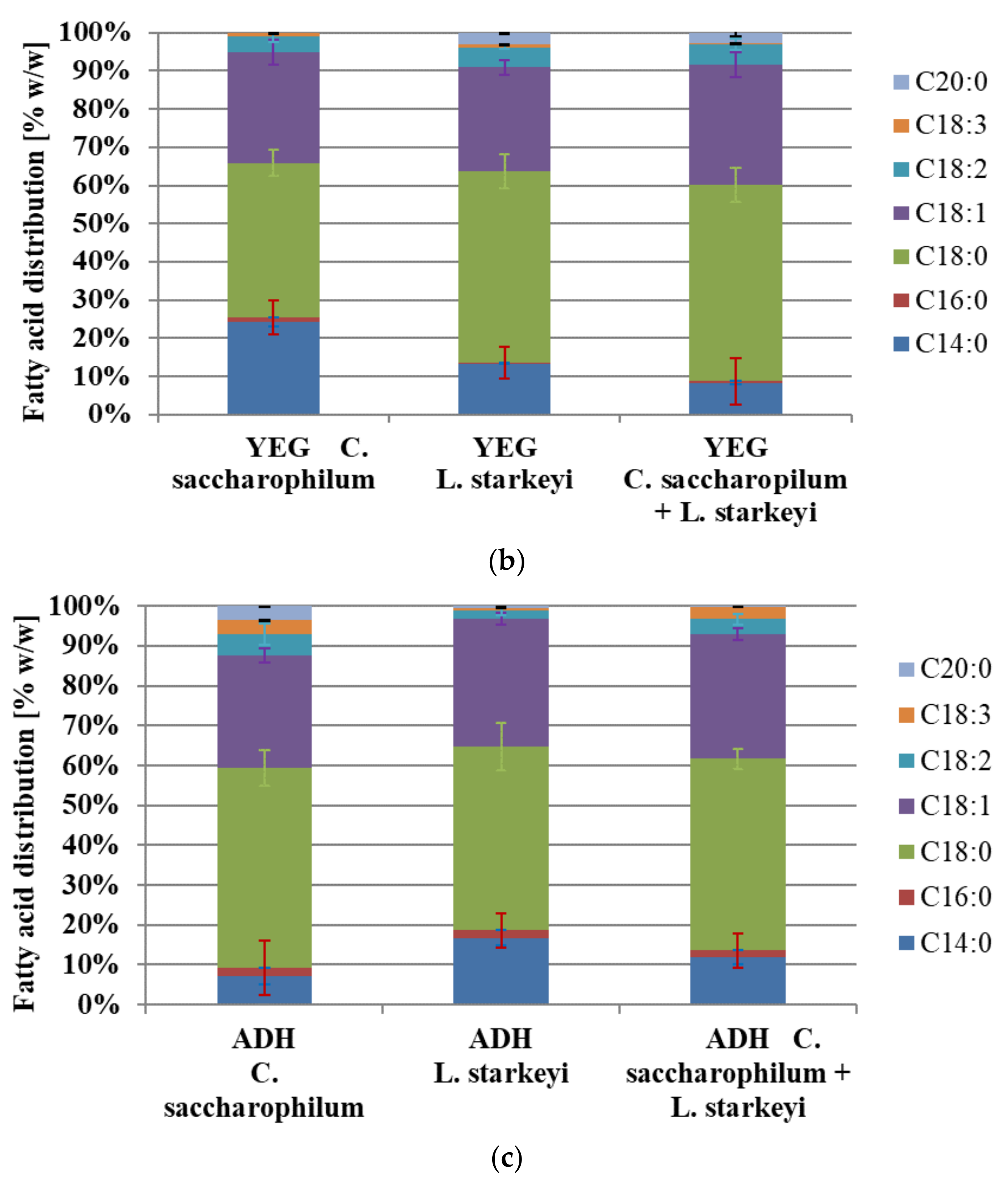
3.3. Lipid Production and Fatty Acid Distribution
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pérez, A.T.E.; Camargo, M.; Rincón, P.C.N.; Marchant, M.A. Key challenges and requirements for sustainable and industrialized biorefinery supply chain design and management: A bibliographic analysis. Renew. Sustain. Energy Rev. 2017, 69, 350–359. [Google Scholar] [CrossRef]
- Abghari, A.; Chen, S. Yarrowia lipolytica as an oleaginous cell factory platform for production of fatty acid-based biofuel and bioproducts. Front. Energy Res. 2014, 2, 1–21. [Google Scholar] [CrossRef]
- Huang, C.; Chen, X.F.; Xiong, L.; Chen, X.D.; Ma, L.L.; Chen, Y. Single cell oil production from low-cost substrates: The possibility and potential of its industrialization. Biotechnol. Adv. 2013, 31, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Patel, A.; Methani, J.; Pruthi, P.A.; Pruthi, V.; Poluri, K.M. Co-culturing of oleaginous microalgae and yeast: Paradigm shift towards enhanced lipid productivity. Environ. Sci. Pollut. Res. 2019, 26, 16952–16973. [Google Scholar] [CrossRef]
- Naidoo, R.K.; Simpson, Z.F.; Oosthuizen, J.R.; Bauer, F.F. Nutrient exchange of carbon and nitrogen promotes the formation of stable mutualisms between Chlorella sorokiniana and Saccharomyces cerevisiae under engineered synthetic growth conditions. Front. Microbiol. 2019, 10, 609. [Google Scholar] [CrossRef]
- Hays, S.G.; Yan, L.L.W.; Silver, P.A.; Ducat, D.C. Synthetic photosynthetic consortia define interactions leading to robustness and photoproduction. J. Biol. Eng. 2017, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, C.T.; Butler, K.; Hays, S.G.; Guarnieri, M.T.; Oyler, G.A.; Betenbaugh, M.J. Mimicking lichens: Incorporation of yeasts strains together with sucrose secreting cyanobacteria improves survival, growth, ROS removal, and lipid production in a stable mutualistic co-culture production platform. Biotechnol. Biofuels 2017, 10, 55. [Google Scholar] [CrossRef]
- Liu, J.; Huang, J.; Sun, Z.; Zhong, Y.; Jiang, Y.; Chen, F. Differential lipid and fatty acid profiles of photoautotrophic and heterotrophic Chlorella zofingiensis: Assessment of algal oils for biodiesel production. Bioresour. Technol. 2011, 102, 106–110. [Google Scholar] [CrossRef]
- Wilhelm, C.; Jakob, T. From photons to biomass and biofuels: Evaluation of different strategies for the improvement of algal biotechnology based on comparative energy balances. Appl. Microbiol. Biotechnol. 2011, 92, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Zheng, Y.; Lucker, B.; Chen, S. Integrated System for Production of Biofuel Feedstock. U.S. Patent WO/2010/014797, 4 February 2010. [Google Scholar]
- Beopoulos, A.; Nicaud, J.M. Yeast: A new oil producer? OCL 2012, 19, 22–28. [Google Scholar] [CrossRef]
- Qin, L.; Liu, L.; Zeng, A.-P.; Wei, D. From low-cost substrates to Single Cell Oils synthesized by oleaginous yeasts. Bioresour. Technol. 2017, 245, 1507–1519. [Google Scholar] [CrossRef]
- Lodder, J.; Kreger-van Rij, N.J.W. The Yeast: A Taxonomic Study, 1st ed.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1952. [Google Scholar]
- Darienko, T.; Gustavs, L.; Mudimu, O.; Menendez, C.R.; Schumann, R.; Karsten, U.; Friedl, T.; Proschold, T. Chloroidium, a common terrestrial coccoid green alga previously assigned to Chlorella (Trebouxiophyceae, Chlorophyta). Eur. J. Phycol. 2010, 45, 79–95. [Google Scholar] [CrossRef]
- Calvey, C.H.; Su, Y.K.; Willis, L.B.; McGee, M.; Jeffries, T.W. Nitrogen limitation, oxygen limitation, and lipid accumulation in Lipomyces Starkeyi. Bioresour. Technol. 2016, 200, 780–788. [Google Scholar] [CrossRef]
- McNeil, B.A.; Stuart, D.T. Optimization of C16 and C18 fatty alcohol production by an engineered strain of Lipomyces starkeyi. J. Ind. Microbiol. Biotechnol. 2018, 45, 1–14. [Google Scholar] [CrossRef]
- Xavier, M.C.A.; Coradini, A.L.V.; Deckmann, A.C.; Franco, T.T. Lipid production from hemicellulose hydrolysate and acetic acid by Lipomyces starkeyi and the ability of yeast to metabolize inhibitors. Biochem. Eng. J. 2017, 118, 11–19. [Google Scholar] [CrossRef]
- Beardall, J. CO2 accumulation by Chlorella saccharophilum (Chlorophyceae) at low external pH: Evidence for active transport of inorganic carbon at the chloroplast envelope. J. Phycol. 1981, 17, 371–373. [Google Scholar] [CrossRef]
- Herrera-Valencia, V.A.; Contreras-Pool, P.Y.; Lopez-Adrian, S.J.; Peraza-Echeverria, S.; Barahona-Perez, L.F. The green microalga Chlorella saccharophilum as a suitable source of oil for biodiesel production. Curr. Microbiol. 2011, 63, 151–157. [Google Scholar] [CrossRef]
- Tan, C.K.; Johns, M.R. Fatty acid production by heterotrophic Chlorella saccharophilum. Hydrobiologia 1991, 215, 13–19. [Google Scholar] [CrossRef]
- Zhang, T.-Y.; Hu, H.-Y.; Wu, Y.-H.; Zhuang, L.-L.; Xu, X.-Q.; Wang, X.-X.; Dao, G.-H. Promising solutions to solve te bottlenecks in the large-scale cultivation of microalgae for biomass/bioenergy production. Renew. Sustain. Energy Rev. 2016, 60, 1602–1614. [Google Scholar] [CrossRef]
- Fagnano, M.; Impagliazzo, A.; Mori, M.; Fiorentino, N. Agronomic and Environmental Impacts of Giant Reed (Arundo donax L.): Results from a Long-Term Field Experiment in Hilly Areas Subject to Soil Erosion. Bioenergy Res. 2015, 8, 415–422. [Google Scholar] [CrossRef]
- Zuccaro, G.; Travaglini, G.; Caputo, G.; Pirozzi, D. Enzymatic Hydrolysis and Oleaginous Fermentation of Steam-Exploded Arundo donax and Lipomyces Starkeyi in a Single Bioreactor for Microbial Oil Accumulation. J. Multidiscip. Eng. Sci. Stud. 2019, 5, 2577–2586. [Google Scholar]
- Garrote, G.; Domínguez, H.; Parajó, J.C. Hydrothermal processing of lignocellulosic materials. Holz Roh-Werkst 1999, 57, 191–202. [Google Scholar] [CrossRef]
- Adney, B.; Baker, J. Measurement of Cellulase Activities; NREL Analytical Procedure LAP-006; National Renewable Energy Laboratory: Golden, CO, USA, 1996. [Google Scholar]
- Wood, T.M.; Bhat, K.M. Methods for measuring cellulase activities. Methods Enzymol. 1988, 160, 87–112. [Google Scholar]
- Singleton, V.L.; Orhofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteau reagent. Methods Enzym. 1999, 299, 152–178. [Google Scholar]
- Martinez, A.; Rodriguez, M.E.; York, S.W.; Preston, J.F.; Ingram, L.O. Use of UV absorbance to monitor furans in dilute acid hydrolysates of biomass. Biotechnol. Prog. 2000, 16, 637–641. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Redfield, A.C. On the Proportions of Organic Derivatives in Sea Water and Their Relation to the Composition of Plankton. In James Johnstone Memorial Volume; University Press of Liverpool: Liverpool, UK, 1934; pp. 176–192. [Google Scholar]
- Zuccaro, G.; Steyer, J.-P.; van Lis, R. The algal trophic mode affects the interaction and oil production of a synergistic microalga-yeast consortium. Bioresour. Technol. 2019, 273, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Wijsman, M.R.; van Dijken, J.P.; van Kleeff, B.H.A.; Scheffers, W.A. Inhibition of fermentation and growth in batch cultures of the yeast Brettanomyces intermedius upon a shift from aerobic to anaerobic conditions (Custers effect). Antonie Van Leeuwenhoek 1984, 50, 183–192. [Google Scholar] [CrossRef]
- Li, H.; Zhong, Y.; Lu, Q.; Zhang, X.; Wang, Q.; Liu, H.; Diao, Z.; Yao, C.; Liu, H. Co-cultivation of Rhodotorula glutinis and Chlorella pyrenoidosa to improve nutrient removal and protein content by their synergistic relationship. RSC Adv. 2019, 9, 14331–14342. [Google Scholar] [CrossRef]
- Ling, J.; Nip, S.; Cheok, W.L.; Alves de Toledo, R.; Shim, H. Lipid production by a mixed culture of oleaginous yeast and microalga from distillery and domestic mixed wastewater. Bioresour. Technol. 2014, 173, 132–139. [Google Scholar] [CrossRef]
- Zuccaro, G.; Pirozzi, D.; Yousuf, A. Lignocellulosic biomass to biodiesel. In Lignocellulosic Biomass to Liquid Biofuels; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2020; pp. 127–167. [Google Scholar]
- Miazek, K.; Remacle, C.; Richel, A.; Goffin, D. Effect of lignocellulose related compounds on microalgae growth and product biosynthesis: A review. Energies 2014, 7, 4446–4481. [Google Scholar] [CrossRef]
- Bajguz, A.; Czerpak, R.; Piotrowska, A.; Polecka, M. Effect of isomers of hydroxybenzoic acid on the growth and metabolism of Chlorella vulgaris Beijerinck (Chlorophyceae). Acta Soc. Bot. Pol. 2001, 70, 253–259. [Google Scholar] [CrossRef]
- Larson, L.J. Effect of phenolic acids on growth of Chlorella pyrenoidosa. Hydrobiologia 1989, 183, 217–222. [Google Scholar] [CrossRef]
- Larsson, S.; Palmqvist, E.; Hahn-Hägerdal, B.; Tengborg, C.; Stenberg, K.; Zacchi, G.; Nilvebrant, N.O. The generation of fermentation inhibitors during dilute acid hydrolysis of softwood. Enz. Microb. Technol. 1988, 24, 151–159. [Google Scholar] [CrossRef]
- Cea Barcia, G.E.; Imperial Cervantes, R.A.; Torres Zuniga, I.; van Den Hende, S. Converting tequila vinasse diluted with tequila process water into microalgae-yeast flocs and dischargeable effluent. Bioresour. Technol. 2020, 300, 122644. [Google Scholar] [CrossRef]
- Gong, Z.; Zhou, W.; Shen, H.; Yang, Z.; Wang, G.; Zuo, Z.; Huo, Y.; Zhao, Z.K. Co-fermentation of acetate and sugars facilitating microbial lipid production on acetate-rich biomass hydrolysates. Bioresour. Technol. 2016, 207, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.B. Another explanation for the toxicity of fermentation acids at low pH: Anion accumulation versus uncoupling. J. Appl. Bacteriol. 1992, 73, 363–370. [Google Scholar] [CrossRef]
- Anschau, A.; Xavier, M.C.; Hernalsteens, S.; Franco, T.T. Effect of feeding strategies on lipid production by Lipomyces starkeyi. Bioresour. Technol 2014, 157, 214–222. [Google Scholar] [CrossRef]
- Gong, Z.; Wang, Q.; Shen, H.; Hu, C.; Jin, G.; Zhao, Z.K. Co-fermentation of cellobiose and xylose by Lipomyces starkeyi for lipid production. Bioresour. Technol. 2012, 117, 20–24. [Google Scholar] [CrossRef]
- Yang, X.B.; Jin, G.J.; Gong, Z.W.; Shen, H.W.; Song, Y.H.; Bai, F.W.; Zhao, Z.B.K. Simultaneous utilization of glucose and mannose from spent yeast cell mass for lipid production by Lipomyces starkeyi. Bioresour. Technol. 2014, 158, 383–387. [Google Scholar] [CrossRef]
- Pirozzi, D.; Ausiello, A.; Yousuf, A.; Zuccaro, G.; Toscano, G. Exploitation of oleaginous yeasts for the production of microbial oils from agricultural biomass. Chem. Eng. Trans. 2014, 37, 469–474. [Google Scholar]
- Probst, K.V.; Vadlani, P.V. Production of single cell oil from Lipomyces starkeyi ATCC 56304 using biorefinery by-products. Bioresour. Technol. 2015, 198, 268–275. [Google Scholar] [CrossRef]
- Matsakas, L.; Sterioti, A.A.; Rova, U.; Christakopoulos, P. Use of dried sweet sorghum for the efficient production of lipids by the yeast Lipomyces starkeyi CBS 1807. Ind. Crop. Prod. 2014, 62, 367–372. [Google Scholar] [CrossRef]
- Liu, L.; Chen, J.; Lim, P.-E.; Wei, D. Enhanced single cell oil production by mixed culture of Chlorella pyrenoidosa and Rhodotorula glutinis using cassava bagasse hydrolysate as carbon source. Bioresour. Technol. 2018, 255, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wei, D.; Wang, Z.; Alam, M.A. Advantage assessment of mixed culture of Chlorella vulgaris and Yarrowia lipolytica for treatment of liquid digestate of year industry and cogeneration of biofuel feedstock. Appl. Biochem. Biotechnol. 2019, 187, 856–869. [Google Scholar] [CrossRef]
- Zhang, Z.; Pang, Z.; Xu, S.; Wei, T.; Song, L.; Wang, G.; Zhang, J.; Yang, X. Improved carotenoid productivity and COD removal efficiency by co-culture of Rhodotorula glutinis and Chlorella vulgaris using starch wastewaters as raw material. Appl. Biochem. Biotechnol. 2019, 189, 193–205. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Liu, H.; Zhang, J. Enhanced lipid production with undetoxified corncob hydrolysate by Rhodotorula glutinis using a high cell density culture strategy. Bioresour. Technol. 2015, 180, 32–39. [Google Scholar] [CrossRef]
- Younes, S.; Bracharz, F.; Awad, D.; Qoura, F.; Mehlmer, N.; Brueck, T. Microbial lipid production by oleaginous yeasts grown on Scenedesmus otusiusculus microalgae biomass hydrolysate. Bioprocess. Biosyst. Eng. 2020, 43, 1629–1638. [Google Scholar] [CrossRef]
- Wang, S.; Wu, Y.; Wang, X. Heterotrophic cultivation of Chlorella pyrenoidosa using sucrose as the sole carbon source by co-culture with Rhodotorula glutinis. Bioresour. Technol. 2016, 220, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Iasimone, F.; Zuccaro, G.; D’Oriano, V.; Franci, G.; Galdiero, M.; Pirozzi, D. Combined yeast and microalgal cultivation in a pilot-scale raceway pond for urban wastewater treatment and potential biodiesel production. Water Sci. Technol. 2018, 77, 1062–1071. [Google Scholar] [CrossRef]
- Tkachenko, A.F.; Tigunova, S.M.; Shulga, S.M. Microbial lipids as a source of biofuel. Cytol. Genet. 2013, 47, 343–348. [Google Scholar] [CrossRef]
- Stansell, G.R.; Gray, V.M.; Sym, S.D. Microalgal fatty acid composition: Implications for biodiesel quality. J. Appl. Phycol. 2011, 24, 791–801. [Google Scholar] [CrossRef]
- Parsons, S.; Allen, M.; Chuck, C.J. Coproducts of algae and yeast-derived single cell oils: A critical review of their role in improving biorefinery sustainability. Bioresour. Technol. 2020, 303, 122862. [Google Scholar] [CrossRef]
- Asraful, A.M.; Masjuki, H.H.; Kalam, M.A.; Rizwanul Fattah, I.M.; Imtenan, S.; Shahir, S.A.; Mobarak, H.M. Production and comparison of fuel properties, engine performance, and emission characteristics of biodiesel from various non-edible vegetable oils: A review. Energy Convers. Manag. 2014, 80, 202–228. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | μx [d−1] | Yx/s [g·g−1] | μs [d−1] | PCO2 [mg·L−1·d−1] | m Lipid [g] | Lipid Content [glipid·gbiomass−1] | Lipid Yield max Ylipid max [g·L−1] | Biomass Productivity [mg·L−1·d−1] | Lipid Productivity [mg·L−1·d−1] | Cellsmax C. saccharophila [106 cells·mL−1] | Cellsmax L. starkeyi [106 cells·mL−1] | Lipid Yield max/Cell max C. saccharophila [μg·cells−1·10−6] | Lipid Yield max/Cell max L. starkeyi [μg·cells−1·10−6] |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BBM + G Chl | 0.381 | 0.471 | 0.689 | 610.0 | 0.018 | 0.076 ± 0.03 | 0.175 ± 0.04 | 328.57 | 25.00 | 36.0 ± 1.76 | 4.9 | ||
| BBM + G Lip | 0.037 | 0.095 | 0.279 | 49.9 | 0.002 | 0.093 ± 0.01 | 0.020 ± 0.04 | 30.65 | 2.87 | 2.0 ± 0.65 | 10.0 | ||
| BBM + G Chl Lip | 0.472 | 0.308 | 1.437 | 474.7 | 0.021 | 0.064 ± 0.02 | 0.211 ± 0.17 | 366.20 | 30.14 | 58.7 ± 1.93 | 309.3 ± 5.91 | 3.6 | 0.7 |
| YEG Chl | 0.944 | 0.583 | 1.065 | 1168.6 | 0.022 | 0.051 ± 0.01 | 0.221 ± 0.24 | 625.00 | 31.57 | 33.0 ± 1.27 | 1.5 | ||
| YEP Lip | 0.803 | 0.596 | 0.978 | 1095.9 | 0.021 | 0.052 ± 0.03 | 0.212 ± 0.07 | 586.31 | 30.29 | 31.5 ± 0.77 | 6.7 | ||
| YEG Chl Lip | 1.166 | 0.440 | 1.118 | 925.4 | 0.032 | 0.068 ± 0.01 | 0.32 ± 0.28 | 611.90 | 35.56 | 66.7 ± 3.21 | 561.3 ± 16.64 | 1.0 | 0.1 |
| ADH Chl | 0.003 | 0.000 * | 0.065 | 3.8 | 0.002 | 0.069 ± 0.03 | 0.016 ± 0.00 | 26.75 | 1.86 | 3.2 ± 0.37 | 5.0 | ||
| ADH Lip | 0.614 | 0.422 * | 0.883 | 837.2 | 0.032 | 0.074 ± 0.01 | 0.318 ± 0.05 | 552.38 | 35.33 | 32.45 ± 1.87 | 9.8 | ||
| ADH Chl Lip | 0.791 | 0.539 * | 0.786 | 822.2 | 0.034 | 0.081 ± 0.04 | 0.335 ± 0.14 | 365.47 | 37.22 | 20.9 ± 1.23 | 110.0 ± 3.83 | 16.0 | 3.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuccaro, G.; del Mondo, A.; Pinto, G.; Pollio, A.; De Natale, A. Biorefinery-Based Approach to Exploit Mixed Cultures of Lipomyces starkeyi and Chloroidium saccharophilum for Single Cell Oil Production. Energies 2021, 14, 1340. https://doi.org/10.3390/en14051340
Zuccaro G, del Mondo A, Pinto G, Pollio A, De Natale A. Biorefinery-Based Approach to Exploit Mixed Cultures of Lipomyces starkeyi and Chloroidium saccharophilum for Single Cell Oil Production. Energies. 2021; 14(5):1340. https://doi.org/10.3390/en14051340
Chicago/Turabian StyleZuccaro, Gaetano, Angelo del Mondo, Gabriele Pinto, Antonino Pollio, and Antonino De Natale. 2021. "Biorefinery-Based Approach to Exploit Mixed Cultures of Lipomyces starkeyi and Chloroidium saccharophilum for Single Cell Oil Production" Energies 14, no. 5: 1340. https://doi.org/10.3390/en14051340
APA StyleZuccaro, G., del Mondo, A., Pinto, G., Pollio, A., & De Natale, A. (2021). Biorefinery-Based Approach to Exploit Mixed Cultures of Lipomyces starkeyi and Chloroidium saccharophilum for Single Cell Oil Production. Energies, 14(5), 1340. https://doi.org/10.3390/en14051340