
Recent Progress and Trends in the Development of Microbial Biofuels from Solid Waste—A Review



Abstract
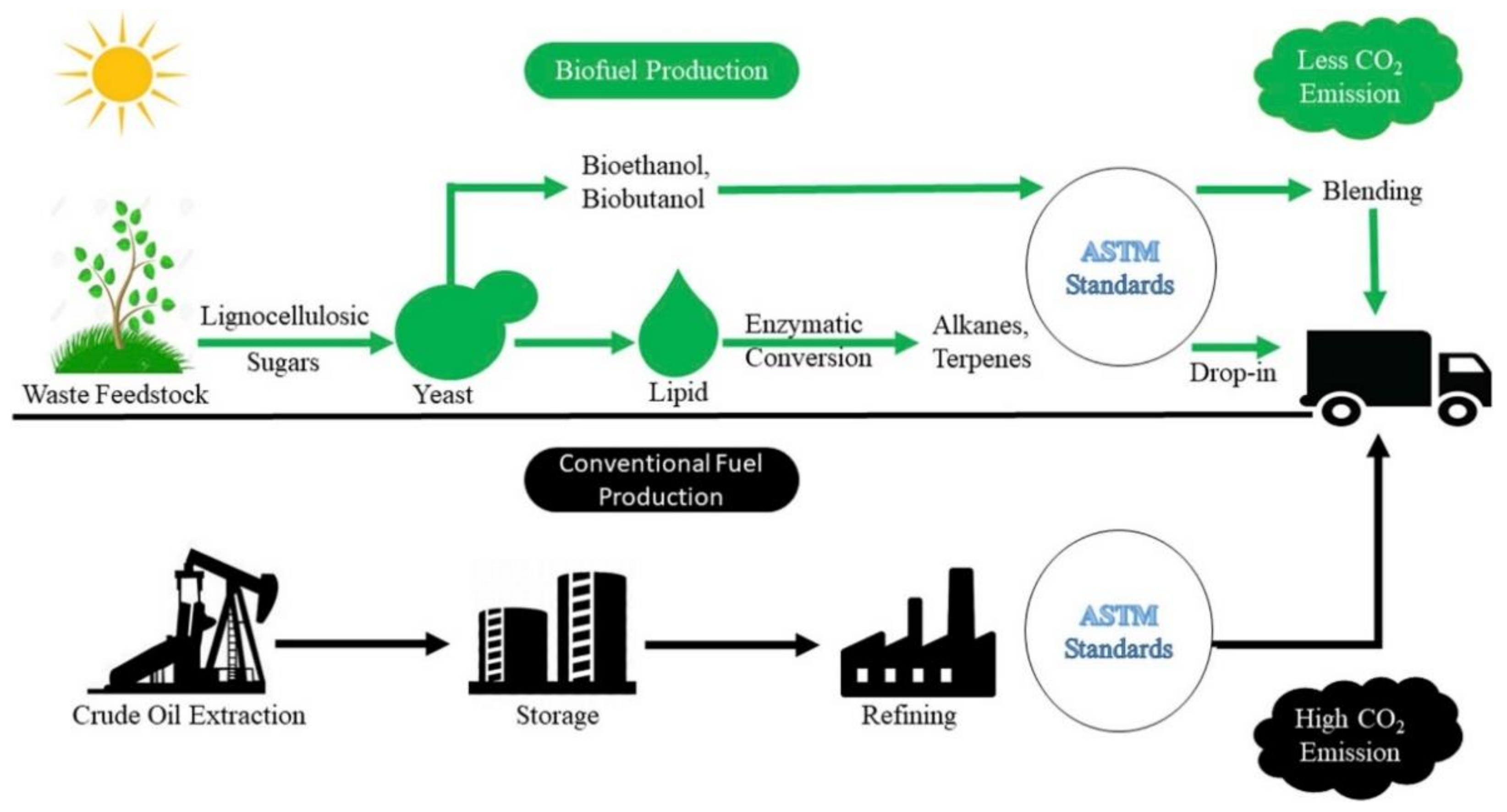
:1. Introduction
2. Residue and Biomass
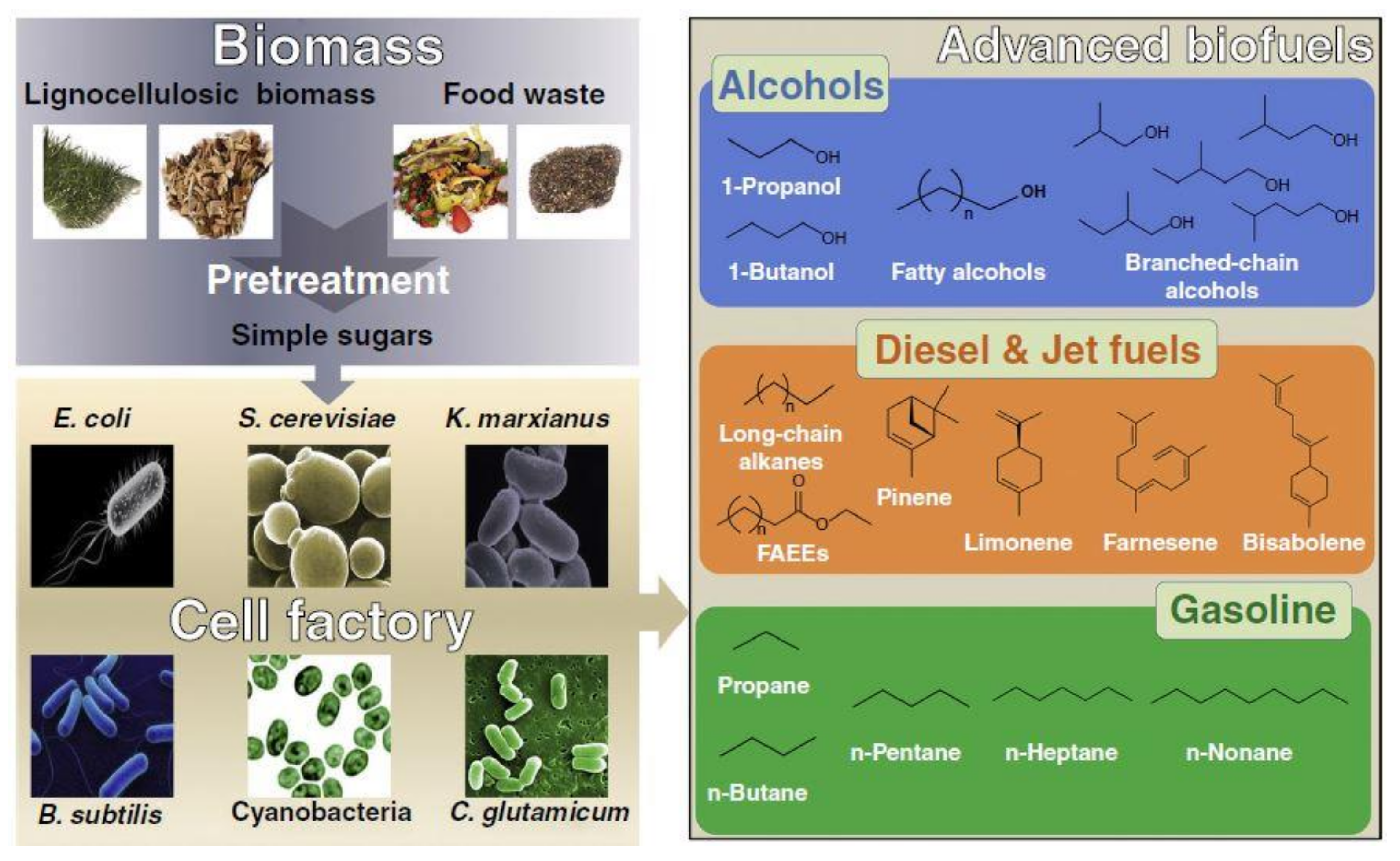
3. Biochemical Pathways for Fuel Production
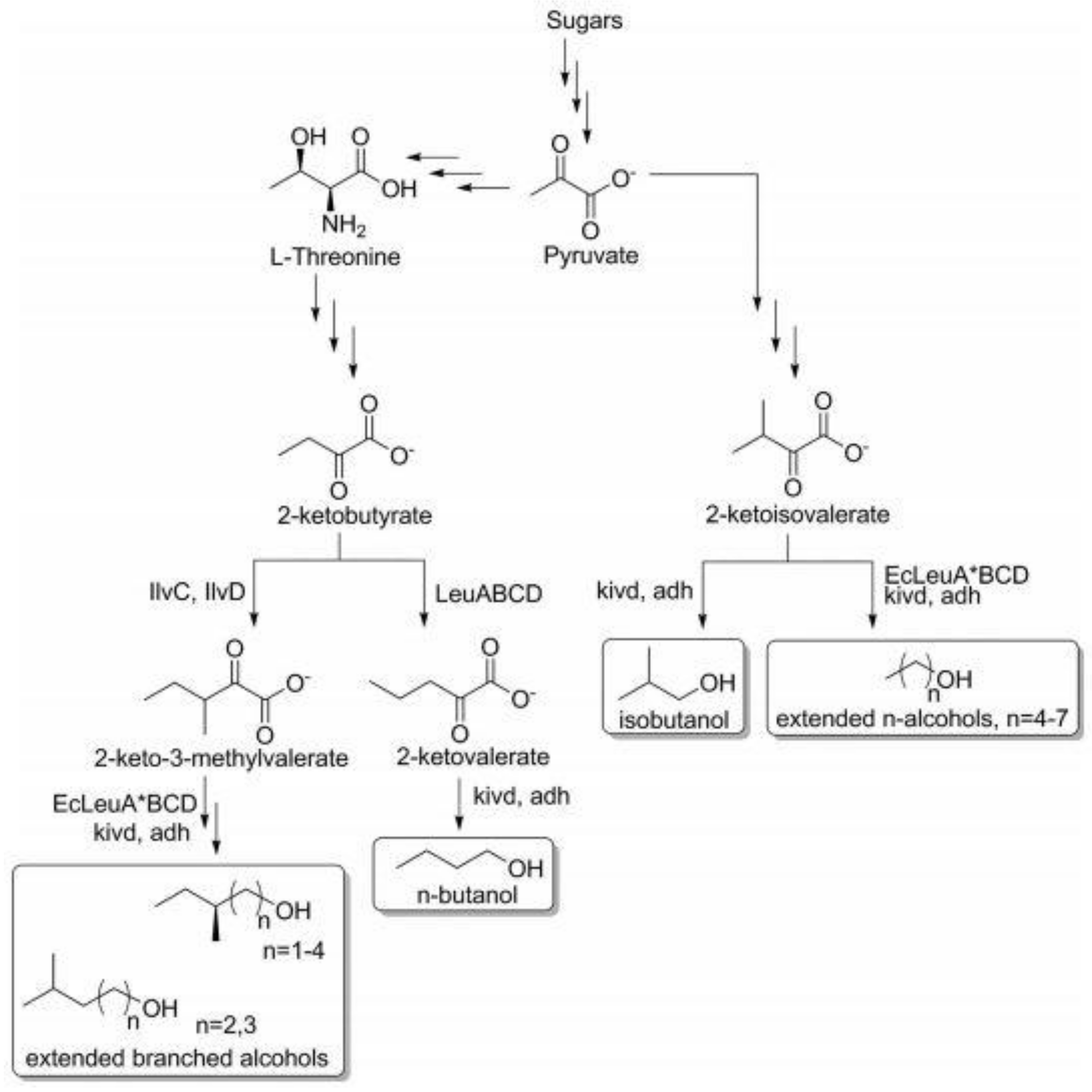
3.1. Pathways for Alcohol-Sourced Fuels
3.2. Pathways for Isoprenoid-Derived Fuels
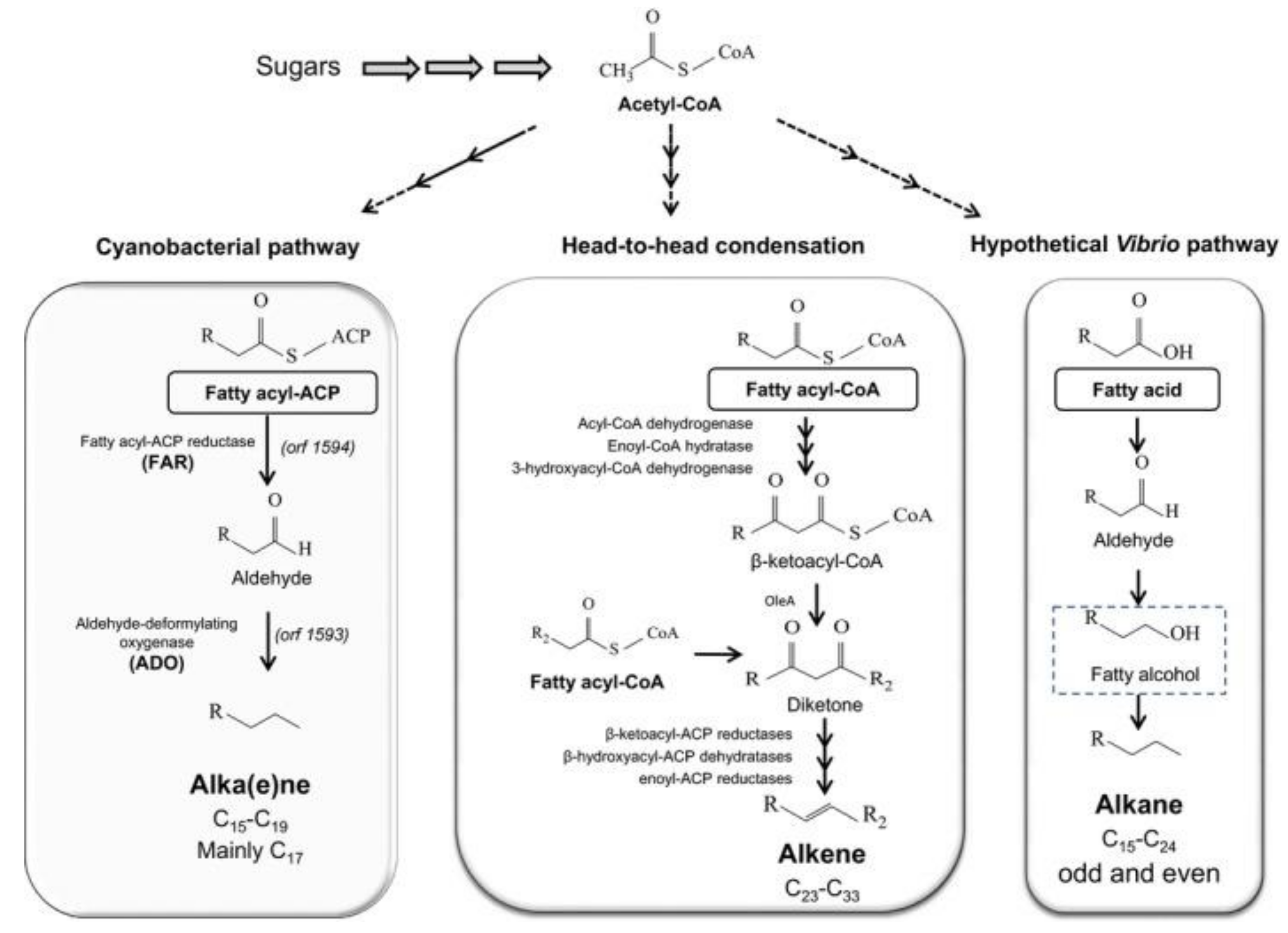
3.3. Pathways for Fatty Acid-Derived Fuels
3.4. Pathways for Polyketide-Derived Fuels
4. Types of Microorganisms: Wild Type vs. Engineered
GMM Risks and Regulations
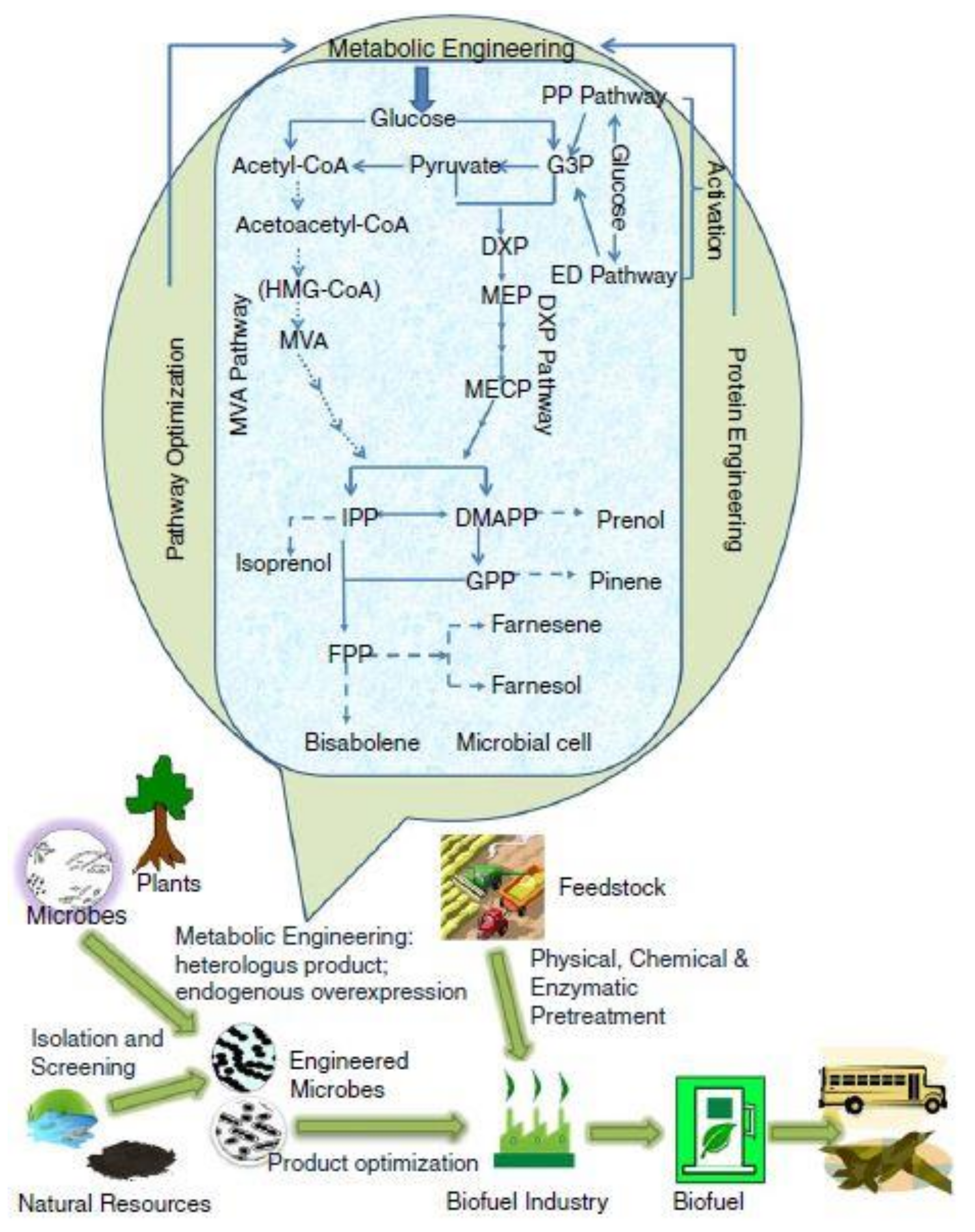
5. Microbial Engineering for the Development of Innovative Biofuels and Biosynthetic Pathways
5.1. Engineered Metabolic Pathway to Produce Fuels from Isoprenoids
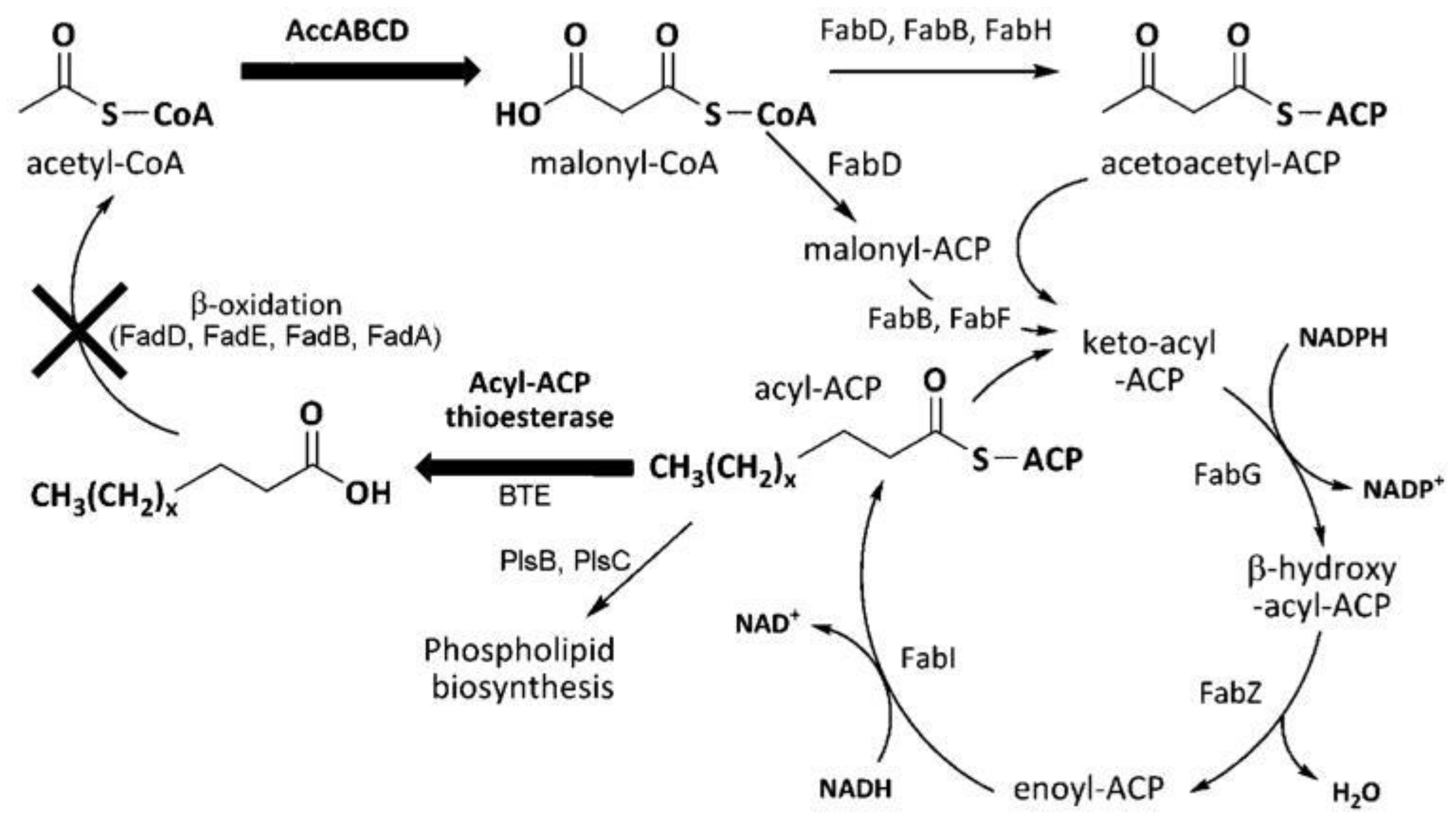
5.2. Engineered Metabolic Pathway to Produce Fuels from Fatty Acids
5.3. Engineered Metabolic Pathway to Produce Fuels from Alkanes
5.4. Engineered Metabolic Pathway to Produce Fuels from Alcohol
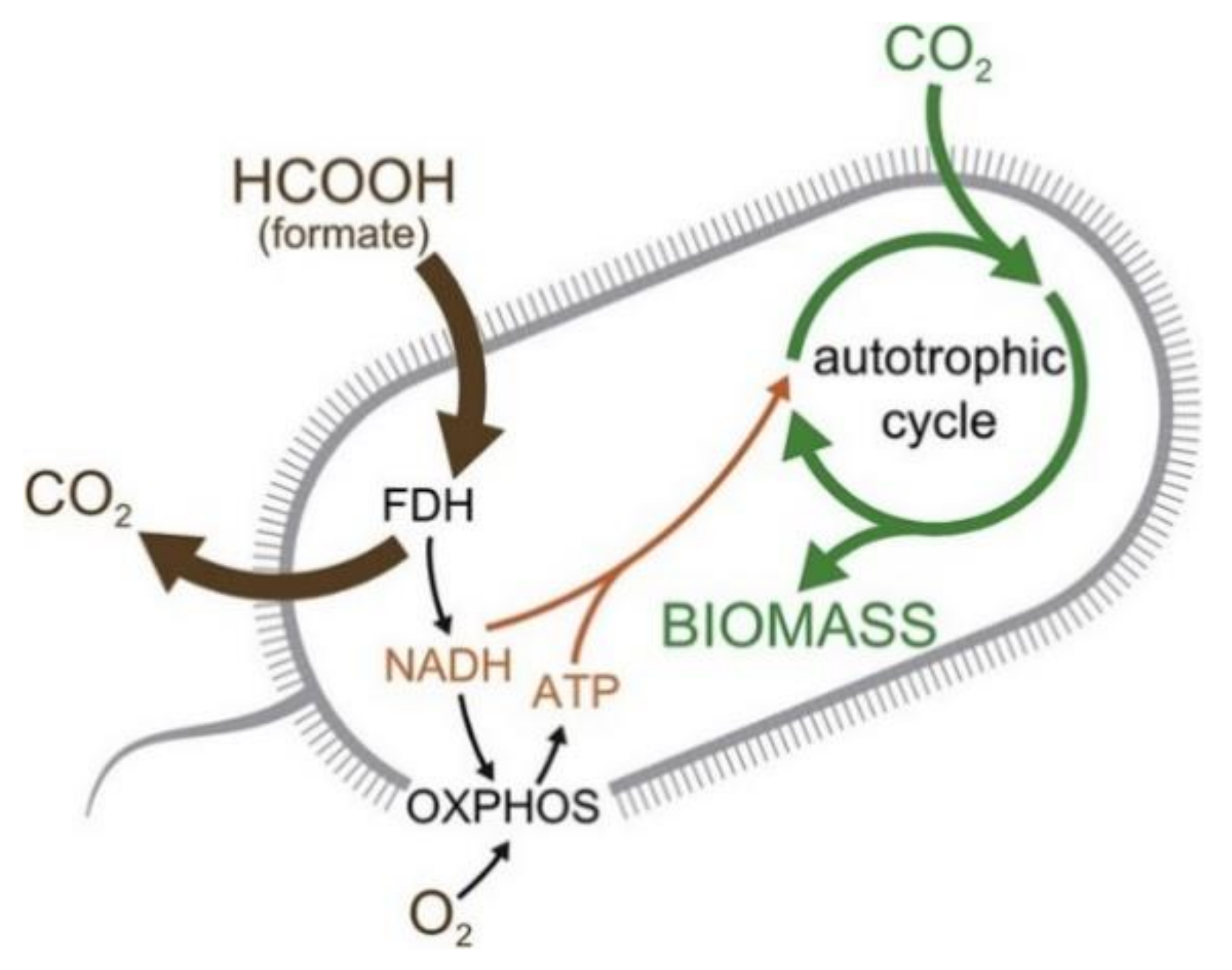
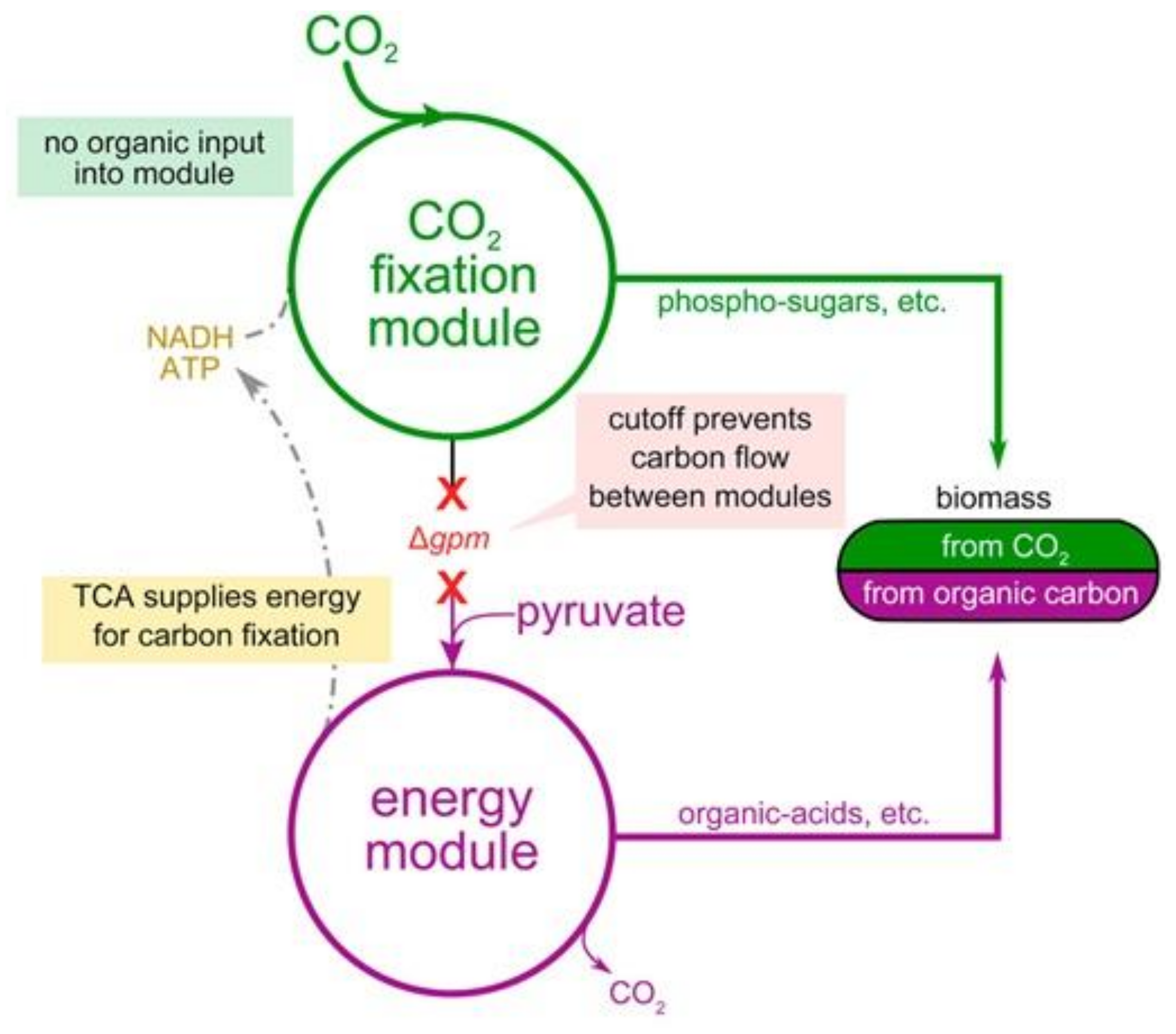
5.5. Sugar Synthesis from CO2 by Microorganisms to Produce Bioenergy Carrying Molecules
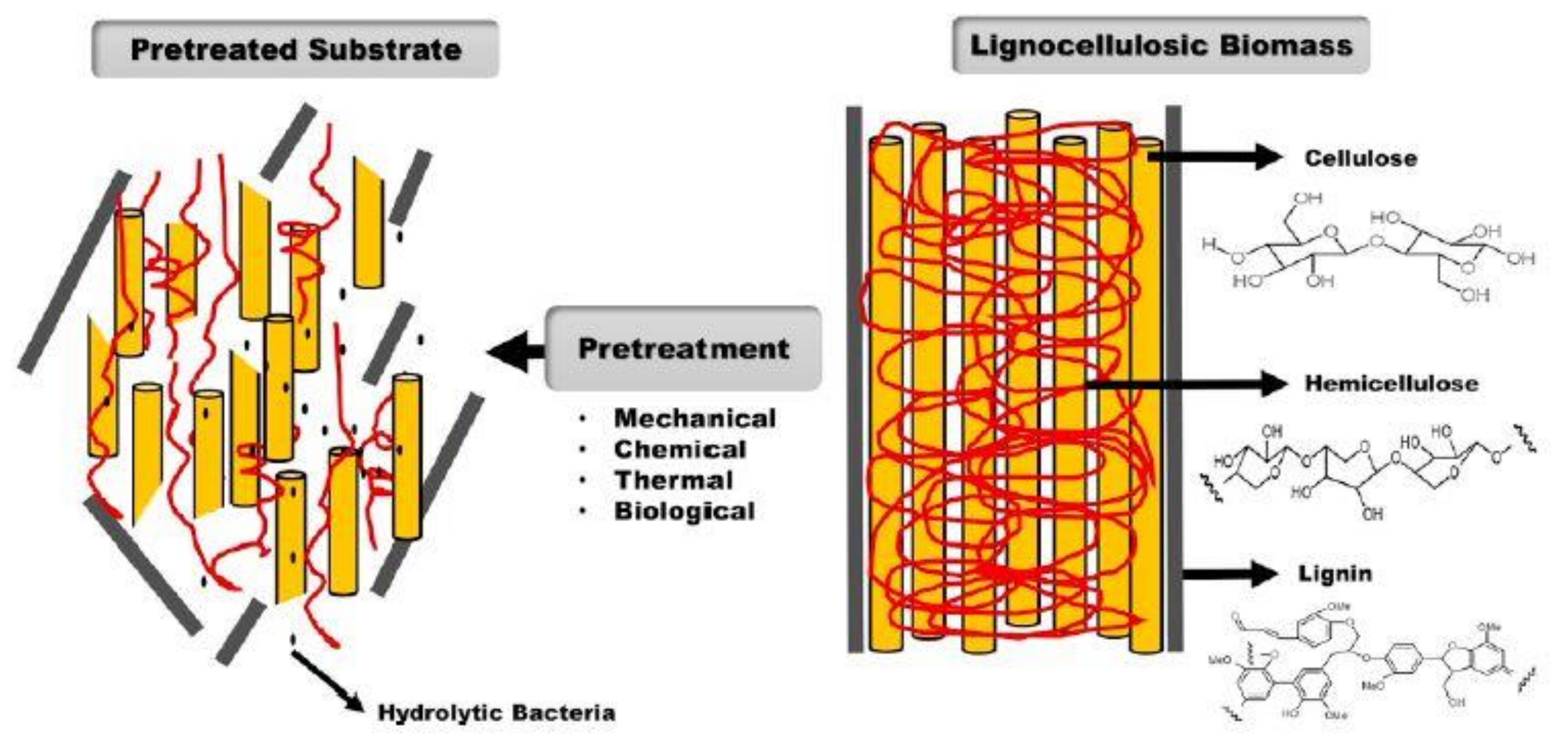
6. Pretreatment Processes
6.1. Pretreatment of Lignocellulosic Biomass
6.2. Biological Pretreatment of Lignocellulosic Biomass
6.3. Enzymatic Pretreatment
6.4. Fungal Pretreatment
6.5. Bacterial Pretreatment
6.6. Pretreatment of the Organic Fraction of Municipal Solid Waste
6.7. Biological Pretreatment to Produce Biofuels from OFMSW
6.8. Biological Pretreatment of Wastewater
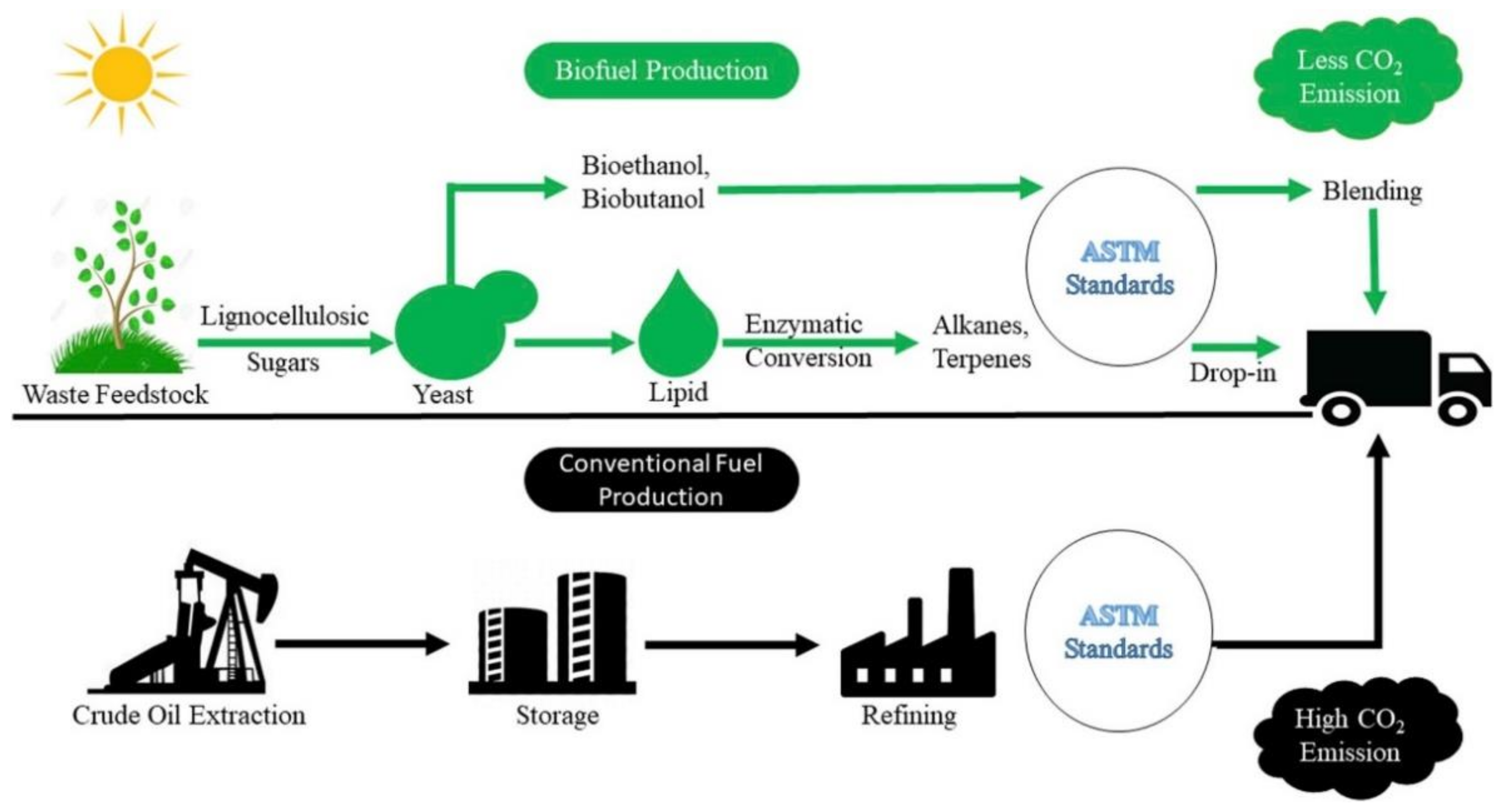
7. Outlook and Possibilities
7.1. Application of Microbial Fuels in Transport, Aviation, and Shipping
7.2. Application of Microbial Fuels in Energy Conversion and Storage
7.3. Application of Microbial Fuels in a Built Environment
8. Conclusions
- The early separation of OFMSW from commercial and demotic buildings will avoid some processes, such as autoclaving, and gain more lignocellulosic material, leading to the greater production of biofuels with less production cost;
- Greater attention should be given to residues with the most lignocellulosic content as a sustainable feedstock for microbial fuels;
- Enzymatic pretreatment appeared to be one of the most appropriate pretreatment methods, as it requires minimal chemical additives, low energy consumption, and a short reaction time;
- Pentose and simple sugar utilization and lignin management should be essential considerations in plants of the next generation;
- Efforts for laboratory studies should be guided in this field by employing genetic engineering techniques to selected species of organisms.
Author Contributions
Funding
Conflicts of Interest
References
- International Energy Agency. Net Zero by 2050-a Roadmap for the Global Energy Sector. Special Report. Available online: https://www.iea.org/reports/net-zero-by-2050 (accessed on 18 August 2021).
- Zhang, F.; Rodriguez, S.; Keasling, J. Metabolic engineering of microbial pathways for advanced biofuels production. Curr. Opin. Biotechnol. 2011, 22, 775–783. [Google Scholar] [CrossRef] [Green Version]
- Harvey, B.; Meylemans, H. The role of butanol in the development of sustainable fuel technologies. J. Chem. Technol. Biotechnol. 2010, 86, 2–9. [Google Scholar] [CrossRef]
- Sims, R.; Mabee, W.; Saddler, J.; Taylor, M. An overview of second-generation biofuel technologies. Bioresour. Technol. 2010, 101, 1570–1580. [Google Scholar] [CrossRef]
- Harvey, B.; Meylemans, H.; Gough, R.; Quintana, R.; Garrison, M.; Bruno, T. High-density biosynthetic fuels: The intersection of heterogeneous catalysis and metabolic engineering. Phys. Chem. Chem. Phys. 2014, 16, 9448–9457. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Mi, L.; Pontrelli, S.; Luo, S. Fuelling the future: Microbial engineering for the production of sustainable biofuels. Nat. Rev. Microbiol. 2016, 14, 288–304. [Google Scholar] [CrossRef]
- Searle, S.; Malins, C. Waste and residue availability for advanced biofuel production in EU Member States. Biomass Bioenergy 2016, 89, 2–10. [Google Scholar] [CrossRef]
- Uçkun Kiran, E.; Trzcinski, A.; Ng, W.; Liu, Y. Bioconversion of food waste to energy: A review. Fuel 2014, 134, 389–399. [Google Scholar] [CrossRef]
- Saini, R.; Hegde, K.; Kaur Brar, S.; Vezina, P. Advanced biofuel production and road to commercialization: An insight into bioconversion potential of Rhodosporidium sp. Biomass Bioenergy 2020, 132, 105439. [Google Scholar] [CrossRef]
- Howard, T.P.; Middelhaufe, S.; Moore, K.; Edner, C.; Kolak, D.M.; Taylor, G.N.; Parker, D.A.; Lee, R.; Smirnoff, N.; Aves, S.J.; et al. Synthesis of customized petroleum-replica fuel molecules by targeted modification of free fatty acid pools in Escherichia coli. Proc. Natl. Acad. Sci. USA 2013, 110, 7636–7641. [Google Scholar] [CrossRef] [Green Version]
- Fulton, L.; Lynd, L.; Körner, A.; Greene, N.; Tonachel, L. The need for biofuels as part of a low carbon energy future. Biofuel Bioprod. Biorefin. 2015, 9, 476–483. [Google Scholar] [CrossRef]
- Peralta-Yahya, P.; Zhang, F.; del Cardayre, S.; Keasling, J. Microbial engineering for the production of advanced biofuels. Nature 2012, 488, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.; Kim, H.; Gustavsson, M.; Lee, S. Recent trends in metabolic engineering of microorganisms for the production of advanced biofuels. Curr. Opin. Chem. Biol. 2016, 35, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chou, H.; Ham, T.; Lee, T.; Keasling, J. Metabolic engineering of microorganisms for biofuels production: From bugs to synthetic biology to fuels. Curr. Opin. Biotechnol. 2008, 19, 556–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, N.; Hills, C.D.; Singh, R.S.; Atkinson, C.J. Biomass waste utilisation in low-carbon products: Harnessing a major potential resource. Climate and Atmospheric Science 2019, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Antizar-Ladislao, B.; Khraisheh, M. Bioconversion of municipal solid waste to glucose for bio-ethanol production. Bioprocess. Biosyst. Eng. 2007, 30, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.W.; Paari, K.A.; Malaviya, A.; Jang, Y.S. Synthetic Biology Tools for Genome and Transcriptome Engineering of Solventogenic Clostridium. Front. Bioeng. Biotechnol. 2020, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Köpke, M.; Held, C.; Hujer, S.; Liesegang, H.; Wiezer, A.; Wollherr, A.; Ehrenreich, A.; Liebl, W.; Gottschalk, G.; Dürre, P. Clostridium ljungdahlii represents a microbial production platform based on syngas. Proc. Natl. Acad Sci. USA 2010, 107, 13087–13092. [Google Scholar] [CrossRef] [Green Version]
- Kirby, J.; Keasling, J.D. Biosynthesis of plant isoprenoids: Perspectives for microbial engineering. Annu. Rev. Plant. Biol. 2009, 60, 335–355. [Google Scholar] [CrossRef]
- Harvey, B.; Wright, M.; Quintana, R. High-density renewable fuels based on the selective dimerization of pinenes. Energy Fuels 2010, 24, 267–273. [Google Scholar] [CrossRef]
- Yuzawa, S.; Kim, W.; Katz, L.; Keasling, J. Heterologous production of polyketides by modular type I polyketide synthases in Escherichia coli. Curr. Opin. Biotechnol. 2012, 23, 727–735. [Google Scholar] [CrossRef]
- Hwang, S.; Lee, N.; Cho, S.; Palsson, B.; Cho, B.K. Repurposing Modular Polyketide Synthases and Non-ribosomal Peptide Synthetases for Novel Chemical Biosynthesis. Front. Mol. Biosci. 2020, 7, 87. [Google Scholar] [CrossRef]
- Shaw, A.J.; Lam, F.H.; Hamilton, M.; Consiglio, A.; MacEwen, K.; Brevnova, E.E.; Greenhagen, E.; LaTouf, W.G.; South, C.R.; van Dijken, H.; et al. Metabolic engineering of microbial competitive advantage for industrial fermentation processes. Science 2016, 353, 583–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westfall, P.; Gardner, T. Industrial fermentation of renewable diesel fuels. Curr. Opin. Biotechnol. 2011, 22, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Loh, J.; Aves, S.J.; Howard, T.P. Alkane biosynthesis in bacteria. Biogenesis of Hydrocarbons. In Handbook of Hydrocarbon and Lipid Microbiology; Stams, A., Sousa, D., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Jovanovic Tews, I.; Jones, S.B.; Santosa, D.M.; Ziyu, D.; Ramasamy, K.; Zhu, Y. A survey of opportunities for microbial conversion of biomass to hydrocarbon compatible fuels. Pac. Northwest Natl. Lab. 2010, 19704, 01–33. [Google Scholar]
- Li, Q.; Cai, H.; Hao, B.; Zhang, C.; Yu, Z.; Zhou, S.; Chenjuan, L. Enhancing Clostridial acetone-butanol-ethanol (ABE) production and improving fuel properties of ABE-enriched biodiesel by extractive fermentation with biodiesel. Appl. Biochem. Biotechnol. 2010, 162, 2381–2386. [Google Scholar] [CrossRef]
- Alper, H.; Moxley, J.; Nevoigt, E.; Fink, G.; Stephanopoulos, G. Engineering yeast transcription machinery for improved ethanol tolerance and production. Science 2006, 314, 1565–1568. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Yu, H.; Zhu, K. Improving alkane synthesis in Escherichia coli via metabolic engineering. Appl. Microbiol. Biotechnol. 2015, 100, 757–767. [Google Scholar] [CrossRef]
- Ezeji, T.; Qureshi, N.; Blaschek, H.P. Bioproduction of butanol from biomass: From genes to bioreactors. Curr. Opin. Biotechnol. 2007, 18, 220–227. [Google Scholar] [CrossRef]
- Ladygina, N.; Dedyukhina, E.G.; Vainshtein, M.B. A review on microbial synthesis of hydrocarbons. Process Biochem. 2006, 41, 1001–1014. [Google Scholar] [CrossRef]
- Davison, J. Genetic exchange between bacteria in the environment. Plasmid 1999, 42, 73–91. [Google Scholar] [CrossRef]
- Torres, B.; Garcia, J.; Diaz, E. Plasmids as tools for containment. In Plasmid Biology; Funnell, B., Phillips, G., Eds.; ASM: Washington, DC, USA, 2003; pp. 589–601. [Google Scholar]
- Davison, J. Genetic tools for Pseudomonads, Rhizobia and other Gram-negative bacteria. Biotechniques 2002, 32, 386–401. [Google Scholar] [CrossRef] [Green Version]
- Davison, J. Towards safer vectors for field release of recombinant bacteria. Environ. Biosafety Res. 2002, 1, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Secretariat of the Convention on Biological Diversity (2000). Cartagena Protocol on Biosafety to the Convention on Biological Diversity: Text and annexes. Montreal 2000, 1–19, ISBN 92-807-1924-6. [Google Scholar]
- Food and Agriculture Organisation of the United Nations. International Plant Protection Convention. Available online: https://www.ippc.int/en/ (accessed on 31 August 2021).
- Andrianantoandro, E.; Basu, S.; Karig, D.; Weiss, R. Synthetic biology: New engineering rules for an emerging discipline. Mol. Syst. Biol. 2006, 2, 1744–4292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfleger, B.F.; Pitera, D.J.; Smolke, C.D.; Keasling, J.D. Combinatorial engineering of intergenic regions in operons tunes expression of multiple genes. Nat. Biotechnol. 2006, 24, 1027–1032. [Google Scholar] [CrossRef]
- Li, M.; Elledge, S. Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC. Nat. Methods 2007, 4, 251–256. [Google Scholar] [CrossRef]
- Shetty, R.; Endy, D.; Knight, T. Engineering BioBrick vectors from BioBrick parts. J. Biol. Eng. 2008, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Carbonell, P.; Le Feuvre, R.; Takano, E.; Scrutton, N.S. In silico design and automated learning to boost next-generation smart biomanufacturing. Synth. Biol. 2020, 5, ysaa020. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Phulara, S.C. Metabolic engineering for isoprenoid-based biofuel production. J. Appl. Microbiol. 2015, 119, 605–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hull, A.; Golubkov, I.; Kronberg, B.; Marandzheva, T.; van Stam, J. An alternative fuel for spark ignition engines. Int. J. Engine Res. 2006, 7, 203–214. [Google Scholar] [CrossRef]
- Withers, S.; Gottlieb, S.; Lieu, B.; Newman, J.; Keasling, J. Identification of isopentenol biosynthetic genes from Bacillus subtilis by a screening method based on isoprenoid precursor toxicity. Appl. Environ. Microbiol. 2007, 73, 6277–6283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yoon, S.H.; Shah, A.A.; Chung, Y.R.; Kim, J.Y.; Choi, E.S.; Keasling, J.D.; Kim, S.W. Farnesol production from Escherichia coli by harnessing the exogenous mevalonate pathway. Biotechnol. Bioeng. 2010, 107, 421–429. [Google Scholar] [CrossRef]
- Meadows, C.; Kang, A.; Lee, T. Metabolic engineering for advanced biofuels production and recent advances toward commercialization. Biotechnol. J. 2018, 13, 1600433. [Google Scholar] [CrossRef]
- Dugar, D.; Stephanopoulos, G. Relative potential of biosynthetic pathways for biofuels and bio-based products. Nat. Biotechnol. 2011, 29, 1074–1078. [Google Scholar] [CrossRef]
- Krishnan, A.; Bonnie, A.; McNeil, B.A.; Stuart, D.T. Biosynthesis of Fatty Alcohols in Engineered Microbial Cell Factories: Advances and Limitations. Front. Bioeng. Biotechnol. 2020, 8, 610936. [Google Scholar] [CrossRef]
- Röttig, A.; Wenning, L.; Bröker, D.; Steinbüchel, A. Fatty acid alkyl esters: Perspectives for production of alternative biofuels. Appl. Microbiol. Biotechnol. 2009, 85, 1713–1733. [Google Scholar] [CrossRef] [PubMed]
- Lennen, R.; Braden, D.; West, R.; Dumesic, J.; Pfleger, B. A process for microbial hydrocarbon synthesis: Overproduction of fatty acids in Escherichia coli and catalytic conversion to alkanes. Biotechnol. Bioeng. 2010, 106, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Vora, H.; Khosla, C. Quantitative analysis and engineering of fatty acid biosynthesis in E. coli. Metab. Eng. 2010, 12, 378–386. [Google Scholar] [CrossRef]
- Pfleger, B.F.; Gossing, M.; Nielsen, J. Metabolic engineering strategies for microbial synthesis of oleochemicals. Metab. Eng. 2015, 29, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Matsuoka, H.; Hirooka, K. Regulation of fatty acid metabolism in bacteria. Mol. Microbiol. 2007, 66, 829–839. [Google Scholar] [CrossRef]
- Zhang, F.; Carothers, J.M.; Keasling, J.D. Design of a dynamic sensor-regulator system for production of chemicals and fuels derived from fatty acids. Nat. Biotechnol. 2012, 30, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, C.; Feng, D.; Cheng, T.; Meng, X.; Liu, W.; Zou, H.; Xian, M. Production of extracellular fatty acid using engineered Escherichia coli. Microb. Cell Fact. 2012, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Dellomonaco, C.; Clomburg, J.M.; Miller, E.N.; Gonzalez, R. Engineered reversal of the beta-oxidation cycle for the synthesis of fuels and chemicals. Nature 2011, 476, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Bowen, C.H.; Liu, D.; Zhang, F.Z. Exploiting nongenetic cell-to-cell variation for enhanced biosynthesis. Nat. Chem. Biol. 2016, 12, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Rude, M.; Schirmer, A. New microbial fuels: A biotech perspective. Curr. Opin. Microbiol. 2009, 12, 274–281. [Google Scholar] [CrossRef]
- Beller, H.; Goh, E.; Keasling, J. Genes involved in long-chain alkene biosynthesis in Micrococcus luteus. Appl. Environ. Microbiol. 2009, 76, 1212–1223. [Google Scholar] [CrossRef] [Green Version]
- Schirmer, A.; Rude, M.A.; Li, X.; Popova, E.; del Cardayre, S.B. Microbial biosynthesis of alkanes. Science 2010, 329, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Rude, M.; Baron, T.; Brubaker, S.; Alibhai, M.; del Cardayre, S.; Schirmer, A. Terminal olefin (1-alkene) biosynthesis by a novel P450 Fatty Acid Decarboxylase from Jeotgalicoccus Species. Appl. Environ. Microbiol. 2011, 77, 1718–1727. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Díaz, L.; Caballero, A.; Pérez-Hernández, N.; Segura, A. Microbial alkane production for jet fuel industry: Motivation, state of the art and perspectives. Microb. Biotechnol. 2016, 10, 103–124. [Google Scholar] [CrossRef]
- Rabinovitch-Deere, C.A.; Oliver, J.W.K.; Rodriguez, G.M.; Atsumi, S. Synthetic biology and metabolic engineering approaches to produce biofuels. Chem. Rev. 2013, 113, 4611–4632. [Google Scholar] [CrossRef]
- Atsumi, S.; Cann, A.F.; Connor, M.R.; Shen, C.R.; Smith, K.M.; Brynildsen, M.P.; Chou, K.J.; Hanai, T.; Liao, J.C. Metabolic engineering of Escherichia coli for 1-butanol production. Metab. Eng. 2008, 10, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Wu, T.Y.; Eckl, E.M.; Hawkins, S.D.; Buelter, T.; Liao, J.C. Engineering the isobutanol biosynthetic pathway in Escherichia coli by comparison of three aldehyde reductase/alcohol dehydrogenase genes. Appl. Microbiol. Biotechnol. 2010, 85, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Bastian, S.; Liu, X.; Meyerowitz, J.T.; Snow, C.D.; Chen, M.M.Y.; Arnold, F.H. Engineered ketol-acid reductoisomerase and alcohol dehydrogenase enable anaerobic 2-methylpropan1-ol production at theoretical yield in Escherichia coli. Metab. Eng. 2011, 13, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.R.; Lan, E.I.; Dekishima, Y.; Baez, A.; Cho, K.M.; Liao, J.C. Driving forces enable hightiter anaerobic 1-butanol synthesis in Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 2905–2915. [Google Scholar] [CrossRef] [Green Version]
- Lian, J.Z.; Si, T.; Nair, N.U.; Zhao, H.M. Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains. Metab. Eng. 2014, 24, 139–149. [Google Scholar] [CrossRef]
- Si, T.; Luo, Y.Z.; Xiao, H.; Zhao, H.M. Utilizing an endogenous pathway for 1- butanol production in Saccharomyces cerevisiae. Metab. Eng. 2014, 22, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Gleizer, S.; Ben-Nissan, R.; Bar-On, Y.M.; Antonovsky, N.; Noor, E.; Zohar, Y.; Jona, G.; Krieger, E.; Shamshoum, M.; Bar-Even, A.; et al. Conversion of Escherichia coli to generate all biomass carbon from CO2. Cell 2019, 179, 1255–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, J.E.; Jang, Y.S. Metabolic engineering of microorganisms for the production of ethanol and butanol from oxides of carbon. Appl. Microbiol. Biotechnol. 2019, 103, 8283–8292. [Google Scholar] [CrossRef]
- Antonovsky, N.; Gleizer, S.; Noor, E.; Zohar, Y.; Herz, E.; Barenholz, U.; Zelcbuch, L.; Amram, S.; Wides, A.; Tepper, N.; et al. Sugar synthesis from CO2 in Escherichia coli. Cell 2016, 166, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Guadalupe-Medina, V.; Wisselink, H.W.; Luttik, M.A.; de Hulster, E.; Daran, J.M.; Pronk, J.T.; van Maris, A.J. Carbon dioxide fixation by Calvin-Cycle enzymes improves ethanol yield in yeast. Biotechnol. Biofuels 2013, 6, 125. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Opgenorth, P.H.; Wernick, D.G.; Rogers, S.; Wu, T.Y.; Higashide, W.; Malati, P.; Huo, Y.X.; Cho, K.M.; Liao, J.C. Integrated electromicrobial conversion of CO2 to higher alcohols. Science 2012, 335, 1596. [Google Scholar] [CrossRef]
- Atsumi, S.; Hanai, T.; Liao, J. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 2008, 451, 86–89. [Google Scholar] [CrossRef]
- Larson, E.D. Biofuel Production Technologies: Status, Prospects and Implications for Trade and Development; United Nations Conference on Trade and Development (UNCTAD): New York, NY, USA, 2008. [Google Scholar]
- Sánchez, O.J.; Cardona, C.A. Trends in biological production of fuel ethanol from different feedstocks. Bioresour Technol. 2008, 99, 5270–5295. [Google Scholar] [CrossRef] [PubMed]
- Hosseini Koupaie, E.; Dahadha, S.; Bazyar Lakeh, A.A.; Azizi, A.; Elbeshbishy, E. Enzymatic pretreatment of lignocellulosic biomass for enhanced biomethane production-A review. J. Environ. Manag. 2019, 233, 774–784. [Google Scholar] [CrossRef]
- Taherzadeh, M.J.; Karimi, K. Pretreatment of lignocellulosic wastes to improve ethanol and biogas production: A review. Int. J. Mol. Sci. 2008, 9, 1621–1651. [Google Scholar] [CrossRef] [Green Version]
- Saritha, M.; Arora, A.; Lata. Biological pretreatment of lignocellulosic substrates for enhanced delignification and enzymatic digestibility. Indian J. Microbiol. 2011, 52, 122–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, H.; Jensen, P.; Batstone, D. Pre-treatment mechanisms during thermophilic–mesophilic temperature phased anaerobic digestion of primary sludge. Water Res. 2010, 44, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Alvira, P.; Tomás-Pejó, E.; Ballesteros, M.; Negro, M. Pretreatment technologies for an efficient bioethanol production process based on enzymatic hydrolysis: A review. Bioresour Technol. 2010, 101, 4851–4861. [Google Scholar] [CrossRef]
- Sharma, H.; Xu, C.; Qin, W. Biological pretreatment of lignocellulosic biomass for biofuels and bioproducts: An overview. Waste Biomass Valorization 2017, 10, 235–251. [Google Scholar] [CrossRef]
- Mishra, S.; Singh, P.K.; Dash, S.; Pattnaik, R. Microbial pretreatment of lignocellulosic biomass for enhanced biomethanation and waste management. 3 Biotech 2018, 8, 458. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Xu, F.; Li, Y. Pretreatment of lignocellulosic biomass for enhanced biogas production. Prog. Energy Combust. Sci. 2014, 42, 35–53. [Google Scholar] [CrossRef]
- Isroi, I.; Millati, R.; Syamsiah, S.; Niklasson, C.; Cahyanto, M.N.; Ludquist, K. Biological pretreatment of lignocelluloses with white-rot fungi and its applications: A review. BioResources 2011, 6, 5224–5259. [Google Scholar] [CrossRef]
- Patel, H.; Gupte, A.; Gupte, S. Effect of different culture conditions and inducers on production of laccase by a basidiomycete fungal isolate Pleurotus ostreatus HP-1 under solid state fermentation. BioResources 2009, 4, 268–284. [Google Scholar]
- Romano, R.; Zhang, R.; Teter, S.; McGarvey, J. The effect of enzyme addition on anaerobic digestion of Jose Tall Wheat Grass. Bioresour. Technol. 2009, 100, 4564–4571. [Google Scholar] [CrossRef]
- Yunqin, L.; Dehan, W.; Lishang, W. Biological pretreatment enhances biogas production in the anaerobic digestion of pulp and paper sludge. Waste Manag. Res. 2010, 28, 800–810. [Google Scholar] [CrossRef]
- Ziemiński, K.; Romanowska, I.; Kowalska, M. Enzymatic pretreatment of lignocellulosic wastes to improve biogas production. Waste Manag. 2012, 32, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Mackuľak, T.; Prousek, J.; Švorc, Ľ.; Drtil, M. Increase of biogas production from pretreated hay and leaves using wood-rotting fungi. Chem. Papers 2012, 66, 649–653. [Google Scholar] [CrossRef]
- Tangnu, S.; Blanch, H.; Wilke, C. Enhanced production of cellulase, hemicellulase, and ?-glucosidase by Trichoderma reesei (Rut C-30). Biotechnol. Bioeng. 1981, 23, 1837–1849. [Google Scholar] [CrossRef]
- Shi, J.; Sharma-Shivappa, R.; Chinn, M.; Howell, N. Effect of microbial pretreatment on enzymatic hydrolysis and fermentation of cotton stalks for ethanol production. Biomass Bioenergy 2009, 33, 88–96. [Google Scholar] [CrossRef]
- Liu, J.; Wang, M.; Tonnis, B.; Habteselassie, M.; Liao, X.; Huang, Q. Fungal pretreatment of switchgrass for improved saccharification and simultaneous enzyme production. Bioresour. Technol. 2013, 135, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Silveira, M.H.L.; Morais, A.R.C.; da Costa Lopes, A.M.; Olekszyszen, D.N.; Bogel-Łukasik, R.; Andreaus, J.; Pereira Ramos, L. Current pretreatment technologies for the development of cellulosic ethanol and biorefineries. ChemSusChem 2015, 8, 3366–3390. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Yang, N.; Xu, C.; Yu, H.; Wu, J.; Zhang, X. Combination of biological pretreatment with mild acid pretreatment for enzymatic hydrolysis and ethanol production from water hyacinth. Bioresour. Technol. 2010, 101, 9600–9604. [Google Scholar] [CrossRef] [PubMed]
- Shirkavand, E.; Baroutian, S.; Gapes, D.J.; Young, B.R. Combination of fungal and physicochemical processes for lignocellulosic biomass pretreatment—a review. Renew. Sustain. Energy Rev. 2016, 54, 217–234. [Google Scholar] [CrossRef]
- Shi, Y.; Chai, L.; Tang, C.; Yang, Z.; Zhang, H.; Chen, R.; Chen, Y.; Zheng, Y. Characterization and genomic analysis of kraft lignin biodegradation by the beta-proteobacterium Cupriavidus basilensis B-8. Biotechnol. Biofuels 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Chai, L.; Tang, C.; Yang, Z.; Zheng, Y.; Shi, Y.; Zhang, H. Kraft lignin biodegradation by Novosphingobium sp. B-7 and analysis of the degradation process. Bioresour. Technol. 2012, 123, 682–685. [Google Scholar] [CrossRef]
- Zhuo, S.; Yan, X.; Liu, D.; Si, M.; Zhang, K.; Liu, M.; Peng, B.; Shi, Y. Use of bacteria for improving the lignocellulose biorefinery process: Importance of pre-erosion. Biotechnol. Biofuels 2018, 11, 146. [Google Scholar] [CrossRef] [Green Version]
- Masai, E.; Katayama, Y.; Fukuda, M. Genetic and biochemical investigations on bacterial catabolic pathways for lignin-derived aromatic compounds. Biosci. Biotechnol. Biochem. 2007, 71, 1–15. [Google Scholar] [CrossRef]
- Maki, M.; Leung, K.T.; Qin, W. The prospects of cellulase-producing bacteria for the bioconversion of lignocellulosic biomass. Int. J. Biol. Sci. 2009, 5, 500–516. [Google Scholar] [CrossRef] [PubMed]
- Miron, J.; Ben-Ghedalia, D.; Morrison, M. Invited review: Adhesion mechanisms of rumen cellulolytic bacteria. J. Dairy Sci. 2001, 84, 1294–1309. [Google Scholar] [CrossRef]
- Duff, S.; Murray, W. Bioconversion of forest products industry waste cellulosics to fuel ethanol: A review. Bioresour. Technol. 1996, 55, 1–33. [Google Scholar] [CrossRef]
- Dien, B.; Cotta, M.; Jeffries, T. Bacteria engineered for fuel ethanol production: Current status. Appl. Microbiol. Biotechnol. 2003, 63, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Yang, Y.; Liu, C.C.; Yang, S.; Feng-Wu Bai, F.W. Engineering Zymomonas mobilis for Robust Cellulosic Ethanol Production. Trends Biotechnol. 2019, 37, 960. [Google Scholar] [CrossRef]
- de Gonzalo, G.; Colpa, D.; Habib, M.; Fraaije, M. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Chandra, R.; Chowdhary, P. Properties of bacterial laccases and their application in bioremediation of industrial wastes. Environ. Sci. Process. Impacts 2015, 17, 326–342. [Google Scholar] [CrossRef]
- Picart, P.; de María, P.; Schallmey, A. From gene to biorefinery: Microbial β-etherases as promising biocatalysts for lignin valorization. Front. Microbiol. 2015, 6, 00916. [Google Scholar] [CrossRef] [Green Version]
- Bala, R.; Mondal, M. Study of biological and thermo-chemical pretreatment of organic fraction of municipal solid waste for enhanced biogas yield. Environ. Sci. Pollut. Res. 2019, 27, 27293–27304. [Google Scholar] [CrossRef]
- Xu, S.; Kong, X.; Liu, J.; Zhao, K.; Zhao, G.; Bahdolla, A. Effects of high-pressure extruding pretreatment on MSW upgrading and hydrolysis enhancement. Waste Manag. 2016, 58, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Bala, R.; Gautam, V.; Mondal, M. Improved biogas yield from organic fraction of municipal solid waste as preliminary step for fuel cell technology and hydrogen generation. Int. J. Hydrogen Energy 2019, 44, 164–173. [Google Scholar] [CrossRef]
- Li, Y.; Jin, Y. Effects of thermal pretreatment on acidification phase during two-phase batch anaerobic digestion of kitchen waste. Renew. Energy 2015, 77, 550–557. [Google Scholar] [CrossRef]
- Fdez.-Güelfo, L.; Álvarez-Gallego, C.; Sales Márquez, D.; Romero García, L. Biological pretreatment applied to industrial organic fraction of municipal solid wastes (OFMSW): Effect on anaerobic digestion. Chem. Eng. J. 2011, 172, 321–325. [Google Scholar] [CrossRef]
- Tyagi, V.; Lo, S.; Rajpal, A. Chemically coupled microwave and ultrasonic pre-hydrolysis of pulp and paper mill waste-activated sludge: Effect on sludge solubilisation and anaerobic digestion. Environ. Sci. Pollut. Res. 2014, 21, 6205–6217. [Google Scholar] [CrossRef]
- Rudakiya, D.; Gupte, A. Degradation of hardwoods by treatment of white rot fungi and its pyrolysis kinetics studies. Int. Biodeterior. Biodegrad. 2017, 120, 21–35. [Google Scholar] [CrossRef]
- Fdez-Güelfo, L.; Álvarez-Gallego, C.; Sales, D.; Romero, L. The use of thermochemical and biological pretreatments to enhance organic matter hydrolysis and solubilization from organic fraction of municipal solid waste (OFMSW). Chem. Eng. J. 2011, 168, 249–254. [Google Scholar] [CrossRef]
- Cammarota, M.; Teixeira, G.; Freire, D. Enzymatic pre-hydrolysis and anaerobic degradation of wastewaters with high oil contents. Biotechnol. Lett. 2001, 23, 1591–1595. [Google Scholar] [CrossRef]
- Koltermann, A.; Kraus, M.; Rarbach, M.; Reisinger, C.; Zavrel, M.; Soltl, Y. Cellulosic ethanol from agricultural residues–an advanced biofuel and biobased chemical platform. JSM Biotechnol. Bioeng. 2014, 2, 1024. [Google Scholar]
- Verma, A.; Kumar, S.; Jain, P.K. Key pre-treatment technologies on cellulosic ethanol production. J. Sci. Res. 2011, 55, 57–63. [Google Scholar]
- Kumar, P.D.; Barrett, D.M.; Delwiche, M.J.; Stroeve, P. Methods for pretreatment of lignocellulosic biomass for efficient hydrolysis and biofuel production. Ind. Eng. Chem. Res. 2009, 48, 3713–3729. [Google Scholar] [CrossRef]
- Lee, J. Biological conversion of lignocellulosic biomass to ethanol. J. Biotechnol. 1997, 56, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Ray, M.J.; Leak, D.J.; Spanu, P.D.; Murphy, R.J. Brown rot fungal early stage decay mechanism as a biological pretreatment for softwood biomass in biofuel production. Biomass Bioenerg. 2010, 34, 1257–1262. [Google Scholar] [CrossRef]
- Sanchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Dashtban, M.; Schraft, H.; Qin, W. Fungal bioconversion of lignocellulosic residues; Opportunities & perspectives. Int. J. Biol Sci. 2009, 5, 578–595. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Kathiresan, K. Bioconversion of lignocellulosic waste to bioethanol by Trichoderma and yeast fermentation. Biotech 2013, 4, 493–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramaniam, M.K.; Rajarathinam, R. Implementation of white rot fungal pretreated rice straw for sustainable bioethanol production by Saccharomyces cerevisiae. Int. J. Eng. Res. Technol. 2013, 2, 4047–4053. [Google Scholar]
- Eggeman, T.; Elander, R.T. Process and economic analysis of pretreatment technologies. Bioresour. Technol. 2005, 96, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.R.; Khalid, H.; Zhang, H.; Rahman, S.; Zhang, R.; Liu, G.; Chen, C. Pretreatment methods of lignocellulosic biomass for anaerobic digestion. AMB Exp. 2017, 7, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghani, M.; Karimi, K.; Sadeghi, M. Pretreatment of Rice Straw for the Improvement of Biogas Production. Energy Fuels 2015, 29, 3770–3775. [Google Scholar] [CrossRef]
- Galbe, M.; Wallberg, O. Pretreatment for biorefineries: A review of common methods for efficient utilization of lignocellulosic materials. Biotechnol. Biofuels 2019, 12, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baruah, J.; Nath, B.K.; Sharma, R.; Sachin Kumar, S.; Deka, R.C.; Baruah, D.C.; Kalita, E. Recent Trends in the Pretreatment of Lignocellulosic Biomass for Value-Added Products. Front. Energy Res. 2018, 6, 141. [Google Scholar] [CrossRef]
- Anitescu, G.; Bruno, T.J. Liquid biofuels: Fluid properties to optimize feedstock selection, processing, refining/blending, storage/transportation, and combustion. Energy Fuels 2012, 26, 324–348. [Google Scholar] [CrossRef]
- Keasling, J.D. Manufacturing molecules through metabolic engineering. Science 2010, 330, 1355–1358. [Google Scholar] [CrossRef]
- Peralta-Yahya, P.P.; Ouellet, M.; Chan, R.; Mukhopadhyay, A.; Keasling, J.D.; Lee, T.S. Identification and microbial production of a terpene-based advanced biofuel. Nat. Commun. 2011, 2, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, D. Microbes to Generate Electricity. Indian J. Microbiol. 2013, 53, 120–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Class | Range | Concentration (g/L) | Reference |
|---|---|---|---|---|
| E. coli BL21(DE3) (modified) | Alkanes | C13, C15, and C17 | 0.30 | [29] |
| C. acetobutylicum PJC4BK (GMM) | Mixed Oxygenates | acetone-butanol-ethanol | 25–33 | [30] |
| C. acetobutylicum P260 (GMM) | Mixed Oxygenates | acetone-butanol-ethanol | 25–34 | [30] |
| C. beijerinckii BA101 (GMM) | Mixed Oxygenates | acetone-butanol-ethanol | 25–35 | [30] |
| C. beijerinckii BA101 (GMM) | Oxygenates alcohol | butanol | 11.9–14.3 | [30] |
| N. muscorum | Alkanes | n-C15-C18 | 0.025–0.12 | [31] |
| Rhodopseudomonas | Isoprenoids | n-C15-C20 | 0.006 | [31] |
| D. desulfuricans | Alkanes | n-C11-C35 | 0.8–2.25 | [31] |
| Bacillus sp. | Mixed | C14-C34 | 0.33 | [31] |
| Pretreatment Method | Considerations | Pretreatment Conditions | Results | Ref |
|---|---|---|---|---|
| Physical | Physical pretreatment techniques have been shown to increase the available surface area and pore size necessary for successful enzymatic reactions. The majority of the techniques used, however, are unable to remove the lignin. Before applying thermal or chemical pretreatments, physical techniques are usually employed as a step. | Feedstock: Cellulose, hemicellulose. Conditions: Mechanical pulverization, particle sizes of 33 to 6 mm | Increased methane yield 11–13% Energy cost is high; particle diameter should be 6 mm for high methane yield | [131] |
| Chemical | Chemical techniques have shown to be highly efficient in expanding the available surface area of lignocellulosic biomass, as well as providing partial-to-complete delignification. It is also possible to obtain a considerable reduction in cellulose crystallinity, as well as partial-to-total hemicellulose solubilization. Pretreatment using chemicals is usually linked with the greatest pretreatment rate. The most significant drawback of chemical pretreatment techniques is the significant chemical consumption linked with high costs and environmental issues. | Feedstock: Rice straw Conditions: 0.25 and 0.5 M of sodium carbonate at 90, 110, and 130 °C and for 1, 2, and 3 h | The rice straw pretreated with 0.5 M of sodium carbonate at 110 °C for 2 h demonstrated the highest improvement among the pretreatments, resulting in the production of 292 mL/g of methane (125% improvement compared with the untreated straw). | [132] |
| Thermal | Previous study has shown that heat pretreatment can reduce cellulose crystallinity, lower the degree of polymerization, and solubilize lignin. When combined with high pressure, the impact of temperature has been observed to be superior. The thermal method’s high energy-de-manding nature is the primary impediment to its industrial use. The combined thermos-chemical pretreatment has proven to be a viable alternative to just thermal and chemical pretreatments, combining the benefits of both procedures while reducing chemical and energy usage. | Feedstock: Wheat straw/corn stover (mixed)Conditions: Soaking 0.2% H2SO4, 190 °C, 5 min. | >50 g L−1 Ethanol > 50 g/L Overall yield 74–78% | [133] |
| Biological | Biological techniques have proven to be highly effective in decreasing the degree of cellulose polymerization, hydrolyzing hemicellulose, and delignification. The advantages of this approach over other pretreatment methods include a minimal chemical need and low energy usage. They are recognized for being the most eco-friendly pretreatment method. Biological techniques, on the other hand, are typically limited in their commercial use due to their poor hydrolysis rate and high cost (when specific enzymes are used). | Feedstock: Wheat straw Microorganism/Enzyme: Ceriporiopsis subvermispora Conditions: 7 weeks of solid-state fermentation incubated at 24 °C. | In vitro gas production of 297 mL/g. | [134] |
| Enzymatic | Recent research has focused more emphasis on the enzymatic pretreatment approach because of the potential it offers for increasing biofuel production, particularly biomethane synthesis. The enzymatic pretreatment of lignocellulosic biomass employs oxidative and hydrolytic enzymes generated mostly by bacteria and fungi. This pre-treatment approach is gaining popularity because of the quick reaction time, low nutrient requirements for enzymatic reactions, and the fact that most enzymes are unaffected by inhibitors and other microbial metabolisms. Furthermore, the enzymatic pretreatment does not necessitate the use of costly processing equipment. However, the high cost of enzymes continues to be a barrier to the economic viability of this pretreatment approach for increased fuel generation on a large scale. | Feedstock: Agricultural residue; sugar beet pulp, spent hops, manure, biofiber, MSW, paper pulp, wheat grass. Microorganism/Enzyme: Laccase, mixture of cellulases, hemicellulases and β-glucosidase, pectinase, Trichoderma enzyme complex Conditions: 37 °C for 4–24 h. Autoclave. Aerobic/anaerobic | 0–34% improved methane production | [85] |
| Fungal | Fungal pretreatment utilizing lignin-degrading microorganisms, such as preliminary white-rot fungi, has resurfaced as a viable option to thermal/chemical pretreatment for cellulosic ethanol production, both economically and environmentally. It has been demonstrated that fungal pretreatment may improve the enzymatic digestibility of a variety of biomass feedstocks, including maize stover, wheat straw, rice straw, cotton stalks, and woody biomass. Simple procedures, minimal energy needs, decreased waste streams, lower downstream processing costs, and reduced inhibitors to ethanol fermentation are all advantages of this approach over thermochemical pretreatments. Despite the benefits, the main drawbacks of fungal pretreatment include significant cellulose and hemicellulose loss and a protracted pretreatment period. | Feedstock: Agricultural residue; chestnut leaves and sisal leaves Microorganism/Enzyme: Fungi (white, brown, and soft rot) and Basidiomycetes Conditions: 28–37 °C for 12 days to 8 weeks. Autoclave/no autoclave. Aerobic | 15% to fivefold increased methanation | [85] |
| Bacterial | Bacterial pretreatment is a screening and application method that uses microorganisms isolated from the natural environment to aid in the hydrolysis of biomass during anaerobic digestion. Bacterial consortia used in pretreatment should obviously have the ability to degrade cellulose and hemicellulose. In anaerobic digestion of distillery wastewater-fortified cassava waste, a bacterial community isolated from a thermophilic straw-decomposing landfill exhibited a 96% greater methanation at 55 °C than its untreated equivalent. | Feedstock: Agricultural residue; corn straw, corn stalk, cassava residue, and cotton stalk. Microorganism/Enzyme: Complex mixture containing yeast and cellulolytic bacteria, heat-treated sludge, Clostridium thermocellum, mixture of fungi and compost-forming microorganism. Conditions: 20–55 °C for 12 h to 20 days. Autoclave/no autoclave. Aerobic | 25–96.63% enhanced methane production | [85] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azimov, U.; Okoro, V.; Hernandez, H.H. Recent Progress and Trends in the Development of Microbial Biofuels from Solid Waste—A Review. Energies 2021, 14, 6011. https://doi.org/10.3390/en14196011
Azimov U, Okoro V, Hernandez HH. Recent Progress and Trends in the Development of Microbial Biofuels from Solid Waste—A Review. Energies. 2021; 14(19):6011. https://doi.org/10.3390/en14196011
Chicago/Turabian StyleAzimov, Ulugbek, Victor Okoro, and Hector H. Hernandez. 2021. "Recent Progress and Trends in the Development of Microbial Biofuels from Solid Waste—A Review" Energies 14, no. 19: 6011. https://doi.org/10.3390/en14196011
APA StyleAzimov, U., Okoro, V., & Hernandez, H. H. (2021). Recent Progress and Trends in the Development of Microbial Biofuels from Solid Waste—A Review. Energies, 14(19), 6011. https://doi.org/10.3390/en14196011