An Assessment of Methanotrophs Producing Industrial-Grade Lipids for Biofuels and Other Commercial Chemicals



Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Lipids
3. Methanotrophic Bacteria
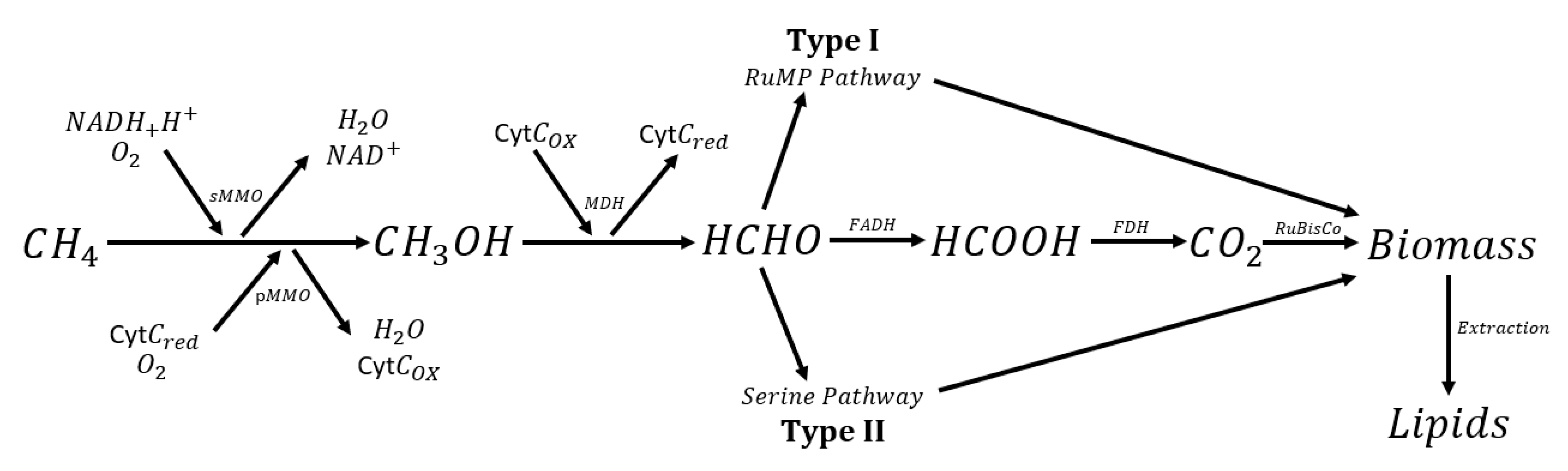
3.1. Biological Processes
3.2. Other Uses of Methanotrophic Bacteria
3.3. Species of Methanotrophs
3.4. Sources of Methanotrophs
4. Cultivation Methods for Methanotrophic Bacteria
5. Lipids Production and Extraction of Lipids from Methanotrophic Bacteria
5.1. Lipids in Methanotrophs
5.2. Microbial Lipid Extraction Techniques
5.3. Lipid Conversion to Biofuels
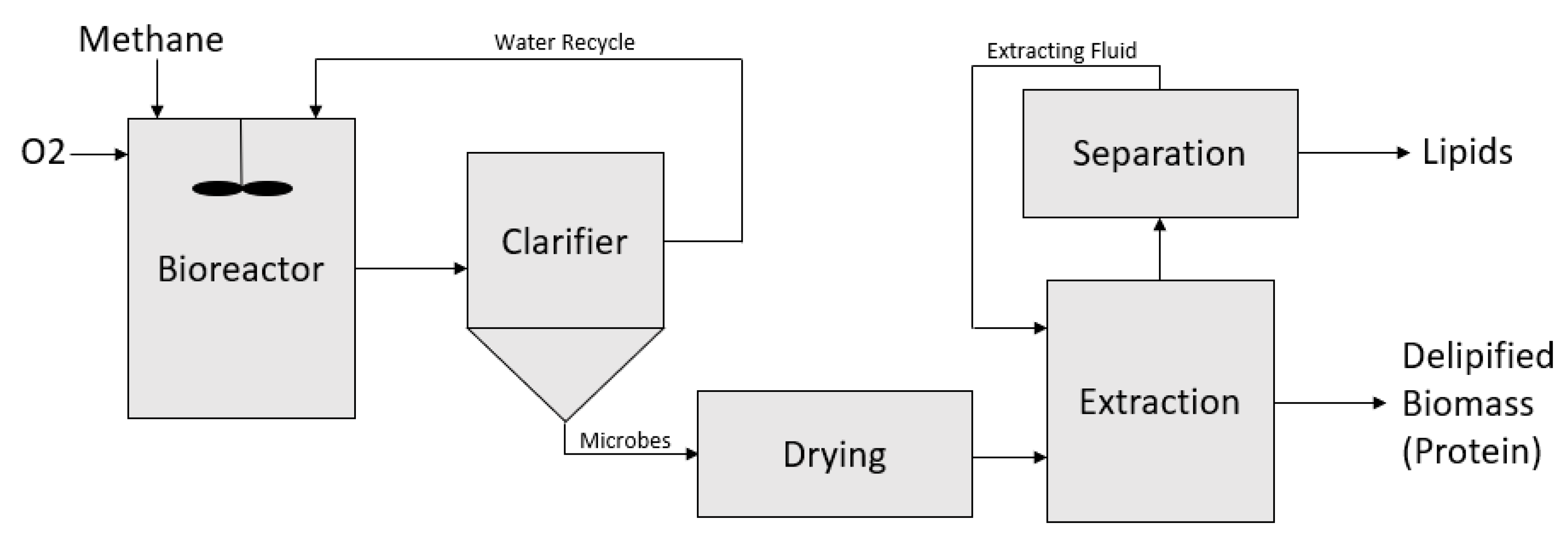
6. Bioreactor Designs and Case Studies Using Methanotrophs
7. Other Products from Methanotrophic Bacteria
8. Challenges of Large-Scale Use of Methanotrophic Bacteria for Lipids and Other Products
8.1. Culturing
8.2. Medium Selection and Application
8.3. Lipid-Extraction Methods
9. Summary
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilman, A.; Laurens, L.M.; Puri, A.W.; Chu, F.; Pienkos, P.T.; Lidstrom, M.E.; Factories, M.C. Bioreactor performance parameters for an industrially-promising methanotroph Methylomicrobium buryatense 5GB1. Microb. Cell Fact. 2015, 14, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henard, C.A.; Franklin, T.G.; Youhenna, B.; But, S.; Alexander, D.; Kalyuzhnaya, M.G.; Guarnieri, M.T. Biogas Biocatalysis: Methanotrophic Bacterial Cultivation, Metabolite Profiling, and Bioconversion to Lactic Acid. Front. Microbiol. 2018, 9, 2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Zhao, X.; Turcotte, F.; Deschênes, J.; Tremblay, R.; Jolicoeur, M. Current lipid extraction methods are significantly enhanced adding a water treatment step in Chlorella protothecoides. Microb. Cell Fact. 2017, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.L.; Preta, G. Lipids in the cell: Organisation regulates function. Cell. Mol. Life Sci. 2018, 75, 1909–1927. [Google Scholar] [CrossRef]
- Cheng, M.H.; Rosentrater, K.A. Profitability analysis of soybean oil processes. Bioengineering 2017, 4, 83. [Google Scholar] [CrossRef] [Green Version]
- Matthaus, B.; Özcan, M.M.; Juhaimi, F.A.l. Some rape/canola seed oils: Fatty acid composition and tocopherols. Z. Fur Nat.-Sect. C J. Biosci. 2016, 71, 73–77. [Google Scholar] [CrossRef]
- Beef Tallow Price-Beef Tallow Price Manufacturers, Suppliers and Exporters on Alibaba.comAnimal Extract. Available online: https://www.alibaba.com/products/beef_tallow_price.html?IndexArea=product_en (accessed on 12 July 2020).
- List, G. Sunflower Seed and Oil. Lipid Technol. 2017, 29, 16. [Google Scholar] [CrossRef] [Green Version]
- US Sunflower Oil Price. Available online: https://ycharts.com/indicators/us_sunflower_oil_price (accessed on 13 July 2020).
- Yamada, M. Lipid Transfer Proteins in Plants and Microorganisms. Plant Cell Physiol. 1992, 33, 1–6. [Google Scholar] [CrossRef]
- Roddy, D.J.; Sayigh, A. Comprehensive Renewable Energy; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Trabold, T.A.; Babbitt, C.W. Sustainable Food Waste-to-Energy Systems; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Hofstrand, D. Tracking Biodiesel Profitability. Available online: https://www.extension.iastate.edu/agdm/energy/html/d1-15.html (accessed on 13 July 2020).
- Monthly Diesel Fuel Retail Prices U.S. 2020|Statista. Available online: https://www.statista.com/statistics/204169/retail-prices-of-diesel-fuel-in-the-united-states-since-2009/ (accessed on 13 July 2020).
- Dahiya, A. Cutting-edge Biofuel Conversion Technologies to Integrate into Petroleum-based Infrastructure and Integrated Biorefineries. In Bioenergy; Elsevier: Amsterdam, The Netherlands, 2015; pp. 467–485. [Google Scholar]
- Rosen, M.J.; Kunjappu, J.T. Surfactants and Interfacial Phenomena; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Surfactants Market Size Growth & Share|Global Industry Report, 2022. Available online: https://www.grandviewresearch.com/industry-analysis/surfactants-market (accessed on 13 July 2020).
- Barnes, D.K.A.; Galgani, F.; Thompson, R.C.; Barlaz, M. Accumulation and fragmentation of plastic debris in global environments. Philos. Trans. R Soc. B Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [Green Version]
- Hanson, R.S.; Hanson, T.E. Methanotrophic bacteria. Microbiol. Rev. 1996, 60, 439–471. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, H.; Yurimoto, H.; Sakai, Y. Interactions of Methylotrophs with Plants and Other Heterotrophic Bacteria. Microorganisms 2015, 3, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Murrell, J.C. Methanotrophy/methane oxidation. In Encyclopedia of Microbiology; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 293–298. [Google Scholar]
- Anthony, C.; York Paris, N.; Diego, S.; Francisco, S. The Biochemistry of Methylotrophs; Academic Press: London, UK; New York, NY, USA, 1982. [Google Scholar]
- Jiang, H.; Chen, Y.; Jiang, P.; Zhang, C.; Smith, T.J.; ColinMurrell, J.; Xing, X.-H. Methanotrophs: Multifunctional bacteria with promising applications in environmental bioengineering. Biochem. Eng. J. 2010, 49, 277–288. [Google Scholar] [CrossRef]
- Canfield, D.E.; Kristensen, E.; Thamdrup, B. Aquatic geomicrobiology (1st edition). Adv. Mar. Biol. 2005. [Google Scholar]
- Aimen, H.; Khan, A.S.; Kanwal, N. Methanotrophs: The Natural Way to Tackle Greenhouse Effect. J. Bioremediat. Biodegrad. 2018, 9. [Google Scholar] [CrossRef]
- Donga, T.; Fei, Q.; Genelot, M.; Smith, H.; Laurens, L.M.L.; Watson, M.J.; Pienkos, P.T. A novel integrated biorefinery process for diesel fuel blendstock production using lipids from the methanotroph, Methylomicrobium buryatense. Energy Convers. Manag. 2017, 140, 62–70. [Google Scholar] [CrossRef]
- Messerschmidt, A. Enzymes and Enzyme Mechanisms. Compr. Nat. Prod. II (1st Edition) 2010. [Google Scholar]
- Theisen, A.R.; Murrell, J.C. Facultative methanotrophs revisited. J. Bacteriol. 2005, 187, 4303–4305. [Google Scholar] [CrossRef] [Green Version]
- Ross, M.O.; Rosenzweig, A.C. A tale of two methane monooxygenases. J. Biol. Inorg. Chem. 2017, 22, 307–319. [Google Scholar] [CrossRef]
- Castresana, J.; Lübben, M.; Saraste, M.; Higgins, D.G. Evolution of cytochrome oxidase, an enzyme older than atmospheric oxygen. EMBO J. 1994, 13, 2516–2525. [Google Scholar] [CrossRef]
- Rasigraf, O.; Kool, D.M.; Jetten, M.S.M.; Damsté, J.S.S.; Ettwig, K.F. Autotrophic carbon dioxide fixation via the Calvin-Benson-Bassham cycle by the denitrifying methanotroph “Candidatus Methylomirabilis oxyfera”. Appl. Environ. Microbiol. 2014, 80, 2451–2460. [Google Scholar] [CrossRef] [Green Version]
- Greenhouse Gas Inventory|California Air Resources Board. Available online: https://www2.arb.ca.gov/our-work/programs/greenhouse-gas-inventory (accessed on 12 July 2020).
- Overview of Greenhouse Gases|Greenhouse Gas (GHG) Emissions|US EPA. Available online: https://www.epa.gov/ghgemissions/overview-greenhouse-gases (accessed on 13 July 2020).
- Pandey, V.C.; Singh, J.S.; Singh, D.P.; Singh, R.P. Methanotrophs: Promising bacteria for environmental remediation. Int. J. Environ. Sci. Technol. 2014, 11, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.S.; Singh, D.P. Methanotrophs: An emerging bioremediation tool with unique broad spectrum methane monooxygenase (MMO) enzyme. In Agro-Environmental Sustainability; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 1–18. [Google Scholar]
- Semrau, J.D. Bioremediation via methanotrophy: Overview of recent findings and suggestions for future research. Front. Microbiol. 2011, 2, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- US EPA O Basic Information about Landfill Gas. Available online: https://www.epa.gov/lmop/basic-information-about-landfill-gas (accessed on 25 June 2020).
- Cébron, A.; Bodrossy, L.; Chen, Y.; Singer, A.C.; Thompson, I.P.; Prosser, J.I.; Murrell, J.C. Identity of active methanotrophs in landfill cover soil as revealed by DNA-stable isotope probing. FEMS Microbiol. Ecol. 2007, 62, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Li, Y.; Lidstrom, M. The oxidative TCA cycle operates during methanotrophic growth of the Type I methanotroph Methylomicrobium buryatense 5GB1. Metab. Eng. 2017, 42, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Puri, A.W.; Owen, S.; Chu, F.; Chavkin, T.; Beck, D.A.C.; Kalyuzhnaya, M.G.; Lidstrom, M.E. Genetic tools for the industrially promising methanotroph Methylomicrobium buryatense. Appl. Environ. Microbiol. 2015, 81, 1775–1781. [Google Scholar] [CrossRef] [Green Version]
- Karlsen, O.A.; Berven, F.S.; Jensen, H.B.; Fjellbirkeland, A. Methods in Methane Metabolism, Part B: Methanotrophy; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Tikhonova, E.N.; Kravchenko, I.K. Activity and diversity of aerobic methanotrophs in thermal springs of the russian far east. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Biotechnology in Agro-Environmental Sustainability; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–30. [Google Scholar]
- Wang, F.; Zhang, Y.; Chen, Y.; He, Y.; Qi, J.; Hinrichs, K.; Zhang, X.; Xiao, X.; Boon, N. Methanotrophic archaea possessing diverging methane-oxidizing and electron-Transporting pathways. ISME J. 2014, 8, 1069–1078. [Google Scholar] [CrossRef] [Green Version]
- Cassarini, C.; Zhang, Y.; Lens, P.N.L. Pressure Selects Dominant Anaerobic Methanotrophic Phylotype and Sulfate Reducing Bacteria in Coastal Marine Lake Grevelingen Sediment. Front. Environ. Sci. 2019, 6, 162. [Google Scholar] [CrossRef] [Green Version]
- Timmers, P.H.A.; Welte, C.U.; Koehorst, J.J.; Plugge, C.M.; Jetten, M.S.M.; Stams, A.J.M. Reverse Methanogenesis and Respiration in Methanotrophic Archaea. Archaea 2017, 2017. [Google Scholar] [CrossRef]
- Knittel, K.; Lösekann, T.; Boetius, A.; Kort, R.; Amann, R. Diversity and distribution of methanotrophic archaea at cold seeps. Appl. Environ. Microbiol. 2005, 71, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Fradet, D.T.; Tavormina, P.L.; Orphan, V.J. Members of the methanotrophic genus Methylomarinum inhabit inland mud pots. PeerJ 2016, 4, e2116. [Google Scholar] [CrossRef] [Green Version]
- Burdette, M. Production of Biodiesel-Like Components by the Type I Methanotroph Methylomonas Methanica. Master’s Thesis, Clemson University, Clemson, SC, USA, August 2013. [Google Scholar]
- Sundstrom, E.R.; Criddle, C.S. Optimization of methanotrophic growth and production of poly(3-hydroxybutyrate) in a high-throughput microbioreactor system. Appl. Environ. Microbiol. 2015, 81, 4767–4773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bussmann, I.; Rahalkar, M.; Schink, B. Cultivation of methanotrophic bacteria in opposing gradients of methane and oxygen. FEMS Microbiol. Ecol. 2006, 56, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Farhan Ul Haque, M.; Crombie, A.T.; Ensminger, S.A.; Baciu, C.; Murrell, J.C. Facultative methanotrophs are abundant at terrestrial natural gas seeps. Microbiome 2018, 6, 118. [Google Scholar] [CrossRef]
- Kolb, S.; Knief, C.; Stubner, S.; Conrad, R. Quantitative detection of methanotrophs in soil by novel pmoA-targeted real-time PCR Assays. Appl. Environ. Microbiol. 2003, 69, 2423–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skennerton, C.T.; Ward, L.M.; Michel, A.; Metcalfe, K.; Valiente, C.; Mullin, S.; Chan, K.Y.; Gradinaru, V.; Orphan, V.J. Genomic Reconstruction of an Uncultured Hydrothermal Vent Gammaproteobacterial Methanotroph (Family Methylothermaceae) Indicates Multiple Adaptations to Oxygen Limitation. Front. Microbiol. 2015, 6, 1425. [Google Scholar] [CrossRef]
- Merkel, A.Y.; Huber, J.A.; Chernyh, N.A.; Bonch-Osmolovskaya, E.A.; Lebedinsky, A.V. Detection of putatively thermophilic anaerobic methanotrophs in diffuse hydrothermal vent fluids. Appl. Environ. Microbiol. 2013, 79, 915–923. [Google Scholar] [CrossRef] [Green Version]
- Singleton, M.; McCalley, C.K.; Woodcroft, B.J.; Boyd, J.A.; Evans, P.N.; Hodgkins, S.B.; Chanton, J.P.; Frolking, S.; Crill, P.M.; Saleska, S.R. Methanotrophy across a natural permafrost thaw environment. ISME J. 2018, 12, 2544–2558. [Google Scholar] [CrossRef] [Green Version]
- Crevecoeur, S.; Vincent, W.F.; Comte, J.; Matveev, A.; Lovejoy, C. Diversity and potential activity of methanotrophs in high methane-emitting permafrost thaw ponds. PLoS ONE 2017, 12, e0188223. [Google Scholar] [CrossRef] [Green Version]
- General Culturing Tips|Methanotroph Commons. Available online: http://www.methanotroph.org/wiki/culturing-tips/ (accessed on 12 July 2020).
- Akberdin, I.R.; Thompson, M.; Hamilton, R.; Desai, N.; Alexander, D.; Henard, C.A.; Guarnieri, M.T.; Kalyuzhnaya, M.G. Methane utilization in Methylomicrobium alcaliphilum 20ZR: A systems approach. Sci. Rep. 2018, 8, 2512. [Google Scholar] [CrossRef] [Green Version]
- Tays, C.; Guarnieri, M.T.; Sauvageau, D.; Stein, L.Y. Combined Effects of Carbon and Nitrogen Source to Optimize Growth of Proteobacterial Methanotrophs. Front. Microbiol. 2018, 9, 2239. [Google Scholar] [CrossRef]
- Kuźniar, A.; Furtak, K.; Włodarczyk, K.; Stępniewska, Z.; Wolińska, A. Methanotrophic bacterial biomass as potential mineral feed ingredients for animals. Int. J. Environ. Res. Public Health 2019, 16, 2674. [Google Scholar] [CrossRef] [Green Version]
- Roell, G.W.; Zha, J.; Carr, R.R.; Koffas, M.A.; Fong, S.S.; Tang, Y.J. Engineering microbial consortia by division of labor. Microb. Cell Fact. 2019, 18, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawed, K.; Yazdani, S.S.; Koffas, M.A. Advances in the development and application of microbial consortia for metabolic engineering. Metab. Eng. Commun. 2019, 9, e00095. [Google Scholar] [CrossRef] [PubMed]
- Yalkowsky, S.H.; He, Y.; Jain, P. Handbook of Aqueous Solubility Data; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Chemical Composition of Natural Gas - Union Gas. Available online: https://www.uniongas.com/about-us/about-natural-gas/chemical-composition-of-natural-gas (accessed on 12 July 2020).
- Fei, Q.; Guarnieri, M.T.; Tao, L.; Laurens, L.M.L.; Dowe, N.; Pienkos, P.T. Bioconversion of natural gas to liquid fuel: Opportunities and challenges. Biotechnol. Adv. 2014, 32, 596–614. [Google Scholar] [CrossRef] [Green Version]
- Just The Facts - Average Price of Natural Gas - The Public Policy Institute. Available online: http://www.ppinys.org/reports/jtf2004/naturalgas.htm (accessed on 12 July 2020).
- What is Renewable Natural Gas?|SoCalGas. Available online: https://www.socalgas.com/smart-energy/renewable-gas/what-is-renewable-natural-gas (accessed on 13 July 2020).
- Parker, N.; Williams, R.; Dominguez-Faus, R.; Scheitrum, D. Renewable natural gas in California: An assessment of the technical and economic potential. Energy Policy 2017, 111, 235–245. [Google Scholar] [CrossRef]
- Renewable Natural Gas Rates. Available online: https://www.fortisbc.com/services/sustainable-energy-options/renewable-natural-gas/renewable-natural-gas-rates (accessed on 13 July 2020).
- Whittenbury, R.; Phillips, K.C.; Wilkinson, J.F. Enrichment, isolation and some properties of methane-utilizing bacteria. J. Gen. Microbiol. 1970, 61, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, O.P.; Saravanan, N.; Cirés, S.; Alvarez-Roa, C.; Razaghi, A.; Chidambarampadmavathy, K.; Velu, C.; Subashchandrabose, G.; Heimann, K. Culture scale-up and immobilisation of a mixed methanotrophic consortium for methane remediation in pilot-scale bio-filters. Environ. Technol. (U. K.) 2017, 38, 474–482. [Google Scholar] [CrossRef]
- Hoefman, S.; van der Ha, D.; de Vos, P.; Boon, N.; Heylen, K. Miniaturized extinction culturing is the preferred strategy for rapid isolation of fast-growing methane-oxidizing bacteria. Microb. Biotechnol. 2012, 5, 368–378. [Google Scholar] [CrossRef] [Green Version]
- Prabandono, K.; Amin, S. Biofuel Production from Microalgae. In Handbook of Marine Microalgae, Biotechnology Advances; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 145–158. [Google Scholar]
- Lee, E.Y. METHANOTROPHS: Microbiology fundamentals and biotechnological applications. Springer Nat. 2019, 1, 163–203. [Google Scholar]
- Jahnke, L.L.; Summons, R.E.; Hope, J.M.; Des Marais, D.J. Carbon isotopic fractionation in lipids from methanotrophic bacteria II: The effects of physiology and environmental parameters on the biosynthesis and isotopic signatures of biomarkers. Geochim. Cosmochim. Acta 1999, 63, 79–93. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.; Shim, S.; Ryu, Y.; Yang, J.; Lim, S.; Lee, C. Lipid Extraction from Tetraselmis sp. Microalgae for Biodiesel Production Using Hexane-based Solvent Mixtures. Biotechnol. Bioprocess. Eng. 2018, 23, 16–22. [Google Scholar] [CrossRef]
- Supercritical Fluid Extraction (SFE) Systems: Waters. Available online: https://www.waters.com/waters/en_US/Supercritical-Fluid-Extraction-(SFE)-Systems/nav.htm?cid=10146521&locale=en_US (accessed on 13 July 2020).
- Chester, T.L.; Pinkston, J.D.; Raynie, D.E. Supercritical Fluid Chromatography/Mass Spectrometry. In Practical Supercritical Fluid Chromatography and Extraction; American Chemical Society: Washington, DC, USA, 2018; pp. 161–178. [Google Scholar]
- Breil, C.; Vian, M.A.; Zemb, T.; Kunz, W.; Chemat, F. “Bligh and Dyer” and Folch methods for solid–liquid–liquid extraction of lipids from microorganisms. Comprehension of solvatation mechanisms and towards substitution with alternative solvents. Int. J. Mol. Sci. 2017, 18, 708. [Google Scholar] [CrossRef] [Green Version]
- Gorgich, M.; Mata, T.M.; Martins, A.A.; Branco-Vieira, M.; Caetano, N.S. Comparison of different lipid extraction procedures applied to three microalgal species. In Energy Reports; Elsevier Ltd.: Amsterdam, The Netherlands, 2020; pp. 477–482. [Google Scholar]
- Kwangdinata, R.; Raya, I.; Zakir, M. Production of Biodiesel from Lipid of Porphyridium cruentum through Ultrasonic Method. IRSN Renew. Energy 2014, 2014, 107278. [Google Scholar] [CrossRef] [Green Version]
- Transesterification to Biodiesel. Available online: https://www.etipbioenergy.eu/value-chains/conversion-technologies/conventional-technologies/transesterification-to-biodiesel (accessed on 13 July 2020).
- Raut, G.; Kamat, S.; Ravikumar, A. Trends in production and fuel properties of biodiesel from heterotrophic microbes. In Advances in Biological Science Research: A Practical Approach; Elsevier: Amsterdam, The Netherlands, 2019; pp. 247–273. [Google Scholar]
- Levett, I.; Birkett, G.; Davies, N.; Bell, A.; Langford, A.; Laycock, B.; Lant, P.; Pratt, S. Techno-economic assessment of poly-3-hydroxybutyrate (PHB) production from methane - The case for thermophilic bioprocessing. J. Environ. Chem. Eng. 2016, 4, 3724–3733. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.L.; Alvarez-Cohen, L. Two-stage methanotrophic bioreactor for the treatment of chlorinated organic wastewater. Water Res. 1997, 31, 2026–2036. [Google Scholar] [CrossRef]
- Steinberg, L.M.; Kronyak, R.E.; House, C.H. Coupling of anaerobic waste treatment to produce protein- and lipid-rich bacterial biomass. Life Sci. Space Res. 2017, 15, 32–42. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Lee, E.Y. Biological conversion of methane to putrescine using genome-scale model-guided metabolic engineering of a methanotrophic bacterium Methylomicrobium alcaliphilum 20Z. Biotechnol. Biofuels 2019, 12, 147. [Google Scholar] [CrossRef]
- Broadwith, P. Magnificent Molecules (Putrescine and Cadaverine), 2012. Royal Society of Chemistry. Available online: http://www.rsc.org/images/TM0512-putrescine-cadaverine_tcm18-221656.pdf (accessed on 30 July 2020).
- Strong, P.J.; Xie, S.; Clarke, W.P. Methane as a resource: Can the methanotrophs add value? Environ. Sci. Technol. 2015, 49, 4001–4018. [Google Scholar] [CrossRef]
- Chemical Safety Facts About Methanol. Available online: https://www.chemicalsafetyfacts.org/methanol/ (accessed on 13 July 2020).
- Knief, C. Diversity and habitat preferences of cultivated and uncultivated aerobic methanotrophic bacteria evaluated based on pmoA as molecular marker. Front. Microbiol. 2015, 6, 1346. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Xu, X.; Dai, M.; Yuan, X.; Guo, R. Hydrogen and methane production from vinasse using two-stage anaerobic digestion. Process. Saf. Environ. Prot. 2017, 107, 81–86. [Google Scholar] [CrossRef]
- Calysta. Producing Protein and Plastics from Methane, a Sustainable Platform for Biotechnology, 2017. Bio.org. Available online: https://www.bio.org/sites/default/files/0830AM-Lori%20Giver.pdf (accessed on 30 July 2020).
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zappi, A.; Fortela, D.L.; Holmes, W.E. An Assessment of Methanotrophs Producing Industrial-Grade Lipids for Biofuels and Other Commercial Chemicals. Energies 2020, 13, 3887. https://doi.org/10.3390/en13153887
Zappi A, Fortela DL, Holmes WE. An Assessment of Methanotrophs Producing Industrial-Grade Lipids for Biofuels and Other Commercial Chemicals. Energies. 2020; 13(15):3887. https://doi.org/10.3390/en13153887
Chicago/Turabian StyleZappi, Alex, Dhan Lord Fortela, and William E. Holmes. 2020. "An Assessment of Methanotrophs Producing Industrial-Grade Lipids for Biofuels and Other Commercial Chemicals" Energies 13, no. 15: 3887. https://doi.org/10.3390/en13153887
APA StyleZappi, A., Fortela, D. L., & Holmes, W. E. (2020). An Assessment of Methanotrophs Producing Industrial-Grade Lipids for Biofuels and Other Commercial Chemicals. Energies, 13(15), 3887. https://doi.org/10.3390/en13153887