Biomass Production of Three Biofuel Energy Plants’ Use of a New Carbon Resource by Carbonic Anhydrase in Simulated Karst Soils: Mechanism and Capacity


 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Substrate
2.2. Experimental Design and Growth Conditions
2.3. Physiological Characteristics
2.4. Biomass Production
2.5. CA Activity
2.6. Stable Carbon–Isotope Ratio Analysis
2.7. Proportion of Bicarbonate Use
2.8. Statistical Analyses
3. Results and Discussion
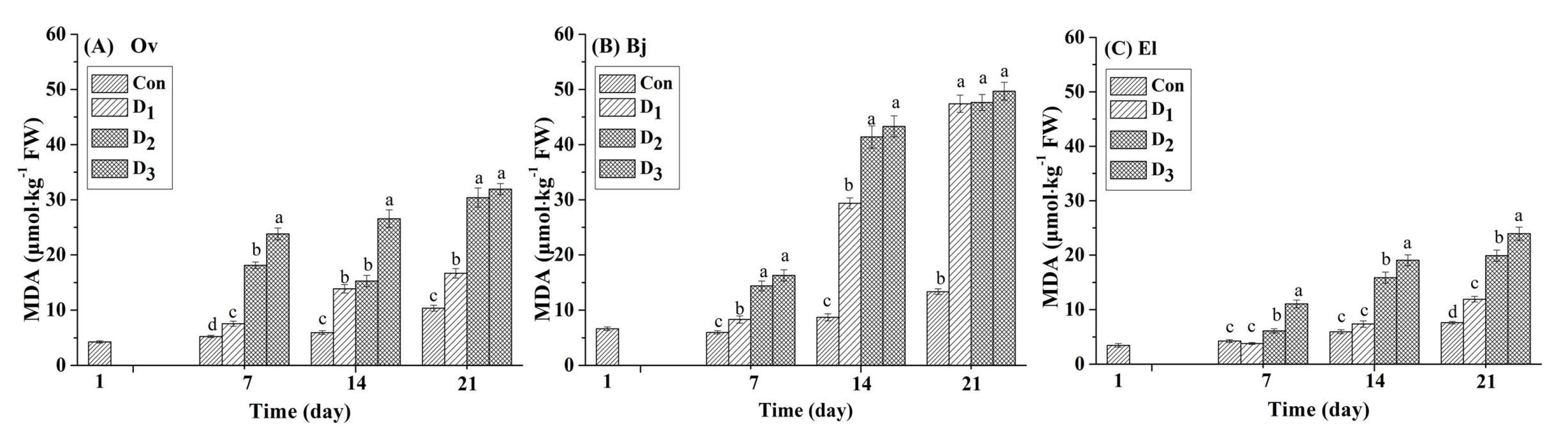
3.1. MDA Content of Plants Leaves Under Drought Stress
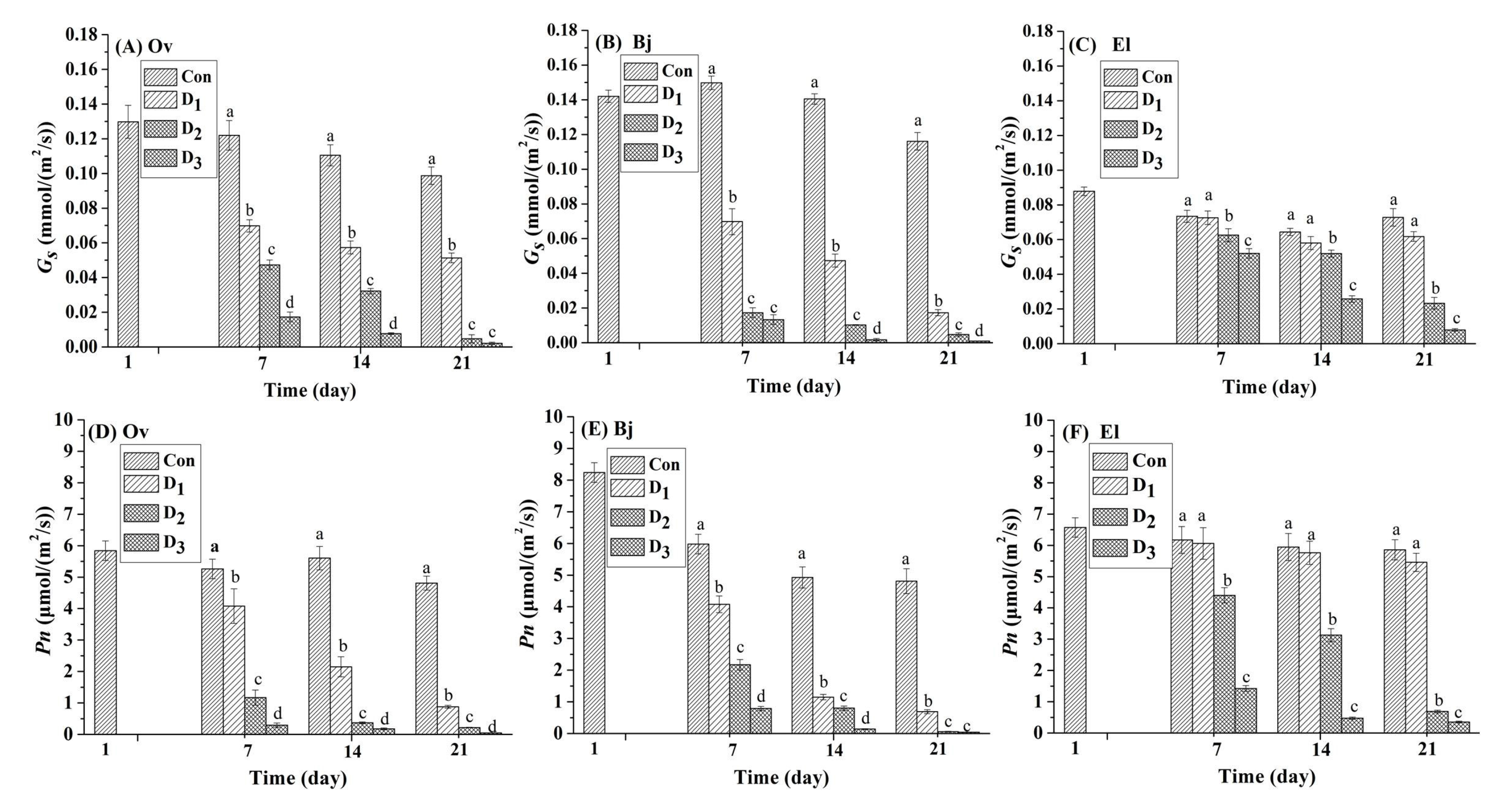
3.2. Photosynthesis of Plant Leaves under Drought Stress
3.3. Biomass Production
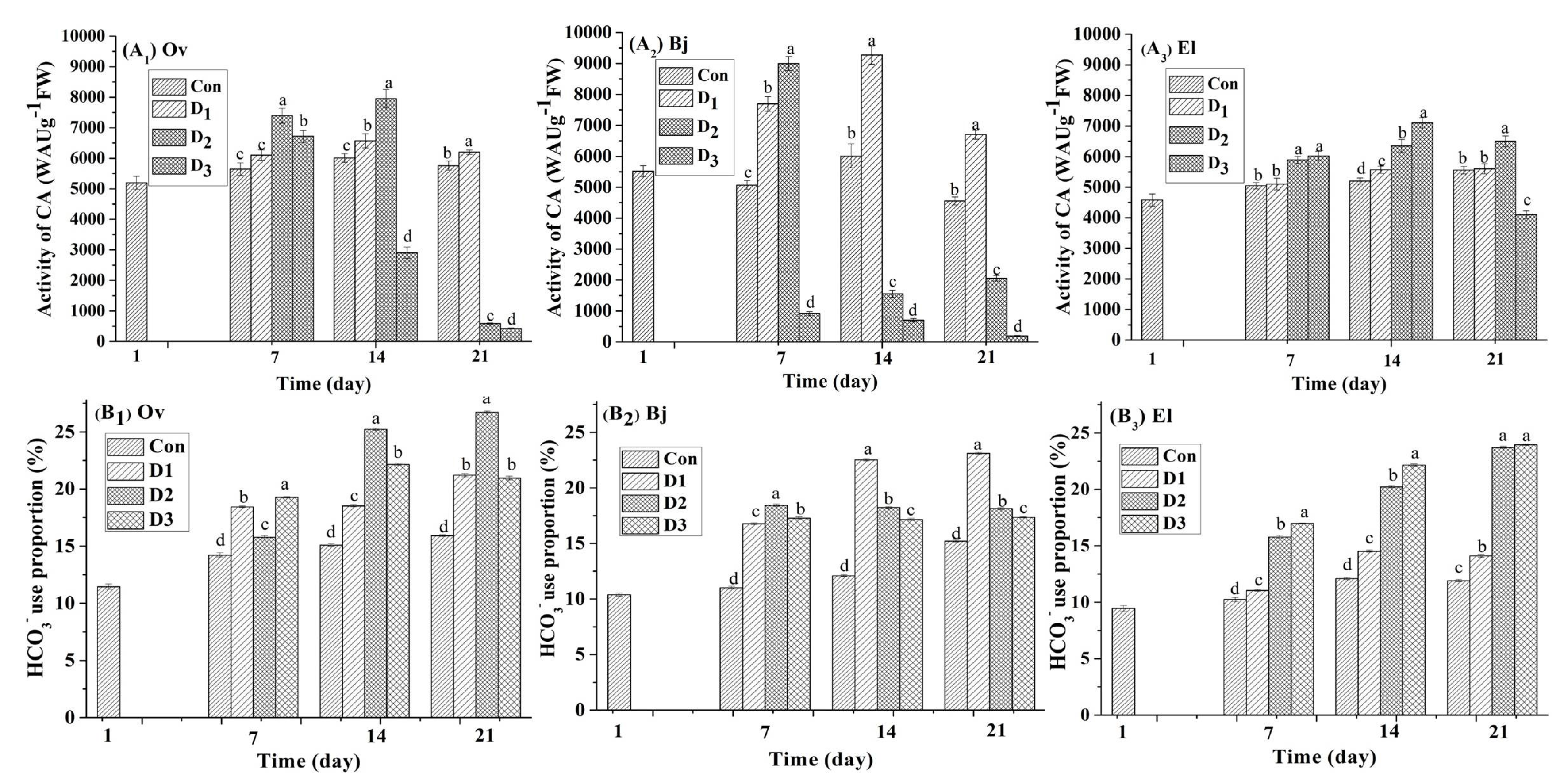
3.4. CA Activity and Proportion of Bicarbonate Use
4. Conclusions
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FAC (wt %) | Palmitic (C16:0) | Palmitoleic (C16:1) | Stearic (C18:0) | Oleic (C18:1) | Linoleic (C18:2) | Linolenic (C18:3) | Gadoleic (C20:1) | Erucic (C22:1) | |
|---|---|---|---|---|---|---|---|---|---|
| Plants/Treatment | |||||||||
| Ov | Con | 13.27 | 0.63 | 9.82 | 22.47 | 42.58 | 4.26 | 3.79 | 3.79 |
| D1 | 13.22 | 0.65 | 9.79 | 22.41 | 41.91 | 4.27 | 4.62 | 4.62 | |
| D2 | 14.37 | 0.23 | 11.53 | 25.23 | 40.43 | 2.12 | 3.94 | 3.95 | |
| D3 | 15.11 | 0.37 | 13.77 | 27.94 | 33.94 | 0.99 | 4.51 | 4.51 | |
| Bj | Con | 5.95 | 2.02 | 3.01 | 29.17 | 20.41 | 11.91 | 9.11 | 9.11 |
| D1 | 6.70 | 2.49 | 2.92 | 28.89 | 21.13 | 10.54 | 10.01 | 10.01 | |
| D2 | 6.92 | 2.91 | 2.61 | 31.77 | 15.19 | 7.43 | 11.97 | 11.97 | |
| D3 | - a | - a | - a | - a | - a | - a | - a | - a | |
| El | Con | 5.11 | 1.79 | 2.17 | 79.58 | 3.83 | 2.37 | 1.73 | 1.73 |
| D1 | 5.09 | 1.78 | 2.02 | 80.51 | 2.59 | 2.49 | 0.94 | 1.74 | |
| D2 | 7.41 | 1.79 | 3.51 | 79.13 | 3.41 | 1.31 | 1.45 | 1.45 | |
| D3 | 9.66 | 0.63 | 5.19 | 82.79 | 1.02 | 0.88 | 1.63 | 1.63 |
| FAC (wt %) | Saturation (Cn:0) | Mono-Unsaturation (Cn:1) | Poly-Unsaturation (Cn:2,3) | Degree of Unsaturation a | CN | HHV MJ/kg | KV mm2/s; 40 °C | |
|---|---|---|---|---|---|---|---|---|
| Plants/Treatment | ||||||||
| Ov | Con | 23.09 | 28.56 | 46.84 | 122.24 | 49.53 | 39.19 | 4.08 |
| D1 | 23.01 | 29.27 | 46.18 | 121.63 | 50.75 | 39.18 | 4.07 | |
| D2 | 25.9 | 30.08 | 42.55 | 115.18 | 53.75 | 39.21 | 4.13 | |
| D3 | 28.88 | 34.75 | 33.53 | 101.81 | 54.08 | 39.26 | 4.18 | |
| Bj | Con | 8.96 | 56.77 | 32.32 | 121.41 | 49.24 | 39.23 | 4.34 |
| D1 | 9.62 | 57.11 | 31.67 | 120.45 | 50.81 | 39.36 | 4.36 | |
| D2 | 9.53 | 66.44 | 22.62 | 107.68 | 50.12 | 39.52 | 4.56 | |
| D3 | - b | - b | - b | - b | - b | - b | - b | |
| El | Con | 7.28 | 83.89 | 6.2 | 96.29 | 59.17 | 38.82 | 4.28 |
| D1 | 7.11 | 84.14 | 5.08 | 94.3 | 60.20 | 38.41 | 4.22 | |
| D2 | 10.92 | 82.76 | 4.72 | 92.2 | 58.68 | 39.24 | 4.38 | |
| D3 | 14.85 | 85.74 | 1.9 | 89.54 | 60.06 | 40.87 | 4.71 |
| Indexes | δ13C Values (‰) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Plants/Label | Day 1 | Day 7 | Day 14 | Day 21 | |||||
| −27.28 | −18.81 | −27.28 | −18.81 | −27.28 | −18.81 | −27.28 | −18.81 | ||
| Ov | Con | −30.74 ± 1.47 | −29.77 ± 1.28 | 30.53 ± 1.32 | 29.32 ± 1.32 | 30.88 ± 1.05 | 29.60 ± 1.28 | 30.99 ± 1.03 | −29.642 ± 1.08 |
| D1 | 30.44 ± 1.25 | 28.87 ± 1.11 | 30.87 ± 1.52 | 29.31 ± 1.06 | 30.95 ± 0.89 | −29.15 ± 1.05 | |||
| D2 | 29.68 ± 1.13 | 28.34 ± 1.28 | 30.60 ± 1.40 | 28.46 ± 0.67 | 30.80 ± 1.12 | −28.54 ± 1.00 | |||
| D3 | 29.55 ± 1.45 | 27.92 ± 1.05 | 30.22 ± 1.27 | 28.34 ± 0.89 | 30.45 ± 1.23 | −28.68 ± 0.78 | |||
| Bj | Con | −33.57 ± 1.61 | −32.69 ± 1.55 | 33.37 ± 1.60 | 32.44 ± 1.30 | 33.54 ± 1.00 | 32.52 ± 0.89 | 33.87 ± 1.54 | −32.58 ± 0.98 |
| D1 | 32.68 ± 1.29 | 31.26 ± 1.43 | 32.78 ± 1.03 | 32.87 ± 1.15 | 32.90 ± 1.28 | −30.94 ± 1.04 | |||
| D2 | 33.19 ± 1.56 | 31.63 ± 1.67 | 33.35 ± 0.99 | 31.81 ± 0.77 | 33.56 ± 1.11 | −32.03 ± 0.99 | |||
| D3 | 32.93 ± 1.34 | 31.47 ± 0.99 | 33.18 ± 1.23 | 31.73 ± 1.35 | 33.43 ± 1.52 | −31.96 ± 1.30 | |||
| El | Con | −31.38 ± 1.52 | −30.58 ± 1.40 | 31.58 ± 1.07 | 30.72 ± 1.10 | 31.76 ± 1.37 | 30.51 ± 1.14 | 31.77 ± 1.34 | −30.76 ± 0.96 |
| D1 | 31.41 ± 1.65 | 30.47 ± 1.23 | 31.51 ± 1.42 | 30.28 ± 1.08 | 31.67 ± 1.22 | −30.47 ± 1.27 | |||
| D2 | 31.22 ± 1.66 | 29.88 ± 1.30 | 31.30 ± 1.50 | 29.59 ± 0.89 | 31.44 ± 1.45 | −29.45 ± 0.94 | |||
| D3 | 31.01 ± 1.29 | 29.57 ± 1.14 | 29.59 ± 1.03 | −29.23 ± 0.96 | −31.21 ± 1.03 | −29.18 ± 0.97 | |||
Acknowledgments
Author Contributions
Conflicts of Interest
Nomenclature
| CA | Carbonic anhydrase |
| Ov | Orychophragmus violaceus L. |
| Bj | Brassica juncea L. |
| El | Euphorbia lathyris L. |
| CN | Cetane number |
| HHV | Higher heating value |
| KV | Kinematic viscosity |
| FAC | Fatty acid composition |
References
- Gelfand, I.; Sahajpal, R.; Zhang, X.; Izaurralde, R.C.; Gross, K.L.; Robertson, G.P. Sustainable bioenergy production from marginal lands in the US Midwest. Nature 2013, 493, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.; Hensgen, F.; Wachendorf, M. Using Grass Cuttings from Sports Fields for Anaerobic Digestion and Combustion. Energies 2017, 10, 388–399. [Google Scholar] [CrossRef]
- Cotana, F.; Cavalaglio, G.; Coccia, V.; Petrozzi, A. Energy Opportunities from Lignocellulosic Biomass for a Biorefinery Case Study. Energies 2016, 9, 748–758. [Google Scholar] [CrossRef]
- Roy, P.; Dutta, A.; Deen, B. An approach to identify the suitable plant location for Miscanthus-based ethanol industry: A case study in Ontario, Canada. Energies 2015, 8, 9266–9281. [Google Scholar] [CrossRef]
- Stehr, M.O. Sustainability assessment of a self-consumption wood-energy chain on small scale for heat generation in central Italy. Energies 2015, 8, 5182–5197. [Google Scholar]
- Zhao, L.L.; Zhang, X.L.; Xu, J.; Ou, X.; Chang, S.; Wu, M. Techno-economic analysis of bioethanol production from lignocellulosic biomass in China: Dilute-acid pretreatment and enzymatic hydrolysis of corn stover. Energies 2015, 8, 4096–4117. [Google Scholar] [CrossRef]
- Tang, Y.; Xie, J.S.; Geng, S. Marginal land-based biomass energy production in China. J. Integr. Plant Biol. 2010, 52, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, D.; Jiang, D.; Liu, L.; Huang, Y. Assessment of bioenergy potential on marginal land in China. Renew. Sustain. Energy Rev. 2011, 15, 1050–1056. [Google Scholar] [CrossRef]
- Liu, C.Q. Biogeochemical Processes and the Surface Material Cycle: The Cycles of Biogenic Elements of Soil–Vegetation System in the Southwestern Karst China; Science Press: Beijing, China, 2009. [Google Scholar]
- Yan, J.; Li, J.; Ye, Q.; Li, K. Concentrations and exports of solutes from surface runoff in Houzhai Karst Basin, southwest China. Chem. Geol. 2012, 304, 1–9. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Xing, D.K. Effect of bicarbonate treatment on photosynthetic assimilation of inorganic carbon in two plant species of Moraceae. Photosynthetica 2012, 50, 587–594. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Bicarbonate supplementation enhanced biofuel production potential as well as nutritional stress mitigation in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 193, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Boisson-Dernier, A.; Israelsson-Nordstrom, M.; Bohmer, M.; Xue, S.; Ries, A.; Godoski, J.; Kuhn, J.M.; Schroeder, J.I. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells. Nat. Cell Biol. 2010, 12, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.G.; Qiang, L.; Schröder, H.; Hönig, N.; Yuan, D.; Grebenjuk, V.; Mussino, F.; Giovine, M.; Wang, X. Carbonic anhydrase: A key regulatory and detoxifying enzyme for Karst plants. Planta 2014, 239, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Perez-Martin, A.; Michelazzo, C.; Torres-Ruiz, J.M.; Flexas, J.; Fernandez, J.E.; Sebastiani, L.; Diaz-Espejo, A. Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: Correlation with gene expression of carbonic anhydrase and aquaporins. J. Exp. Bot. 2014, 65, 3143–3156. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, R.; Kannan, K.; Deshkar, A.; Yadav, R.; Chakrabarti, T. Enhanced algal CO2 sequestration through calcite deposition by Chlorella sp. and Spirulina platensis in a mini-raceway pond. Bioresour. Technol. 2010, 101, 2616–2622. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.-F.; Qiu, M.; Yang, J.-C. Enhanced tolerance to drought in transgenic rice plants overexpressing C4 photosynthesis enzymes. Crop J. 2013, 1, 105–114. [Google Scholar] [CrossRef]
- Jiang, C.; Tholen, D.; Xu, J.; Xin, C.; Zhang, H.; Zhu, X.; Zhao, Y. Increased expression of mitochondria-localized carbonic anhydrase activity resulted in an increased biomass accumulation in Arabidopsis thaliana. J. Plant Biol. 2014, 57, 366–374. [Google Scholar] [CrossRef]
- Xie, T.; Wu, Y. The role of microalgae and their carbonic anhydrase on the biological dissolution of limestone. Environ. Earth Sci. 2014, 71, 5231–5239. [Google Scholar] [CrossRef]
- Swarnalatha, G.V.; Hegde, N.S.; Chauhan, V.S.; Sarada, R. The effect of carbon dioxide rich environment on carbonic anhydrase activity, growth and metabolite production in indigenous freshwater microalgae. Algal Res. 2015, 9, 151–159. [Google Scholar] [CrossRef]
- Wu, Y.; Xing, D.; Liu, Y. The characteristics of bicarbonate used by plants. Earth Environ. 2011, 39, 273–277. [Google Scholar]
- Motomura, H.; Ueno, O.; Kagawa, A.; Yukawa, T. Carbon isotope ratios and the variation in the diurnal pattern of malate accumulation in aerial roots of CAM species of Phalaenopsis (Orchidaceae). Photosynthetica 2008, 46, 531–536. [Google Scholar] [CrossRef]
- Zuo, Y.; Ren, L.; Zhang, F.; Jiang, R.F. Bicarbonate concentration as affected by soil water content controls iron nutrition of peanut plants in a calcareous soil. Plant Physiol. Biochem. 2007, 45, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Mir, B.A.; Mir, S.A.; Khazir, J.; Tonfack, L.B.; Cowan, D.A.; Vyas, D.; Koul, S. Cold stress affects antioxidative response and accumulation of medicinally important withanolides in Withania somnifera (L.) Dunal. Ind. Crops Prod. 2015, 74, 1008–1016. [Google Scholar] [CrossRef]
- Wang, R.; Hanna, M.A.; Zhou, W.-W.; Bhadury, P.S.; Chen, Q.; Song, B.-A.; Yang, S. Production and selected fuel properties of biodiesel from promising non-edible oils: Euphorbia lathyris L., Sapium sebiferum L. and Jatropha curcas L. Bioresour. Technol. 2011, 102, 1194–1199. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Verduzco, L.F.; Rodríguez-Rodríguez, J.E.; del Jaramillo-Jacob, A.R. Predicting cetane number, kinematic viscosity, density and higher heating value of biodiesel from its fatty acid methyl ester composition. Fuel 2012, 91, 102–111. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernández, C.M.; Casas, A.; Rodríguez, L.; Pérez, Á. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Pinzi, S.; Garcia, I.; Lopez-Gimenez, F.; Luque de Castro, M.; Dorado, G.; Dorado, M. The ideal vegetable oil-based biodiesel composition: A review of social, economical and technical implications. Energy Fuels 2009, 23, 2325–2341. [Google Scholar] [CrossRef]
| Indexes | Dry Biomass Productivity (g) | Oil Content (wt %) | Acid Value (mg KOH/g) | ||||
|---|---|---|---|---|---|---|---|
| Plants/Treatment | Seed | Root/Stem/Leaf | Total Biomass | Average Total Biomass Under Stress | |||
| Ov | Con | 8.93 | 113.28 | 122.21 | 111.38 | 30.17 ± 2.21a | 4.02 ± 0.09d |
| D1 | 7.27 | 111.08 | 118.35 | 29.17 ± 1.34a | 4.61 ± 0.10c | ||
| D2 | 6.68 | 103.54 | 110.22 | 22.35 ± 1.41b | 5.02 ± 0.07b | ||
| D3 | 4.42 | 101.15 | 105.57 | 16.79 ± 0.73c | 7.41 ± 0.11a | ||
| Bj | Con | 11.31 | 135.12 | 146.43 | 127.55 | 31.42 ± 1.93a | 5.07 ± 0.14d |
| D1 | 6.22 | 131.51 | 137.73 | 26.27 ± 1.58b | 5.74 ± 0.12c | ||
| D2 | 3.17 | 125.47 | 128.64 | 13.46 ± 0.83c | 7.07 ± 0.07b | ||
| D3 | 2.34 | 113.93 | 116.27 | 10.17 ± 1.83c | 9.17 ± 0.19a | ||
| El | Con | 10.90 | 76.88 | 86.29 | 81.65 | 29.83 ± 1.33a | 14.11 ± 0.13c |
| D1 | 9.59 | 73.14 | 84.18 | 29.13 ± 1.51a | 13.90 ± 0.22c | ||
| D2 | 9.43 | 74.75 | 82.73 | 28.83 ± 1.19a | 15.29 ± 0.21b | ||
| D3 | 6.67 | 71.38 | 78.05 | 22.83 ± 2.07c | 16.11 ± 0.08a | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Wu, Y.; Xing, D.; Hang, H.; Xie, X.; Yang, X.; Zhang, K.; Rao, S. Biomass Production of Three Biofuel Energy Plants’ Use of a New Carbon Resource by Carbonic Anhydrase in Simulated Karst Soils: Mechanism and Capacity. Energies 2017, 10, 1370. https://doi.org/10.3390/en10091370
Wang R, Wu Y, Xing D, Hang H, Xie X, Yang X, Zhang K, Rao S. Biomass Production of Three Biofuel Energy Plants’ Use of a New Carbon Resource by Carbonic Anhydrase in Simulated Karst Soils: Mechanism and Capacity. Energies. 2017; 10(9):1370. https://doi.org/10.3390/en10091370
Chicago/Turabian StyleWang, Rui, Yanyou Wu, Deke Xing, Hongtao Hang, Xiaolin Xie, Xiuqun Yang, Kaiyan Zhang, and Sen Rao. 2017. "Biomass Production of Three Biofuel Energy Plants’ Use of a New Carbon Resource by Carbonic Anhydrase in Simulated Karst Soils: Mechanism and Capacity" Energies 10, no. 9: 1370. https://doi.org/10.3390/en10091370
APA StyleWang, R., Wu, Y., Xing, D., Hang, H., Xie, X., Yang, X., Zhang, K., & Rao, S. (2017). Biomass Production of Three Biofuel Energy Plants’ Use of a New Carbon Resource by Carbonic Anhydrase in Simulated Karst Soils: Mechanism and Capacity. Energies, 10(9), 1370. https://doi.org/10.3390/en10091370