Abstract
Arabinoxylan (AX) has been associated with alleviating intestinal barrier damage, and different structures of AX give rise to different effects on the intestinal barrier. This study investigated the main structural characteristics of AX, whose functional properties are attributed to alleviating intestinal barrier damage, and clarified their underlying mechanisms. An in vitro Caco-2 cell model was established to investigate the intestinal barrier effects of AX with various degrees of substitution (Ds) and molecular weight (Mw), with an added MyD88 inhibitor to verify the signaling pathways. Arabinoxylan treated with endo-1,4-β-xylanase (AXX) with higher Ds and Mw showed stronger physiological activity, which might be correlated with the uronic acid and bound ferulic acid contents in AXX. Moreover, AXX alleviated the intestinal barrier damage by upregulating the transepithelial electrical resistance (TER) and alleviating the decrease of claudin-1 (p < 0.05). AXX regulated the expression of inflammatory factors IL-2, TNF-α, IL-6 and IL-10 (p < 0.05). In addition, AXX reduced the intestinal barrier damage induced via inhibiting the TLRs/MyD88/NF-κB pathway and activating the TLRs/PKC pathway. Thus, AX with higher Ds and Mw might be better in alleviating intestinal barrier damage, and MyD88 might be the key point of AXX to identify these signaling pathways.
1. Introduction
The integrity of epithelial cell and tight junction (TJ) proteins is often referred to as the “intestinal barrier” [1,2]. The intestinal epithelial barrier plays a key role in preventing the transfer of pathogenic microorganisms and toxic substances from the intestinal lumen to the systemic circulation [3]. The loss of barrier integrity can lead to the invasion of harmful substances in the intestinal lumen, which can cause different diseases such as inflammatory bowel disease and extraintestinal autoimmune diseases [4,5]. TJ connectivity plays an important role in maintaining the structural and functional integrity of the intestinal barrier by regulating paracellular transport. In recent studies, increased intestinal permeability due to decreased TJ-related protein (such as zonula occludens family proteins, occludin and claudins) expression and epithelial barrier function was observed [6].
Toll-like receptors (TLRs) are a class of transmembrane proteins that trigger cell signal transduction. Toll-like receptor 2 (TLR2) is a potential therapeutic target for gastrointestinal diseases, in which TJ-related intestinal epithelial barrier disruption might be the main feature [7,8]. Toll-like receptor 4 (TLR4) is the best characterization protein of pathogen recognition receptors and plays a homeostasis role in maintaining TJ protein expression and affecting intestinal permeability [9]. Myeloid differentiation factor 88 (MyD88) is the core of transduction of extracellular stimulation via TLRs. After specific recognition, intracellular signal transduction mainly depends on MyD88-dependent and MyD88-independent pathways. Nuclear factor-κB (NF-κB) is an MyD88-dependent node of TLR’s downstream inflammatory signaling pathway, and the activation of NF-κB and synthesis of inflammatory cytokines not only intensify the inflammatory response but also compromise intestinal integrity [10]. Protein kinase C (PKC) is a family of serine- and threonine-specific protein kinases that mediates numerous cellular processes in a tissue-specific manner. Studies have shown that the activation of PKC enhances the integrity of TJ, and the PKC pathway might enhance the intestinal TJ barrier [11].
Arabinoxylan (AX) is a complex polymer that is composed of a backbone of β-d-xylopyranose residues connected by β-(1,4)-glycosidic bonds, which is substituted by arabinose residues at the C(O)-2 and/or C(O)-3 position [12]. In addition, arabinose can be attached with some phenolic acids. For example, ferulic acid (FA) can be covalently linked to the C5 position of the α-L-arabinofuranosidase residue via an ester bond [13]. Many in vivo studies have shown that AX has a protective effect on the intestinal barrier [14,15,16]; however, its specific mechanisms in vitro remain unclear. Studies have shown that the biological activity of AX is highly dependent on the chemical structure of AX [17]. In vitro, Mendis, et al. [18] found that increased arabinose substitution in AX might be better at reducing inflammation in colon cancer cells. AX with low-branched hydroxycinnamate and higher DS exhibit stronger antioxidant activity [19]. Our previous study showed that triticale bran AX with higher molecular weight had higher antioxidant levels [12]. These studies showed that the Mw or degree of arabinose substitution is closely related to the physiological function of AX. Previous studies tended to focus on the effect of a single factor (Mw or Ds) on the physiological function of AX. However, little information is available regarding the combined effect of these two factors on physiological function, and further verification is needed. Therefore, in the present study, four arabinoxylan hydrolysates (AXHs) with different degrees of substitution and molecular weight were prepared for cell culture to investigate the relationship between different unique structures of AX and the effects of alleviating Caco-2 cell permeability. Furthermore, the potential pathways of action were explored.
2. Materials and Methods
2.1. Materials and Reagents
The bran was acquired from Xinxiang Agricultural Development Company Limited (Xinxinag China). Neutral protease (activity: 100 μ/mg protein) was acquired from Shanghai Yuanye Biotechnology Company Limited (Shanghai, China). α-amylase from Aspergillus oryzae and endo-1,4-β-xylanase from Penicillium were acquired from Shanghai Ryon Biological Technology Company Limited (Shanghai, China). α-L-arabinofuranosidase B21 from Bacteroides ovatus was acquired from Megazyme International Ireland (Wicklow, Ireland). Solvents and reagents were of analytical grade throughout the experiment.
2.2. Preparation of Arabinoxylan Hydrolysates
AX was dispersed in citric acid solutions with the content of 0.1 M and 0.15 M [20,21]. After a two hour 95 °C water bath, NaOH was added. The mixture was centrifuged, precipitated with ethanol, redissolved and lyophilized to yield the hydrolysate AXC1 and AXC. AXC was added with 0.15 M HCl to obtain hydrolysate AXH2. α-L-arabinofuranosidase was added in AX solution and oscillated in a 40 °C water bath for 24 h. After centrifugation, samples were freeze-dried to obtain hydrolysate AXAE. Endo-1,4-β-xylanase was added in the AX suspension. After centrifugation, collected and added anhydrous ethanol. After centrifugation for 15 min, the precipitation was collected and redissolved. The supernatant was extracted after repeated centrifugation for 15 min and freeze-dried to obtain hydrolysate AXX.
2.3. Structural Characterisations of Arabinoxylan Hydrolysates
2.3.1. Molecular Weight Measurement
Samples were dissolved and the sample solution was filtrated [17]. A Refractive index detector (Shimadzu RID-20, Shimadzu, Kyoto, Japan) and a gel filtration column (TSK GMPWXL, 7.8 mm × 300 mm, TOSOH, Yamaguchi, Japan) were used for analysis. A quantity of 0.1 N NaNO3 and 0.06% NaN3 aqueous solution as mobile phase and flow rate of 0.6 mL/min. Standard dextrans were used to obtain the standard curve.
2.3.2. Monosaccharide Composition Measurement
Determination with the method illustrated by Yuan, et al. [22]. A Dionex UltiMate 3000 HPLC system (Thermo Fisher Scientific, Waltham, MA, USA) was used to analyze 1-Phenyl-3-methyl-5-pyrazolone (PMP) derivatives. A quantity of 20 μL of PMP derivatives were injected into the HPLC system.
2.3.3. 1H NMR Analysis
The analysis was conducted in accordance with a previous article [23]. Here, after exchange with D2O, the samples were finally dissolved in pure D2O. A 400.00 MHz spectrometer (JNM-ECZ400S, JEOL, Tokyo, Japan) was used to obtain the 1H spectra of samples. The 1H chemical shifts (ppm) were referenced to a D2O signal at 4.790 ppm at 25 °C.
2.3.4. FT-IR Analysis
The analysis was conducted in accordance with a previous article [22]. Fourier transform infrared spectra of freeze-dried samples were recorded at room temperature with an FTIR spectrometer (Thermo Fisher Scientific, Waltham, MA, USA).
2.3.5. Ferulic Acid Determination
FA content was measured by spectrophotometry [24]. The sample was dissolved in 1 mL of ultrapure water. A quantity of 900 μL of glycine-NaOH buffer (pH 10, 0.04 M) was added to 100 μL of the sample solution. The absorbance values were at 345 and 375 nm.
2.4. Establishment of the Cell Model
2.4.1. Cell Culture
Caco-2 cell culture was performed based on Fang, et al. [25]. Human colon cells (Caco-2) were selected as the research object. The Caco-2 cell line (ATCC, Manassas, VA, USA), derived from human colon cancer cells, in DMEM containing 10% fetal bovine serum, 50 U/L penicillin-streptomycin and 1% non-essential amino acid at 37 °C, 5% CO2 high-sugar medium, was cultured in a 25 cm2 cell culture flask, replacing the culture medium every 1–2 days. After 5–7 days, the cells had grown to confluence and passaged at 1:2 or 1:3.
2.4.2. Treatment of Cells with Arabinoxylan Hydrolysates
The treatment was performed based on Wu, et al. [26]. The cells were starved for 12 h before the experiment on serum-free media, and the treatment groups were added with AXHs (400 μg/mL) and different concentrations of lipopolysaccharide (LPS, μg/mL) after 1 h preincubation. The control group (without LPS and AXH), negative control group (with LPS) and treatment groups (with LPS and AXH) were set. The resistance values of each group were detected after 24 h of culture, and the experiment was repeated three times.
2.4.3. Treatment of Cells with MyD88 Inhibitor
The treatment was performed based on Song, et al. [27]. The cells were starved for 12 h before the experiment (adding serum-free medium), and all groups were added with MyD88 inhibitor. After 1 h, the AXHs with the best screening effect was added (μg/mL). After 2 h, LPS (μg/mL) was added. The control group (no LPS and no AXH), negative control group (only LPS), treatment group I (LPS and AXH, no MyD88 inhibitor) and treatment group II (LPS, the best effect AXH and MyD88 inhibitor) were set. The resistance values of each group were detected after 24 h of culture, and the experiment was repeated three times.
2.5. Determination of Epithelial Monolayer Resistance
Determination was conducted with the method illustrated by Wu, et al. [28]. The cells were starved for 12 h before the experiment (adding serum-free medium), pre-incubated for 1 h and then added with LPS (μg/mL). Solvent was used as the control group, and the resistance value of each group was detected after 24 h of culture. The short and long arms of the cell resistor electrode were inserted into the upper and lower chambers of the transwell chamber, respectively. Three different points of the chambers were measured, and the average value of the measured values was taken.
2.6. Assessment of Protein Expression by Western Blot
Assessment by Western blot was performed according to Wu, et al. [29]. The Caco-2 cell culture medium was aspirated and rinsed with PBS twice. The cells were digested by trypsin, and the pipetted cells were transferred into a 1.5 mL EP tube. A total of 150 μL of RIPA lysate containing PMSF was added to the centrifuge tube and centrifuged at 4 °C, and the supernatant after centrifugation was prepared with 10% polyacrylamide gel to prepare SDS-PAGE gel. About 50 μg of protein sample was prepared and loaded. Firstly, electrophoresis was performed at a constant voltage of 80 V and then 100 V. The protein in the gel was transferred to the PVDF membrane by transfer membrane and blocked with TBST buffer containing 5% skimmed milk powder. Then, the primary antibody was incubated overnight at 4 °C. The secondary antibody was incubated at room temperature for 1 h. ImageJ image processing software was used to analyze the Mw and net optical density value of the target band. The average of five experimental results was regarded as the relative protein content. β-actin was the internal reference gene.
2.7. Quantification of Gene Expression Using Real-Time PCR
The cells were collected and washed with PBS buffer three times. Then, 1 mL of Trizol reagent was added to each well to extract the total RNA of the cells. RNA concentration and purity were detected and reversed transcription was synthesized into cDNA. The target gene sequence was acquired from the NCBI biological information website, and Primer 5 primer design software was used to construct the gene primers. A 10 μL SYBR Green reaction system was used for RT-PCR detection, β-actin and GADPH were used as internal reference genes for calculation, and the target gene level was expressed in terms of relative expression. The calculation method of the results was reported in the study of Pfaffl, et al. [30].
2.8. Enzyme-Linked Immunosorbent Assay
The culture medium was discarded. The cells were collected and washed three times with PBS buffer. An ELISA kit was used to determine the cytokines IL-2, IL-6, IL-10 and TNF-α in the Caco-2 cell culture medium content. The determination method was carried out in accordance with the ELISA kit’s instructions.
2.9. Statistical Analysis
All data were entered into Excel 2019. OriginPro 9.1 was used for mapping. IBM SPSS Statistics 26 mathematical software was used for statistical analysis of experimental data, and analysis of variance (ANOVA) was used to verify the statistical differences between groups. The difference was statistically significant with p < 0.05, and the results were expressed as mean ± standard deviation.
3. Results
3.1. Structural Characterisations of Arabinoxylan Hydrolysates
3.1.1. Molecular Weight and Degree of Substitution of Arabinoxylan Hydrolysates
The Mw and Ds of AXHs are shown in Table 1. The monosaccharide composition of AXH was mainly composed of xylose and arabinose, following with glucose and galactose. Different AXHs had a different ratio of arabinose to xylose and AXX and AXAE contained more arabinose. Furthermore, the Mw of AXH acquired by endo-1,4-β-xylanase treatment was higher than that acquired by α-L-arabinofuranosidase treatment. The Mw is mainly distributed between 2.67 × 103 Da to 6.43 × 105 Da. Furthermore, in the acid-treated AXC1 and AXH2, the Mw of AXH2 was lower than that of AXC1.

Table 1.
Molecular weight and monosaccharide composition of AXHs.
3.1.2. 1H NMR Analysis of Arabinoxylan Hydrolysates
The structural change of AXH was verified by 1H NMR. The NMR spectra of all samples showed 3–6 main peaks in the chemical shift range of 5.15–5.40 ppm. The signal peak of the arabinose residue was connected to the C(O)-3 position of the xylose residue on the main chain at a chemical shift of 5.38 ppm. The signal peak around 5.27 and/or 5.21 ppm corresponding to xylose residues was monosubstituted by arabinose residues at the C(O)-2 and/or C(O)-3 position. The chemical shift of xylose residues was monosubstituted at the C(O)-3 and/or C(O)-2 position at 4.50 and/or 4.60 ppm. The chemical shift of xylose residues was disubstituted at 4.62 ppm. As shown in Figure 1, the contents of xylose residues in AXH2 and AXAE were higher than those in AXX and AXC1.
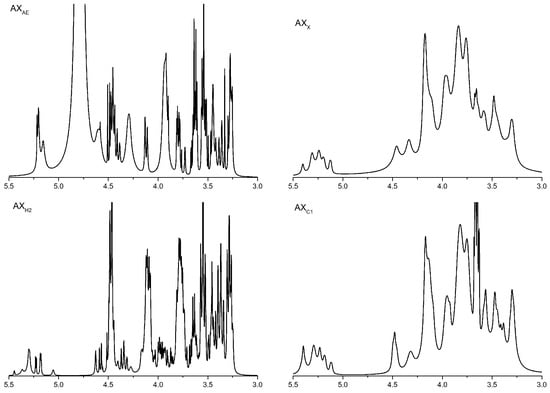
Figure 1.
1H NMR analysis of AXH. AXAE, triticale bran arabinoxylan was treated with α-L-arabinofuranosidase; AXC1, triticale bran arabinoxylan was treated with 0.1 M citric acid; AXH2, triticale bran arabinoxylan was treated with 0.15 M HCL; AXX, triticale bran arabinoxylan was treated with endo-1,4-β-xylanase.
3.1.3. FT-IR Spectrum Analysis of Arabinoxylan Hydrolysates
As shown in Figure 2, a broad and strong peak appeared near 3410 cm−1. The range of 3000–2800 cm−1 were produced by C-H vibration. The band between 1700–1600 cm−1 was the C=O asymmetric stretching vibration in COO-. The absorption band between 1400–1300 cm−1 was the C=O symmetric stretching vibration in COO-. Frequency band signals between 1200 cm−1 and 800 cm−1 were associated with a specific polysaccharide structure composed of pyranose ring vibration, which overlapped with C-C and C-OH stretching vibration and glycosidic bond (C-O-C) vibration. AXAE had a relatively strong absorption peak at 1074.50 cm−1. These were the typical structural features of AX.
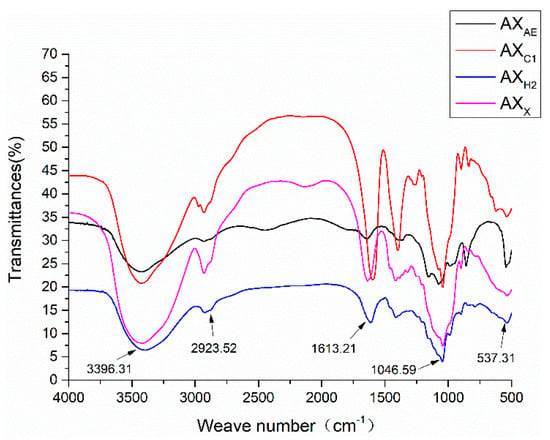
Figure 2.
FT-IR spectra of AXH. AXAE, triticale bran arabinoxylan was treated with α-L-arabinofuranosidase; AXC1, triticale bran arabinoxylan was treated with 0.1 M citric acid; AXH2, triticale bran arabinoxylan was treated with 0.15 M HCL; AXX, triticale bran arabinoxylan was treated with endo-1,4-β-xylanase.
3.1.4. Ferulic Acid Content Analysis
FA was a major bound phenolic acid linked to AX mainly with ester bounds, which affected its physical and chemical properties. Free FA content was higher than that of bound FA (Figure 3). Bound FA content in AXX was higher than that acquired by acid and α-L-arabinofuranosidase treatment (p < 0.05).
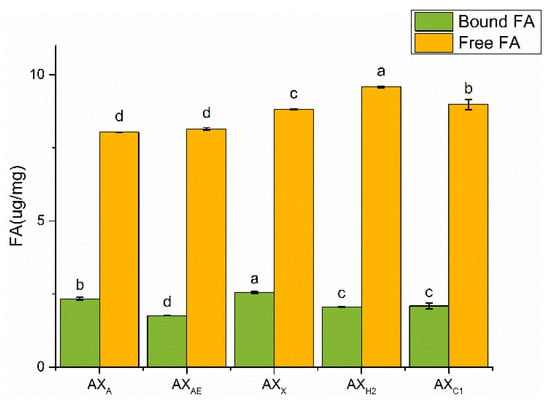
Figure 3.
The content of FA in AXH. AXAE, triticale bran arabinoxylan was treated with α-L-arabinofuranosidase; AXC1, triticale bran arabinoxylan was treated with 0.1 M citric acid; AXH2, triticale bran arabinoxylan was treated with 0.15 M HCL; AXX, triticale bran arabinoxylan was treated with endo-1,4-β-xylanase. Values represented in this figure consist of mean ± standard deviation (n = 3). Values with different superscript letter are significantly different (p < 0.05).
3.2. Regulating Effects of AXH on the Intestinal Barrier in the Caco-2 Model
3.2.1. Transepithelial Electrical Resistance Measurement
About 1 μg/mL of LPS was used as the optimal LPS emergency concentration for our study. The alleviating effect of each sample on LPS-induced intestinal epithelial barrier integrity destruction was monitored by transepithelial electrical resistivity (Figure 4). Our research showed that after treatment with the LPS group, the resistivity of Caco-2 intestinal epithelial cells decreased by 9%. The TER value of the LPS + AXX group was higher than the LPS + AXC1 group (407.12 ± 34.08 Ω/cm2), and there was no significant difference among other groups.
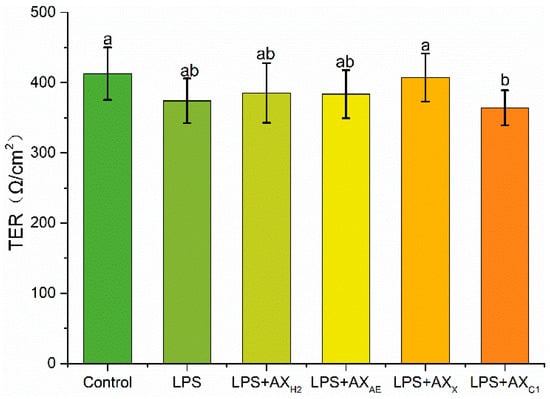
Figure 4.
Transepithelial electrical resistance after treated with LPS. Control, without LPS or arabinoxylan hydrolysates; AXH2, triticale bran arabinoxylan was treated with 0.15 M HCL; AXAE, triticale bran arabinoxylan was treated with α-L-arabinofuranosidase; AXC1, triticale bran arabinoxylan was treated with 0.1 M citric acid; AXX, triticale bran arabinoxylan was treated with endo-1,4-β-xylanase. Values represented in this figure consist of mean ± standard deviation (n = 3). Values with different superscript letter are significantly different (p < 0.05).
3.2.2. Tight Junction Protein Expression in Caco-2 Cells
Claudin-1, occludin and zonula occludens (ZO)-1 were down-regulated under LPS conditions (p < 0.05), and this effect was changed by the different structures of AXHs (Figure 5A). Compared with LPS, AXX increased the expression of claudin-1 (p < 0.05), and there was no significant expression of occludin and ZO-1 in the AXX group (p > 0.05). As shown in Figure 5B, the expression of claudin-1 was down-regulated in the LPS + AXX + MyD88 inhibitor group (p < 0.05), whereas there was no significant change in occludin and ZO-1 (p > 0.05).
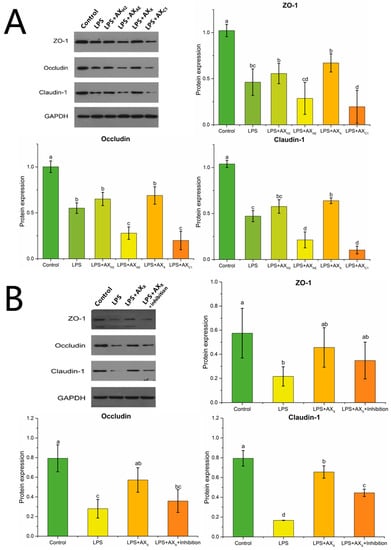
Figure 5.
Tight junction protein expression in Caco-2 cells treated with (A) AXHs and (B) MyD88 inhibitor. Control, without LPS or arabinoxylan hydrolysates; AXH2, triticale bran arabinoxylan was treated with 0.15 M HCl; AXAE, triticale bran arabinoxylan was treated with α-L-arabinofuranosidase; AXC1, triticale bran arabinoxylan was treated with 0.1 M citric acid; AXX, triticale bran arabinoxylan was treated with endo-1,4-β-xylanase; Inhibitor, MyD88 inhibitor. Values represented in this figure consist of mean ± standard deviation (n = 3). Values with different superscript letter are significantly different (p < 0.05).
3.2.3. Signaling Pathway in Caco-2 Cells
As shown in Figure 6A,B, compared with the LPS group, the AXX and AXH2 groups up-regulated the expression of TLR2 and TLR4 (p < 0.05). After MyD88 inhibitor was added, the expression of TLR2 and TLR4 was down-regulated compared with the LPS group (p < 0.05). The LPS + AXX group up-regulated the expression of MyD88 compared with the LPS group, whereas this change decreased in the MyD88 inhibitor group (p < 0.05) (Figure 6C). Figure 6D,E shows that LPS-induced PKC and NF-κB expression in Caco-2 cells was upregulated and inhibited, respectively, compared with that in the AXX group (p < 0.05). In the LPS + AXX + MyD88 inhibitor group, NF-κB expression was up-regulated (p < 0.05), whereas PKC expression was not significantly changed compared with that in the AXX group (p > 0.05) (Figure 6D,E).
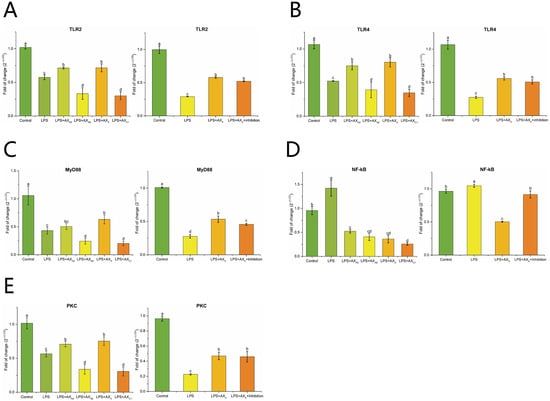
Figure 6.
The effect of AXHs and MyD88 inhibitor on the expression of TLR2 (A), TLR4 (B), MyD88 (C), NF-κB (D), PKC (E) in LPS-induced Caco-2 cells. Control, without LPS or arabinoxylan hydrolysates; AXH2, triticale bran arabinoxylan was treated with 0.15 M HCl; AXAE, triticale bran arabinoxylan was treated with α-L-arabinofuranosidase; AXC1, triticale bran arabinoxylan was treated with 0.1 M citric acid; AXX, triticale bran arabinoxylan was treated with endo-1,4-β-xylanase; Inhibitor, MyD88 inhibitor. Values represented in this figure consist of mean ± standard deviation (n = 3). Values with different superscript letter are significantly different (p < 0.05).
3.2.4. Inflammatory Cytokines
As shown in Figure 7A–D, the LPS + AXX group down-regulated TNF-α, IL-6 and IL-10 expression and up-regulated IL-2 expression compared with the LPS group (p < 0.05). In addition, compared with the LPS + AXX group, the addition of MyD88 inhibitor up-regulated TNF-α, IL-6 and IL-10 expression and down-regulated IL-2 expression (p < 0.05) (Figure 7A–D).
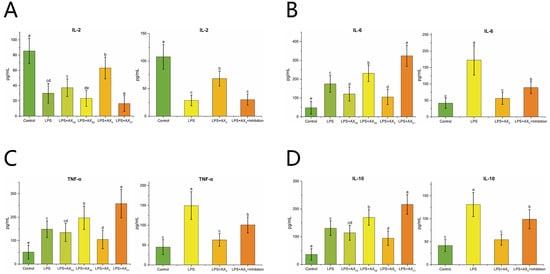
Figure 7.
The effect of AXHs and MyD88 inhibitor on the expression of IL-2 (A), IL-6 (B), TNF-α (C), IL-10 (D) in LPS-induced Caco-2 cells Control, without LPS or arabinoxylan hydrolysates; AXAE, triticale bran arabinoxylan was treated with α-L-arabinofuranosidase; AXX, triticale bran arabinoxylan was treated with endo-1,4-β-xylanase; AXH2, triticale bran arabinoxylan was treated with 0.15 M HCl; AXC1, triticale bran arabinoxylan was treated with 0.1 M citric acid; Inhibitor, MyD88 inhibitor. Values represented in this figure consist of mean ± standard deviation (n = 3). Values with different superscript letter are significantly different (p < 0.05).
4. Discussion
Our previous studies marked the effect of different dietary fibers on improving the intestinal barrier, and found that wheat bran fiber had the best effect [31]. Further studies found that the intestinal epithelial barrier function improvement exerted by wheat bran was related to its AX content [32]. AX with different structural characteristics exhibited different biological activities, and there was a strong correlation between the molecular structure and physiological functions of AX. In our previous study, AXHs with different structures were prepared by different methods [21]. According to their substitution degree and molecular weight difference, four representative AXHs were identified, including AXX with high Ds and high Mw, AXAE with high Ds and low Mw, AXC1 with low Ds and high Mw and AXH2 with low Ds and low Mw. The arabinose residue of AX performed different structures along the xylose backbone [23]. The 1H NMR results showed that AXHs except AXAE had disubstituted arabinose residues, which indicated the role of α-L-arabinofuranosidase, and the higher A/X ratio of AXAE might be related to the region of monosubstituted arabinose residue and disubstituted xylose residues. In addition, AXHs except AXX had mono- and/or disubstituted xylose residues, indicating that endo-1,4-β-xylanase selectively removes xylose residues, and the higher degree of AXX could be confirmed (Ara/Xyl ratio: 1.17). Although the A/X ratio of AXAE and AXX were similar, the substitution patterns of these polymers were not the same. The A/X ratio of AXC1 was higher than AXH2 due to the former monosubstituted xylose residue. FT-IR spectrum results showed that the high Ds of arabinose at xylose residue C3 was indicated by the low-intensity peaks at 988.18 and 1154.83 cm−1, which proved that AXAE had more branched structures and was consistent with the high Ds of AXAE. A strong absorption peak of AXX at 1650 cm−1 was associated with water in the sample, which might be related to the fact that AXX had a highly mono- and/or disubstituted arabinose. The solubility of AX was reported to be determined by its structure, with a higher A/X ratio promoting its solubility [17]. Moreover, the solubilization of arabinose substituents might be due to the prevention of intermolecular aggregation of unsubstituted xylose residues.
Uronic acid was observed in AXX and AXC1. Studies had shown that uronic acid polysaccharides activated MAPK and NF-κB signaling pathways by inducing the release of inflammatory cytokines. Therefore, AX with uronic acid might have more physiological properties [33,34]. Hromádková, et al. [35] revealed that the aggregation formed through the crosslinking of FA substituents might be responsible for the high Mw of AX, which was verified by the higher proportion of FA binding of AXX in our work. FA played an important role in the physiological functions of AX, especially in the antioxidant capacity of AX, but it contributed less to the effect of AX on the Caco-2 cell barrier. Zhang, et al. [16] suggested that FA bound to arabinose in AX to regulate intestinal barrier damage through gut microbiota. We speculated that AXX was more biologically active in the intestinal epithelial cell due to its unique structure. The molecular mechanisms behind this effect remains to be elucidated.
A decrease in TER was generally considered a reference index for cell damage or death, whereas the TER values might be restored and increased by LPS co-culture with AXX, which might be better for improving the permeability of Caco-2 cells. TJs surrounded and anchored adjacent cells. The depletion of TJ protein expression impaired intestinal barrier permeability and the ability to maintain TER [36]. Claudin-1, occludin and ZO-1 were three important TJ proteins. Claudin-1 interacted with other connexins in neighboring cells and formed fences that regulated the permeability of TJ complexes. Occludin could bind with adjacent cells through the outer part of the cell membrane, which conduced to control the permeability between cells. ZO-1 was mainly located at the boundary of adjacent epithelial cells, which anchored occludin and claudin-1 to the cytoskeleton and co-controlled intestinal barrier permeability [37,38]. In this study, LPS reduced the expression of ZO-1, occludin and claudin-1. Claudin was offset by AXX. Claudin has been reported to largely determine the paracellular ion permeability at TJs [39]. We suggested that AXX maintained the integrity of the intestinal epithelial barrier by promoting the expression of claudin-1.
Pro-inflammatory cytokines (IL-1, IL-2, IL-6, etc.) had been proven to be interfering factors of the intestinal barrier. They lead to the further deterioration of intestinal epithelial function by inhibiting TJ protein expression [40]. In this study, AXX induced the expression of interleukin-2 (IL-2), inhibited the expression of interleukin-6 (IL-6), interleukin-10 (IL-10) and tumor necrosis factor-α (TNF-α) to enhance the intestinal immune function of the host, which was consistent with the study of Mendis, et al. [18]. These results suggested that AXX production played an important role in improving intestinal barrier and permeability by regulating TJ protein expression and inflammatory cytokine secretion.
Next, we tried to reveal the mechanism of AXX in alleviating the intestinal barrier in Caco-2 cells. Our previous study found that the up-regulation of TJ protein gene expression in intestinal epithelial cells by wheat bran dietary fiber was accompanied by the up-regulation of TLR2 expression [31]. Dietary fiber inulin-type fructans could modulate TLR2 to enhance the intestinal barrier and prevented pathogens from entering the host [41]. TLR4-mediated intestinal barrier dysfunction was considered to be a key factor in the initiation and enhancement of gastrointestinal injury [42]. In this study, the expression of TLR2 and TLR4 was up-regulated in the AXX and AXH2 groups, which suggested that structurally different AX had similarities in regulating intestinal barrier damage. AXX and AXH2 might induce cytokine production in intestinal epithelial cells in a TLR2- and/or TLR4-dependent manner. The similarity of AXH biologically affected with different Mw and Ds might be due to the similarity of glycosidic bonds and changes in the structure and number of arabinose substitutions in the xylose backbone [43]. In addition, we found that the infrared spectra of AXH2 and AXX were similar. However, according to the data of this study, AXX with high Ds might exert steric hindrance on the formation of intermolecular crosslinks and thus be easily dispersed into the reactive mixture to obtain a better performance.
MyD88 was an important signaling pathway regulating LPS-induced intestinal epithelial TJ permeability [44]. It had been widely reported that polysaccharides exerted their regulatory role by acting on MyD88. Based on these studies, a model with additional MyD88 inhibitor in the LPS + AXX group was proposed to explore and verify the signaling pathways by which AXX alleviated LPS-induced changes in barrier permeability. Our results confirmed that the expression of claudin-1 was affected by the addition of MyD88 inhibitor. The altered expression of inflammatory factors after MyD88 inhibitor suggested that this response might be related to MyD88-dependent TLR signaling. NF-κB activation might be an important pathway of TJ barrier dysfunction induced by pro-inflammatory cytokines. Supplementation with AXX reduced the damage of intestinal mucosa primarily by inhibiting the MyD88-dependent NF-κB pathway and promoting TJ protein expression, which was demonstrated with the addition of MyD88 inhibitor. The phosphorylation of downstream PKC was associated with the activation of TLR2 [45]. Studies had shown that PKC activation enhanced the integrity of TJs, and the PKC pathway might enhance the intestinal TJ barrier [40]. PKC expression was not significantly changed in the LPS + AXX + MyD88 inhibitor group, which suggested that the function of PKC in a pathway was not dependent on MyD88. Given that the addition of AXH2 had no effect on MyD88 expression, we hypothesized that AXX alleviated intestinal barrier damage by stimulating the TLRs/PKC and inhibiting the TLRs/MyD88/NF-κB signaling pathways.
5. Conclusions
The results of this study demonstrated that the function of AXH to alleviate Caco-2 cell permeability was related to its fine structure and physicochemical properties. AXX with higher Mw, Ds and bound FA levels was more beneficial to the improvement of barrier function. Furthermore, AXX promoted epithelial barrier integrity by influencing TER, claudin-1 and inflammatory cytokines via the TLRs/MyD88/NF-κB and TLRs/PKC signaling pathways, which was verified by additional MyD88 inhibitor. Whether TLR2 and TLR4 are involved in the two signaling pathways remains unclear. As far as the current results are concerned, further research is required to obtain AX with detailed structure and characteristics, which will lay a solid foundation on the precise regulation of AX.
Author Contributions
J.L.: investigation, data curation, formal analysis, writing—original draft; Q.J.: software, formal analysis, writing—review and editing; Y.L. (Ying Liu): conceptualization, data curation; D.C.: project administration; Z.F.: project administration; Y.L. (Yuntao Liu): supervision; S.L.: supervision; B.H.: investigation; C.W.: investigation; H.C.: conceptualization, methodology, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the State Key Program of the National Natural Science Foundation of China (No. 31730091).
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ji, J.; Qu, H.; Shu, D. Crosstalk between bioactive peptide and intestinal barrier in gut homeostasis. Curr. Protein Pept. Sci. 2015, 16, 604–612. [Google Scholar] [CrossRef]
- Thadi, A.; Patwa, V.; Joshi, A.; Foss, J.A.; Eddy, E.P.; Palejwala, V.A.; Shailubhai, K. Plecanatide and dolcanatide, guanylate cyclase-C agonists, attenuate paracellular permeability and enhance normal localization of tight junctional proteins to maintain intestinal barrier function. Gastroenterology 2017, 152, S506. [Google Scholar] [CrossRef]
- Wang, B.; Wu, Z.; Ji, Y.; Sun, K.; Dai, Z.; Wu, G. L-glutamine enhances tight junction integrity by activating CaMK kinase 2-AMP-activated protein kinase signaling in intestinal porcine epithelial cells. J. Nutr. 2016, 146, 501–508. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- König, J.; Wells, J.; Cani, P.D.; Ródenas, C.L.G.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.J. Human intestinal barrier function in health and disease. Clin. Transl. Gastroenterol. 2016, 7, e196. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight junction in the intestinal epithelium: Its association with diseases and regulation by phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podlsky, D.K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef]
- Kaminsky, L.; Sadi, R.A.; Ma, T. Lactobacillus acidophilus causes enhancement of the intestinal tight junction barrier by a toll-like receptor-2-dependent increase in occludin. J. Allergy Clin. Immunol. 2022, 149, AB99. [Google Scholar] [CrossRef]
- Bruning, E.E.; Coller, J.K.; Wardill, H.R.; Bowen, J.M. Site-specific contribution of Toll-like receptor 4 to intestinal homeostasis and inflammatory disease. J. Cell. Physiol. 2021, 236, 877–888. [Google Scholar] [CrossRef]
- Dong, N.; Xue, C.; Zhang, L.; Zhang, T.; Wang, C.; Bi, C.; Shan, A. Oleanolic acid enhances tight junctions and ameliorates inflammation in Salmonella typhimurium-induced diarrhea in mice via the TLR4/NF-kappaB and MAPK pathway. Food Funct. 2020, 11, 1122–1132. [Google Scholar] [CrossRef]
- Li, C.; Ai, G.; Wang, Y.; Lu, Q.; Luo, C.; Tan, L.; Lin, G.; Liu, Y.; Li, Y.; Zeng, H.; et al. Oxyberberine, a novel gut microbiota-mediated metabolite of berberine, possesses superior anti-colitis effect: Impact on intestinal epithelial barrier, gut microbiota profile and TLR4-MyD88-NF-kappaB pathway. Pharmacol. Res. 2020, 152, 104603. [Google Scholar] [CrossRef]
- Chen, H.; Chen, Z.; Fu, Y.; Liu, J.; Lin, S.; Zhang, Q.; Liu, Y.; Wu, D.; Lin, D.; Han, G.; et al. Structure, antioxidant, and hypoglycemic activities of arabinoxylans extracted by multiple methods from triticale. Antioxidants 2019, 8, 584. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Courtin, C.M.; Verbeke, K.; Wiele, T.V.; Verstraete, W.; Delcour, J.A. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2014, 51, 178–194. [Google Scholar] [CrossRef]
- Zhao, Z.; Cheng, W.; Qu, W.; Wang, K. Arabinoxylan rice bran (MGN-3/Biobran) alleviates radiation-induced intestinal barrier dysfunction of mice in a mitochondrion-dependent manner. Biomed. Pharmacother. 2020, 124, 109855. [Google Scholar] [CrossRef]
- Salden, B.N.; Troost, F.J.; Wilms, E.; Truchado, P.; Vargas, R.V.; Pieper, D.H.; Jáuregui, R.; Marzorati, M.; Wiele, T.; Possemiers, S.; et al. Reinforcement of intestinal epithelial barrier by arabinoxylans in overweight and obese subjects: A randomized controlled trial: Arabinoxylans in gut barrier. Clin. Nutr. 2018, 37, 471–480. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, P.; Zhao, J. Ferulic acid mediates prebiotic responses of cereal-derived arabinoxylans on host health. Anim. Nutr. 2022, 9, 31–38. [Google Scholar] [CrossRef]
- Chen, Z.; Li, S.; Fu, Y.; Li, C.; Chen, D.; Chen, H. Arabinoxylan structural characteristics, interaction with gut microbiota and potential health functions. J. Funct. Foods 2019, 54, 536–551. [Google Scholar] [CrossRef]
- Mendis, M.; Leclerc, E.; Simsek, S. Arabinoxylan hydrolyzates as immunomodulators in Caco-2 and HT-29 colon cancer cell lines. Food Funct. 2017, 8, 220–231. [Google Scholar] [CrossRef]
- Bijalwan, V.; Ali, U.; Kesarwani, A.K.; Yadav, K.; Mazumder, K. Hydroxycinnamic acid bound arabinoxylans from millet brans-structural features and antioxidant activity. Int. J. Biol. Macromol. 2016, 88, 296–305. [Google Scholar] [CrossRef]
- Mendis, M.; Simsek, S. Production of structurally diverse wheat arabinoxylan hydrolyzates using combinations of xylanase and arabinofuranosidase. Carbohyd. Polym. 2015, 132, 452–459. [Google Scholar] [CrossRef]
- Chen, H.; Liu, Y.; Yang, T.; Chen, D.; Xiao, Y.; Qin, W.; Wu, D.; Zhang, Q.; Lin, D.; Liu, Y.; et al. Interactive effects of molecular weight and degree of substitution on biological activities of arabinoxylan and its hydrolysates from triticale bran. Int. J. Biol. Macromol. 2021, 166, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Lin, S.; Fu, Y.; Nie, X.; Liu, W.; Su, Y.; Han, Q.; Zhao, L.; Zhang, Q.; Lin, D.; et al. Effects of extraction methods on the physicochemical characteristics and biological activities of polysaccharides from okra (Abelmoschus esculentus). Int. J. Biol. Macromol. 2019, 127, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, L.; Tuomainen, P.; Virkki, L.; Tenkanen, M. Molecular characterization and solution properties of enzymatically tailored arabinoxylans. Int. J. Biol. Macromol. 2011, 49, 963–969. [Google Scholar] [CrossRef]
- Malunga, L.N.; Beta, T. Antioxidant capacity of arabinoxylan oligosaccharide fractions prepared from wheat aleurone using Trichoderma viride or Neocallimastix patriciarum xylanase. Food Chem. 2015, 167, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Liang, F.; Liu, K.; Qaiser, S.; Pan, S.; Xu, X. Structure characteristics for intestinal uptake of flavonoids in Caco-2 cells. Food Res. Int. 2018, 105, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Luo, T.; Li, Y.; Gao, Z.; Zhang, K.; Song, J.; Xiao, J.; Cao, Y. Granny Smith apple procyanidin extract upregulates tight junction protein expression and modulates oxidative stress and inflammation in lipopolysaccharide-induced Caco-2 cells. Food Funct. 2018, 9, 3321–3329. [Google Scholar] [CrossRef]
- Song, J.; Chen, D.; Pan, Y.; Shi, X.; Liu, Q.; Lu, X.; Xu, X.; Chen, G.; Cai, Y. Discovery of a novel MyD88 inhibitor M20 and its protection against sepsis-mediated acute lung injury. Front. Pharmacol. 2021, 12, 775117. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, Y.; Ouyang, Y.; He, Y.; Xiao, J.; Zhang, L.; Feng, N. Effect of catechin on dietary AGEs absorption and cytotoxicity in Caco-2 cells. Food Chem. 2021, 355, 129574. [Google Scholar] [CrossRef]
- Wu, Q.; Li, S.; Li, X.; Sui, Y.; Yang, Y.; Dong, L.; Xie, B.; Sun, Z. Inhibition of advanced glycation endproduct formation by lotus seedpod oligomeric procyanidins through RAGE–MAPK signaling and NF-κB activation in high-fat-diet rats. J. Agric. Food Chem. 2015, 63, 6989–6998. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Lange, I.G.; Daxenberger, A.; Meyer, H.H.D. Tissue-specific expression pattern of estrogen receptors (ER): Quantification of ERα and ERβ mRNA with real-time RT-PCR. APMIS 2001, 109, 345–355. [Google Scholar] [CrossRef]
- Chen, H.; Mao, X.; He, J.; Yu, B.; Huang, Z.; Yu, J.; Zheng, P.; Chen, D. Dietary fibre affects intestinal mucosal barrier function and regulates intestinal bacteria in weaning piglets. Br. J. Nutr. 2013, 110, 1837–1848. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, W.; Degroote, J.; Possemiers, S.; Chen, D.; de Smet, S.; Michiels, J. Arabinoxylan in wheat is more responsible than cellulose for promoting intestinal barrier function in weaned male piglets. J. Nutr. 2015, 145, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhou, D.; Xiao, H.; Fu, X.; Kong, Q.; Zhu, C.; Han, Z.; Mou, H. Marine-derived uronic acid-containing polysaccharides: Structures, sources, production, and nutritional functions. Trends Food Sci. Technol. 2022, 122, 1–12. [Google Scholar] [CrossRef]
- Köwitsch, A.; Zhou, G.; Groth, T. Medical application of glycosaminoglycans: A review. J. Tissue Eng. Regen. Med. 2018, 12, e23–e41. [Google Scholar] [CrossRef] [PubMed]
- Hromádková, Z.; Paulsen, B.S.; Polovka, M.; Kostálová, Z.; Ebringerová, A. Structural features of two heteroxylan polysaccharide fractions from wheat bran with anti-complementary and antioxidant activities. Carbohyd. Polym. 2013, 93, 22–30. [Google Scholar] [CrossRef]
- Wu, R.Y.; Abdullah, M.; Määttänen, P.; Pilar, A.V.C.; Scruten, E.; Henry, K.C.J.; Napper, S.; Brien, C.; Jones, N.L.; Sherman, P.M. Protein kinase C delta signaling is required for dietary prebiotic-induced strengthening of intestinal epithelial barrier function. Sci. Rep. 2017, 7, 40820. [Google Scholar] [CrossRef]
- Itallie, C.M.V.; Anderson, J.M. Architecture of tight junctions and principles of molecular composition. Semin. Cell Dev. Biol. 2014, 36, 157–165. [Google Scholar] [CrossRef]
- Garcia, M.A.; Nelson, W.J.; Chavez, N. Cell-cell junctions organize structural and signaling networks. Csh. Perspect. Biol. 2018, 10, a029181. [Google Scholar] [CrossRef]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef]
- Jo, H.; Hwang, D.; Kim, J.K.; Lim, Y.H. Oxyresveratrol improves tight junction integrity through the PKC and MAPK signaling pathways in Caco-2 cells. Food Chem. Toxicol. 2017, 108, 203–213. [Google Scholar] [CrossRef]
- Vogt, L.M.; Meyer, D.; Pullens, G.; Faas, M.M.; Venema, K.; Ramasamy, U.; Schols, H.A.; Vos, P. Toll-like receptor 2 activation by beta2→1-fructans protects barrier function of T84 human intestinal epithelial cells in a chain length-dependent manner. J. Nutr. 2014, 144, 1002–1008. [Google Scholar] [CrossRef] [PubMed]
- Wardill, H.R.; Gibson, R.J.; Sebille, Y.Z.A.V.; Secombe, K.R.; Coller, J.K.; White, I.A.; Manavis, J.; Hutchinson, M.R.; Staikopoulos, V.; Logan, R.M.; et al. Irinotecan-induced gastrointestinal dysfunction and pain are mediated by common TLR4-dependent mechanisms. Mol. Cancer Ther. 2016, 15, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Paesani, C.; Degano, A.L.; Zalosnik, M.I.; Fabi, J.P.; Perez, G.T. Enzymatic modification of arabinoxylans from soft and hard Argentinian wheat inhibits the viability of HCT-116 cells. Food Res. Int. 2021, 147, 110466. [Google Scholar] [CrossRef] [PubMed]
- Nighot, M.; Sadi, R.A.; Guo, S.; Rawat, M.; Nighot, P.; Watterson, M.D.; Ma, T.Y. Lipopolysaccharide-Induced Increase in Intestinal Epithelial Tight Permeability Is Mediated by Toll-Like Receptor 4/Myeloid Differentiation Primary Response 88 (MyD88) Activation of Myosin Light Chain Kinase Expression. Am. J. Pathol. 2017, 187, 2698–2710. [Google Scholar] [CrossRef]
- Yuki, T.; Yoshida, H.; Akazawa, Y.; Komiya, A.; Sugiyama, Y.; Inoue, S. Activation of TLR2 enhances tight junction barrier in epidermal keratinocytes. J. Immunol. 2011, 187, 3230–3237. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).