

Anthocyanins in Dairy Cow Nutrition: A Review


Abstract
1. Introduction
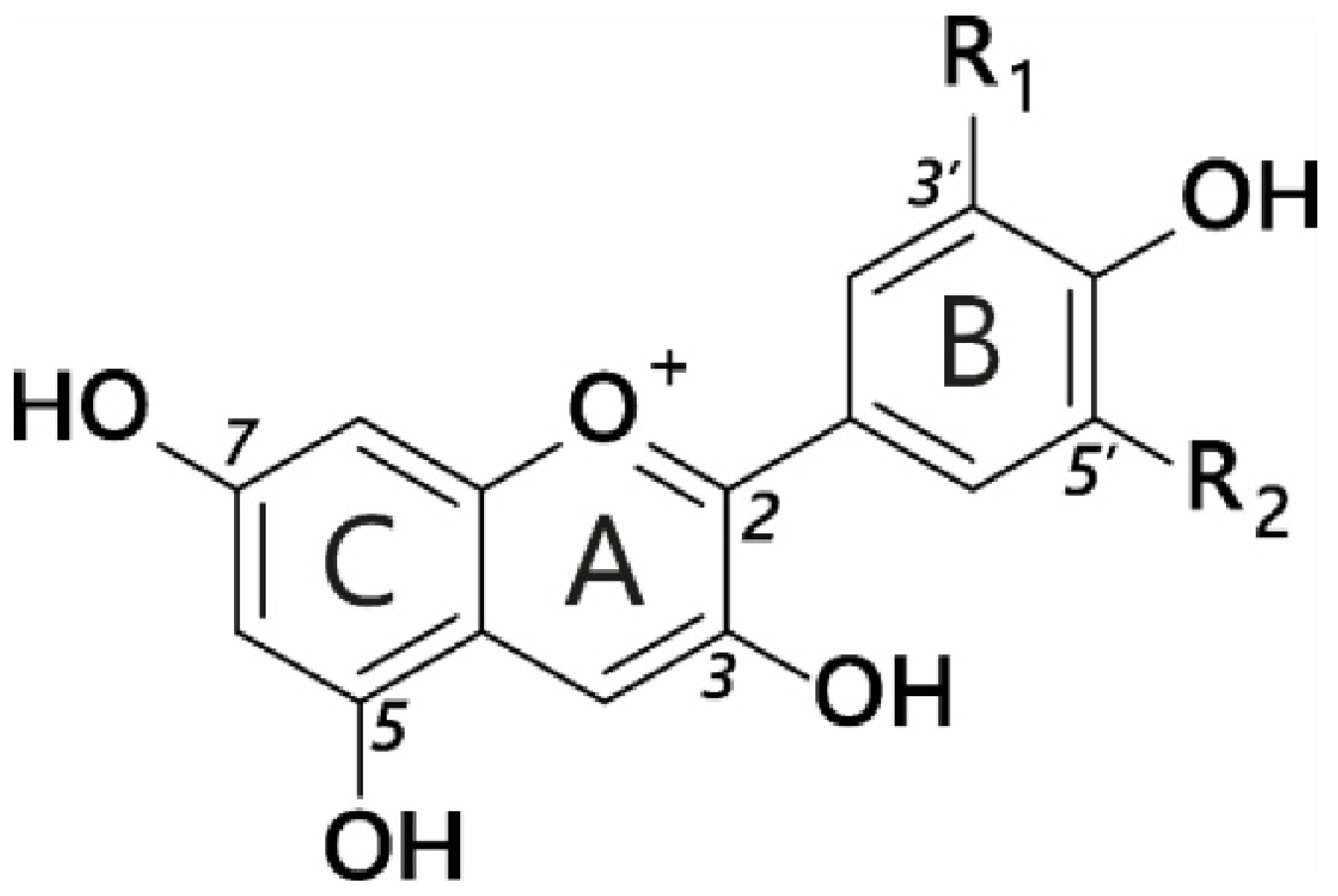
2. Definition of Anthocyanins
3. Possible Absorption and Metabolic Pathways in Dairy Cows
4. Effect of Anthocyanins on Rumen Fermentation and Methane Production
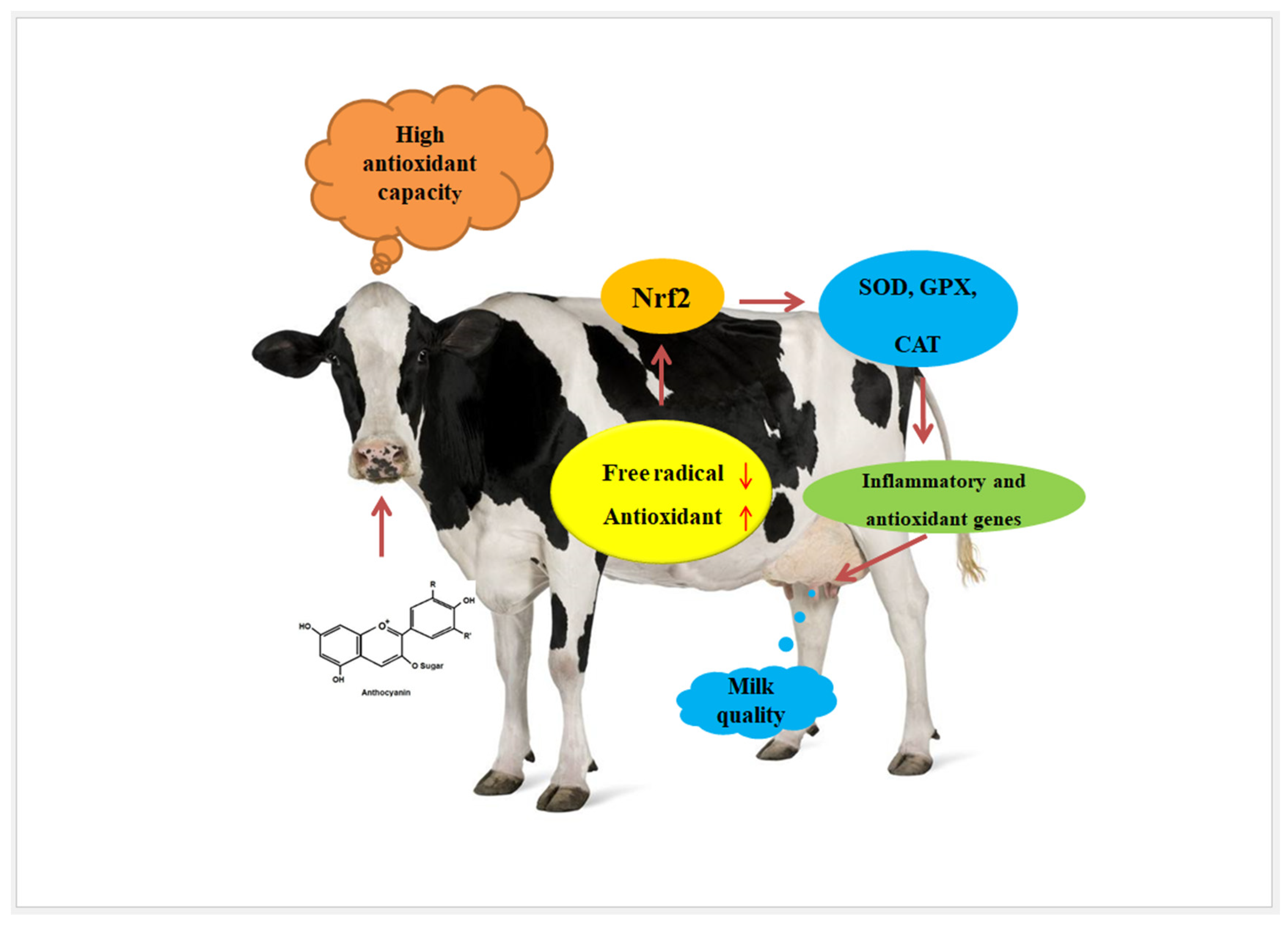
5. Effect of Anthocyanins on Antioxidant Activity
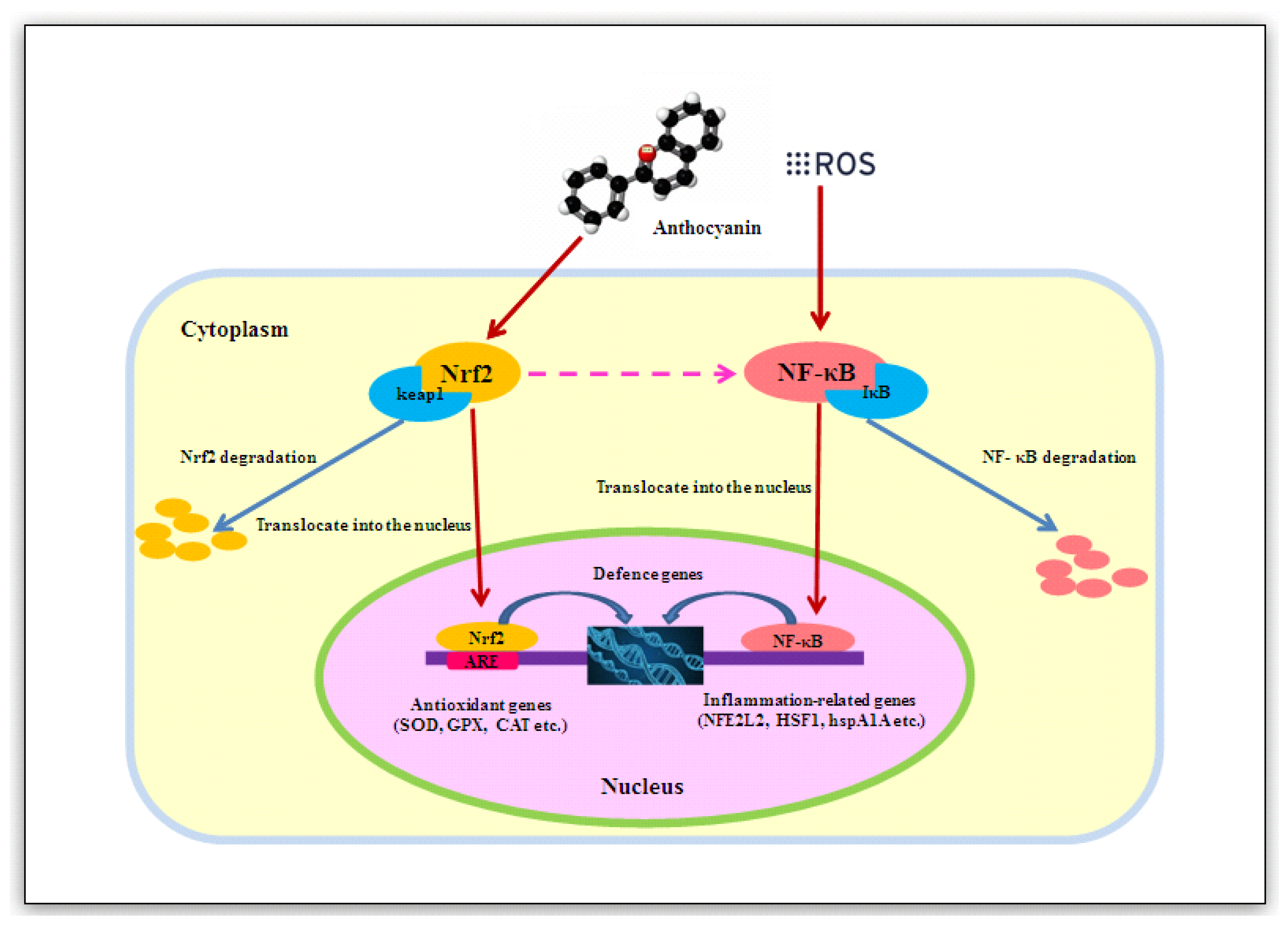
6. Effect of Anthocyanins on the Nrf2 and NF-κB Signalling Pathways
7. Effect of Anthocyanins on Milk Production
8. Effect of Anthocyanins on Milk Oxidation
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Filomeni, G.; Zio, D.D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef]
- Sundrum, A. Metabolic disorders in the transition period indicate that the dairy cows’ ability to adapt is overstressed. Animals 2015, 5, 978–1020. [Google Scholar] [CrossRef]
- Pedernera, M.; Celi, P.; García, S.C.; Salvin, H.E.; Barchia, I.; Fulkerson, W.J. Effect of diet, energy balance and milk production on oxidative stress in early-lactating dairy cows grazing pasture. Vet. J. 2010, 186, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Contreras, G.A.; Aitken, S.L. Metabolic factors affecting the inflammatory response of periparturient dairy cows. Anim. Health Res. Rev. 2009, 10, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [CrossRef]
- Tian, X.Z.; Li, J.X.; Luo, Q.Y.; Wang, X.; Xiao, M.M.; Zhou, D.; Lu, Q.; Chen, X. Effect of supplementation with selenium-yeast on muscle antioxidant activity, meat quality, fatty acids and amino acids in goats. Front. Vet. Sci. 2022, 8, 813672. [Google Scholar] [CrossRef]
- Tian, X.; Wang, X.; Li, J.; Luo, Q.; Ban, C.; Lu, Q. The effects of selenium on rumen fermentation parameters and microbial metagenome in goats. Fermentation 2022, 8, 240. [Google Scholar] [CrossRef]
- De Beer, D.; Joubert, E.; Gelderblom, W.C.A.; Manley, M. Phenolic compounds: A review of their possible role as in vivo antioxidants of wine. S. Afr. J. Enol. Vitic. 2017, 23, 48–61. [Google Scholar] [CrossRef][Green Version]
- Xiao, J.; Khan, M.Z.; Ma, Y.; Alugongo, G.M.; Ma, J.; Chen, T.; Khan, A.; Cao, Z. The antioxidant properties of selenium and vitamin E; their role in periparturient dairy cattle health regulation. Antioxidants 2021, 10, 1555. [Google Scholar] [CrossRef]
- Suman, M.; Tyagi, A.K.; Phondba, B.T. Polyphenols rich plants extract supplementation to enhance the desaturation and antioxidant activity in goat kids. Indian J. Anim. Sci. 2015, 85, 593–600. [Google Scholar]
- Stoldt, A.K.; Mielenz, M.; Nürnberg, G.; Sauerwein, H.; Esatbeyoglu, T.; Wagner, A.E.; Rimbach, G.; Starke, A.; Wolffram, S.; Metges, C.C. Effects of a six-week intraduodenal supplementation with quercetin on liver lipid metabolism and oxidative stress in peripartal dairy cows. J. Anim. Sci. 2016, 94, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.Z.; Lu, Q.; Zhao, S.G.; Li, J.X.; Luo, Q.Y.; Wang, X.; Zhang, Y.D.; Zheng, N. Purple corn anthocyanin affects lipid mechanism, flavor compound profiles, and related gene expression of longissimus thoracis et lumborum muscle in goats. Animals 2021, 11, 2407. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Li, J.; Li, H.; Zhou, D.; Wang, X.; Tian, Y.; Qin, J.; Tian, X.; Lu, Q. The effects of purple corn pigment on growth performance, blood biochemical indices, meat quality, muscle amino acids, and fatty acids of growing chickens. Foods 2022, 11, 1870. [Google Scholar] [CrossRef] [PubMed]
- Canuto, G.A.; Oliveira, D.R.; da Conceição, L.S.; Farah, J.P.; Tavares, M.F. Development and validation of a liquid chromatography method for anthocyanins in strawberry (Fragaria spp.) and complementary studies on stability, kinetics and antioxidant power. Food Chem. 2016, 192, 566–574. [Google Scholar] [CrossRef]
- Tian, X.Z.; Paengkoum, P.; Paengkoum, S.; Thongpe, S.; Ban, C. Comparison of forage yield, silage fermentative quality, anthocyanin stability, antioxidant activity, and in vitro rumen fermentation of anthocyanin-rich purple corn (Zea mays L.) stover and sticky corn stover. J. Integr. Agric. 2018, 17, 2082–2095. [Google Scholar] [CrossRef]
- Leatherwood, W.L. The Effect of Anthocyanins from Purple-Fleshed Sweetpotato on In Vitro Fermentation by Rumen Microbial Cultures. Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 2013. [Google Scholar]
- Lv, Y.; Chen, J.; Zhu, M.; Liu, Y.; Wu, X.; Xiao, X.; Yuyama, N.; Liu, F.; Jing, H.; Cai, H. The wall-associated kinase-like gene RL1 contributes to red leaves in sorghum. Plant J. 2022, 112, 135–150. [Google Scholar] [CrossRef]
- Houghton, A.; Appelhagen, I.; Martin, C. Natural blues: Structure meets function in anthocyanins. Plants 2021, 10, 726. [Google Scholar] [CrossRef]
- Sancho, R.A.S.; Pastore, G.M. Evaluation of the effects of anthocyanins in type 2 diabetes. Food Res. Int. 2012, 46, 378–386. [Google Scholar] [CrossRef]
- Konczak, I.; Zhang, W. Anthocyanins—More than nature’s colours. BioMed Res. Int. 2004, 5, 239–240. [Google Scholar] [CrossRef]
- Castaneda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; Rodríguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Reque, P.M.; Steffens, R.S.; Jablonski, A.; Flôres, S.H.; Rios, A.D.; de Jong, E.V. Cold storage of blueberry (Vaccinium spp.) fruits and juice: Anthocyanin stability and antioxidant activity. J. Food Compos. Anal. 2014, 33, 111–116. [Google Scholar] [CrossRef]
- Clifford, M.N. Anthocyanins–nature, occurrence and dietary burden. J. Sci. Food Agr. 2000, 80, 1063–1072. [Google Scholar] [CrossRef]
- Taethaisong, N.; Paengkoum, S.; Nakharuthai, C.; Onjai-uea, N.; Thongpea, S.; Sinpru, B.; Surakhunthod, J.; Meethip, W.; Paengkoum, P. Consumption of purple neem foliage rich in anthocyanins improves rumen fermentation, growth performance and plasma antioxidant activity in growing goats. Fermentation 2022, 8, 373. [Google Scholar] [CrossRef]
- Suong, N.T.; Paengkoum, S.; Purba, R.A.; Paengkoum, P. Optimizing anthocyanin-rich black cane (Saccharum sinensis Robx.) silage for ruminants using molasses and iron sulphate: A sustainable alternative. Fermentation 2022, 8, 248. [Google Scholar] [CrossRef]
- Tian, X.; Li, J.; Luo, Q.; Wang, X.; Wang, T.; Zhou, D.; Xie, L.; Ban, C.; Lu, Q. Effects of purple corn anthocyanin on growth performance, meat quality, muscle antioxidant status, and fatty acid profiles in goats. Foods 2022, 11, 1255. [Google Scholar] [CrossRef]
- Laleh, G.H.; Frydoonfar, H.; Heidary, R.; Jameei, R.; Zare, S. The effect of light, temperature, pH and species on stability of anthocyanin pigments in four Berberis species. Pakistan J. Nutr. 2006, 5, 90–92. [Google Scholar]
- Reyes, L.F.; Cisneros-Zevallos, L. Degradation kinetics and colour of anthocyanins in aqueous extracts of purple-and red-flesh potatoes (Solanum tuberosum L.). Food Chem. 2007, 100, 885–894. [Google Scholar] [CrossRef]
- Brouillard, R.; Cheminat, A. Flavonoids and flower color. Prog. Clin. Biol. Res. 1988, 280, 93–106. [Google Scholar]
- Krause, K.M.; Oetzel, G.R. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Technol. 2006, 126, 215–236. [Google Scholar] [CrossRef]
- Passamonti, S.; Vrhovsek, U.; Vanzo, A.; Mattivi, F. The stomach as a site for anthocyanins absorption from food. FEBS Lett. 2003, 544, 210–213. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [PubMed]
- Kamalak, A.D.E.M.; Canbolat, O.; Sahin, M.; Gurbuz, Y.; Ozkan, C.O. The effect of polyethylene glycol (PEG 8000) supplementation on in vitro gas production kinetics of leaves from tannin containing trees. S. Afr. J. Anim. Sci. 2005, 35, 229–353. [Google Scholar] [CrossRef]
- Hosoda, K.; Eruden, B.; Matsuyama, H.; Shioya, S. Silage fermentative quality and characteristics of anthocyanin stability in anthocyanin-rich corn (Zea mays L.). Asian-Austral. J. Anim. Sci. 2009, 22, 528–533. [Google Scholar]
- Ichiyanagi, T.; Rahman, M.M.; Kashiwada, Y.; Ikeshiro, Y.; Shida, Y.; Hatano, Y.; Matsumoto, H.; Hirayama, M.; Konishi, T. Absorption and metabolism of delphinidin 3-o-β-d-glucoside in rats. Biofactors 2010, 21, 411–413. [Google Scholar] [CrossRef]
- Dijkstra, J.; Forbes, J.M.; France, J. Quantitative Aspects of Ruminant Digestion and Metabolism; CABI: Cambridge, UK, 2005. [Google Scholar]
- Fang, J. Bioavailability of anthocyanins. Drug Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef]
- Tian, X.Z.; Paengkoum, P.; Paengkoum, S.; Chumpawadee, S.; Ban, C.; Thongpea, S. Short communication: Purple corn (Zea mays L.) stover silage with abundant anthocyanins transferring anthocyanin composition to the milk and increasing antioxidant status of lactating dairy goats. J. Dairy Sci. 2019, 102, 413–418. [Google Scholar] [CrossRef]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef]
- Li, S.; Khafipour, E.; Krause, D.O.; Kroeker, A.; Rodriguez-Lecompte, J.C.; Gozho, G.N.; Plaizier, J.C. Effects of subacute ruminal acidosis challenges on fermentation and endotoxins in the rumen and hindgut of dairy cows. J. Dairy Sci. 2012, 95, 294–303. [Google Scholar] [CrossRef]
- Hosoda, K.; Matsuo, M.; Miyaji, M.; Matsuyama, H.; Maeda, H.; Ohta, H.; Kato, H.; Nonaka, K. Fermentative quality of purple rice (Oryza sativa L.) silage and its effects on digestibility, ruminal fermentation and oxidative status markers in sheep: A preliminary study. Grassl. Sci. 2012, 58, 161–169. [Google Scholar] [CrossRef]
- Francisco, A.; Alves, S.P.; Portugal, P.V.; Dentinho, M.T.; Jerónimo, E.; Sengo, S.; Almeida, J.; Bressan, M.C.; Pires, V.M.R.; Alfaia, C.M.; et al. Effects of dietary inclusion of citrus pulp and rockrose soft stems and leaves on lamb meat quality and fatty acid composition. Animal 2017, 12, 872–881. [Google Scholar]
- Tian, X.Z.; Li, J.X.; Luo, Q.Y.; Zhou, D.; Long, Q.M.; Wang, X.; Lu, Q.; Wen, G.L. Effects of purple corn anthocyanin on blood biochemical indexes, ruminal fluid fermentation, and rumen microbiota in goats. Front. Vet. Sci. 2021, 8, 715710. [Google Scholar] [CrossRef]
- Astuti, T.; Juandes, P.; Yelni, G.; Amir, Y.S. The effect of a local biotechnological approach on rumen fluid characteristics (pH, NH3, VFA) of the oil palm fronds as ruminant feed. Int. J. Agr. Innov. Res. 2015, 3, 1703–1705. [Google Scholar]
- Suong, N.T.; Paengkoum, S.; Schonewille, J.T.; Purba, R.A.; Paengkoum, P. Growth performance, blood biochemical indices, rumen bacterial community, and carcass characteristics in goats fed anthocyanin-rich black cane silage. Front. Vet. Sci. 2022, 9, 880838. [Google Scholar] [CrossRef]
- Tayengwa, T.; Mapiye, C. Citrus and winery wastes: Promising dietary supplements for sustainable ruminant animal nutrition, health, production, and meat quality. Sustainability 2018, 10, 3718. [Google Scholar] [CrossRef]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Correddu, F.; Lunesu, M.F.; Buffa, G.; Atzori, A.S.; Nudda, A.; Battacone, G.; Pulina, G. Can agro-industrial by-products rich in polyphenols be advantageously used in the feeding and nutrition of dairy small ruminants? Animals 2020, 10, 131. [Google Scholar] [CrossRef]
- Lu, Q.; Luo, Q.; Li, J.; Wang, X.; Ban, C.; Qin, J.; Tian, Y.; Tian, X.; Chen, X. Evaluation of the chemical composition, bioactive substance, gas production, and rumen fermentation parameters of four types of distiller’s grains. Molecules 2022, 27, 6134. [Google Scholar] [CrossRef]
- Moate, P.J.; Williams, S.R.; Torok, V.A.; Hannah, M.C.; Ribaux, B.E.; Tavendale, M.H.; Eckard, R.J.; Jacobs, J.L.; Auldist, M.J.; Wales, W.J. Grape marc reduces methane emissions when fed to dairy cows. J. Dairy Sci. 2014, 97, 5073–5087. [Google Scholar] [CrossRef]
- Lazalde-Cruz, R.; Miranda-Romero, L.A.; Tirado-González, D.N.; Carrillo-Díaz, M.I.; Medina-Cuéllar, S.E.; Mendoza-Martínez, G.D.; Lara-Bueno, A.; Tirado-Estrada, G.; Salem, A.Z. Potential effects of delphinidin-3-o-sambubioside and cyanidin-3-o-sambubioside of Hibiscus sabdariffa L. on ruminant meat and milk quality. Animals 2021, 11, 2827. [Google Scholar] [CrossRef]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsable for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Juniper, D.T. Revisiting oxidative stress and the use of organic selenium in dairy cow nutrition. Animals 2019, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Majlesi, A.; Yasini, S.P.; Azimpour, S.; Mottaghian, P. Evaluation of oxidative and antioxidant status in dairy calves before and after weaning. Bulg. J. Vet. Med. 2021, 24, 184–190. [Google Scholar] [CrossRef]
- Miller, J.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C. Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Sharma, N. Alternative approach to control intramammary infection in dairy cows-A review. Asian J. Anim. Vet. Adv. 2007, 2, 50–62. [Google Scholar] [CrossRef][Green Version]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, N.K.; Singh, O.P.; Pandey, V.; Verma, P.K. Oxidative stress and antioxidant status during transition period in dairy cows. Asian-Austral. J. Anim. Sci. 2011, 24, 479–484. [Google Scholar] [CrossRef]
- Puppel, K.; Kapusta, A.; Kuczyńska, B. The etiology of oxidative stress in the various species of animals, a review. J. Sci. Food Agr. 2015, 95, 2179–2184. [Google Scholar] [CrossRef]
- Celi, P.; Merlo, M.; Barbato, O.; Gabai, G. Relationship between oxidative stress and the success of artificial insemination in dairy cows in a pasture-based system. Vet. J. 2012, 193, 498–502. [Google Scholar] [CrossRef]
- Gorniak, T.; Meyer, U.; Südekum, K.H.; Dänicke, S. Impact of mild heat stress on dry matter intake, milk yield and milk composition in mid-lactation Holstein dairy cows in a temperate climate. Arch. Anim. Nutr. 2014, 68, 358–369. [Google Scholar] [CrossRef]
- Das, R.; Sailo, L.; Verma, N.; Bharti, P.; Saikia, J. Impact of heat stress on health and performance of dairy animals: A review. Vet. World. 2016, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Garg, A.; Krohn, R.L.; Bagchi, M.; Tran, M.X.; Stohs, S.J. Oxygen free radical scavenging abilities of vitamins C and E, and a grape seed proanthocyanidin extract in vitro. Res. Commun. Mol. Path. 1997, 95, 179–189. [Google Scholar]
- Han, K.H.; Sekikawa, M.; Shimada, K.I.; Hashimoto, M.; Hashimoto, N.; Noda, T.; Tanaka, H.; Fukushima, M. Anthocyanin-rich purple potato flake extract has antioxidant capacity and improves antioxidant potential in rats. Brit. J. Nutr. 2006, 96, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Sakano, K.; Mizutani, M.; Murata, M.; Oikawa, S.; Hiraku, Y.; Kawanishi, S. Procyanidin B2 has anti-and pro-oxidant effects on metal-mediated DNA damage. Free Radic. Biol. Med. 2005, 39, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Pieszka, M.; Gogol, P.; Pietras, M.; Pieszka, M. Valuable components of dried pomaces of chokeberry, black currant, strawberry, apple and carrot as a source of natural antioxidants and nutraceuticals in the animal diet. Ann. Anim. Sci. 2015, 15, 475. [Google Scholar] [CrossRef]
- Li, N.; Alam, J.; Venkatesan, M.I.; Eiguren-Fernandez, A.; Schmitz, D.; Di Stefano, E.; Slaughter, N.; Killeen, E.; Wang, X.; Huang, A.; et al. Nrf2 is a key transcription factor that regulates antioxidant defense in macrophages and epithelial cells: Protecting against the proinflammatory and oxidizing effects of diesel exhaust chemicals. J. Immunol. 2004, 173, 3467–3481. [Google Scholar] [CrossRef]
- Brewer, M.S. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Aboonabi, A.; Singh, I. Chemopreventive role of anthocyanins in atherosclerosis via activation of Nrf2-ARE as an indicator and modulator of redox. Biomed. Pharmacother. 2015, 72, 30–36. [Google Scholar] [CrossRef]
- Tian, X.Z.; Xin, H.L.; Paengkoum, P.; Siwaporn, P.; Ban, C.; Sorasak, T. Effects of anthocyanin-rich purple corn (Zea mays L.) stover silage on nutrient utilization, rumen fermentation, plasma antioxidant capacity, and mammary gland gene expression in dairy goats. J. Anim. Sci. 2019, 97, 1384–1397. [Google Scholar] [CrossRef]
- Hosoda, K.; Eruden, B.; Matsuyama, H.; Shioya, S. Effect of anthocyanin-rich corn silage on digestibility, milk production and plasma enzyme activities in lactating dairy cows. Anim. Sci. J. 2012, 83, 453–459. [Google Scholar]
- Gessner, D.K.; Koch, C.; Romberg, F.J.; Winkler, A.; Dusel, G.; Herzog, E.; Most, E.; Eder, K. The effect of grape seed and grape marc meal extract on milk performance and the expression of genes of endoplasmic reticulum stress and inflammation in the liver of dairy cows in early lactation. J. Dairy Sci. 2015, 98, 8856–8868. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. The importance of the oxidative status of dairy cattle in the periparturient period: Revisiting antioxidant supplementation. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1003–1016. [Google Scholar] [CrossRef]
- Xiao, W. Advances in NF-kappaB signaling transduction and transcription. Cell Mol. Immunol. 2004, 1, 425–435. [Google Scholar]
- Cho, W.; Jin, X.; Pang, J.; Wang, Y.; Mivechi, N.F.; Moskophidis, D. The molecular chaperone heat shock protein 70 controls liver cancer initiation and progression by regulating adaptive DNA damage and mitogen-activated protein kinase/extracellular signal-regulated kinase signaling pathways. Mol. Cell. Biol. 2019, 39, e00391-18. [Google Scholar] [CrossRef]
- Imbert, V.; Rupec, R.A.; Livolsi, A.; Pahl, H.L.; Traenckner, E.B.M.; Mueller-Dieckmann, C.; Farahifar, D.; Rossi, B.; Auberger, P.; Baeuerle, P.A.; et al. Tyrosine phosphorylation of IκB-α activates NF-κB without proteolytic degradation of IκB-α. Cell 1996, 86, 787–798. [Google Scholar] [CrossRef]
- Kuntz, S.; Asseburg, H.; Dold, S.; Römpp, A.; Fröhling, B.; Kunz, C.; Rudloff, S. Inhibition of low-grade inflammation by anthocyanins from grape extract in an in vitro epithelial-endothelial co-culture model. Food Funct. 2015, 6, 1136–1149. [Google Scholar] [CrossRef]
- Speciale, A.; Cimino, F.; Saija, A.; Canali, R.; Virgili, F. Bioavailability and molecular activities of anthocyanins as modulators of endothelial function. Genes Nutr. 2014, 9, 404. [Google Scholar] [CrossRef]
- Gessner, D.K.; Winkler, A.; Koch, C.; Dusel, G.; Liebisch, G.; Ringseis, R.; Eder, K. Analysis of hepatic transcript profile and plasma lipid profile in early lactating dairy cows fed grape seed and grape marc meal extract. BMC Genom. 2017, 18, 253. [Google Scholar] [CrossRef]
- Safa, S.; Kargar, S.; Moghaddam, G.A.; Ciliberti, M.G.; Caroprese, M. Heat stress abatement during the postpartum period: Effects on whole lactation milk yield, indicators of metabolic status, inflammatory cytokines, and biomarkers of the oxidative stress. J. Anim. Sci. 2019, 97, 122–132. [Google Scholar] [CrossRef]
- Matsuba, T.; Kubozono, H.; Saegusa, A.; Obata, K.; Gotoh, K.; Miki, K.; Akiyama, T.; Oba, M. Short communication: Effects of feeding purple corn (Zea mays L.) silage on productivity and blood superoxide dismutase concentration in lactating cows. J. Dairy Sci. 2019, 102, 7179–7182. [Google Scholar] [CrossRef]
- Matra, M.; Wanapat, M. Phytonutrient pellet supplementation enhanced rumen fermentation efficiency and milk production of lactating Holstein-Friesian crossbred cows. Anim Nutr. 2022, 9, 119–126. [Google Scholar] [CrossRef]
- Yoon, I. Optimising rumen function to manage heat stress and milk fat depression. Intl. Dairy Top. 2015, 14, 15–17. [Google Scholar]
- Sejrsen, K.; Hvelplund, T.; Nielsen, M.O. Ruminant Physiology: Digestion, Metabolism and Impact of Nutrition on Gene Expression, Immunology and Stress; Wageningen Academic Publish: Wageningen, The Netherlands, 2006. [Google Scholar]
- Khonkhaeng, B.; Cherdthong, A.; Chantaprasarn, N.; Harvatine, K.J.; Foiklang, S.; Chanjula, P.; Wanapat, M.; So, S.; Polyorach, S. Comparative effect of Volvariella volvacea-treated rice straw and purple corn stover fed at different levels on predicted methane production and milk fatty acid profiles in tropical dairy cows. Livest. Sci. 2021, 251, 104626. [Google Scholar] [CrossRef]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar] [CrossRef]
- Paraskevakis, N. Effects of dietary dried Greek Oregano (Origanum vulgare ssp. hirtum) supplementation on blood and milk enzymatic antioxidant indices, on milk total antioxidant capacity and on productivity in goats. Anim. Feed Sci. Technol. 2015, 209, 90–97. [Google Scholar] [CrossRef]
- Lindmark-Månsson, H.; Åkesson, B. Antioxidative factors in milk. Brit. J. Nutr. 2000, 84, 103–110. [Google Scholar] [CrossRef]
- Gad, A.S.; Sayd, A.F. Antioxidant properties of rosemary and its potential uses as natural antioxidant in dairy products—A review. Food Nutr. Sci. 2015, 6, 179–193. [Google Scholar] [CrossRef]
- Catalá, A. An overview of lipid peroxidation with emphasis in outer segments of photoreceptors and the chemiluminescence assay. Int. J. Biochem. Cell B 2006, 38, 1482–1495. [Google Scholar] [CrossRef]
- Niki, E. Lipid peroxidation: Physiological levels and dual biological effects. Free Radic. Biol. Med. 2009, 47, 469–484. [Google Scholar] [CrossRef]
- Leopold, J.A.; Loscalzo, J. Oxidative risk for atherothrombotic cardiovascular disease. Free Radic. Biol. Med. 2009, 47, 1673–1706. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.W.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R.L. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.Z.; Wang, X.; Ban, C.; Luo, Q.Y.; Li, J.X.; Lu, Q. Effect of purple corn anthocyanin on antioxidant activity, volatile compound and sensory property in milk during storage and light prevention. Front. Nutr. 2022, 9, 862689. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.Z.; Lu, Q.; Paengkoum, P.; Paengkoum, S. Effect of purple corn pigment on change of anthocyanin composition and unsaturated fatty acids during milk storage. J. Dairy Sci. 2020, 103, 7808–7812. [Google Scholar] [CrossRef]
- Prommachart, R.; Cherdthong, A.; Navanukraw, C.; Pongdontri, P.; Taron, W.; Uriyapongson, J.; Uriyapongson, S. Effect of dietary anthocyanin-extracted residue on meat oxidation and fatty acid profile of male dairy cattle. Animals 2021, 11, 322. [Google Scholar] [CrossRef] [PubMed]
- Bridle, P.; Timberlake, C.F. Anthocyanins as natural food colours—Selected aspects. Food Chem. 1997, 58, 103–109. [Google Scholar] [CrossRef]
- Torel, J.; Cillard, J.; Cillard, P. Antioxidant activity of flavonoids and reactivity with peroxy radical. Phytochemistry 1986, 25, 383–385. [Google Scholar] [CrossRef]
- Narayan, M.S.; Naidu, K.A.; Ravishankar, G.A.; Srinivas, L.; Venkataraman, L.V. Antioxidant effect of anthocyanin on enzymatic and non-enzymatic lipid peroxidation. Prostag. Leukotr. ESS 1999, 60, 1–4. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Anthocyanidin | R1 | R2 |
|---|---|---|
| Pelargonidin | H | H |
| Peonidin | OCH3 | H |
| Cyanidin | H | OH |
| Malvidin | OCH3 | OCH3 |
| Petunidin | OH | OCH3 |
| Delphinidin | OH | OH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, X.; Lu, Q. Anthocyanins in Dairy Cow Nutrition: A Review. Agriculture 2022, 12, 1806. https://doi.org/10.3390/agriculture12111806
Tian X, Lu Q. Anthocyanins in Dairy Cow Nutrition: A Review. Agriculture. 2022; 12(11):1806. https://doi.org/10.3390/agriculture12111806
Chicago/Turabian StyleTian, Xingzhou, and Qi Lu. 2022. "Anthocyanins in Dairy Cow Nutrition: A Review" Agriculture 12, no. 11: 1806. https://doi.org/10.3390/agriculture12111806
APA StyleTian, X., & Lu, Q. (2022). Anthocyanins in Dairy Cow Nutrition: A Review. Agriculture, 12(11), 1806. https://doi.org/10.3390/agriculture12111806