
Effects of Fermented Bamboo Shoot Processing Waste on Growth Performance, Serum Parameters, and Gut Microbiota of Weaned Piglets
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Production of Fermented Feed
2.2. Experimental Design
2.3. Determination of Growth Performance
2.4. Determination of Serum Parameters
2.5. Gut Microbiota Composition Analysis
2.6. Data Analysis
3. Results
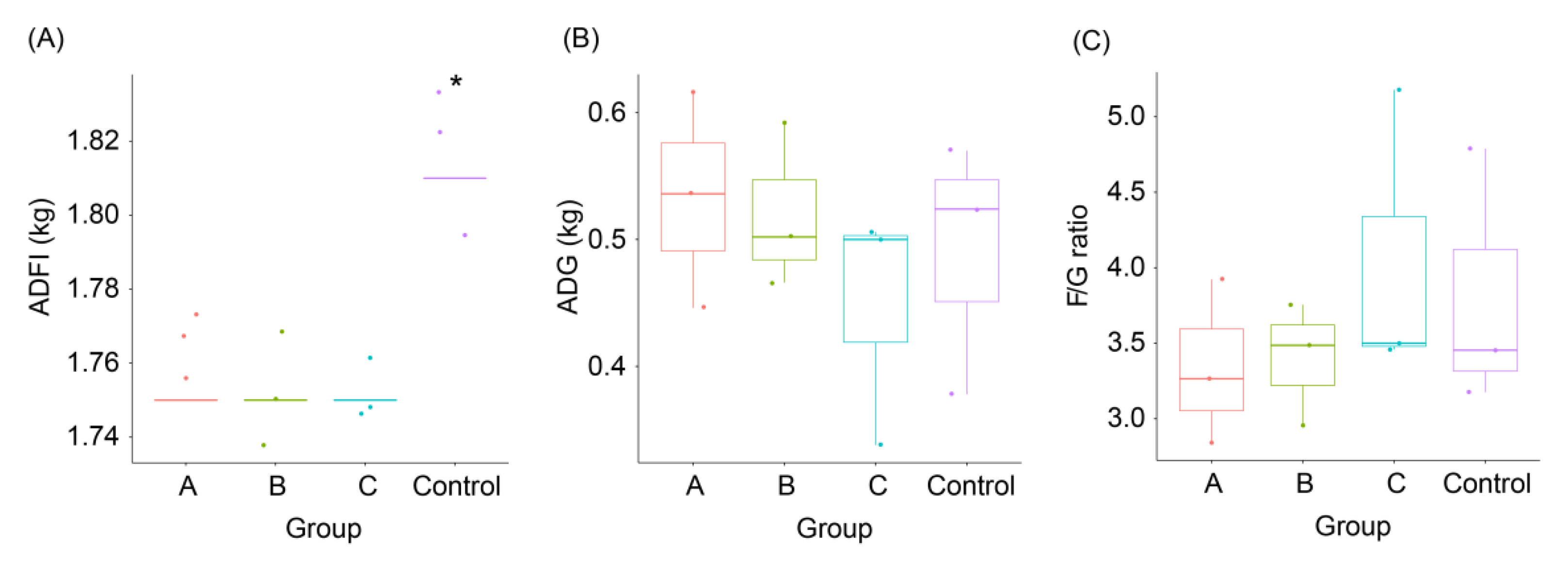
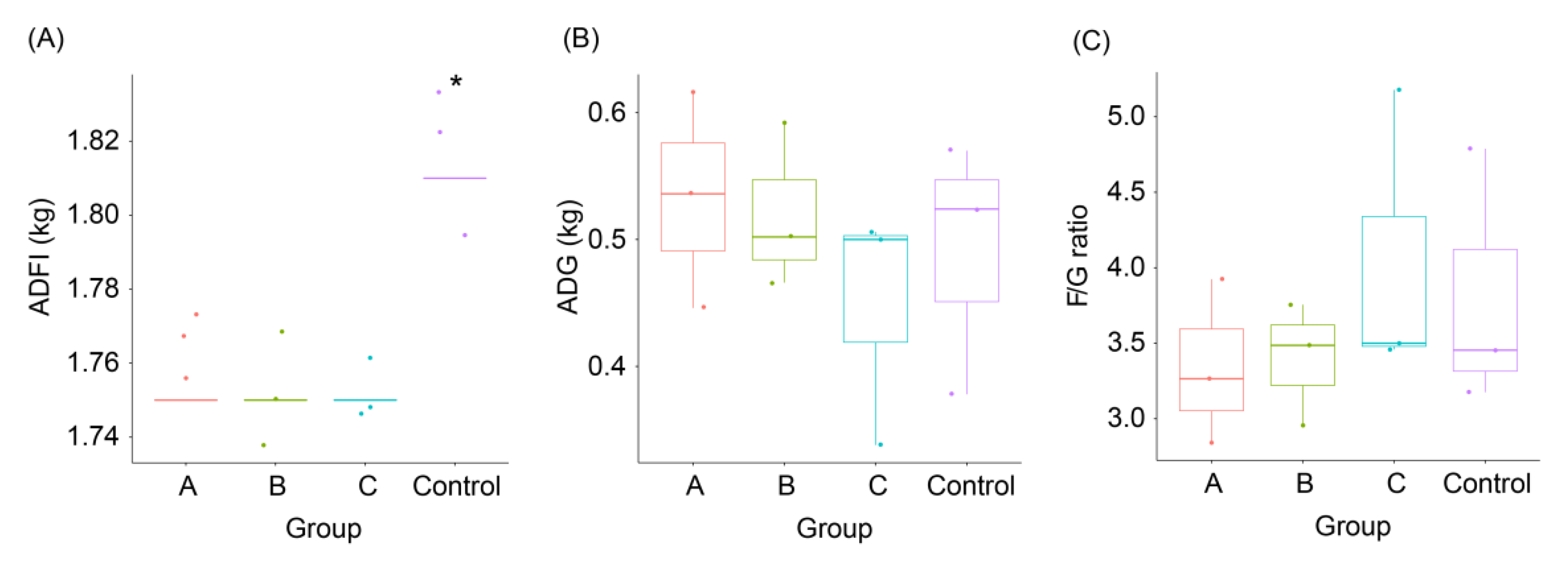
3.1. Effects of FBSPW on Growth Performances of Weaned Piglets
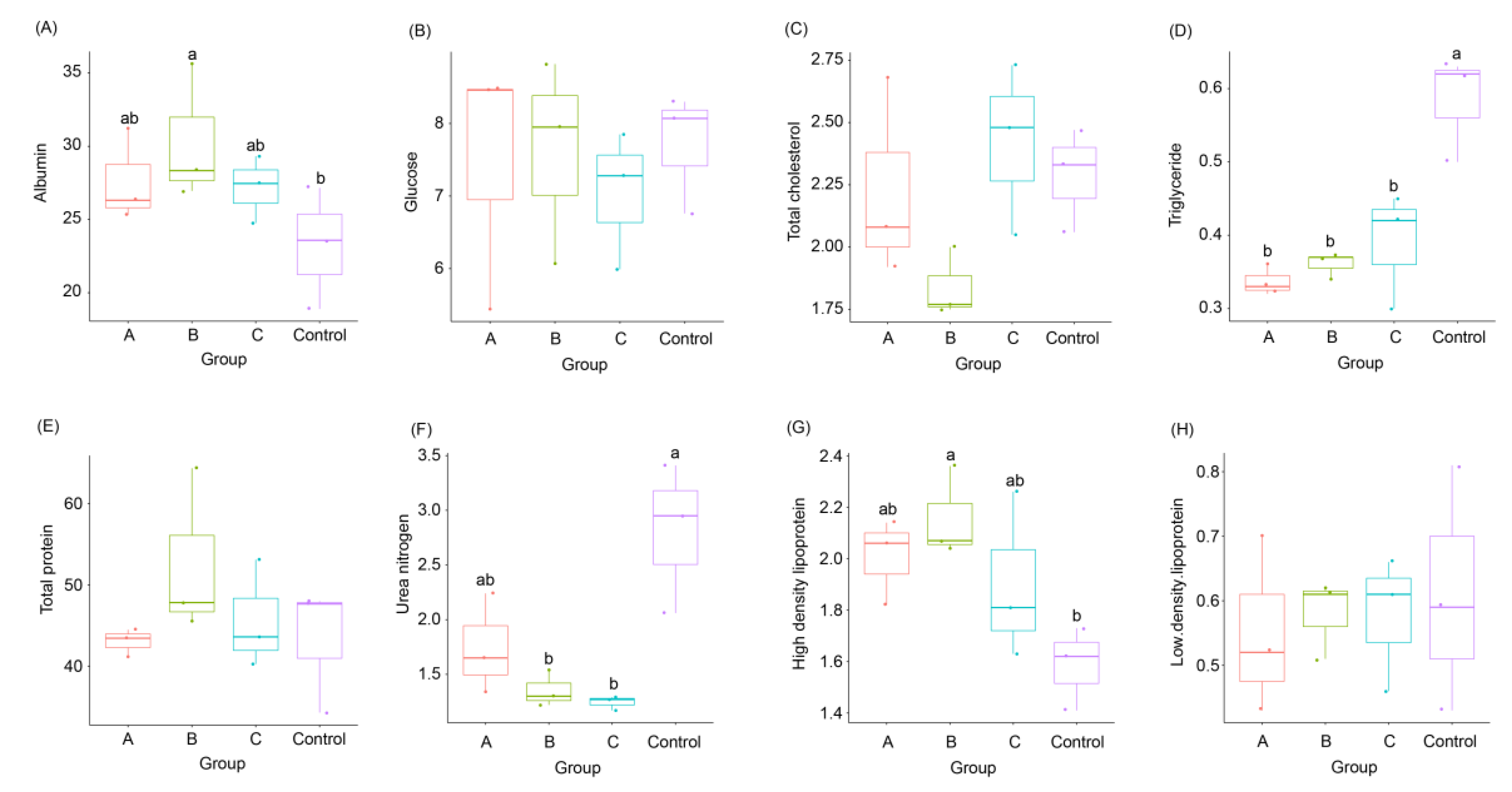
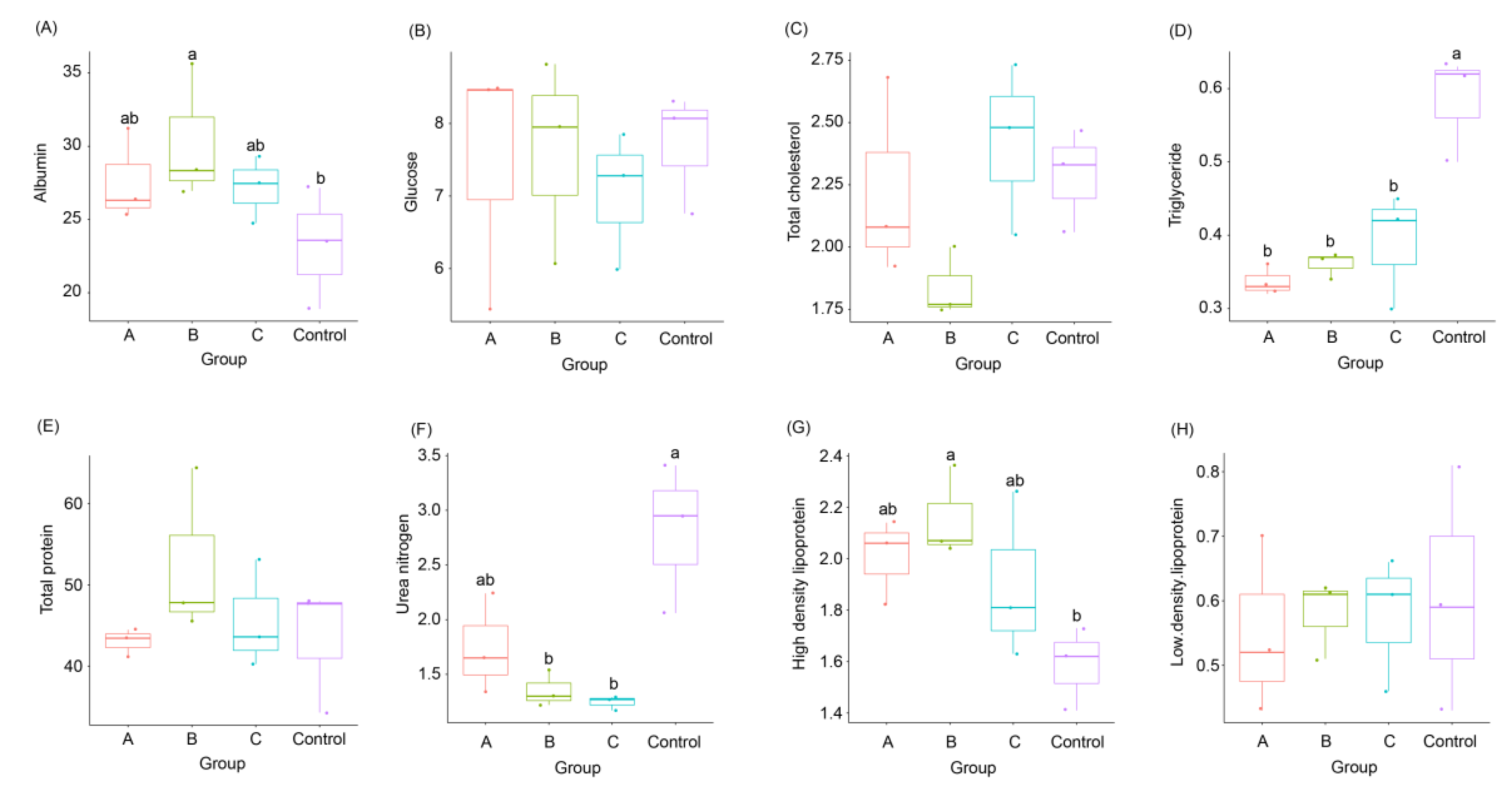
3.2. Effects of FBSPW on Serum Parameters of Weaned Piglets
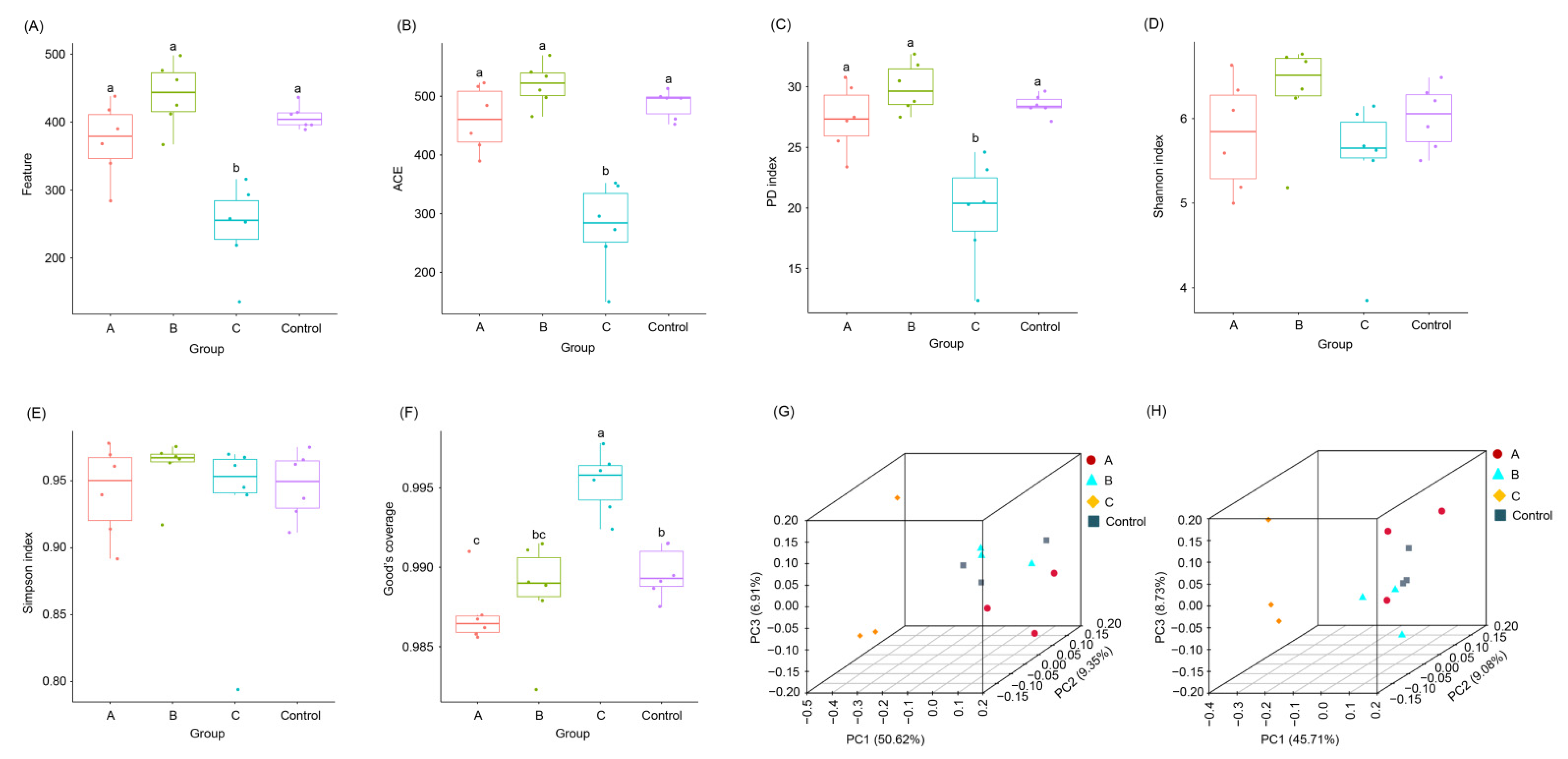
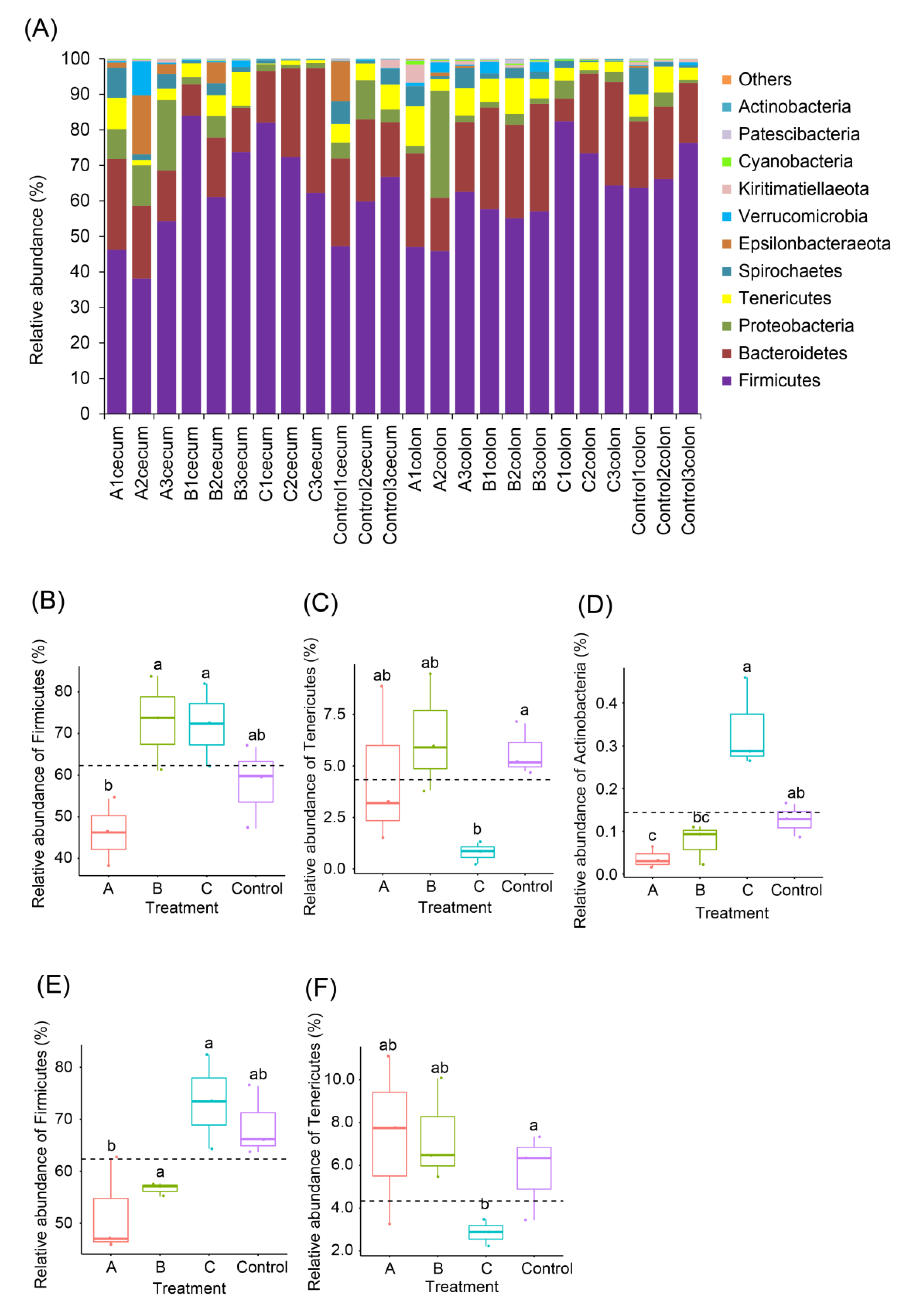
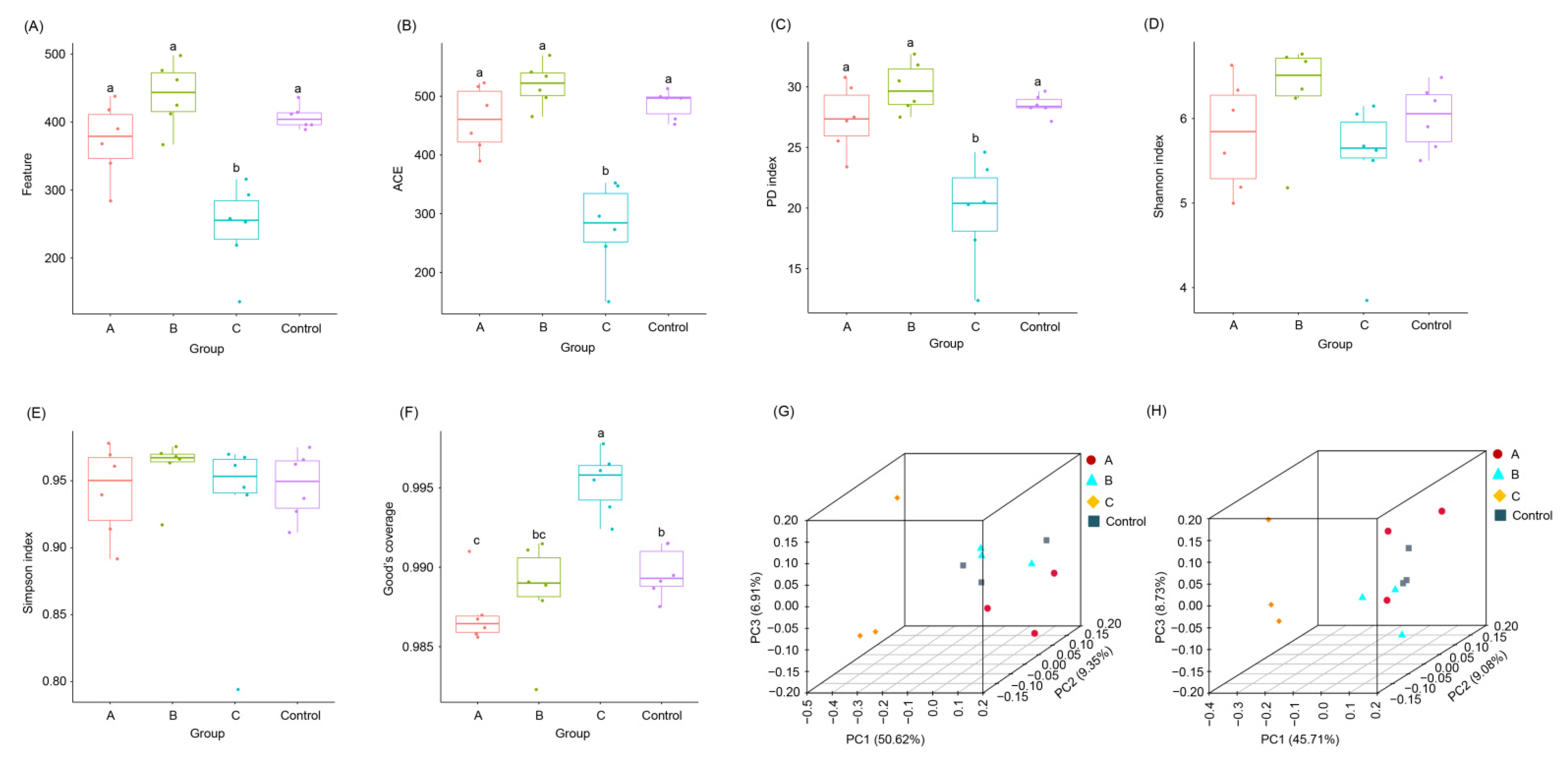
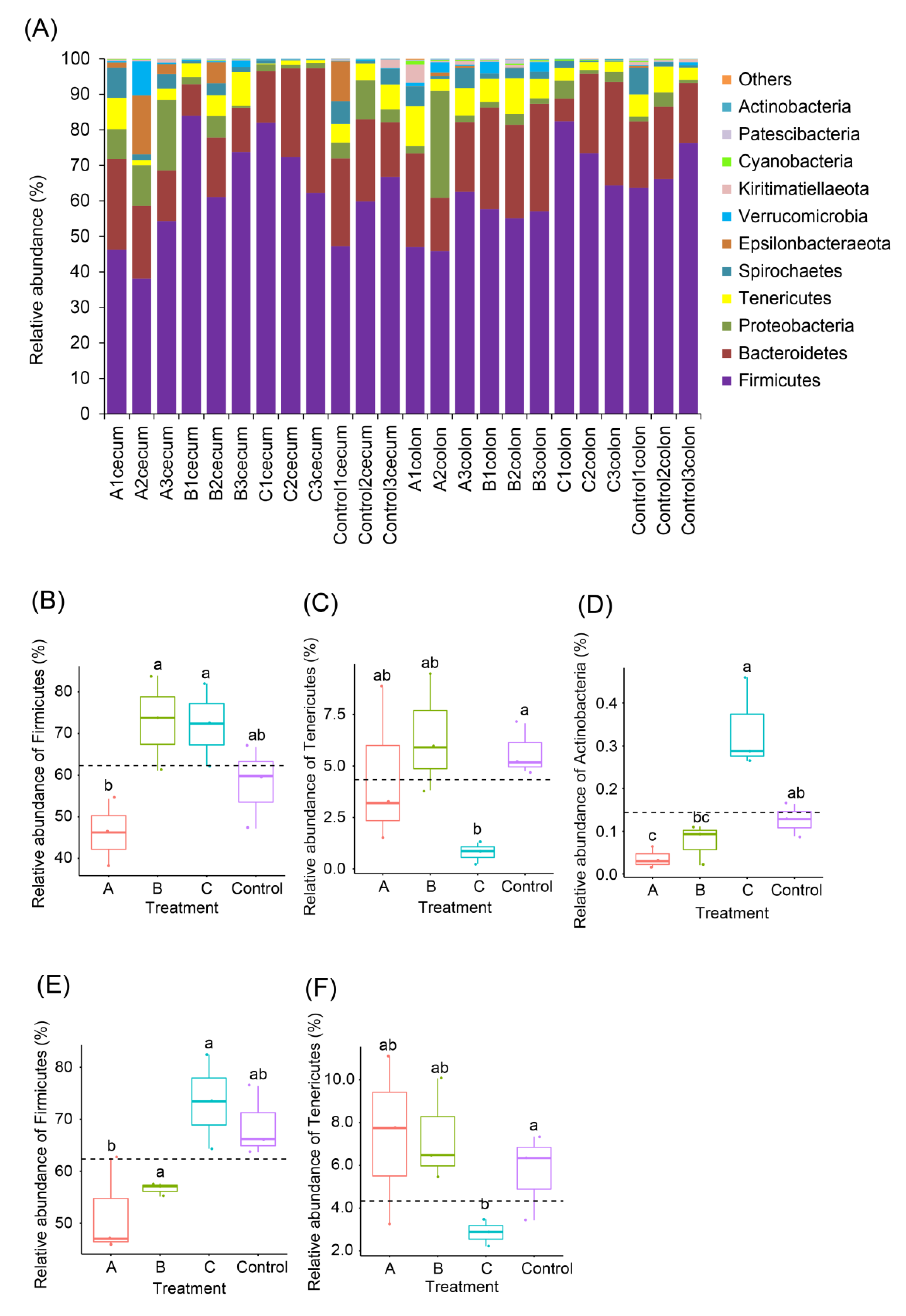
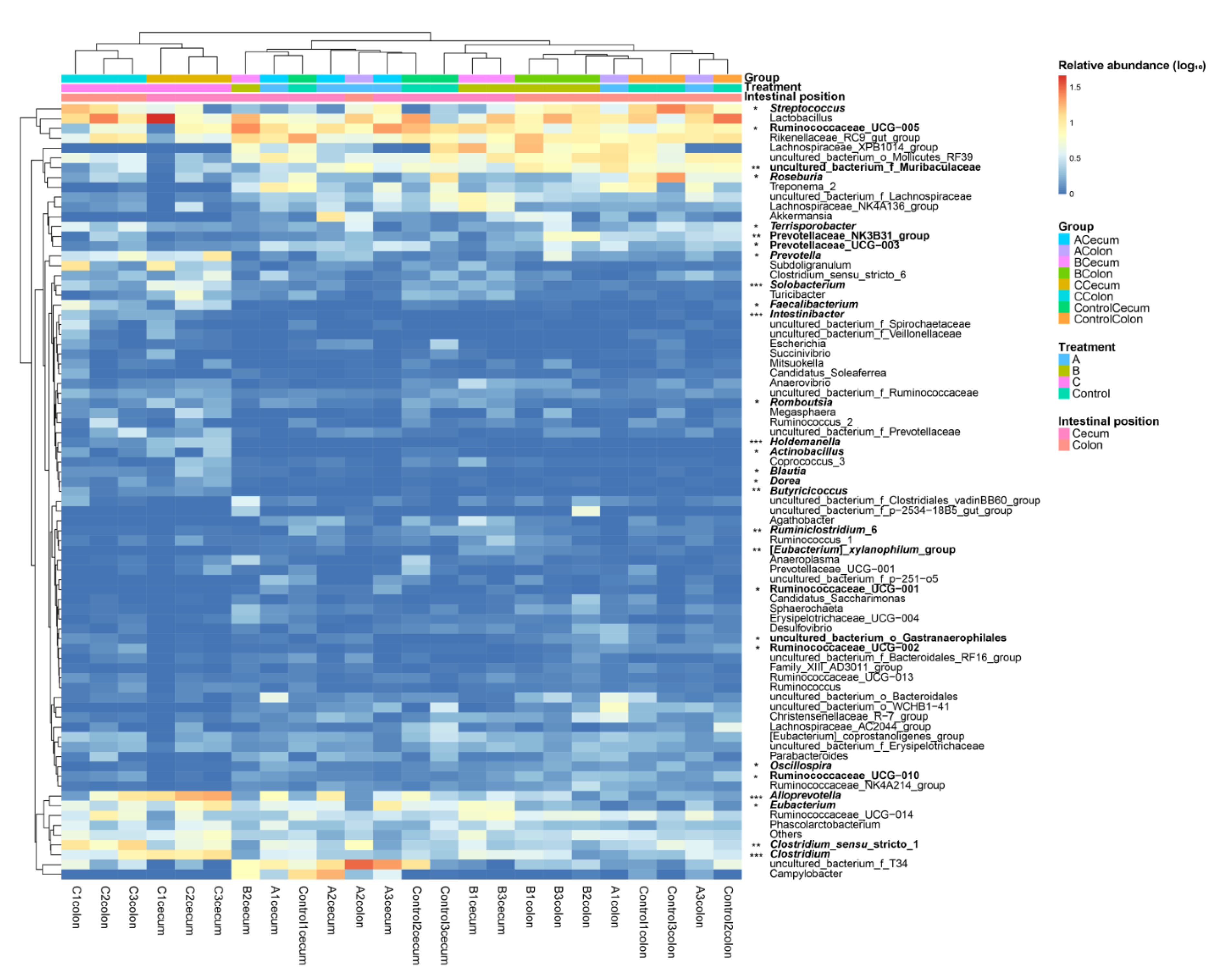
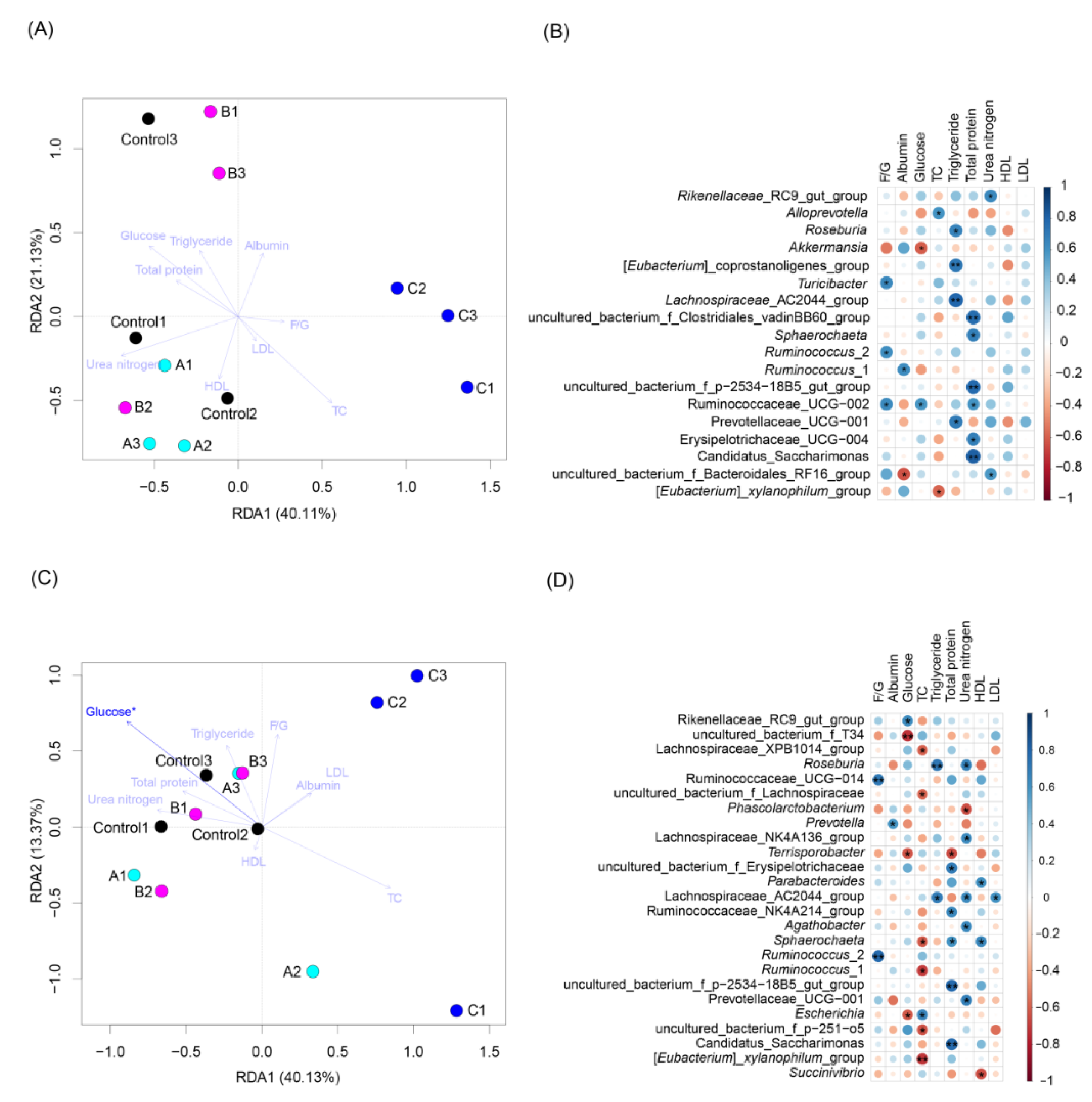
3.3. Effects of FBSPW on Gut Microbiota of Weaned Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choudhury, D.; Sahu, J.K.; Sharma, G.D. Value addition to bamboo shoots: A review. J. Food Sci. Technol. 2012, 49, 407–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.F. Research on the Utilization of Fibers from Bamboo Shoots Processing and Recycling Industry Chain Design. Master’s Thesis, Zhejiang University, Hangzhou, China, 2011. [Google Scholar]
- Xu, L.; Huang, L.; Wang, P. Research progress in extraction method of the dietary fiber from bamoo shoots. China Brew. 2013, 32, 16–18. [Google Scholar]
- Lin, Z.; Chen, J.; Zhang, J.; Brooks, M.S.L. Potential for value-added utilization of bamboo shoot processing waste-recommendations for a biorefinery approach. Food Bioprocess Technol. 2018, 11, 901–912. [Google Scholar] [CrossRef]
- Qi, Y. The Silage and Fermentation of Bamboo Shoot Scraps and Its Naturitional Value Evaluation. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2013. [Google Scholar]
- Wang, Y. The Screening of Strain for Bamboo Shoot Processing Waste Fermentation and Optimization of Fermentation Conditions. Master’s Thesis, Ningbo University, Ningbo, China, 2014. [Google Scholar]
- Luo, X.; Wang, Q.; Zheng, B.; Lin, L.; Chen, B.; Zheng, Y.; Xiao, J. Hydration properties and binding capacities of dietary fibers from bamboo shoot shell and its hypolipidemic effects in mice. Food Chem. Toxicol. 2017, 109, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Bakare, A.G.; Madzimure, J.; Ndou, S.P.; Chimonyo, M. Growth performance and behaviour in grouped pigs fed fibrous diet. Asian-Australas. J. Anim. Sci. 2014, 27, 1204–1210. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-G.; Yu, H.; Zhang, L.; Liu, M.; Qiao, X.-Y.; Cui, W.; Jiang, Y.-P.; Wang, L.; Li, Y.-J.; Tang, L.-J. Probiotic properties of genetically engineered Lactobacillus plantarum producing porcine lactoferrin used as feed additive for piglets. Proc. Biochem. 2016, 51, 719–724. [Google Scholar] [CrossRef]
- Chi, C.-H.; Cho, S.-J. Improvement of bioactivity of soybean meal by solid-state fermentation with Bacillus amyloliquefaciens versus Lactobacillus spp. and Saccharomyces cerevisiae. LWT Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Zhao, J.; Dong, Z.; Chen, L.; Wang, S.; Shao, T. The replacement of whole-plant corn with bamboo shoot shell on the fermentation quality, chemical composition, aerobic stability and in vitro digestibility of total mixed ration silage. Anim. Feed Sci. Technol. 2020, 259, 114348. [Google Scholar] [CrossRef]
- Li, G.; Zou, X.; Kuang, G.; Ren, Y.; Deng, C.; Lin, Q.; Zhao, X.; Xu, S.; Song, J.-L. Preventative effects of fermented Chimonobambusa quadrangularis shoot on activated carbon-induced constipation. Exp. Ther. Med. 2017, 13, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Shi, C.; Zhang, Y.; Song, D.; Lu, Z.; Wang, Y. Microbiota in fermented feed and swine gut. Appl. Microbiol. Biotech. 2018, 102, 2941–2948. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, X.; Liu, Y.; Cao, H.; Han, Q.; Xie, B.; Fan, L.; Li, X.; Hu, J.; Yang, G.; et al. Effect of fermented corn-soybean meal on serum immunity, the expression of genes related to gut immunity, gut microbiota, and bacterial metabolites in grower-finisher pigs. Front. Microbiol. 2019, 10, 2620. [Google Scholar] [CrossRef]
- Qu, Q.; Yang, F.; Zhao, C.; Liu, X.; Yang, P.; Li, Z.; Han, L.; Shi, X. Effects of fermented ginseng on the gut microbiota and immunity of rats with antibiotic-associated diarrhea. J. Ethnopharmacol. 2021, 267, 113594. [Google Scholar] [CrossRef]
- Xu, Q.; Yang, Z.; Chen, S.; Zhu, W.; Xiao, S.; Liu, J.; Wang, H.; Lan, S. Effects of replacing dietary fish meal by soybean meal co-fermented using Bacillus subtilis and Enterococcus faecium on serum antioxidant indices and gut microbitoa of crucian carp Carassius auratus. Fishes 2022, 7, 54. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Agrawal, A.; Gopal, L. Application of diversity index in measurement of species diversity. In Biomonitoring of Water and Waste Water; Springer: New Delhi, India, 2013. [Google Scholar]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Satya, S.; Bal, L.M.; Singhal, P.; Naik, S.N. Bamboo shoot processing: Food quality and safety aspect (a review). Trends Food Sci. Technol. 2010, 21, 181–189. [Google Scholar] [CrossRef]
- Tang, J.; Li, K.; Zhang, Z.; Zheng, S.; Li, Z.; Tong, G. Progress on utilization of bomboo shoot by-products and functional components. Food Mach. 2021, 37, 233–240. [Google Scholar]
- Sarangthem, K.; Singh, T.N. Fermentation decreases the antinutritional content in bamboo shoots. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 361–369. [Google Scholar]
- Farnworth, E.R.; Modler, H.W.; Jones, J.D.; Cave, N.; Yamazaki, H.; Rao, A.V. Feeding Jerusalem artichoke flour rich in fructooligosaccharides to weanling pigs. Can. J. Anim. Sci. 1992, 72, 977–980. [Google Scholar] [CrossRef]
- Johnston, L.J.; Noll, S.; Renteria, A.; Shurson, J. Feeding by-products high in concentration of fiber to nonruminants. Natl. Symp. Altern. Feeds Livest. Poul. 2003, 2003, 169–186. [Google Scholar]
- Kim, J.S.; Ingale, S.L.; Lee, S.H.; Kim, K.H.; Kim, J.S.; Lee, J.H.; Chae, B.J. Effects of energy levels of diet and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites in growing pigs. Anim. Feed Sci. Technol. 2013, 186, 64–70. [Google Scholar] [CrossRef]
- Li, P.; Xue, L.; Zhang, R.; Piao, X.; Zhan, J.S. Effects of fermented potato pulp on performance, nutrient digestibility, carcass traits and plasma parameters of growing-finishing pigs. Asian-Australas. J. Anim. Sci. 2011, 24, 1456–1463. [Google Scholar] [CrossRef]
- Cho, J.H.; Min, B.J.; Chen, Y.J.; Yoo, J.S.; Wang, Q.; Kim, J.D.; Kim, I.H. Evaluation of FSP (fermented soy protein) to replace soybean meal in weaned pigs: Growth performance, blood urea nitrogen and total protein concentrations in serum and nutrient digestibility. Asian-Australas. J. Anim. Sci. 2007, 20, 1874–1879. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Zhao, Y.; Liu, H.; Liu, J.; Makkar, H.P.S.; Becker, K. Effects of replacing soybean meal by detoxified Jatropha curcas kernel meal in the diet of growing pigs on their growth, serum biochemical parameters and visceral organs. Anim. Feed Sci. Technol. 2011, 170, 141–146. [Google Scholar] [CrossRef]
- Lee, C.H.; Kim, H.B.; Ahn, J.H.; Jung, H.J.; Yun, W.; Lee, J.H.; Kwak, W.G.; Oh, H.J.; Liu, S.D.; An, J.S.; et al. Effects of restricted feeding with fermented whole-crop barley and wheat on the growth performance, nutrient digestibility, blood characteristic, and fecal microbiota in finishing pigs. Korean J. Agric. Sci. 2018, 45, 665–675. [Google Scholar]
- Hu, C.H.; Zuo, A.Y.; Wang, D.G.; Pan, H.Y.; Zheng, W.B.; Qian, Z.C.; Zou, X.T. Effects of broccoli stems and leaves meal on production performance and egg quality of laying hens. Anim. Feed Sci. Technol. 2011, 170, 117–121. [Google Scholar] [CrossRef]
- Ting, S.; Yeh, H.; Lien, T. Effects of supplemental levels of hesperetin and naringenin on egg quality, serum traits and antioxidant activity of laying hens. Anim. Feed Sci. Technol. 2011, 163, 59–66. [Google Scholar] [CrossRef]
- Wu, T.-M.; Jiang, J.-J.; Lu, R.-M.; Lin, Y.-H. Effects of dietary inclusion of soybean meal and cholesterol on the growth, cholesterol status and metabolism of the giant grouper (Epinephelus lanceolatus). Aquac. Nutr. 2020, 26, 351–357. [Google Scholar] [CrossRef]
- Akanbi, B.; Agarry, O. Hypocholesterolemic and growth promoting effects of Lactobacillus plantarum AK isolated from a Nigerian fermented cereal product on rats fed high fat diet. Adv. Microbiol. 2014, 4, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Pan, L.; Sheng, Q.H.; Ma, X.K.; Long, S.F.; Xu, Y.T.; Piao, X.S. Effects of isomalto-oligosaccharides as potential prebiotics on performance, immune function and gut microbiota in weaned pigs. Anim. Feed Sci. Technol. 2017, 230, 126–135. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Effects of different probiotic complexes of lactic acid bacteria on growth performance and gut environment of weaned piglets. Livest. Sci. 2010, 133, 182–184. [Google Scholar] [CrossRef]
- Deng, J.; Li, Y.; Zhang, J.; Yang, Q. Co-administration of Bacillus subtilis RJGP16 and Lactobacillus salivarius B1 strongly enhances the intestinal mucosal immunity of piglets. Res. Vet. Sci. 2013, 94, 62–68. [Google Scholar] [CrossRef]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [Green Version]
- Pajarillo, E.A.B.; Chae, J.-P.; Balolong, M.P.; Kim, H.B.; Kang, D.-K. Assessment of fecal bacterial diversity among healthy piglets during the weaning transition. J. Gen. Appl. Microbiol. 2014, 60, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.J.; Li, X.J.; Chen, F.; Wu, H.H.; Xu, M.Y. Community structure and potential nitrogen metabolisms of subtropical aquaculture pond microbiota. Appl. Ecol. Environ. Res. 2018, 16, 7687–7697. [Google Scholar] [CrossRef]
- Ni, J.; Huang, R.; Zhou, H.; Xu, X.; Li, Y.; Cao, P.; Zhong, K.; Ge, M.; Chen, X.; Hou, B.; et al. Analysis of the relationship between the degree of dysbiosis in gut microbiota and prognosis at different stages of primary hepatocellular carcinoma. Front. Microbiol. 2019, 10, 1458. [Google Scholar] [CrossRef]
- Ni, J.; Fu, C.; Huang, R.; Li, Z.; Li, S.; Cao, P.; Zhong, K.; Ge, M.; Gao, Y. Metabolic syndrome cannot mask the changes of faecal microbiota compositions caused by primary hepatocellular carcinoma. Lett. Appl. Microbiol. 2021, 73, 73–80. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Ni, J.; Zhang, C.; Jia, J.; Wu, G.; Sun, H.; Wang, S. Berberine relieves metabolic syndrome in mice by inhibiting liver inflammation caused by a high-fat diet and potential association with gut microbiota. Front. Microbiol. 2022, 12, 752512. [Google Scholar] [CrossRef]
- Mao, L.T.; Huang, Z.C.; Lu, Z.Z.; Lin, G.G.; Chen, Z.F.; Yang, Z. Multi-species Bacillus inoculant and effect on potato growth and control of potato (Solanum tuberosum L.) blackleg disease. Appl. Ecol. Environ. Res. 2022, 20, 1571–1583. [Google Scholar] [CrossRef]
- Tap, J.; Furet, J.-P.; Bensaada, M.; Philippe, C.; Roth, H.; Rabot, S.; Lakhdari, O.; Lombard, V.; Henrissat, B.; Corthier, G.; et al. Gut microbiota richness promotes its stability upon increased dietary fibre intake in healthy adults. Environ. Microbiol. 2015, 17, 4954–4964. [Google Scholar] [CrossRef]
- Kelly, J.; Daly, K.; Moran, A.W.; Ryan, S.; Bravo, D.; Shirazi-Beechey, S.P. Composition and diversity of mucosa-associated microbiota along the entire length of the pig gastrointestinal tract; dietary influences. Environ. Microbiol. 2017, 19, 1425–1438. [Google Scholar] [CrossRef] [Green Version]
- Math, R.K.; Islam, S.M.A.; Hong, S.J.; Cho, K.M.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Kim, E.J.; Lee, Y.H.; Yun, H.D. Metagenomic characterization of oyster shell dump reveals predominance of Firmicutes bacteria. Microbiology 2010, 79, 509–519. [Google Scholar] [CrossRef]
- Sharmin, F.; Wakelin, S.; Huygens, F.; Hargreaves, M. Firmicutes dominate the bacterial taxa within sugarcane processing plants. Sci. Rep. 2013, 3, 3107. [Google Scholar] [CrossRef] [Green Version]
- Eeckhaut, V.; Machiels, K.; Perrier, C.; Romero, C.; Maes, S.; Flahou, B.; Steppe, M.; Haesebrouck, F.; Sas, B.; Ducatelle, R.; et al. Butyricicoccus pullicaecorum in inflammatory bowel disease. Gut 2013, 62, 1673. [Google Scholar] [CrossRef]
- Geirnaert, A.; Steyaert, A.; Eeckhaut, V.; Debruyne, B.; Arends, J.B.A.; Van Immerseel, F.; Boon, N.; Van de Wiele, T. Butyricicoccus pullicaecorum, a butyrate producer with probiotic potential, is intrinsically tolerant to stomach and small intestine conditions. Anaerobe 2014, 30, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Su, Y.; Zhu, W. Microbiome-metabolome responses in the cecum and colon of pig to a high resistant starch diet. Front. Microbiol. 2016, 7, 779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Sarmiento, W.; Sáyago-Ayerdi, S.G.; Goñi, I.; Gutiérrez-Miceli, F.A.; Abud-Archila, M.; del Carmen Rejón-Orantes, J.; Rincón-Rosales, R.; Peña-Ocaña, B.A.; Ruíz-Valdiviezo, V.M. Changes in intestinal microbiota and predicted metabolic pathways during colonic fermentation of mango (Mangifera indica L.)—Based bar indigestible fraction. Nutrients 2020, 12, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Kleinsteuber, S.; Centler, F.; Harms, H.; Sträuber, H. Competition between butyrate fermenters and chain-elongating bacteria limits the efficiency of medium-chain carboxylate production. Front. Microbiol. 2020, 11, 336. [Google Scholar] [CrossRef]
- Rodriguez, J.; Hiel, S.; Neyrinck, A.M.; Roy, T.L.; Pötgens, S.A.; Leyrolle, Q.; Pachikian, B.D.; Gianfrancesco, M.A.; Cani, P.D.; Paquot, N.; et al. Discovery of the gut microbial signature driving the efficacy of prebiotic intervention in obese patients. Gut 2020, 69, 1975–1987. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Zhang, W.; Fan, M.; Qian, H.; Zhang, H.; Qi, X.; Wang, L. Source of gut microbiota determines oat β-glucan degradation and short chain fatty acid-producing pathway. Food Biosci. 2021, 41, 101010. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Hatanaka, E.; Lambertucci, R.H.; Newsholme, P.; Curi, R. Effects of short chain fatty acids on effector mechanisms of neutrophils. Cell Biochem. Funct. 2009, 27, 48–55. [Google Scholar] [CrossRef]
- Elamin, E.E.; Masclee, A.A.; Dekker, J.; Pieters, H.-J.; Jonkers, D.M. Short-chain fatty acids activate AMP-activated protein kinase and ameliorate ethanol-induced intestinal barrier dysfunction in caco-2 cell monolayers. J. Nutr. 2013, 143, 1872–1881. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Li, S.; Luo, J.; Wu, X.; Liu, L. Effects of dietary fibers and their mixtures on short chain fatty acids and microbiota in mice guts. Food Funct. 2013, 4, 932. [Google Scholar] [CrossRef] [PubMed]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Kurosaki, M.; Nihei, S.; Hasegawa, H.; Shinoda, S.; Haruki, M.; Hirano, N. Enzymatic diversity of the Clostridium thermocellum cellulosome is crucial for the degradation of crystalline cellulose and plant biomass. Sci. Rep. 2016, 6, 35709. [Google Scholar] [CrossRef]
- Lü, Y.; Li, N.; Yuan, X.; Hua, B.; Wang, J.; Ishii, M.; Igarashi, Y.; Cui, Z. Enhancing the cellulose-degrading activity of cellulolytic bacteria CTL-6 (Clostridium thermocellum) by co-culture with non-cellulolytic bacteria W2-10 (Geobacillus sp.). Appl. Biochem. Biotechnol. 2013, 171, 1578–1588. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Basal Diet | A | B | C |
|---|---|---|---|---|
| Ingredients (%) | ||||
| FBSPW | 0 | 4 | 8 | 12 |
| Corn | 65 | 63 | 61 | 59 |
| Soybean meal | 20 | 19 | 18 | 17 |
| Wheat bran | 8 | 7 | 6 | 5 |
| Calcium hydroxide | 3 | 3 | 3 | 3 |
| Salt | 0.35 | 0.35 | 0.35 | 0.35 |
| Soybean oil | 2 | 2 | 2 | 2 |
| Premix 1 | 1.65 | 1.65 | 1.65 | 1.65 |
| Total | 100 | 100 | 100 | 100 |
| Calculated composition | ||||
| ME (MJ/kg) | 13.79 | 13.62 | 13.47 | 13.30 |
| Crude protein (%) | 19.22 | 19.43 | 19.66 | 19.87 |
| Crude fiber (%) | 5.1 | 5.4 | 5.7 | 6.0 |
| ADF (%) | 7.62 | 8.00 | 8.37 | 8.72 |
| NDF (%) | 16.01 | 16.84 | 17.58 | 18.33 |
| Lysine (%) | 1.07 | 1.09 | 1.11 | 1.13 |
| Met (%) | 0.32 | 0.33 | 0.34 | 0.35 |
| Calcium (%) | 0.77 | 0.76 | 0.75 | 0.74 |
| Total phosphorus (%) | 0.68 | 0.66 | 0.65 | 0.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Peng, Y.; Yang, Z.; Chen, S.; Liu, J.; Wang, Z.; Wang, G.; Lan, S. Effects of Fermented Bamboo Shoot Processing Waste on Growth Performance, Serum Parameters, and Gut Microbiota of Weaned Piglets. Animals 2022, 12, 2728. https://doi.org/10.3390/ani12202728
Huang Y, Peng Y, Yang Z, Chen S, Liu J, Wang Z, Wang G, Lan S. Effects of Fermented Bamboo Shoot Processing Waste on Growth Performance, Serum Parameters, and Gut Microbiota of Weaned Piglets. Animals. 2022; 12(20):2728. https://doi.org/10.3390/ani12202728
Chicago/Turabian StyleHuang, Yuanhao, Yingjie Peng, Zheng Yang, Siyu Chen, Jing Liu, Zheng Wang, Gang Wang, and Shile Lan. 2022. "Effects of Fermented Bamboo Shoot Processing Waste on Growth Performance, Serum Parameters, and Gut Microbiota of Weaned Piglets" Animals 12, no. 20: 2728. https://doi.org/10.3390/ani12202728