
Differential Effect of Chronic Morphine on Neuronal Degeneration in Male vs. Female Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Behavior Tests
2.3. Tissue Preparation
2.4. Immunohistochemical Analysis
2.5. TUNEL Detection
2.6. ELISA Analysis of Plasma Samples
2.7. Statistical Analysis
3. Results
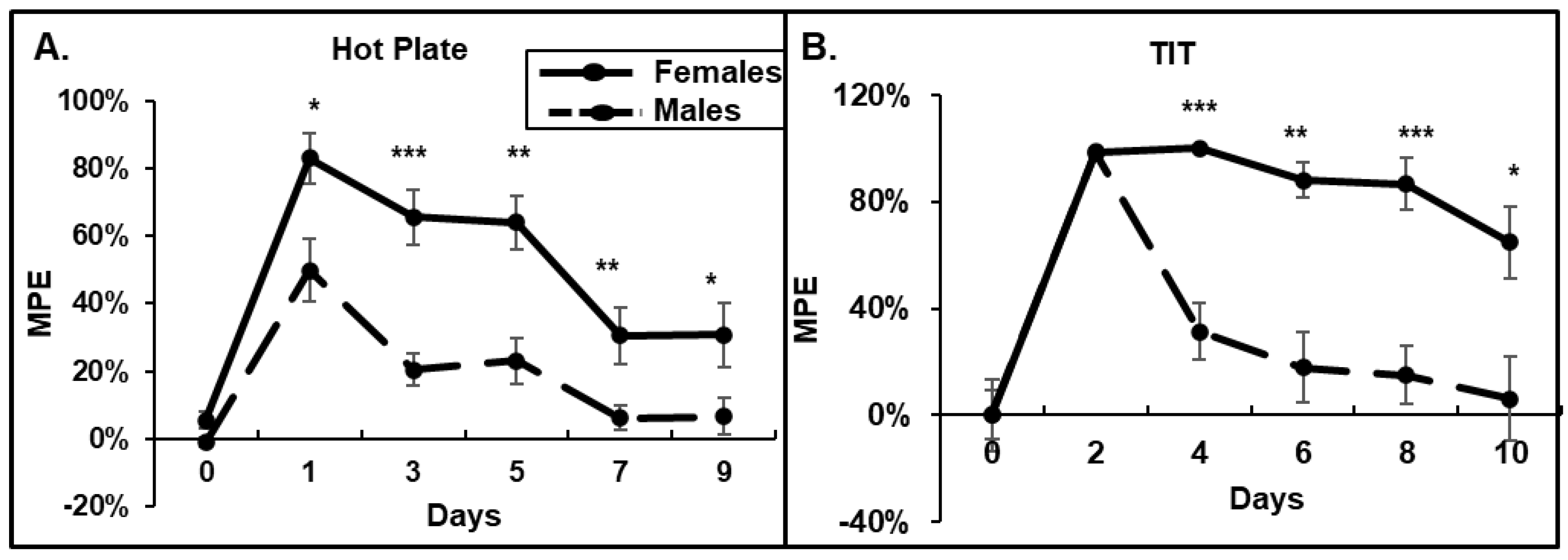
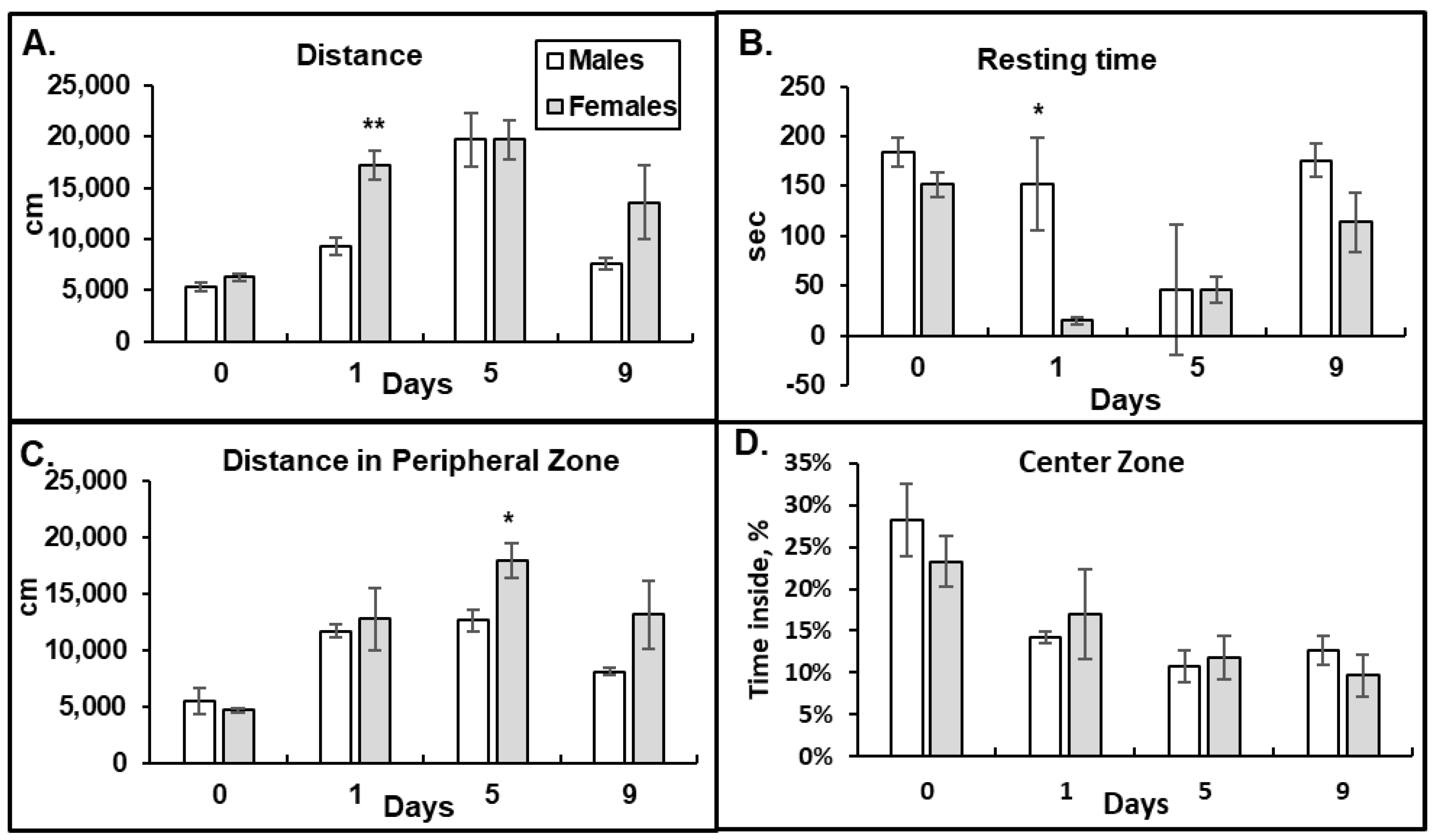
3.1. Behavioral Response to Morphine: Female Mice Have Higher Acute Responses to Morphine and Delayed Developments of Tolerance
3.2. Morphine-Induced Neuronal Degeneration
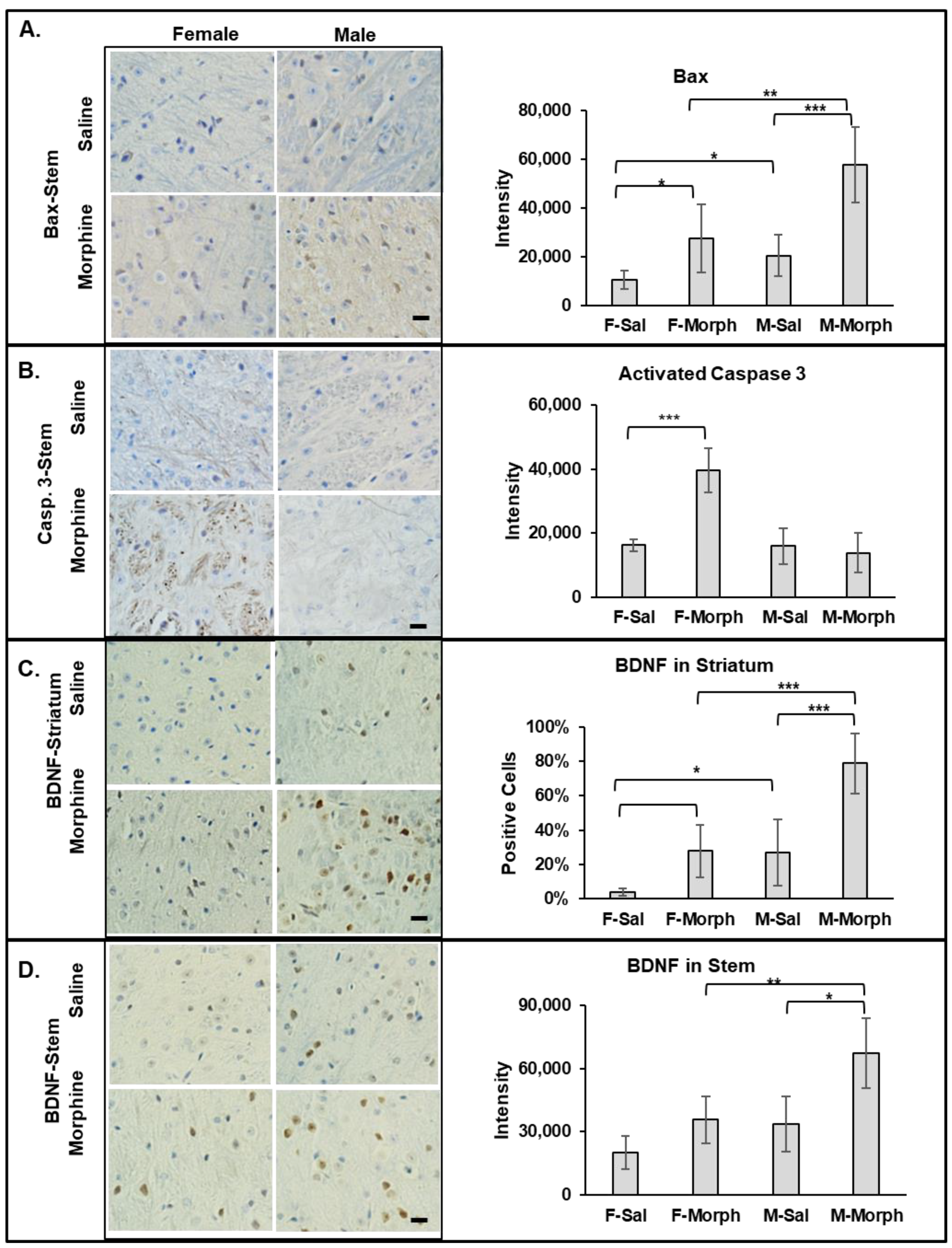
3.2.1. Male Mice Have Higher Constitutive Bax Expression Than Females. Chronic Morphine Increases Bax Expression in Mouse Brains but Not the Number of Pro-Apoptotic Cells
3.2.2. Chronic Morphine Activates Caspase 3 in Female Brains
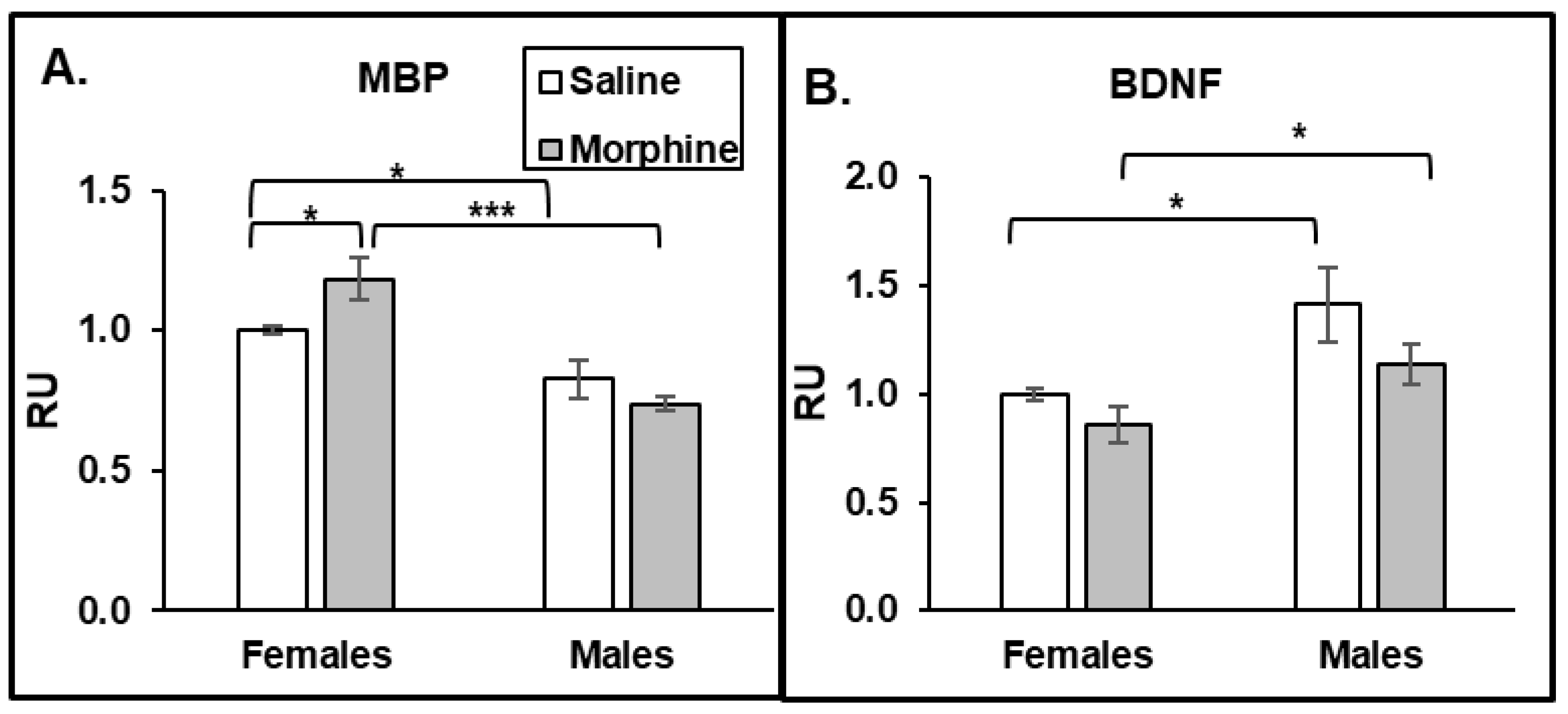
3.2.3. Chronic Morphine Increases BDNF Expression. Male Mice Have Higher Constitutive BDNF Expression Than Females
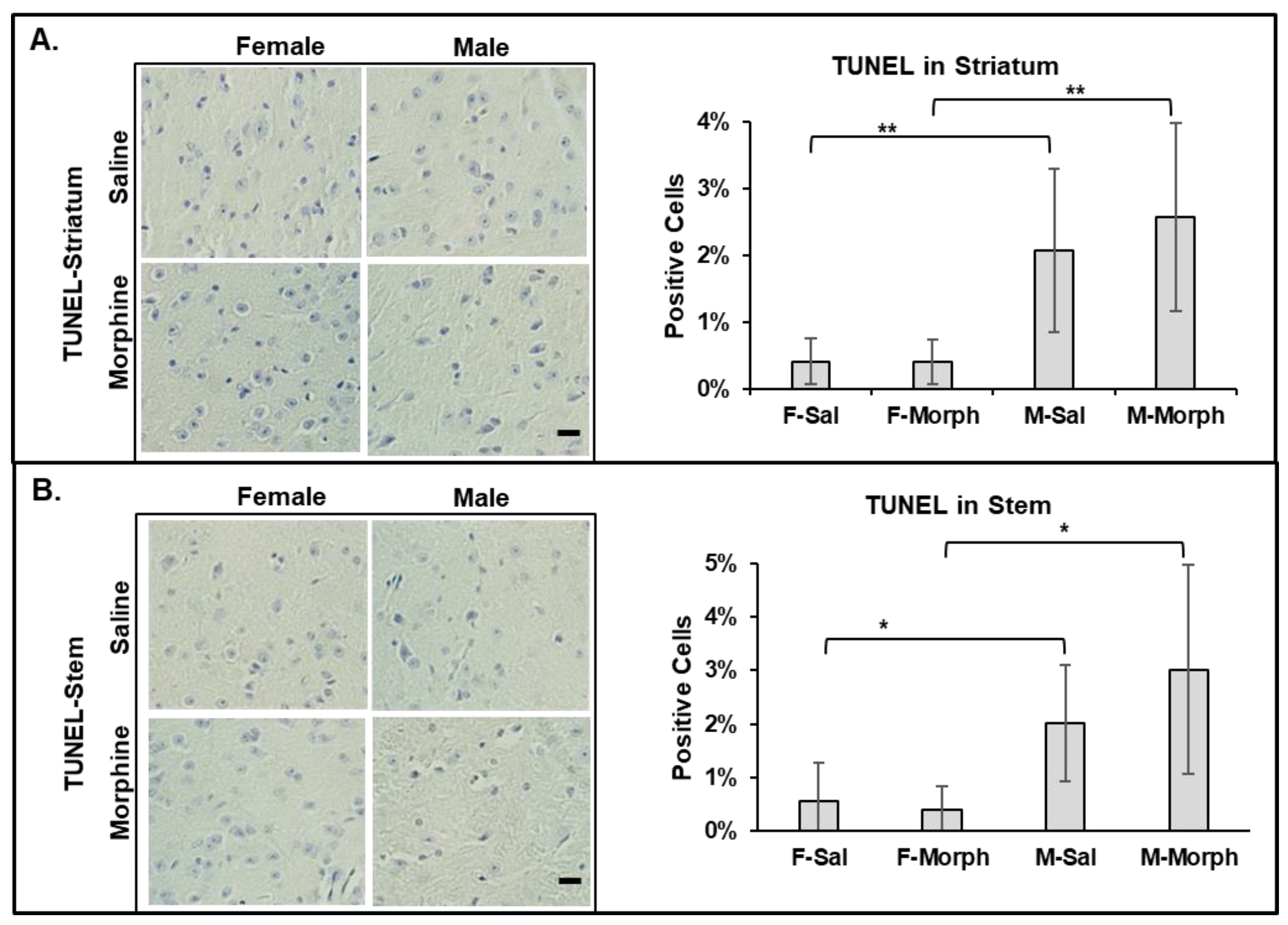
3.2.4. Male Mouse Brains Have Higher Numbers of Apoptotic Cells Than Females: Chronic Morphine Does Not Induce Neuronal Apoptosis
3.2.5. ELISA Plasma
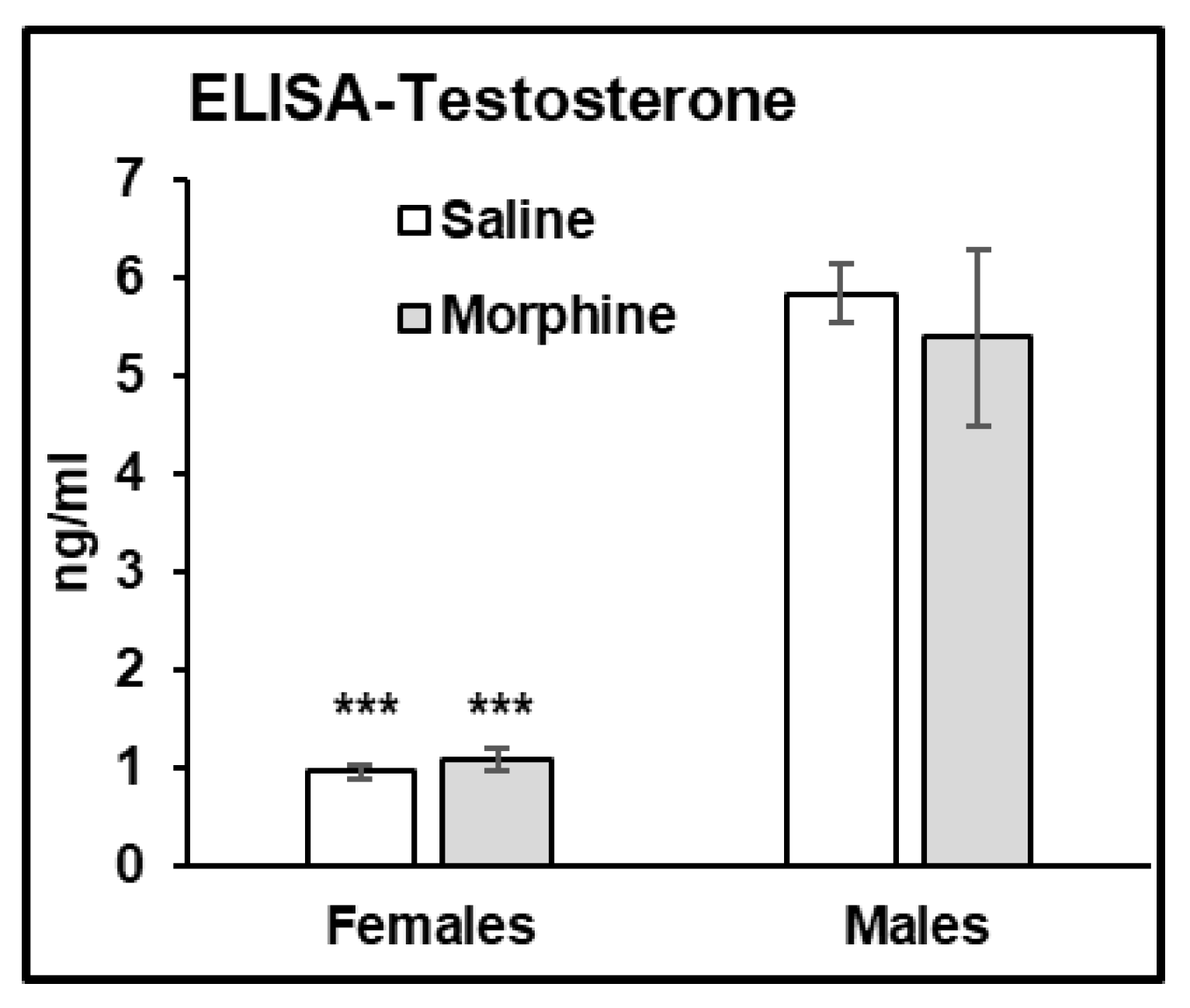
3.2.6. Testosterone Assays
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NIDA. What Is the Scope of Prescription Drug Misuse in the United States? 2023. Available online: https://nida.nih.gov/publications/research-reports/misuse-prescription-drugs/what-scope-prescription-drug-misuse (accessed on 8 December 2023).
- Lee, C.W.; Ho, I.K. Sex differences in opioid analgesia and addiction: Interactions among opioid receptors and estrogen receptors. Mol. Pain 2013, 9, 45. [Google Scholar] [CrossRef]
- Serdarevic, M.; Striley, C.W.; Cottler, L.B. Sex differences in prescription opioid use. Curr. Opin. Psychiatry 2017, 30, 238–246. [Google Scholar] [CrossRef]
- Templeton, K.J. Sex and Gender Issues in Pain Management. J. Bone Joint Surg. Am. 2020, 102 (Suppl. S1), 32–35. [Google Scholar] [CrossRef]
- Goetz, T.G.; Becker, J.B.; Mazure, C.M. Women, opioid use and addiction. Faseb J. 2021, 35, e21303. [Google Scholar] [CrossRef]
- Bobashev, G.V.; Warren, L.K. National polydrug use patterns among people who misuse prescription opioids and people who use heroin. Results from the National Household Survey on Drug Use and Health. Drug Alcohol. Depend. 2022, 238, 109553. [Google Scholar] [CrossRef]
- Loyd, D.R.; Murphy, A.Z. The neuroanatomy of sexual dimorphism in opioid analgesia. Exp. Neurol. 2014, 259, 57–63. [Google Scholar] [CrossRef]
- McHugh, R.K.; Votaw, V.R.; Sugarman, D.E.; Greenfield, S.F. Sex and gender differences in substance use disorders. Clin. Psychol. Rev. 2018, 66, 12–23. [Google Scholar] [CrossRef]
- Krueger, K.; Lamenza, F.; Gu, H.; El-Hodiri, H.; Wester, J.; Oberdick, J.; Fischer, A.J.; Oghumu, S. Sex differences in susceptibility to substance use disorder: Role for X chromosome inactivation and escape? Mol. Cell Neurosci. 2023, 125, 103859. [Google Scholar] [CrossRef]
- NIDA Substance Use in Women DrugFacts. 2020. Available online: https://www.drugabuse.gov/download/19238/substance-use-in-women-drugfacts.pdf?v=8e3444077b0c0e237d1413403790818a (accessed on 12 December 2023).
- Wolters, E.; Stam, F.; Lousberg, R.; Wijngaarden, G.; Rengelink, H.; Schipper, M.; Verbeeten, B. Leucoencephalopathy after inhaling “heroin” pyrolysate. Lancet 1982, 2, 1233–1237. [Google Scholar] [CrossRef]
- Yin, R.; Lu, C.; Chen, Q.; Fan, J.; Lu, J. Microvascular damage is involved in the pathogenesis of heroin induced spongiform leukoencephalopathy. Int. J. Med. Sci. 2013, 10, 299–306. [Google Scholar] [CrossRef]
- Zanin, A.; Masiero, S.; Severino, M.S.; Calderone, M.; Da Dalt, L.; Laverda, A.M. A delayed methadone encephalopathy: Clinical and neuroradiological findings. J. Child. Neurol. 2010, 25, 748–751. [Google Scholar] [CrossRef]
- Mittal, M.; Wang, Y.; Reeves, A.; Newell, K. Methadone-induced delayed posthypoxic encephalopathy: Clinical, radiological, and pathological findings. Case Rep. Med. 2010, 2010, 716494. [Google Scholar] [CrossRef]
- Salgado, R.; Jorens, P.; Baar, I.; Cras, P.; Hans, G.; Parizel, P. Methadone-induced toxic leukoencephalopathy: MR imaging and MR proton spectroscopy findings. AJNR Am. J. Neuroradiol. 2010, 31, 565–566. [Google Scholar] [CrossRef]
- Odia, Y.M.; Jinka, M.; Ziai, W.C. Severe leukoencephalopathy following acute oxycodone intoxication. Neurocrit. Care 2010, 13, 93–97. [Google Scholar] [CrossRef]
- Vestal-Laborde, A.A.; Eschenroeder, A.C.; Bigbee, J.W.; Robinson, S.E.; Sato-Bigbee, C. The opioid system and brain development: Effects of methadone on the oligodendrocyte lineage and the early stages of myelination. Dev. Neurosci. 2014, 36, 409–421. [Google Scholar] [CrossRef]
- Gupta, B.K. Synthetic Heroin-Induced Parkinsonism. Jefferson J. Psychiatry 1986, 4, 11. [Google Scholar] [CrossRef][Green Version]
- Mao, J.; Sung, B.; Ji, R.R.; Lim, G. Neuronal apoptosis associated with morphine tolerance: Evidence for an opioid-induced neurotoxic mechanism. J. Neurosci. 2002, 22, 7650–7661. [Google Scholar] [CrossRef]
- Emeterio, E.P.; Tramullas, M.; Hurlé, M.A. Hurlé, Modulation of apoptosis in the mouse brain after morphine treatments and morphine withdrawal. J. Neurosci. Res. 2006, 83, 1352–1361. [Google Scholar] [CrossRef]
- Chen, Q.; Cui, J.; Zhang, Y.; Yu, L.-C. Prolonged morphine application modulates Bax and Hsp70 levels in primary rat neurons. Neurosci. Lett. 2008, 441, 311–314. [Google Scholar] [CrossRef]
- Liu, L.-W.; Lu, J.; Wang, X.-H.; Fu, S.-K.; Li, Q.; Lin, F.-Q. Neuronal apoptosis in morphine addiction and its molecular mechanism. Int. J. Clin. Exp. Med. 2013, 6, 540–545. [Google Scholar]
- Bajic, D.; Commons, K.G.; Soriano, S.G. Morphine-enhanced apoptosis in selective brain regions of neonatal rats. Int. J. Dev. Neurosci. 2013, 31, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Pu, H.; Wang, X.; Zhang, J.; Ma, C.; Su, Y.; Li, X.; Liu, X.; Su, L. Cerebellar neuronal apoptosis in heroin-addicted rats and its molecular mechanism. Int. J. Clin. Exp. Pathol. 2015, 8, 8260–8267. [Google Scholar]
- Fan, R.; Schrott, L.M.; Snelling, S.; Ndi, J.; Arnold, T.; Korneeva, N.L. Chronic oxycodone induces integrated stress response in rat brain. BMC Neurosci. 2015, 16, 58. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Schrott, L.M.; Arnold, T.; Snelling, S.; Rao, M.; Graham, D.; Cornelius, A.; Korneeva, N.L. Chronic oxycodone induces axonal degeneration in rat brain. BMC Neurosci. 2018, 19, 15. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, M.E.; Morales, P.; Santapau, D.; Ávila, A.; Ponce, C.; Berrios-Cárcamo, P.; Olivares, B.; Gallardo, J.; Ezquer, M.; Herrera-Marschitz, M.; et al. Chronic Voluntary Morphine Intake Is Associated with Changes in Brain Structures Involved in Drug Dependence in a Rat Model of Polydrug Use. Int. J. Mol. Sci. 2023, 24, 17081. [Google Scholar] [CrossRef] [PubMed]
- Zelco, A.; Wapeesittipan, P.; Joshi, A. Insights into Sex and Gender Differences in Brain and Psychopathologies Using Big Data. Life 2023, 13, 1676. [Google Scholar] [CrossRef] [PubMed]
- Beam, C.R.; Kaneshiro, C.; Jang, J.Y.; Reynolds, C.A.; Pedersen, N.L.; Gatz, M. Differences Between Women and Men in Incidence Rates of Dementia and Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 64, 1077–1083. [Google Scholar] [CrossRef]
- Young, J.E.; Wu, M.; Hunsberger, H.C. Editorial: Sex and gender differences in neurodegenerative diseases. Front. Neurosci. 2023, 17, 1175674. [Google Scholar] [CrossRef]
- Whitley, H.; Lindsey, W. Sex-based differences in drug activity. Am. Fam. Physician 2009, 80, 1254–1258. [Google Scholar]
- NIDA. Drug Overdose Death Rates. 2023. Available online: https://nida.nih.gov/research-topics/trends-statistics/overdose-death-rates (accessed on 7 December 2023).
- Smith, Y.R.; Stohler, C.S.; Nichols, T.E.; Bueller, J.A.; Koeppe, R.A.; Zubieta, J.-K. Pronociceptive and antinociceptive effects of estradiol through endogenous opioid neurotransmission in women. J. Neurosci. 2006, 26, 5777–5785. [Google Scholar] [CrossRef]
- Barrett, A.C.; Cook, C.D.; Terner, J.M.; Craft, R.M.; Picker, M.J. Importance of sex and relative efficacy at the mu opioid receptor in the development of tolerance and cross-tolerance to the antinociceptive effects of opioids. Psychopharmacology 2001, 158, 154–164. [Google Scholar] [CrossRef]
- Kest, B.; Palmese, C.; Hopkins, E. A comparison of morphine analgesic tolerance in male and female mice. Brain Res. 2000, 879, 17–22. [Google Scholar] [CrossRef]
- Liu, S.; Seidlitz, J.; Blumenthal, J.D.; Clasen, L.S.; Raznahan, A. Integrative structural, functional, and transcriptomic analyses of sex-biased brain organization in humans. Proc. Natl. Acad. Sci. USA 2020, 117, 18788–18798. [Google Scholar] [CrossRef]
- Wapeesittipan, P.; Joshi, A. Integrated analysis of robust sex-biased gene signatures in human brain. Biol. Sex. Differ. 2023, 14, 36. [Google Scholar] [CrossRef]
- Vegeto, E.; Villa, A.; Della Torre, S.; Crippa, V.; Rusmini, P.; Cristofani, R.; Galbiati, M.; Maggi, A.; Poletti, A. The Role of Sex and Sex Hormones in Neurodegenerative Diseases. Endocr. Rev. 2020, 41, 273–319. [Google Scholar] [CrossRef]
- Ysrraelit, M.C.; Correale, J. Impact of sex hormones on immune function and multiple sclerosis development. Immunology 2019, 156, 9–22. [Google Scholar] [CrossRef]
- Zahaf, A.; Kassoussi, A.; Hutteau-Hamel, T.; Mellouk, A.; Marie, C.; Zoupi, L.; Tsouki, F.; Mattern, C.; Bobé, P.; Schumacher, M.; et al. Androgens show sex-dependent differences in myelination in immune and non-immune murine models of CNS demyelination. Nat. Commun. 2023, 14, 1592. [Google Scholar] [CrossRef]
- Chisu, V.; Manca, P.; Lepore, G.; Gadau, S.; Zedda, M.; Farina, V. Testosterone induces neuroprotection from oxidative stress. Effects on catalase activity and 3-nitro-L-tyrosine incorporation into alpha-tubulin in a mouse neuroblastoma cell line. Arch. Ital. Biol. 2006, 144, 63–73. [Google Scholar] [PubMed]
- Ogata, T.; Nakamura, Y.; Tsuji, K.; Shibata, T.; Kataoka, K. Steroid hormones protect spinal cord neurons from glutamate toxicity. Neuroscience 1993, 55, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Tomassini, V.; Onesti, E.; Mainero, C.; Giugni, E.; Paolillo, A.; Salvetti, M.; Nicoletti, F.; Pozzilli, C. Sex hormones modulate brain damage in multiple sclerosis: MRI evidence. J. Neurol. Neurosurg. Psychiatry 2005, 76, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Bove, R.; Musallam, A.; Healy, B.; Raghavan, K.; Glanz, B.; Bakshi, R.; Weiner, H.; De Jager, P.; Miller, K.; Chitnis, T. Low testosterone is associated with disability in men with multiple sclerosis. Mult. Scler. 2014, 20, 1584–1592. [Google Scholar] [CrossRef]
- Spritzer, M.D.; Roy, E.A. Testosterone and Adult Neurogenesis. Biomolecules 2020, 10, 225. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, M.; Yang, L.; Zeng, M. GPR68 Improves Nerve Damage and Myelination in an Immature Rat Model Induced by Sevoflurane Anesthesia by Activating cAMP/CREB to Mediate BDNF. ACS Chem. Neurosci. 2022, 13, 423–431. [Google Scholar] [CrossRef]
- Nociti, V.; Romozzi, M. The Role of BDNF in Multiple Sclerosis Neuroinflammation. Int. J. Mol. Sci. 2023, 24, 8447. [Google Scholar] [CrossRef]
- Numakawa, T.; Odaka, H. Brain-Derived Neurotrophic Factor Signaling in the Pathophysiology of Alzheimer’s Disease: Beneficial Effects of Flavonoids for Neuroprotection. Int. J. Mol. Sci. 2021, 22, 5719. [Google Scholar] [CrossRef]
- Nejtek, V.; Hilburn, C.; Gangwani, P.; Underwood, W.; Singh, M.; Patel, G.; Forster, M.J. Is serum brain-derived neurotrophic factor related to craving for or use of alcohol, cocaine, or methamphetamine? Neuropsychiatr. Dis. Treat. 2011, 7, 357–364. [Google Scholar] [CrossRef][Green Version]
- Heberlein, A.; Lenz, B.; Opfermann, B.; Gröschl, M.; Janke, E.; Stange, K.; Groh, A.; Kornhuber, J.; Frieling, H.; Bleich, S.; et al. Association of testosterone and BDNF serum levels with craving during alcohol withdrawal. Alcohol 2016, 54, 67–72. [Google Scholar] [CrossRef]
- Lim, S.Y.; Cengiz, P. Opioid tolerance and opioid-induced hyperalgesia: Is TrkB modulation a potential pharmacological solution? Neuropharmacology 2022, 220, 109260. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brazile, C., Jr.; Fan, R.; Benoit, B.; Arnold, T.; Korneeva, N. Differential Effect of Chronic Morphine on Neuronal Degeneration in Male vs. Female Mice. Pathophysiology 2024, 31, 152-165. https://doi.org/10.3390/pathophysiology31010012
Brazile C Jr., Fan R, Benoit B, Arnold T, Korneeva N. Differential Effect of Chronic Morphine on Neuronal Degeneration in Male vs. Female Mice. Pathophysiology. 2024; 31(1):152-165. https://doi.org/10.3390/pathophysiology31010012
Chicago/Turabian StyleBrazile, Chet, Jr., Ruping Fan, Beau Benoit, Thomas Arnold, and Nadejda Korneeva. 2024. "Differential Effect of Chronic Morphine on Neuronal Degeneration in Male vs. Female Mice" Pathophysiology 31, no. 1: 152-165. https://doi.org/10.3390/pathophysiology31010012
APA StyleBrazile, C., Jr., Fan, R., Benoit, B., Arnold, T., & Korneeva, N. (2024). Differential Effect of Chronic Morphine on Neuronal Degeneration in Male vs. Female Mice. Pathophysiology, 31(1), 152-165. https://doi.org/10.3390/pathophysiology31010012