
The Expressions of NF−κB, COX−2, Sp1, and c−Jun in Pancreatic Ductal Adenocarcinoma and Their Associations with Patient Survival

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Location and Period of Study
2.3. Ethical Approval
2.4. Patient Selection
2.5. Immunohistochemistry
2.6. Evaluation of Staining
2.7. Data Analysis
3. Results
3.1. Clinicopathological Characteristics
3.2. NF−κB, COX−2, Sp1, and c−Jun Expressions in PDAC Tissues
3.3. Correlations between NF−κB, COX−2, Sp1, and c−Jun Expressions in PDAC Tissues
3.4. Difference in NF−κB, COX−2, Sp1, and c−Jun Expressions between PDAC and Normal Tissues
3.5. Associations between NF−κB, COX−2, Sp1, and c−Jun Expressions and the Clinicopathological Characteristics of PDAC Patients
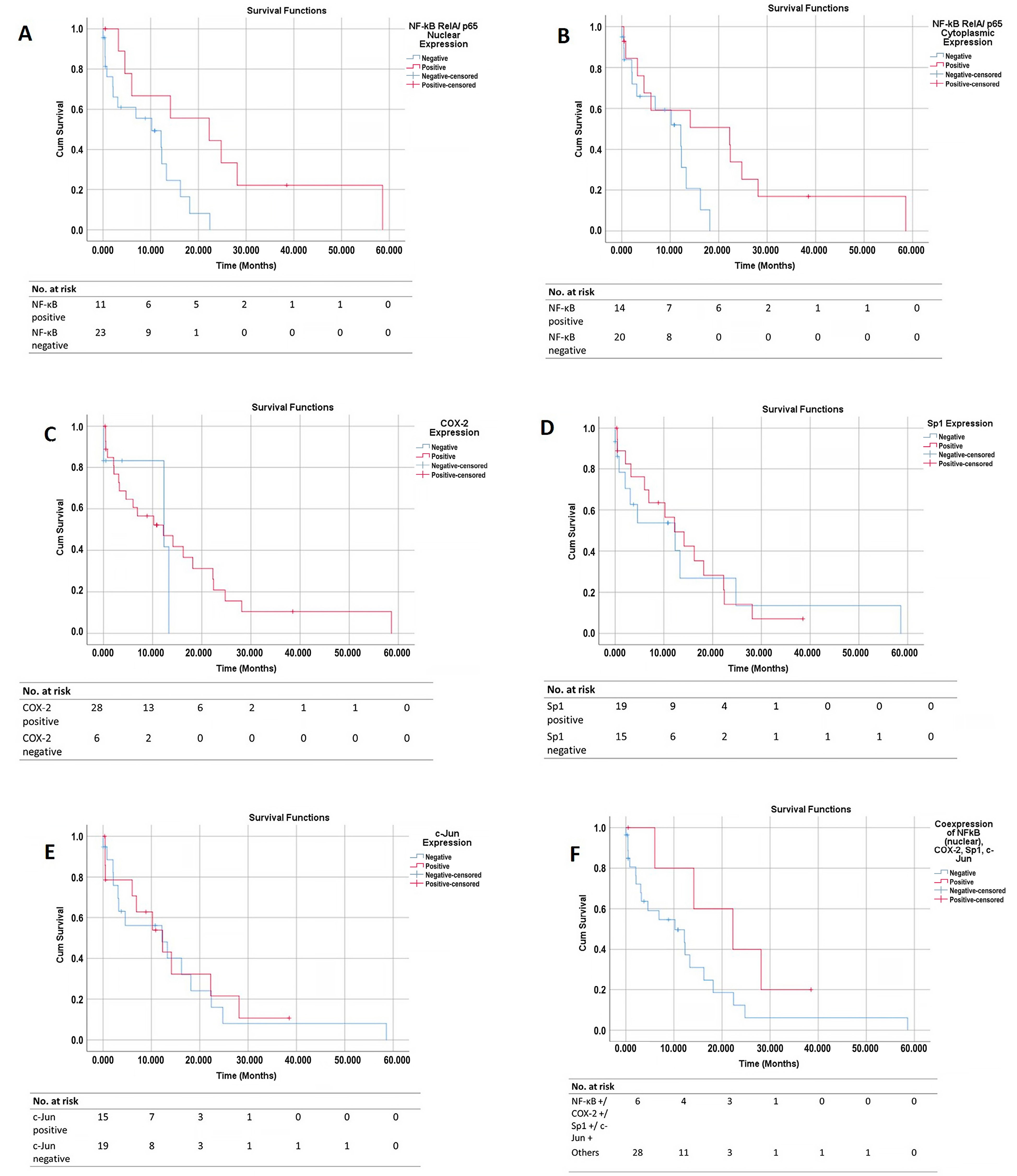
3.6. Survival Curves
3.7. Cox Regression Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pancreas. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/13−Pancreas−fact−sheet.pdf (accessed on 6 June 2022).
- American Cancer Society. Cancer Facts & Figures 2022; American Cancer Society: Atlanta, GA, USA, 2022. [Google Scholar]
- Park, W.; Chawla, A.; O’Reilly, E.M. Pancreatic Cancer: A Review. JAMA 2021, 326, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Eser, S.; Schnieke, A.; Schneider, G.; Saur, D. Oncogenic KRAS signalling in pancreatic cancer. Br. J. Cancer 2014, 111, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Daniluk, J.; Liu, Y.; Chu, J.; Li, Z.; Ji, B.; Logsdon, C.D. Oncogenic K−Ras requires activation for enhanced activity. Oncogene 2014, 33, 532–535. [Google Scholar] [CrossRef] [PubMed]
- Hashemi Goradel, N.; Najafi, M.; Salehi, E.; Farhood, B.; Mortezaee, K. Cyclooxygenase−2 in cancer: A review. J. Cell. Physiol. 2019, 234, 5683–5699. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. Role of prostanoids in gastrointestinal cancer. J. Clin. Investig. 2018, 128, 2732–2742. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.A. Regulation of COX−2 expression in human cancers. Prog. Exp. Tumor Res. 2003, 37, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Maitra, A.; Ashfaq, R.; Gunn, C.R.; Rahman, A.; Yeo, C.J.; Sohn, T.A.; Cameron, J.L.; Hruban, R.H.; Wilentz, R.E. Cyclooxygenase 2 expression in pancreatic adenocarcinoma and pancreatic intraepithelial neoplasia: An immunohistochemical analysis with automated cellular imaging. Am. J. Clin. Pathol. 2002, 118, 194–201. [Google Scholar] [CrossRef]
- Yip−Schneider, M.T.; Barnard, D.S.; Billings, S.D.; Cheng, L.; Heilman, D.K.; Lin, A.; Marshall, S.J.; Crowell, P.L.; Marshall, M.S.; Sweeney, C.J. Cyclooxygenase−2 expression in human pancreatic adenocarcinomas. Carcinogenesis 2000, 21, 139–146. [Google Scholar] [CrossRef]
- Chun, K.S.; Surh, Y.J. Signal transduction pathways regulating cyclooxygenase−2 expression: Potential molecular targets for chemoprevention. Biochem. Pharmacol. 2004, 68, 1089–1100. [Google Scholar] [CrossRef]
- Kang, Y.J.; Mbonye, U.R.; DeLong, C.J.; Wada, M.; Smith, W.L. Regulation of intracellular cyclooxygenase levels by gene transcription and protein degradation. Prog. Lipid Res. 2007, 46, 108–125. [Google Scholar] [CrossRef]
- Li, Q.; Yang, G.; Feng, M.; Zheng, S.; Cao, Z.; Qiu, J.; You, L.; Zheng, L.; Hu, Y.; Zhang, T.; et al. NF−kappaB in pancreatic cancer: Its key role in chemoresistance. Cancer Lett. 2018, 421, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Wu, H.S.; Wang, L.; Tian, Y.; Zhang, J.H.; Wu, H.L. Expression and significance of TLR4 and HIF−1alpha in pancreatic ductal adenocarcinoma. World J. Gastroenterol. 2010, 16, 2881–2888. [Google Scholar] [CrossRef] [PubMed]
- Sankpal, U.T.; Maliakal, P.; Bose, D.; Kayaleh, O.; Buchholz, D.; Basha, R. Expression of specificity protein transcription factors in pancreatic cancer and their association in prognosis and therapy. Curr. Med. Chem. 2012, 19, 3779–3786. [Google Scholar] [CrossRef]
- Hu, J.; Hu, H.; Hang, J.J.; Yang, H.Y.; Wang, Z.Y.; Wang, L.; Chen, D.H.; Wang, L.W. Simultaneous high expression of PLD1 and Sp1 predicts a poor prognosis for pancreatic ductal adenocarcinoma patients. Oncotarget 2016, 7, 78557–78565. [Google Scholar] [CrossRef] [PubMed]
- Eferl, R.; Wagner, E.F. AP−1: A double−edged sword in tumorigenesis. Nat. Rev. Cancer 2003, 3, 859–868. [Google Scholar] [CrossRef]
- Ferrara, C.; Tessari, G.; Poletti, A.; Giacon, C.; Meggiato, T.; Martines, D.; Del Favero, G.; Naccarato, R. Ki−67 and c−jun expression in pancreatic cancer: A prognostic marker? Oncol. Rep. 1999, 6, 1117–1122. [Google Scholar] [CrossRef]
- Fagman, J.B.; Ljungman, D.; Falk, P.; Iresjo, B.M.; Engstrom, C.; Naredi, P.; Lundholm, K. EGFR, but not COX−2, protein in resected pancreatic ductal adenocarcinoma is associated with poor survival. Oncol. Lett. 2019, 17, 5361–5368. [Google Scholar] [CrossRef]
- Juuti, A.; Louhimo, J.; Nordling, S.; Ristimaki, A.; Haglund, C. Cyclooxygenase−2 expression correlates with poor prognosis in pancreatic cancer. J. Clin. Pathol. 2006, 59, 382–386. [Google Scholar] [CrossRef]
- Matsubayashi, H.; Infante, J.R.; Winter, J.; Klein, A.P.; Schulick, R.; Hruban, R.; Visvanathan, K.; Goggins, M. Tumor COX−2 expression and prognosis of patients with resectable pancreatic cancer. Cancer Biol. Ther. 2007, 6, 1569–1575. [Google Scholar] [CrossRef]
- Pomianowska, E.; Schjolberg, A.R.; Clausen, O.P.; Gladhaug, I.P. COX−2 overexpression in resected pancreatic head adenocarcinomas correlates with favourable prognosis. BMC Cancer 2014, 14, 458. [Google Scholar] [CrossRef]
- Wang, D.; Guo, X.Z.; Li, H.Y.; Zhao, J.J.; Shao, X.D.; Wu, C.Y. Prognostic significance of cyclooxygenase−2 protein in pancreatic cancer: A meta−analysis. Tumour Biol. 2014, 35, 10301–10307. [Google Scholar] [CrossRef]
- Renaldi, K.; Simadibrata, M.; Rahadiani, N.; Handjari, D.R.; William, A.; Sinuraya, F.; Makmun, D. Prognostic Value of COX−2, NF−kappaB, and Sp1 Tissue Expressions in Pancreatic Ductal Adenocarcinoma: A Systematic Review and Meta−analysis. Turk. J. Gastroenterol. 2021, 32, 956–970. [Google Scholar] [CrossRef] [PubMed]
- Weichert, W.; Boehm, M.; Gekeler, V.; Bahra, M.; Langrehr, J.; Neuhaus, P.; Denkert, C.; Imre, G.; Weller, C.; Hofmann, H.P.; et al. High expression of RelA/p65 is associated with activation of nuclear factor−kappaB−dependent signaling in pancreatic cancer and marks a patient population with poor prognosis. Br. J. Cancer 2007, 97, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Hang, J.; Hu, H.; Huang, J.; Han, T.; Zhuo, M.; Zhou, Y.; Wang, L.; Wang, Y.; Jiao, F.; Wang, L. Sp1 and COX2 expression is positively correlated with a poor prognosis in pancreatic ductal adenocarcinoma. Oncotarget 2016, 7, 28207–28217. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Han, T.; Zhuo, M.; Wu, L.L.; Yuan, C.; Wu, L.; Lei, W.; Jiao, F.; Wang, L.W. Elevated COX−2 Expression Promotes Angiogenesis Through EGFR/p38−MAPK/Sp1−Dependent Signalling in Pancreatic Cancer. Sci. Rep. 2017, 7, 470. [Google Scholar] [CrossRef]
- Yang, S.H.; Hsu, C.H.; Lee, J.C.; Tien, Y.W.; Kuo, S.H.; Cheng, A.L. Nuclear expression of glioma−associated oncogene homolog 1 and nuclear factor−kappaB is associated with a poor prognosis of pancreatic cancer. Oncology 2013, 85, 86–94. [Google Scholar] [CrossRef]
- Jiang, N.Y.; Woda, B.A.; Banner, B.F.; Whalen, G.F.; Dresser, K.A.; Lu, D. Sp1, a new biomarker that identifies a subset of aggressive pancreatic ductal adenocarcinoma. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1648–1652. [Google Scholar] [CrossRef]
- Hill, R.; Li, Y.; Tran, L.M.; Dry, S.; Calvopina, J.H.; Garcia, A.; Kim, C.; Wang, Y.; Donahue, T.R.; Herschman, H.R.; et al. Cell intrinsic role of COX−2 in pancreatic cancer development. Mol. Cancer Ther. 2012, 11, 2127–2137. [Google Scholar] [CrossRef]
- Bu, X.; Zhao, C.; Dai, X. Involvement of COX−2/PGE(2) Pathway in the Upregulation of MMP−9 Expression in Pancreatic Cancer. Gastroenterol. Res. Pract. 2011, 2011, 214269. [Google Scholar] [CrossRef][Green Version]
- Knab, L.M.; Grippo, P.J.; Bentrem, D.J. Involvement of eicosanoids in the pathogenesis of pancreatic cancer: The roles of cyclooxygenase−2 and 5−lipoxygenase. World J. Gastroenterol. 2014, 20, 10729–10739. [Google Scholar] [CrossRef]
- Prabhu, L.; Mundade, R.; Korc, M.; Loehrer, P.J.; Lu, T. Critical role of NF−kappaB in pancreatic cancer. Oncotarget 2014, 5, 10969–10975. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Asano, T.; Yao, Y.; Zhang, R.; Claret, F.X.; Korc, M.; Sabapathy, K.; Menter, D.G.; Abbruzzese, J.L.; Reddy, S.A.G. Activator protein−1 has an essential role in pancreatic cancer cells and is regulated by a novel Akt−mediated mechanism. Mol. Cancer Res. 2009, 7, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Tempero, M.A.; Malafa, M.P.; Al−Hawary, M.; Behrman, S.W.; Benson, A.B.; Cardin, D.B.; Chiorean, E.G.; Chung, V.; Czito, B.; Del Chiaro, M.; et al. Pancreatic Adenocarcinoma, Version 2.2021, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2021, 19, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D.; Gilmore, T.D. Good cop, bad cop: The different faces of NF−kappaB. Cell. Death Differ. 2006, 13, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Dajee, M.; Lazarov, M.; Zhang, J.Y.; Cai, T.; Green, C.L.; Russell, A.J.; Marinkovich, M.P.; Tao, S.; Lin, Q.; Kubo, Y.; et al. NF−kappaB blockade and oncogenic Ras trigger invasive human epidermal neoplasia. Nature 2003, 421, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Luedde, T.; Beraza, N.; Kotsikoris, V.; van Loo, G.; Nenci, A.; De Vos, R.; Roskams, T.; Trautwein, C.; Pasparakis, M. Deletion of NEMO/IKKgamma in liver parenchymal cells causes steatohepatitis and hepatocellular carcinoma. Cancer Cell. 2007, 11, 119–132. [Google Scholar] [CrossRef]
- Lee, B.L.; Lee, H.S.; Jung, J.; Cho, S.J.; Chung, H.Y.; Kim, W.H.; Jin, Y.W.; Kim, C.S.; Nam, S.Y. Nuclear factor−kappaB activation correlates with better prognosis and Akt activation in human gastric cancer. Clin. Cancer Res. 2005, 11, 2518–2525. [Google Scholar] [CrossRef]
- Jara−Gutierrez, A.; Baladron, V. The Role of Prostaglandins in Different Types of Cancer. Cells 2021, 10, 1487. [Google Scholar] [CrossRef]
- Dragovich, T.; Burris, H., 3rd; Loehrer, P.; Von Hoff, D.D.; Chow, S.; Stratton, S.; Green, S.; Obregon, Y.; Alvarez, I.; Gordon, M. Gemcitabine plus celecoxib in patients with advanced or metastatic pancreatic adenocarcinoma: Results of a phase II trial. Am. J. Clin. Oncol. 2008, 31, 157–162. [Google Scholar] [CrossRef]
- El−Rayes, B.F.; Zalupski, M.M.; Shields, A.F.; Ferris, A.M.; Vaishampayan, U.; Heilbrun, L.K.; Venkatramanamoorthy, R.; Adsay, V.; Philip, P.A. A phase II study of celecoxib, gemcitabine, and cisplatin in advanced pancreatic cancer. Investig. New Drugs 2005, 23, 583–590. [Google Scholar] [CrossRef]
- Chuang, J.Y.; Wu, C.H.; Lai, M.D.; Chang, W.C.; Hung, J.J. Overexpression of Sp1 leads to p53−dependent apoptosis in cancer cells. Int. J. Cancer 2009, 125, 2066–2076. [Google Scholar] [CrossRef]
- Omura, N.; Griffith, M.; Vincent, A.; Li, A.; Hong, S.M.; Walter, K.; Borges, M.; Goggins, M. Cyclooxygenase−deficient pancreatic cancer cells use exogenous sources of prostaglandins. Mol. Cancer Res. 2010, 8, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Treiber, M.; Neuhofer, P.; Anetsberger, E.; Einwachter, H.; Lesina, M.; Rickmann, M.; Liang, S.; Kehl, T.; Nakhai, H.; Schmid, R.M.; et al. Myeloid, but not pancreatic, RelA/p65 is required for fibrosis in a mouse model of chronic pancreatitis. Gastroenterology 2011, 141, 1473–1485.e7. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Primavesi, F.; Stattner, S.; Schlick, K.; Kiesslich, T.; Mayr, C.; Klieser, E.; Urbas, R.; Neureiter, D. Pancreatic cancer in young adults: Changes, challenges, and solutions. Onco Targets Ther. 2019, 12, 3387–3400. [Google Scholar] [CrossRef] [PubMed]
- Wahyudi, D.; Pratiw, S.E. Incidence of pancreatic cancer cases in Dr. Soedarso Hospital Pontianak. IJOC 2021, 15, 6. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, J.; Du, J.; Xu, H.; Zhang, W.; Ni, Q.X.; Yu, H.; Risch, H.A.; Gao, Y.T.; Gao, Y. Urinary prostaglandin E2 metabolite and pancreatic cancer risk: Case−control study in urban Shanghai. PLoS ONE 2015, 10, e0118004. [Google Scholar] [CrossRef]
- Cui, Y.; Shu, X.O.; Li, H.L.; Yang, G.; Wen, W.; Gao, Y.T.; Cai, Q.; Rothman, N.; Yin, H.Y.; Lan, Q.; et al. Prospective study of urinary prostaglandin E2 metabolite and pancreatic cancer risk. Int. J. Cancer 2017, 141, 2423–2429. [Google Scholar] [CrossRef]
- Antonangeli, F.; Natalini, A.; Garassino, M.C.; Sica, A.; Santoni, A.; Di Rosa, F. Regulation of PD−L1 Expression by NF−κB in Cancer. Front. Immunol. 2020, 11, 584626. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Categories | Number (%) |
|---|---|---|
| Mean age in years (standard deviation) | 52.4 (SD 10.9) | |
| Gender | Male | 33 (62.3) |
| Female | 20 (37.7) | |
| Nuclear grade | I | 22 (41.5) |
| II | 16 (30.2) | |
| III | 15 (28.3) | |
| Lymphovascular invasion | Yes | 6 (11.3) |
| No | 47 (88.7) | |
| Perineural invasion | Yes | 15 (28.3) |
| No | 38 (71.7) | |
| Specimen | Resection | 33 (62.3) |
| Biopsy | 20 (37.7) |
| Factors | Total | Univariate Analysis | ||
|---|---|---|---|---|
| HR | 95% CI | p−Value | ||
| Gender | ||||
| Male | 19 | 1.72 | 0.74–4.03 | 0.210 |
| Female | 15 | 1 | ||
| Age | ||||
| ≥60 | 11 | 0.85 | 0.35–2.09 | 0.731 |
| <60 | 23 | 1 | ||
| Grade | ||||
| >1 | 20 | 1.55 | 0.66–3.65 | 0.311 |
| 1 | 14 | 1 | ||
| Perineural invasion | ||||
| Present | 11 | 1.02 | 0.42–2.50 | 0.959 |
| Not Present | 23 | 1 | ||
| Lymphovascular invasion | ||||
| Present | 6 | 0.50 | 0.15–1.70 | 0.268 |
| Not Present | 28 | 1 | ||
| Type of specimen | ||||
| Biopsy | 13 | 1.49 | 0.65–3.41 | 0.341 |
| Resection | 21 | 1 | ||
| Cancer stage | ||||
| III–IV | 24 | 1.82 | 0.60–5.55 | 0.289 |
| I–II | 10 | 1 | ||
| NF−κB (RelA) cytoplasmic status | ||||
| Positive | 14 | 0.37 | 0.14–1.03 | 0.058 |
| Negative | 20 | 1 | ||
| NF−κB (RelA) nuclear status | ||||
| Positive | 11 | 0.29 | 0.10–0.84 | 0.022 * |
| Negative | 23 | 1 | ||
| COX−2 status | ||||
| Positive | 28 | 0.67 | 0.19–2.35 | 0.531 |
| Negative | 6 | 1 | ||
| Sp1 status | ||||
| Positive | 19 | 0.91 | 0.39–2.12 | 0.830 |
| Negative | 15 | 1 | ||
| c−Jun status | ||||
| Positive | 15 | 0.90 | 0.39–2.06 | 0.799 |
| Negative | 19 | 1 | ||
| Co−expressions of NF−κB (nuclear), COX−2, Sp1, c−Jun | ||||
| Yes | 6 | 0.43 | 0.14–1.29 | 0.133 |
| No | 28 | 1 | ||
| Gender | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renaldi, K.; Simadibrata, M.; Rahadiani, N.; Handjari, D.R.; Harahap, A.R.; Harimurti, K.; Zubir, N.; Siregar, L.; Loho, I.M.; Suzanna, E.; et al. The Expressions of NF−κB, COX−2, Sp1, and c−Jun in Pancreatic Ductal Adenocarcinoma and Their Associations with Patient Survival. Pathophysiology 2023, 30, 92-109. https://doi.org/10.3390/pathophysiology30020009
Renaldi K, Simadibrata M, Rahadiani N, Handjari DR, Harahap AR, Harimurti K, Zubir N, Siregar L, Loho IM, Suzanna E, et al. The Expressions of NF−κB, COX−2, Sp1, and c−Jun in Pancreatic Ductal Adenocarcinoma and Their Associations with Patient Survival. Pathophysiology. 2023; 30(2):92-109. https://doi.org/10.3390/pathophysiology30020009
Chicago/Turabian StyleRenaldi, Kaka, Marcellus Simadibrata, Nur Rahadiani, Diah Rini Handjari, Alida Roswita Harahap, Kuntjoro Harimurti, Nasrul Zubir, Lianda Siregar, Imelda Maria Loho, Evlina Suzanna, and et al. 2023. "The Expressions of NF−κB, COX−2, Sp1, and c−Jun in Pancreatic Ductal Adenocarcinoma and Their Associations with Patient Survival" Pathophysiology 30, no. 2: 92-109. https://doi.org/10.3390/pathophysiology30020009
APA StyleRenaldi, K., Simadibrata, M., Rahadiani, N., Handjari, D. R., Harahap, A. R., Harimurti, K., Zubir, N., Siregar, L., Loho, I. M., Suzanna, E., Prawirodihardjo, B., Hidajat, H., Widodo, B., Rahniayu, A., Tambun, R., William, A., & Makmun, D. (2023). The Expressions of NF−κB, COX−2, Sp1, and c−Jun in Pancreatic Ductal Adenocarcinoma and Their Associations with Patient Survival. Pathophysiology, 30(2), 92-109. https://doi.org/10.3390/pathophysiology30020009