

Screening and Characterization of New Acetobacter fabarum and Acetobacter pasteurianus Strains with High Ethanol–Thermo Tolerance and the Optimization of Acetic Acid Production
 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Identification of Bacterial Strains
2.3. Morphological Biochemical and Metabolic Tests
2.4. Acetic Acid Production Capacity on GYC Medium
2.5. Tolerance Analysis of AAB Strains
2.6. Molecular Identification
3. Results
3.1. Screening and Chemicals Characterization of AAB
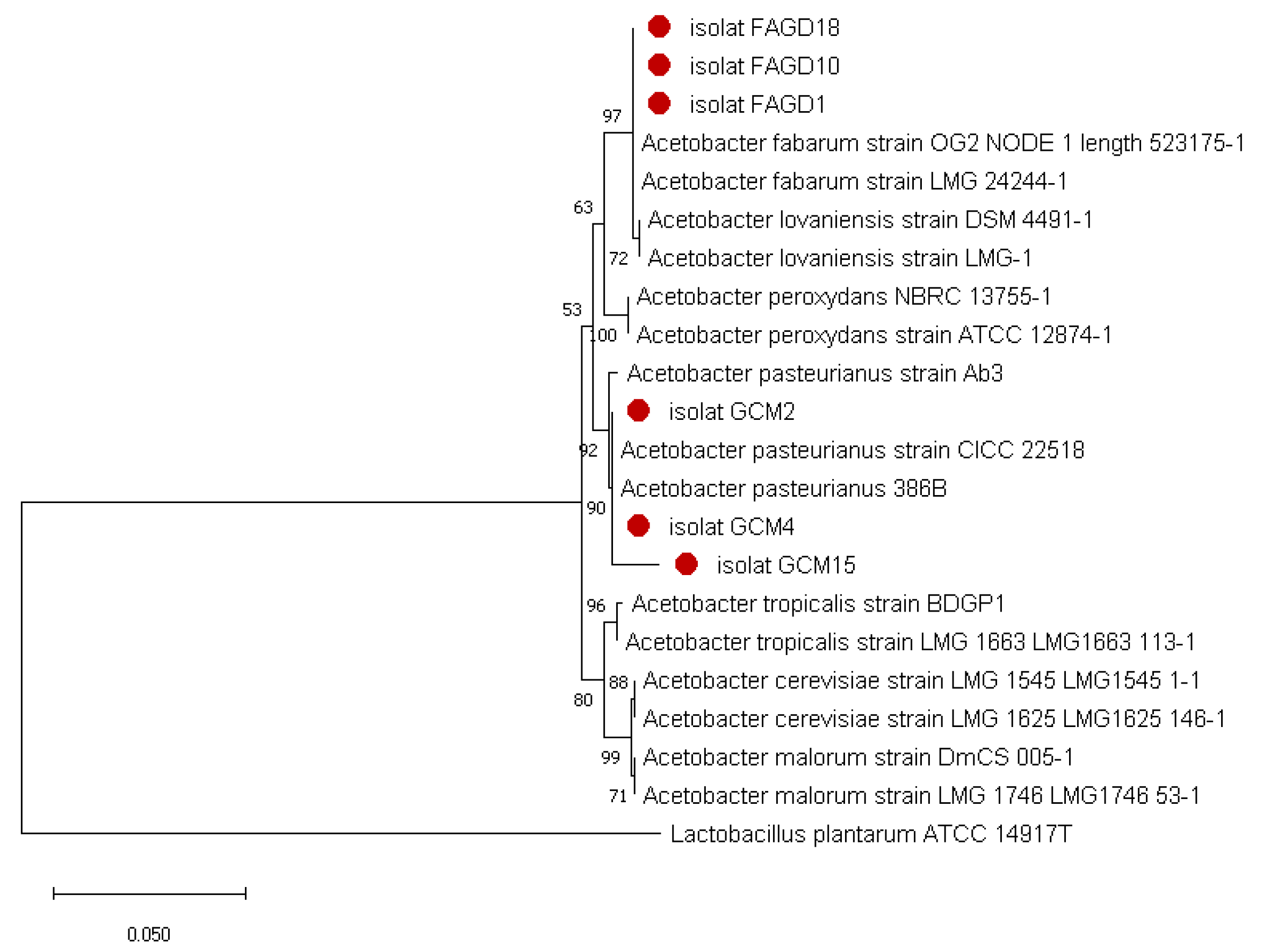
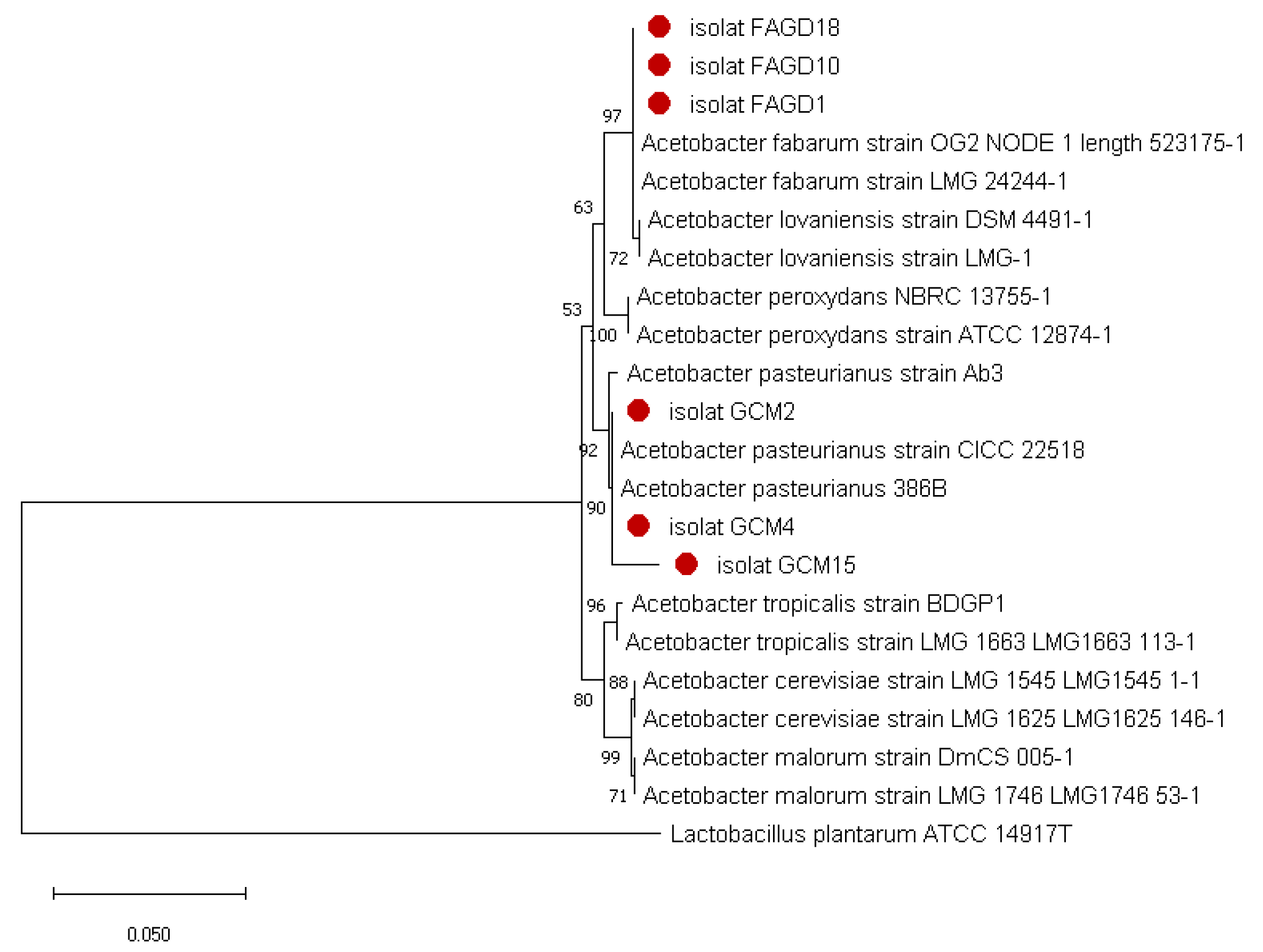
3.2. Molecular Identification of the Isolates by 16S rDNA Genes Sequencing
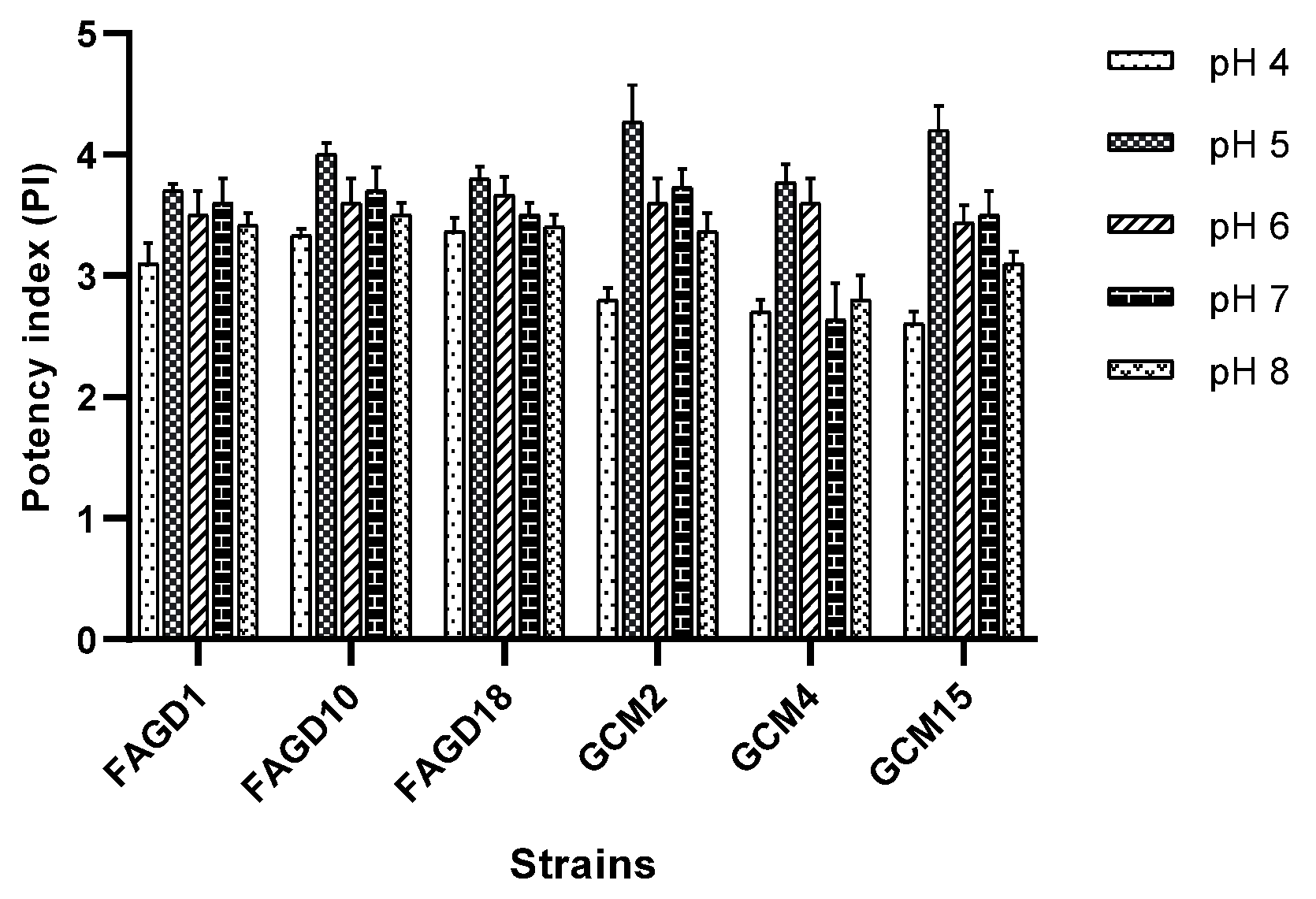
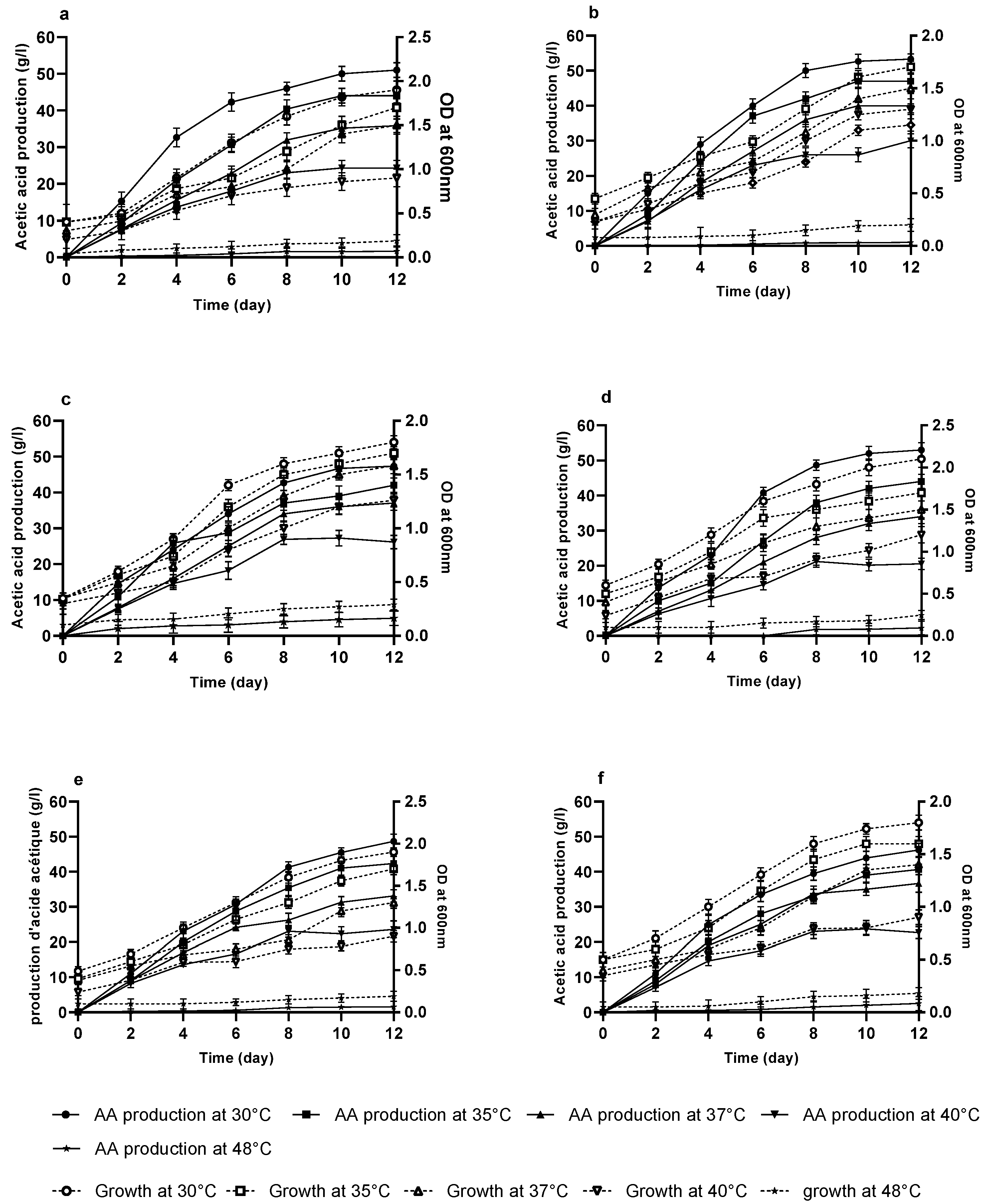
3.3. Effect of pH on Acetic Acid Production in Terms of Potency Index (PI)
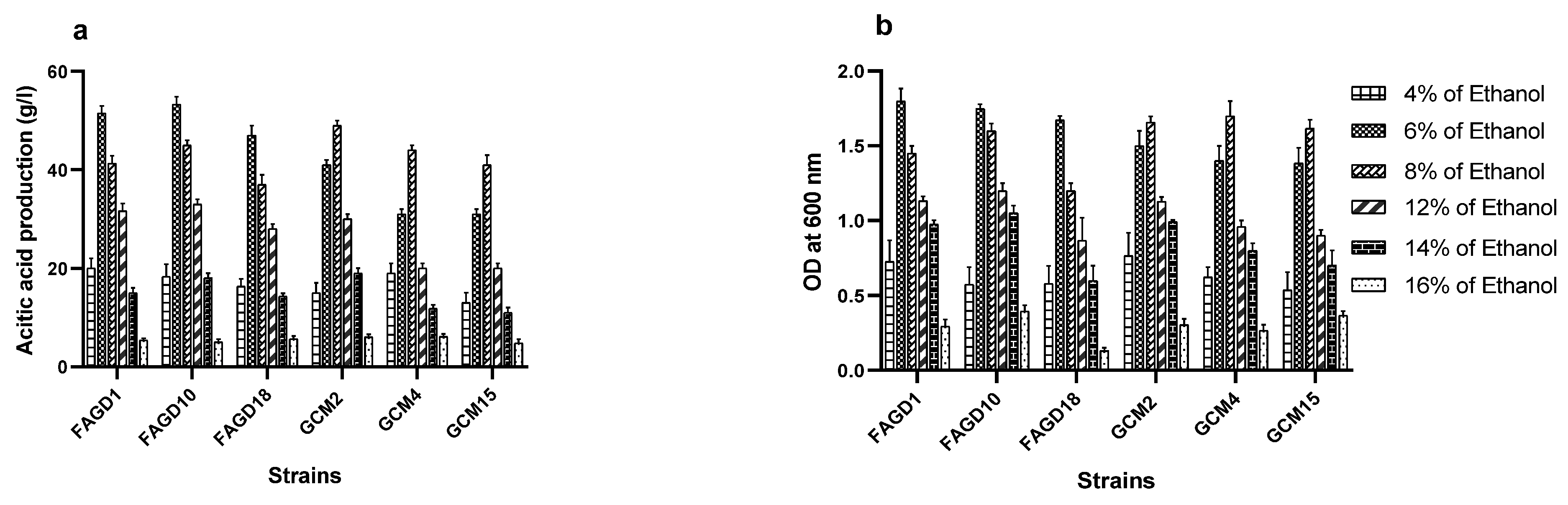
3.4. Ethanol Tolerance of AAB Strains
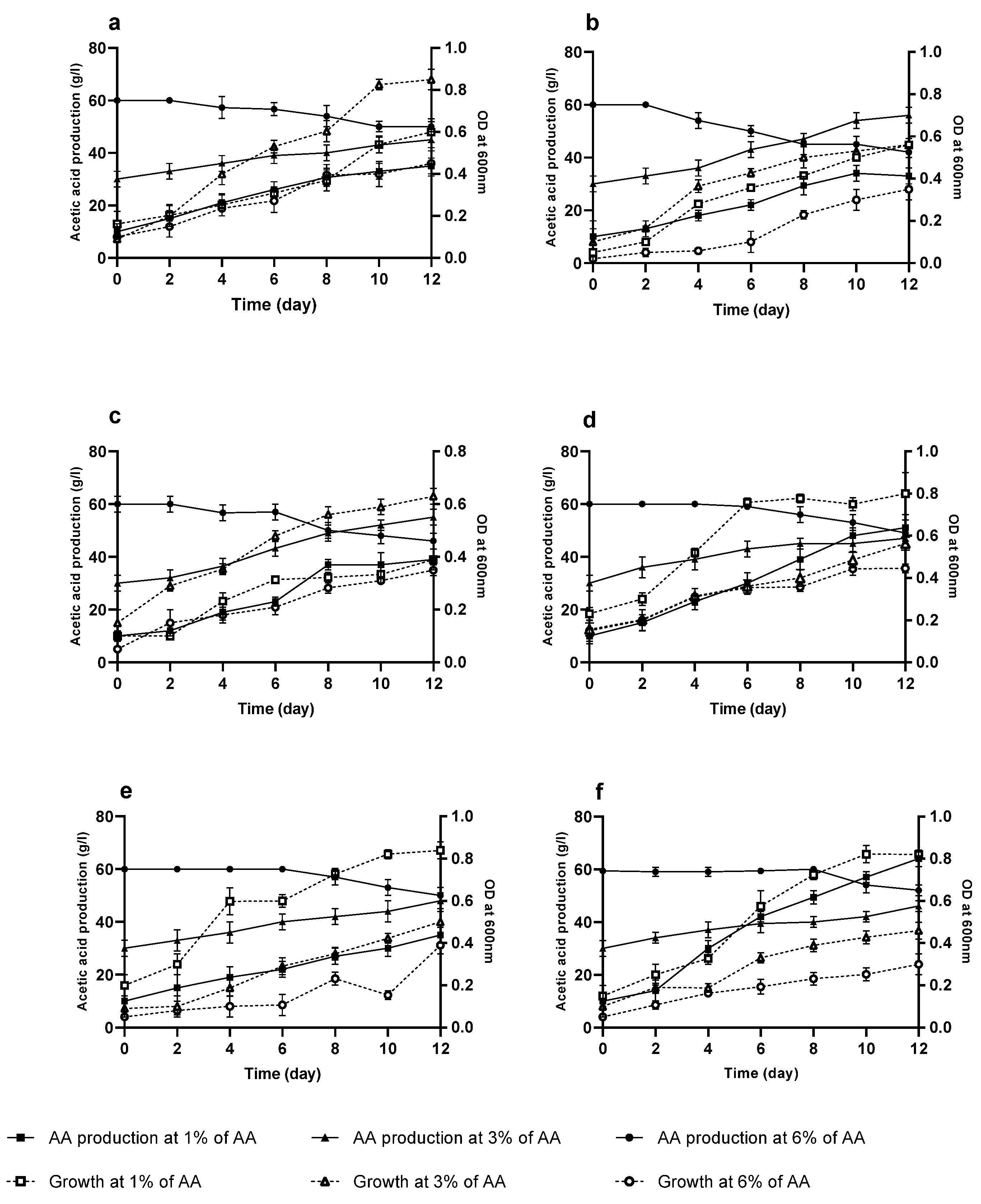
3.5. Acetic Acid Tolerance of AAB Strains
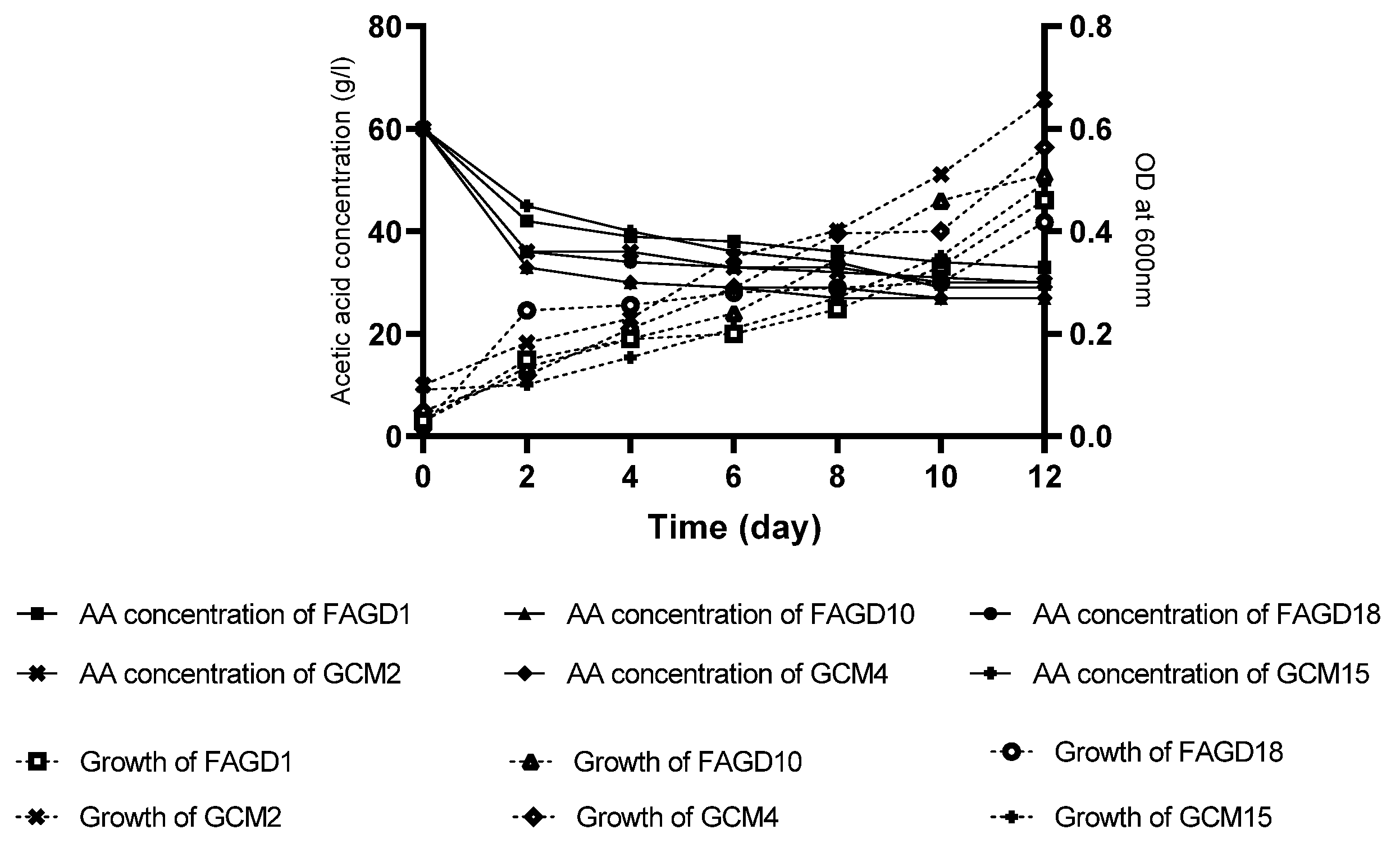
3.6. Growth and Production of Acetic Acid
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Merli, G.; Becci, A.; Amato, A.; Beolchini, F. Acetic acid bioproduction: The technological innovation change. Sci. Total Environ. 2021, 798, 149292. [Google Scholar] [CrossRef] [PubMed]
- De Roos, J.; De Vuyst, L. Acetic acid bacteria in fermented foods and beverages. Curr. Opin. Biotechnol. 2018, 49, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Vashisht, A.; Thakur, K.; Kauldhar, B.S.; Kumar, V.; Yadav, S.K. Waste valorization: Identification of an ethanol tolerant bacterium Acetobacter pasteurianus SKYAA25 for acetic acid production from apple pomace. Sci. Total Environ. 2019, 690, 956–964. [Google Scholar] [CrossRef]
- Pal, P.; Nayak, J. Acetic Acid Production and Purification: Critical Review Towards Process Intensification. Sep. Purif. Rev. 2017, 46, 44–61. [Google Scholar] [CrossRef]
- Mamlouk, D.; Gullo, M. Acetic Acid Bacteria: Physiology and Carbon Sources Oxidation. Indian J. Microbiol. 2013, 53, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Saichana, N.; Matsushita, K.; Adachi, O.; Frébort, I.; Frebortova, J. Acetic acid bacteria: A group of bacteria with versatile biotechnological applications revue. Biotechnol. Adv. 2015, 33, 1260–1271. [Google Scholar] [CrossRef]
- Gomes, R.J.; Borges, M.D.F.; Rosa, M.D.F.; Castro-Gómez, R.J.H.; Spinosa, W.A. Acetic acid bacteria in the food industry: Systematics, characteristics and applications. Food Technol. Biotechnol. 2018, 56, 139–151. [Google Scholar] [CrossRef]
- Trček, J.; Barja, F. Updates on quick identification of acetic acid bacteria with a focus on the 16S−23S rRNA gene internal transcribed spacer and the analysis of cell proteins by MALDI-TOF mass spectrometry. Int. J. Food Microbiol. 2014, 196, 137–144. [Google Scholar] [CrossRef]
- Qiu, X.; Zhang, Y.; Hong, H. Classification of acetic acid bacteria and their acid resistant mechanism. AMB Express 2021, 11, 29. [Google Scholar] [CrossRef]
- Fernández-Pérez, R.; Torres, C.; Sanz, S.; Ruiz-Larrea, F. Rapid molecular methods for enumeration and taxonomical identification of acetic acid bacteria responsible for submerged vinegar production. Eur. Food Res. Technol. 2010, 231, 813–819. [Google Scholar] [CrossRef]
- Andrés-Barrao, C.; Benagli, C.; Chappuis, M.; Ortega Pérez, R.; Tonolla, M.; Barja, F. Rapid identification of acetic acid bacteria using MALDI-TOF mass spectrometry fingerprinting. Syst. Appl. Microbiol. 2013, 36, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Yetiman, A.E.; Kesmen, Z. Identification of acetic acid bacteria in traditionally produced vinegar and mother of vinegar by using different molecular techniques. Int. J. Food Microbiol. 2015, 204, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Feng, F.; Chen, L.; Yao, Q.; Chen, K. Directional isolation of ethanol-tolerant acetic acid bacteria from industrial fermented vinegar. Eur. Food Res. Technol. 2013, 236, 573–578. [Google Scholar] [CrossRef]
- Ndoye, B.; Lebecque, S.; Dubois-Dauphin, R.; Tounkara, L.; Guiro, A.T.; Kere, C.; Diawara, B.; Thonart, P. Thermoresistant properties of acetic acids bacteria isolated from tropical products of Sub-Saharan Africa and destined to industrial vinegar. Enzym. Microb. Technol. 2006, 39, 916–923. [Google Scholar] [CrossRef]
- Raspor, P.; Goranovič, D. Biotechnological applications of acetic acid bacteria. Crit. Rev. Biotechnol. 2008, 28, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Bai, Y.; Li, D.; Wang, C.; Xu, N.; Hu, Y. Screening and characterization of ethanol-tolerant and thermotolerant acetic acid bacteria from Chinese vinegar Pei. World J. Microbiol. Biotechnol. 2016, 32, 14. [Google Scholar] [CrossRef]
- Moryadee, A.; Pathon-Aree, W. Isolation of Thermotolerant Acetic Acid Bacteria from Fruits for Vinegar Production. Res. J. Microbiol. 2008, 3, 209–212. [Google Scholar] [CrossRef]
- Pothimon, R.; Gullo, M.; La China, S.; Thompson, A.K.; Krusong, W. Conducting High acetic acid and temperature acetification processes by Acetobacter pasteurianus UMCC 2951. Process Biochem. 2020, 98, 41–50. [Google Scholar] [CrossRef]
- Kanchanarach, W.; Theeragool, G.; Inoue, T.; Yakushi, T.; Adachi, O.; Matsushita, K. Acetic acid fermentation of acetobacter pasteurianus: Relationship between acetic acid resistance and pellicle polysaccharide formation. Biosci. Biotechnol. Biochem. 2010, 74, 1591–1597. [Google Scholar] [CrossRef]
- Perumpuli, P.A.B.N.; Watanabe, T.; Toyama, H. Identification and characterization of thermotolerant acetic acid bacteria strains isolated from coconut water vinegar in Sri Lanka. Biosci. Biotechnol. Biochem. 2014, 78, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Dartnell, L.R.; Fallaize, D.R.; Whiting, S.J.; Ward, J.M. Desiccation resistance of Antarctic Dry Valley bacteria isolated from contrasting locations. Antarct. Sci. 2010, 22, 171–172. [Google Scholar] [CrossRef]
- Konate, M.; Akpa, E.E.; Koffi, L.B.; Kra, K.A.S.; Megnanou, R.M.; Niamke, S. Isolation of thermotolerant and high acetic acid-producing Acetobacter pasteurianus from Ivorian palm wine. Emir. J. Food Agric. 2014, 26, 773–785. [Google Scholar] [CrossRef]
- Soumahoro, S.; Goualie, B.G.; Adom, J.N.; Ouattara, H.G.; Koua, G.; Doue, G.G.; Niamke, S.L. Effects of culture conditions on acetic acid production by bacteria isolated from Ivoirian fermenting cocoa (Theobroma cacao L.) beans. J. Appl. Biosci. 2016, 95, 8981. [Google Scholar] [CrossRef]
- Matsushita, K.; Azuma, Y.; Kosaka, T.; Yakushi, T.; Hoshida, H.; Akada, R.; Yamada, M. Genomic analyses of thermotolerant microorganisms used for high-temperature fermentations. Biosci. Biotechnol. Biochem. 2016, 80, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Osumi, N.; Matsutani, M.; Phathanathavorn, T.; Kataoka, N.; Theeragool, G.; Yakushi, T.; Shiraishi, Y.; Matsushita, K. Thermal adaptation of acetic acid bacteria for practical high-temperature vinegar fermentation. Biosci. Biotechnol. Biochem. 2021, 85, 1243–1251. [Google Scholar] [CrossRef]
- Mounir, M.; Shafiei, R.; Zarmehrkhorshid, R.; Hamouda, A.; Ismaili Alaoui, M.; Thonart, P. Simultaneous production of acetic and gluconic acids by a thermotolerant Acetobacter strain during acetous fermentation in a bioreactor. J. Biosci. Bioeng. 2016, 121, 166–171. [Google Scholar] [CrossRef]
- Es-sbata, I.; Lakhlifi, T.; Yatim, M.; El-Abid, H.; Belhaj, A.; Hafidi, M.; Zouhair, R. Screening and molecular characterization of new thermo- and ethanol-tolerant Acetobacter malorum strains isolated from two biomes Moroccan cactus fruits. Biotechnol. Appl. Biochem. 2021, 68, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Es-sbata, I.; Castro, R.; Durán-Guerrero, E.; Zouhair, R.; Astola, A. Production of prickly pear (Opuntia ficus-indica) vinegar in submerged culture using Acetobacter malorum and Gluconobacter oxydans: Study of volatile and polyphenolic composition. J. Food Compos. Anal. 2022, 112, 104699. [Google Scholar] [CrossRef]
- Du Toit, W.J.; Pretorius, I.S. The occurrence, control and esoteric effect of acetic acid bacteria in winemaking. Ann. Microbiol. 2002, 52, 155–179. [Google Scholar]
- Trček, J.; Mira, N.P.; Jarboe, L.R. Adaptation and tolerance of bacteria against acetic acid. Appl. Microbiol. Biotechnol. 2015, 99, 6215–6229. [Google Scholar] [CrossRef]
- Gao, L.; Wu, X.; Zhu, C.; Jin, Z.; Wang, W.; Xia, X. Metabolic engineering to improve the biomanufacturing efficiency of acetic acid bacteria: Advances and prospects. Crit. Rev. Biotechnol. 2020, 40, 522–538. [Google Scholar] [CrossRef] [PubMed]
- Kourouma, M.C.; Mbengue, M.; Sarr, N.C.D.; Sarr, K.; Kane, C.T. Thermoresistant, Ethanol-Resistant and Acid-Resistant Properties of Acetic Acid Bacteria Isolated from Fermented Mango Alcohol. Adv. Microbiol. 2022, 12, 177–191. [Google Scholar] [CrossRef]
- Kadere, T.T.; Miyamoto, T.; Oniang, R.K.; Kutima, P.M.; Njoroge, S.M. Isolation and identification of the genera Acetobacter and Gluconobacter in coconut toddy (mnazi). Afr. J. Biotechnol. 2008, 7, 2963–2971. [Google Scholar]
- Yamada, Y.; Hosono, R.; Lisdyanti, P.; Widyastuti, Y.; Saono, S.; Uchimura, T.; Komagata, K. Identification of acetic acid bacteria isolated from Indonesian sources, especially of isolates classified in the genus Gluconobacter. J. Gen. Appl. Microbiol. 1999, 45, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Arifuzzaman, M.; Hasan, M.Z.; Rahman, S.M.B.; Pramanik, M.K. Isolation and characterization of Acetobacter and Gluconobacter spp from sugarcane and rotten fruits. Res. Rev. Biosci. 2014, 8, 359–365. [Google Scholar]
- Kappeng, K.; Pathom-Aree, W. Isolation of acetic acid bacteria from honey. Maejo Int. J. Sci. Technol. 2009, 3, 71–76. [Google Scholar]
- Romero-Cortes, T. Isolation and characterization of acetic acid bacteria in cocoa fermentation. Afr. J. Microbiol. Res. 2012, 6, 339–347. [Google Scholar] [CrossRef]
- Yanti, N.A.; Ahmad, S.W.; Ambardini, S.; Muhiddin, N.H.; Sulaiman, L.O.I. Screening of Acetic Acid Bacteria from Pineapple Waste for Bacterial Cellulose Production using Sago Liquid Waste. Biosaintifika J. Biol. Biol. Educ. 2017, 9, 387. [Google Scholar] [CrossRef]
- Mathew, B.; Agrawal, S.; Nashikkar, N.; Bundale, S.; Upadhyay, A. Isolation of Acetic Acid Bacteria and Preparation of Starter Culture for Apple Cider Vinegar Fermentation. Adv. Microbiol. 2019, 9, 556–569. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, R.; Yin, H.; Bai, X.; Chang, Y.; Xia, M.; Wang, M. Acetobacter pasteurianus metabolic change induced by initial acetic acid to adapt to acetic acid fermentation conditions. Appl. Microbiol. Biotechnol. 2017, 101, 7007–7016. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Cepec, E.; Trček, J. Antimicrobial resistance of acetobacter and komagataeibacter species originating from vinegars. Int. J. Environ. Res. Public Health 2022, 19, 463. [Google Scholar] [CrossRef] [PubMed]
- Beheshti-Maal, K.; Shafiee, N. Isolation and Identification of A Novel Strain of Acetobacter ghanensis KBMNS-IAUF-6 from Banana Fruit, Resistant to High Temperature and Ethanol Concentration. Iran. J. Med. Microbiol. 2019, 13, 251–265. [Google Scholar] [CrossRef]
- Wang, B.; Shao, Y.; Chen, F. Overview on mechanisms of acetic acid resistance in acetic acid bacteria. World J. Microbiol. Biotechnol. 2015, 31, 255–263. [Google Scholar] [CrossRef]
- Nakano, S.; Fukaya, M. Analysis of proteins responsive to acetic acid in Acetobacter: Molecular mechanisms conferring acetic acid resistance in acetic acid bacteria. Int. J. Food Microbiol. 2008, 125, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Fukaya, M.; Horinouchi, S. Enhanced expression of aconitase raises acetic acid resistance in Acetobacter aceti. FEMS Microbiol. Lett. 2004, 235, 315–322. [Google Scholar] [CrossRef]
- Matsumoto, N.; Matsutani, M.; Azuma, Y.; Kataoka, N.; Yakushi, T.; Matsushita, K. In vitro thermal adaptation of mesophilic Acetobacter pasteurianus NBRC 3283 generates thermotolerant strains with evolutionary trade-offs. Biosci. Biotechnol. Biochem. 2020, 84, 832–841. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Isolates | |||||||
|---|---|---|---|---|---|---|---|
| FAGD 1 | FAGD 10 | FAGD 18 | GCM 2 | GCM 4 | GCM 15 | ||
| Morphological characteristics | Shape | Rod | Rod | Rod | Rod | Rod | Rod |
| Gram’s stain | − | − | − | − | − | − | |
| Spore | − | − | − | − | − | − | |
| Motility | + | + | + | − | − | − | |
| Biochemical characteristics | Catalase | + | + | + | + | + | + |
| Oxidase | − | − | − | − | − | − | |
| Production of acetic acid from ethanol | + | + | + | + | + | + | |
| Over oxidation of ethanol to CO2 and H2O | + | + | + | + | + | + | |
| Brown pigmentation | − | − | − | − | − | − | |
| Growth in peptone | + | + | + | + | + | + | |
| D-glucose concentration > 30% | − | − | − | + | + | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Askri, T.; Yatim, M.; Sehli, Y.; Rahou, A.; Belhaj, A.; Castro, R.; Durán-Guerrero, E.; Hafidi, M.; Zouhair, R. Screening and Characterization of New Acetobacter fabarum and Acetobacter pasteurianus Strains with High Ethanol–Thermo Tolerance and the Optimization of Acetic Acid Production. Microorganisms 2022, 10, 1741. https://doi.org/10.3390/microorganisms10091741
El-Askri T, Yatim M, Sehli Y, Rahou A, Belhaj A, Castro R, Durán-Guerrero E, Hafidi M, Zouhair R. Screening and Characterization of New Acetobacter fabarum and Acetobacter pasteurianus Strains with High Ethanol–Thermo Tolerance and the Optimization of Acetic Acid Production. Microorganisms. 2022; 10(9):1741. https://doi.org/10.3390/microorganisms10091741
Chicago/Turabian StyleEl-Askri, Taoufik, Meriem Yatim, Youness Sehli, Abdelilah Rahou, Abdelhaq Belhaj, Remedios Castro, Enrique Durán-Guerrero, Majida Hafidi, and Rachid Zouhair. 2022. "Screening and Characterization of New Acetobacter fabarum and Acetobacter pasteurianus Strains with High Ethanol–Thermo Tolerance and the Optimization of Acetic Acid Production" Microorganisms 10, no. 9: 1741. https://doi.org/10.3390/microorganisms10091741
APA StyleEl-Askri, T., Yatim, M., Sehli, Y., Rahou, A., Belhaj, A., Castro, R., Durán-Guerrero, E., Hafidi, M., & Zouhair, R. (2022). Screening and Characterization of New Acetobacter fabarum and Acetobacter pasteurianus Strains with High Ethanol–Thermo Tolerance and the Optimization of Acetic Acid Production. Microorganisms, 10(9), 1741. https://doi.org/10.3390/microorganisms10091741