The Sex Differences in Uveal Melanoma: Potential Roles of EIF1AX, Immune Response and Redox Regulation

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. The Sex Difference Analyzed by Epidemiological Methods
3.1.1. The Age-Dependent Sex Disparity in Uveal Melanoma from the SEER Dataset
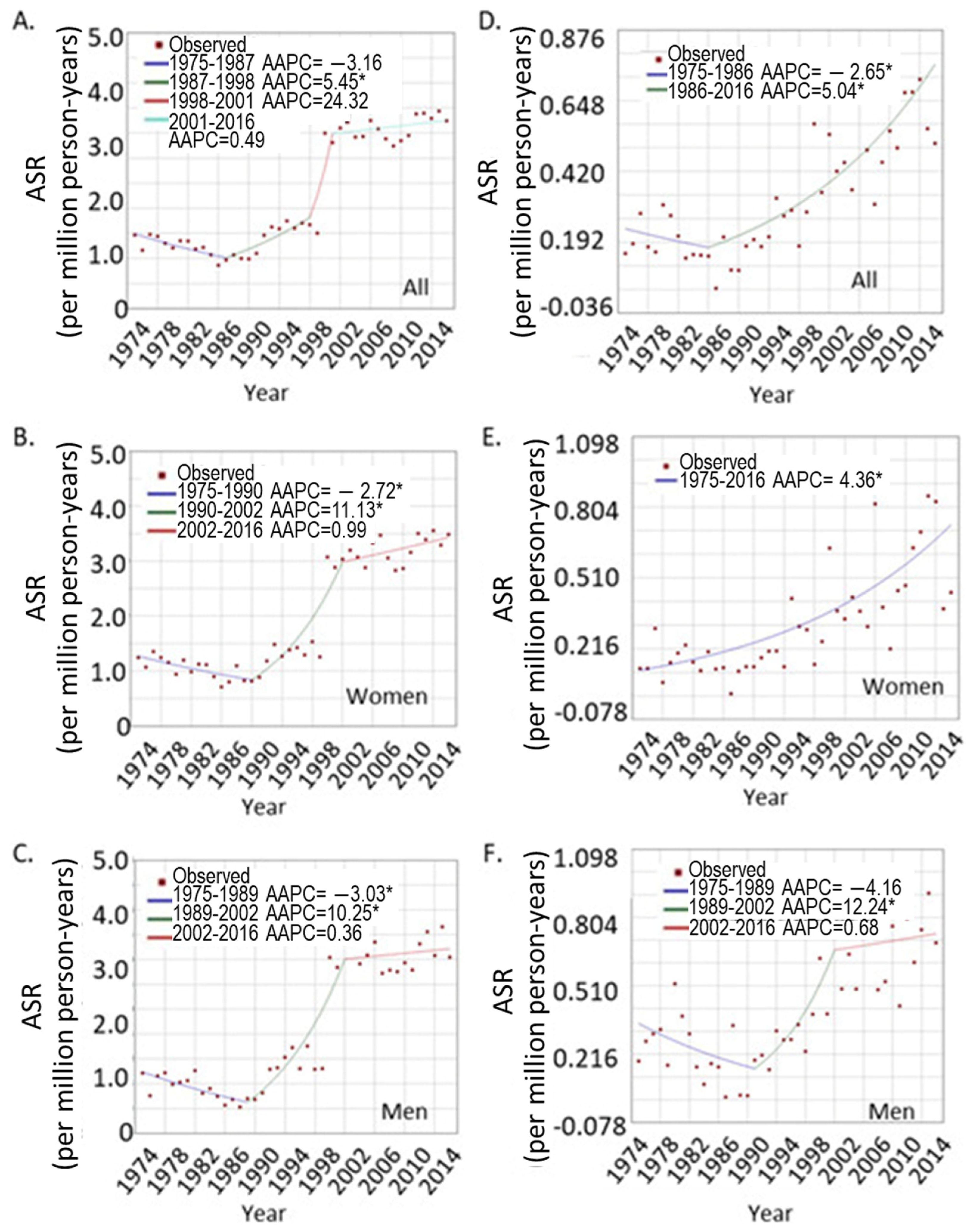
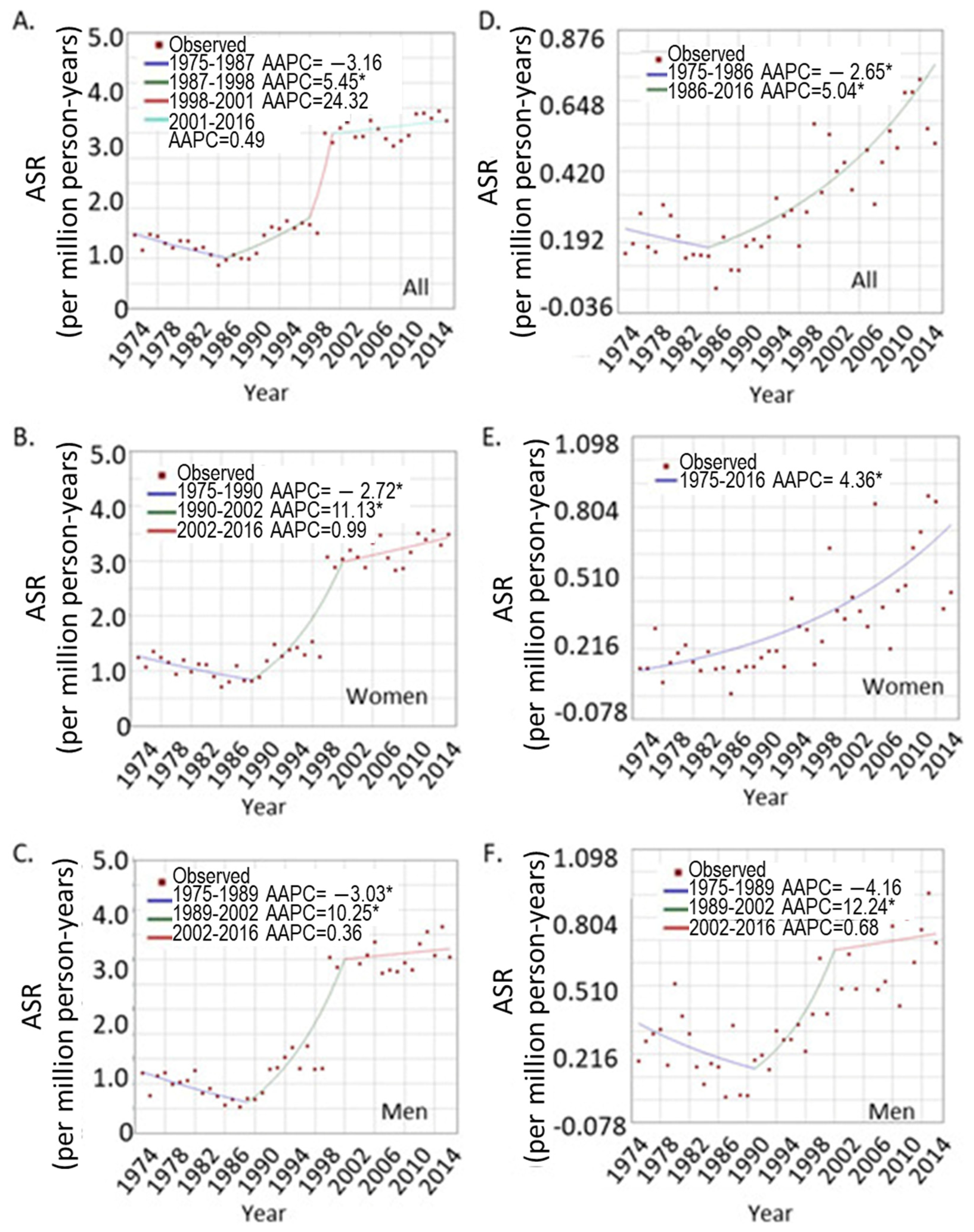
3.1.2. The Trend of UVM Incidence Rates over Years
3.1.3. The UVM Incidence Trend in Different Age Groups
3.2. The Sex Difference in Tumor Genomic Analysis
3.2.1. The Sex Difference in Major Oncogenes from the TCGA UVM Patients: Higher EIF1AX Expression in Female Tumors
3.2.2. The Global Sex-Differentiated Gene Expression Profile in UVM Tumors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaliki, S.; Shields, C.L. Uveal melanoma: Relatively rare but deadly cancer. Eye 2017, 31, 241–257. [Google Scholar] [CrossRef] [Green Version]
- Quintella Mendes, G.L.; Koifman, S. Socioeconomic status as a predictor of melanoma survival in a series of 1083 cases from Brazil: Just a marker of health services accessibility? Melanoma Res. 2013, 23, 199–205. [Google Scholar] [CrossRef]
- Lee, J.A.; Strickland, D. Malignant melanoma: Social status and outdoor work. Br. J. Cancer 1980, 41, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Shildkrot, Y.; Thomas, F.; Al-Hariri, A.; Fry, C.L.; Haik, B.G.; Wilson, M.W. Socioeconomic factors and diagnosis of uveal melanoma in the mid-southern United States. Curr. Eye Res. 2011, 36, 824–830. [Google Scholar] [CrossRef]
- Singh, A.D.; Topham, A. Incidence of uveal melanoma in the United States: 1973–1997. Ophthalmology 2003, 110, 956–961. [Google Scholar] [CrossRef]
- Krantz, B.A.; Dave, N.; Komatsubara, K.M.; Marr, B.P.; Carvajal, R.D. Uveal melanoma: Epidemiology, etiology, and treatment of primary disease. Clin. Ophthalmol. 2017, 11, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, K.M.; Seddon, J.M.; Glynn, R.J.; Gragoudas, E.S.; Albert, D.M. Epidemiologic aspects of uveal melanoma. Surv. Ophthalmol. 1988, 32, 239–251. [Google Scholar] [CrossRef]
- Liu-Smith, F.; Farhat, A.M.; Arce, A.; Ziogas, A.; Taylor, T.; Wang, Z.; Yourk, V.; Liu, J.; Wu, J.; McEligot, A.J.; et al. Sex differences in the association of cutaneous melanoma incidence rates and geographic ultraviolet light exposure. J. Am. Acad. Dermatol. 2017, 76, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu-Smith, F.; Ziogas, A. Age-dependent interaction between sex and geographic ultraviolet index in melanoma risk. J. Am. Acad. Dermatol. 2020, 82, 1102–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foss, A.J.; Alexander, R.A.; Guille, M.J.; Hungerford, J.L.; McCartney, A.C.; Lightman, S. Estrogen and progesterone receptor analysis in ocular melanomas. Ophthalmology 1995, 102, 431–435. [Google Scholar] [CrossRef]
- Collier, V.; Musicante, M.; Patel, T.; Liu-Smith, F. Sex disparity in skin carcinogenesis and potential influence of sex hormones. Ski. Health Dis. 2021. [Google Scholar] [CrossRef]
- Decatur, C.L.; Ong, E.; Garg, N.; Anbunathan, H.; Bowcock, A.M.; Field, M.G.; Harbour, J.W. Driver Mutations in Uveal Melanoma: Associations with Gene Expression Profile and Patient Outcomes. JAMA Ophthalmol. 2016, 134, 728–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoomian, B.; Shields, J.A.; Shields, C.L. Overview of BAP1 cancer predisposition syndrome and the relationship to uveal melanoma. J. Curr. Ophthalmol. 2018, 30, 102–109. [Google Scholar] [CrossRef]
- Liu-Smith, F.; Lu, Y. Opposite Roles of BAP1 in Overall Survival of Uveal Melanoma and Cutaneous Melanoma. J. Clin. Med. 2020, 9, 411. [Google Scholar] [CrossRef] [Green Version]
- Kalirai, H.; Dodson, A.; Faqir, S.; Damato, B.E.; Coupland, S.E. Lack of BAP1 protein expression in uveal melanoma is associated with increased metastatic risk and has utility in routine prognostic testing. Br. J. Cancer 2014, 111, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Chua, V.; Lapadula, D.; Randolph, C.; Benovic, J.L.; Wedegaertner, P.B.; Aplin, A.E. Dysregulated GPCR Signaling and Therapeutic Options in Uveal Melanoma. Mol. Cancer Res. 2017, 15, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A.G.; Shih, J.; Yau, C.; Gibb, E.A.; Oba, J.; Mungall, K.L.; Hess, J.M.; Uzunangelov, V.; Walter, V.; Danilova, L.; et al. Integrative Analysis Identifies Four Molecular and Clinical Subsets in Uveal Melanoma. Cancer Cell 2017, 32, 204–220 e215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Raamsdonk, C.D.; Griewank, K.G.; Crosby, M.B.; Garrido, M.C.; Vemula, S.; Wiesner, T.; Obenauf, A.C.; Wackernagel, W.; Green, G.; Bouvier, N.; et al. Mutations in GNA11 in uveal melanoma. N. Engl. J. Med. 2010, 363, 2191–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Raamsdonk, C.D.; Bezrookove, V.; Green, G.; Bauer, J.; Gaugler, L.; O’Brien, J.M.; Simpson, E.M.; Barsh, G.S.; Bastian, B.C. Frequent somatic mutations of GNAQ in uveal melanoma and blue naevi. Nature 2009, 457, 599–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandiani, C.; Beranger, G.E.; Leclerc, J.; Ballotti, R.; Bertolotto, C. Focus on cutaneous and uveal melanoma specificities. Genes Dev. 2017, 31, 724–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolatshad, H.; Pellagatti, A.; Fernandez-Mercado, M.; Yip, B.H.; Malcovati, L.; Attwood, M.; Przychodzen, B.; Sahgal, N.; Kanapin, A.A.; Lockstone, H.; et al. Disruption of SF3B1 results in deregulated expression and splicing of key genes and pathways in myelodysplastic syndrome hematopoietic stem and progenitor cells. Leukemia 2015, 29, 1092–1103. [Google Scholar] [CrossRef] [PubMed]
- Edmond, V.; Merdzhanova, G.; Gout, S.; Brambilla, E.; Gazzeri, S.; Eymin, B. A new function of the splicing factor SRSF2 in the control of E2F1-mediated cell cycle progression in neuroendocrine lung tumors. Cell Cycle 2013, 12, 1267–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lake, S.L.; Damato, B.E.; Kalirai, H.; Dodson, A.R.; Taktak, A.F.; Lloyd, B.H.; Coupland, S.E. Single nucleotide polymorphism array analysis of uveal melanomas reveals that amplification of CNKSR3 is correlated with improved patient survival. Am. J. Pathol. 2013, 182, 678–687. [Google Scholar] [CrossRef]
- Scholz, S.L.; Moller, I.; Reis, H.; Susskind, D.; van de Nes, J.A.P.; Leonardelli, S.; Schilling, B.; Livingstone, E.; Schimming, T.; Paschen, A.; et al. Frequent GNAQ, GNA11, and EIF1AX Mutations in Iris Melanoma. Investig. Ophthalmol. Vis. Sci. 2017, 58, 3464–3470. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Research, N. Integrated genomic characterization of papillary thyroid carcinoma. Cell 2014, 159, 676–690. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorthy, G.P.; Davidson, N.R.; Leach, S.D.; Zhao, Z.; Lowe, S.W.; Lee, G.; Landa, I.; Nagarajah, J.; Saqcena, M.; Singh, K.; et al. EIF1AX and RAS Mutations Cooperate to Drive Thyroid Tumorigenesis through ATF4 and c-MYC. Cancer Discov. 2019, 9, 264–281. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, Q.; Dang, K.; Ma, S.; Cotton, J.L.; Yang, S.; Zhu, L.J.; Deng, A.C.; Ip, Y.T.; Johnson, R.L.; et al. YAP/TAZ Activation Drives Uveal Melanoma Initiation and Progression. Cell Rep. 2019, 29, 3200–3211. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costin, G.E.; Valencia, J.C.; Wakamatsu, K.; Ito, S.; Solano, F.; Milac, A.L.; Vieira, W.D.; Yamaguchi, Y.; Rouzaud, F.; Petrescu, A.J.; et al. Mutations in dopachrome tautomerase (Dct) affect eumelanin/pheomelanin synthesis, but do not affect intracellular trafficking of the mutant protein. Biochem. J. 2005, 391, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronow, M.E.; Topham, A.K.; Singh, A.D. Uveal Melanoma: 5-Year Update on Incidence, Treatment, and Survival (SEER 1973-2013). Ocul. Oncol. Pathol. 2018, 4, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.I.; Lim, H.; Moon, A. Sex Differences in Cancer: Epidemiology, Genetics and Therapy. Biomol. Ther. 2018, 26, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Mitkov, M.; Joseph, R.; Copland, J., III. Steroid hormone influence on melanomagenesis. Mol. Cell. Endocrinol. 2015, 417, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Lv, N.; Liao, J.; Long, J.; Xue, R.; Ai, N.; Xu, D.; Fan, X. Copy number variation is highly correlated with differential gene expression: A pan-cancer study. BMC Med. Genet. 2019, 20, 175. [Google Scholar] [CrossRef]
- Karunamurthy, A.; Panebianco, F.; Hsiao, S.J.; Vorhauer, J.; Nikiforova, M.N.; Chiosea, S.; Nikiforov, Y.E. Prevalence and phenotypic correlations of EIF1AX mutations in thyroid nodules. Endocr. Relat. Cancer 2016, 23, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Jager, M.J.; Shields, C.L.; Cebulla, C.M.; Abdel-Rahman, M.H.; Grossniklaus, H.E.; Stern, M.H.; Carvajal, R.D.; Belfort, R.N.; Jia, R.; Shields, J.A.; et al. Uveal melanoma. Nat. Rev. Dis. Primers. 2020, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Rathod, K.S.; Kapil, V.; Velmurugan, S.; Khambata, R.S.; Siddique, U.; Khan, S.; Van Eijl, S.; Gee, L.C.; Bansal, J.; Pitrola, K.; et al. Accelerated resolution of inflammation underlies sex differences in inflammatory responses in humans. J. Clin. Investig. 2017, 127, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Seiler, M.P.; Mathew, R.; Liszewski, M.K.; Spooner, C.J.; Barr, K.; Meng, F.; Singh, H.; Bendelac, A. Elevated and sustained expression of the transcription factors Egr1 and Egr2 controls NKT lineage differentiation in response to TCR signaling. Nat. Immunol. 2012, 13, 264–271. [Google Scholar] [CrossRef]
- Galligan, J.J.; Petersen, D.R. The human protein disulfide isomerase gene family. Hum Genom. 2012, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Tenkorang, M.A.; Snyder, B.; Cunningham, R.L. Sex-related differences in oxidative stress and neurodegeneration. Steroids 2018, 133, 21–27. [Google Scholar] [CrossRef]
- Fujimoto, T.; Nakamura, O.; Saito, M.; Tsuru, A.; Matsumoto, M.; Kohno, K.; Inaba, K.; Kadokura, H. Identification of the physiological substrates of PDIp, a pancreas-specific protein-disulfide isomerase family member. J. Biol. Chem. 2018, 293, 18421–18433. [Google Scholar] [CrossRef] [Green Version]
- Turano, C.; Coppari, S.; Altieri, F.; Ferraro, A. Proteins of the PDI family: Unpredicted non-ER locations and functions. J. Cell. Physiol. 2002, 193, 154–163. [Google Scholar] [CrossRef]
- Desilva, M.G.; Notkins, A.L.; Lan, M.S. Molecular characterization of a pancreas-specific protein disulfide isomerase, PDIp. DNA Cell Biol. 1997, 16, 269–274. [Google Scholar] [CrossRef]
- Fu, X.M.; Zhu, B.T. Human pancreas-specific protein disulfide isomerase homolog (PDIp) is an intracellular estrogen-binding protein that modulates estrogen levels and actions in target cells. J. Steroid Biochem. Mol. Biol. 2009, 115, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Bellanti, F.; Matteo, M.; Rollo, T.; De Rosario, F.; Greco, P.; Vendemiale, G.; Serviddio, G. Sex hormones modulate circulating antioxidant enzymes: Impact of estrogen therapy. Redox Biol. 2013, 1, 340–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, A.K.; Soo, K.Y.; Levina, V.; Talbo, G.H.; Atkin, J.D. N-linked glycosylation modulates dimerization of protein disulfide isomerase family A member 2 (PDIA2). FEBS J. 2013, 280, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.N.; Simon, J.D.; Sarna, T. Role of ocular melanin in ophthalmic physiology and pathology. Photochem. Photobiol. 2008, 84, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Luger, T.A.; Scholzen, T.; Grabbe, S. The role of alpha-melanocyte-stimulating hormone in cutaneous biology. J. Investig. Dermatology. Symp. Proc. 1997, 2, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Donahoo, W.T.; Hernandez, T.L.; Costa, J.L.; Jensen, D.R.; Morris, A.M.; Brennan, M.B.; Hochgeschwender, U.; Eckel, R.H. Plasma alpha-melanocyte-stimulating hormone: Sex differences and correlations with obesity. Metabolism 2009, 58, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.K.; Funasaka, Y.; Slominski, A.; Ermak, G.; Hwang, J.; Pawelek, J.M.; Ichihashi, M. Production and release of proopiomelanocortin (POMC) derived peptides by human melanocytes and keratinocytes in culture: Regulation by ultraviolet B. Biochim. Biophys. Acta 1996, 1313, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Baron, M.; Maillet, J.; Huyvaert, M.; Dechaume, A.; Boutry, R.; Loiselle, H.; Durand, E.; Toussaint, B.; Vaillant, E.; Philippe, J.; et al. Loss-of-function mutations in MRAP2 are pathogenic in hyperphagic obesity with hyperglycemia and hypertension. Nat. Med. 2019, 25, 1733–1738. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Case Number | IR (Per Million) * | IRR | p Value | ||||
|---|---|---|---|---|---|---|---|
| Age Category | Female | Male | Total | Female | Male | F/M | One-Sided |
| 01–04 years | 1 | 0 | 1 | 0.007 | 0.000 | n/a | n/a |
| 05–09 years | 0 | 3 | 3 | 0.000 | 0.019 | n/a | n/a |
| 10–14 years | 2 | 8 | 10 | 0.013 | 0.049 | 0.26 | 0.0380 |
| 15–19 years | 28 | 22 | 50 | 0.174 | 0.130 | 1.34 | 0.1540 |
| 20–24 years | 46 | 39 | 85 | 0.276 | 0.225 | 1.23 | 0.1730 |
| 25–29 years | 77 | 66 | 143 | 0.45 | 0.38 | 1.19 | 0.1550 |
| 30–34 years | 122 | 120 | 242 | 0.72 | 0.70 | 1.02 | 0.4450 |
| 35–39 years | 142 | 185 | 327 | 0.88 | 1.15 | 0.76 | 0.0064 |
| 40–44 years | 223 | 282 | 505 | 1.45 | 1.88 | 0.77 | 0.0019 |
| 45–49 years | 327 | 412 | 739 | 2.29 | 2.99 | 0.77 | 0.0002 |
| 50–54 years | 495 | 577 | 1072 | 3.72 | 4.58 | 0.81 | 0.0004 |
| 55–59 years | 601 | 743 | 1344 | 5.04 | 6.74 | 0.75 | 0.0000 |
| 60–64 years | 587 | 652 | 1239 | 5.64 | 7.02 | 0.80 | 0.0001 |
| 65–69 years | 693 | 774 | 1467 | 7.72 | 10.26 | 0.75 | 0.0000 |
| 70–74 years | 578 | 597 | 1175 | 7.79 | 10.38 | 0.75 | 0.0000 |
| 75–79 years | 484 | 518 | 1002 | 8.08 | 12.49 | 0.65 | 0.0000 |
| 80–84 years | 372 | 336 | 708 | 8.47 | 12.85 | 0.66 | 0.0000 |
| 85+ years | 254 | 191 | 445 | 6.00 | 10.14 | 0.59 | 0.0000 |
| Total | 5032 | 5525 | 10,557 | 2.14 | 2.86 | 0.87 | 0.0000 |
| Gene and Status | All | Female | Male | p Value * | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | Mean | Std. Err | N | Mean | Std. Err | N | Mean | Std. Err | ||
| EIF1AX_all | 80 | 958.5 | 59.2 | 35 | 1108.6 | 101 | 45 | 841.7 | 65.9 | 0.024 |
| EFNB1_all | 80 | 322.1 | 9.8 | 35 | 315.6 | 14.9 | 45 | 327.2 | 13.1 | 0.56 |
| EIF2S3_all | 80 | 7033.7 | 342.5 | 35 | 7546.3 | 570.6 | 45 | 6635.1 | 412.9 | 0.19 |
| EIF1AX_-1 | 12 | 1271.0 | 227.2 | 7 | 1562.7 | 96 | 5 | 862.6 | 54.9 | 0.13 |
| EIF1AX_0 | 58 | 921.3 | 57.8 | 21 | 1015.5 | 47.5 | 37 | 867.9 | 49.7 | 0.22 |
| EIF1AX_1 | 10 | 798.8 | 176.3 | 7 | 933.6 | 68.9 | 3 | 484.3 | 24.1 | 0.27 |
| Regression ** | p1 = 0.030; p2 = 0.034 | p1 = 0.047; p2= 0.054 | p1 = 0.35; p2 = 0.32 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu-Smith, F.; Chiu, C.-Y.; Johnson, D.L.; Miller, P.W.; Glazer, E.S.; Wu, Z.; Wilson, M.W. The Sex Differences in Uveal Melanoma: Potential Roles of EIF1AX, Immune Response and Redox Regulation. Curr. Oncol. 2021, 28, 2801-2811. https://doi.org/10.3390/curroncol28040245
Liu-Smith F, Chiu C-Y, Johnson DL, Miller PW, Glazer ES, Wu Z, Wilson MW. The Sex Differences in Uveal Melanoma: Potential Roles of EIF1AX, Immune Response and Redox Regulation. Current Oncology. 2021; 28(4):2801-2811. https://doi.org/10.3390/curroncol28040245
Chicago/Turabian StyleLiu-Smith, Feng, Chi-Yang Chiu, Daniel L. Johnson, Phillip Winston Miller, Evan S. Glazer, Zhaohui Wu, and Matthew W. Wilson. 2021. "The Sex Differences in Uveal Melanoma: Potential Roles of EIF1AX, Immune Response and Redox Regulation" Current Oncology 28, no. 4: 2801-2811. https://doi.org/10.3390/curroncol28040245
APA StyleLiu-Smith, F., Chiu, C.-Y., Johnson, D. L., Miller, P. W., Glazer, E. S., Wu, Z., & Wilson, M. W. (2021). The Sex Differences in Uveal Melanoma: Potential Roles of EIF1AX, Immune Response and Redox Regulation. Current Oncology, 28(4), 2801-2811. https://doi.org/10.3390/curroncol28040245