
Plastic Responses in Growth, Morphology, and Biomass Allocation of Five Subtropical Tree Species to Different Degrees of Shading

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Shade Treatments
2.2. Seedling Functional Traits
2.3. Statistical Analysis
3. Results
3.1. Growth, Morphology, and Biomass Allocation
3.2. Plasticity of Traits
4. Discussion
4.1. Effects of Shading on Growth
4.2. Effects of Shading on Morphology
4.3. Effects of Shading on Allocation
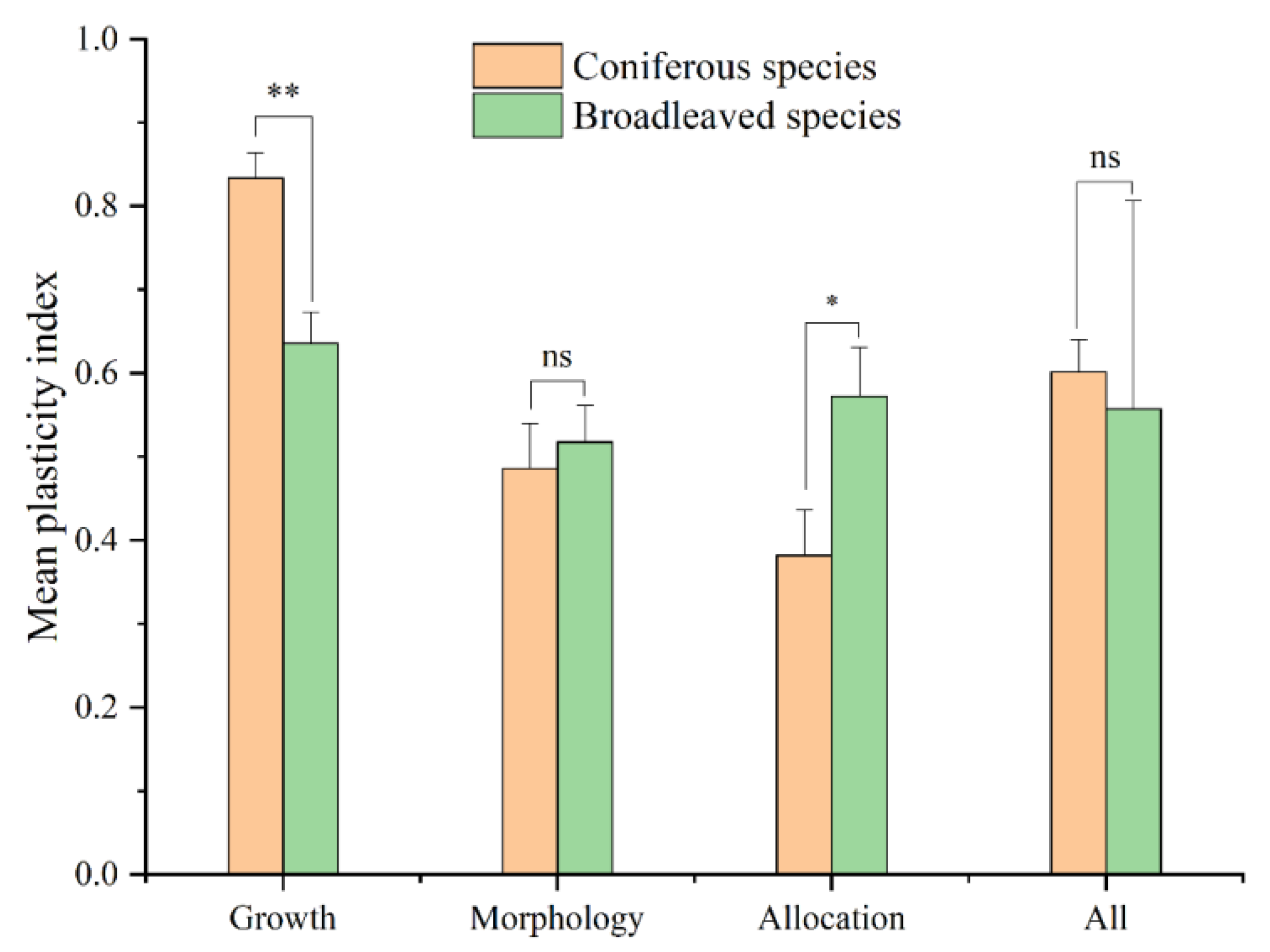
4.4. Interspecific Variation in Plasticity
4.5. Differences in Plasticity across Traits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, B.; Liu, Q.Q.; Daryanto, S.; Guo, S.; Huang, Z.J.; Wang, Z.N.; Wang, L.X.; Ma, X.Q. Responses of Chinese fir and Schimasuperba seedlings to light gradients: Implications for the restoration of mixed broadleaf-conifer forests from Chinese fir monocultures. For. Ecol. Manag. 2018, 419–420, 51–57. [Google Scholar] [CrossRef] [Green Version]
- CSFB. Forestry Resource Statistics for China in 2009–2013; The People’s Republic of China State Forestry Bureau: Beijing, China, 2014. (In Chinese) [Google Scholar]
- Erskine, P.D.; Lamb, D.; Bristow, M. Tree species diversity and ecosystem function: Can tropical multi-species plantations generate greater productivity? For. Ecol. Manag. 2006, 233, 205–210. [Google Scholar] [CrossRef]
- Wang, J.; Ren, H.; Yang, L.; Duan, W.J. Establishment and early growth of introduced indigenous tree species in typical plantations and shrubland in South China. For. Ecol. Manag. 2009, 258, 1293–1300. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.B.; Xu, Q.; Gao, D.Q.; Wang, T.; Sui, M.Z.; Huang, J.; Gu, B.H.; Liu, F.T.; Jiang, J. Soil capacity of intercepting different rainfalls across subtropical plantation: Distinct effects of plant and soil properties. Sci. Total Environ. 2021, 784, 147120. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Jucker, T.; Bouriaud, O.; Avacaritei, D.; Dănilă, I.; Duduman, G.; Valladares, F.; Coomes, D.A. Competition for light and water play contrasting roles in driving diversity-productivity relationships in Iberian forests. J. Ecol. 2014, 102, 1202–1213. [Google Scholar] [CrossRef] [Green Version]
- Alem, S.; Pavlis, J.; Urban, J.; Kucera, J. Pure and mixed plantations of Eucalyptus camaldulensis and Cupressus lusitanica: Their growth interactions and effect on diversity and density of undergrowth woody plants in relation to light. J. For. 2015, 5, 375–386. [Google Scholar]
- Chen, G.S.; Yang, Z.J.; Gao, R.; Xie, J.S.; Guo, J.F.; Huang, Z.Q.; Yang, Y.S. Carbon storage in a chronosequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013, 300, 68–76. [Google Scholar] [CrossRef]
- Yang, Q.F.; Sun, M.; Zheng, Y.Y.; Meng, X.R.; Huang, M.; Huang, Y.M.; Ding, G.C. Profiles of soil carbon, nitrogen of 23 a mixed plantation of Schima superba and Chinese fir. J. Beihua Univ. (Natur. Sci.) 2017, 18, 389–394, (in Chinese with English abstract). [Google Scholar]
- Yang, Y.S.; Wang, L.X.; Yang, Z.J.; Xu, C.; Xie, J.S.; Chen, G.S.; Lin, C.F.; Guo, J.F.; Liu, X.F.; Xiong, D.C.; et al. Large ecosystem service benefits of assisted natural regeneration. J. Geophys. Res. 2018, 123, 676–687. [Google Scholar] [CrossRef]
- Felton, A.; Nilsson, U.; Sonesson, J.; Felton, A.M.; Roberge, J.M.; Ranius, T.; Ahlström, M.; Bergh, J.; Björkman, C.; Boberg, J.; et al. Replacing monocultures with mixed-species stands: Ecosystem service implications of two production forest alternatives in Sweden. Ambio 2016, 45, 124–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Yu, M.; Wang, G.G.; Wu, T.; Zhang, C. Growth, morphology and biomass allocation in response to light gradient in five subtropical evergreen broadleaved tree seedlings. J. Trop. For. Sci. 2013, 25, 537–546. [Google Scholar]
- Myers, J.A.; Kitajima, K. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest. J. Ecol. 2007, 95, 383–395. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Jarcuska, B. Growth, survival, density, biomass partitioning and morphological adaptations of natural regeneration in Fagus sylvatica. A review. Dendrobiology 2009, 61, 3–11. [Google Scholar]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Valladares, F.; Laanisto, L.; Niinemets, Ü.; Zavala, M.A. Shedding light on shade: Ecological perspectives of understorey plant life. Plant Ecol. Divers. 2016, 9, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Sevillano, I.; Short, I.; Grant, J.; O’Reilly, C. Effects of light availability on morphology, growth and biomass allocation of Fagus sylvatica and Quercus robur seedlings. For. Ecol. Manag. 2016, 374, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.M.; Wu, T.; Geng, Y.F.; Chai, Y.; Hao, J.B. Phenotypic plasticity of lianas in response to altered light environment. Ecol. Res. 2016, 31, 375–384. [Google Scholar] [CrossRef]
- Poorter, L. Growth responses of 15 rain-forest tree species to a light gradient: The relative importance of morphological and physiological traits. Funct. Ecol. 1999, 13, 396–410. [Google Scholar] [CrossRef]
- Curt, T.; Coll, L.; Prévosto, B.; Balandier, P.; Kunstler, G. Plasticity in growth, biomass allocation and root morphology in beech seedlings as induced by irradiance and herbaceous competition. Ann. For. Sci. 2005, 62, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Lusk, C.H.; Falster, D.S.; Jara-Vergara, C.K.; Jimenez-Castillo, M.; Saldaña-Mendoza, A. Ontogenetic variation in light requirements of juvenile rainforest evergreens. Funct. Ecol. 2008, 22, 454–459. [Google Scholar] [CrossRef]
- Bastias, C.C.; Valladares, F.; Ricote, M.N.; Benavides, R. Local canopy diversity does not influence phenotypic expression and plasticity of tree seedlings exposed to different resource availabilities. Environ. Exp. Bot. 2018, 156, 38–47. [Google Scholar] [CrossRef]
- Wyka, T.P.; Oleksyn, J.; Żytkowiak, R.; Karolewski, P.; Jagodziński, A.M.; Reich, P.B. Responses of leaf structure and photosynthetic properties to intra-canopy light gradients: A common garden test with four broadleaf deciduous angiosperm and seven evergreen conifer tree species. Oecologia 2012, 170, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.Q.; Heal, K.V.; Liu, A.Q.; Jarvis, P.G. Nutrient cycling and distribution in different-aged plantations of Chinese fir in southern China. For. Ecol. Manag. 2007, 243, 61–74. [Google Scholar] [CrossRef]
- Chen, L.C.; Wang, S.L.; Wang, P.; Kong, C.H. Autoinhibition and soil allelochemical (cyclic dipeptide) levels in replanted Chinese fir (Cunninghamia lanceolata) plantations. Plant Soil. 2014, 374, 793–801. [Google Scholar] [CrossRef]
- Luo, J.; Tian, Y.X.; Zhou, X.L.; Chen, J.H.; Zeng, Z.Q.; Li, X.Q.; Yao, M. Studies on the characteristics of soil seed banks under main forest types of close-to-nature forest management in Hunan. J. Cent. South. Univ. For. Technol. 2014, 34, 56–61, (in Chinese with English abstract). [Google Scholar]
- Zhang, H.D.; Zhang, R.H.; Qi, F.; Liu, X.; Niu, Y.; Fan, Z.F.; Zhang, Q.H.; Li, J.Z.; Yuan, L.; Song, Y.Y.; et al. The CSLE model based soil erosion prediction: Comparisons of sampling density and extrapolation method at the county level. Catena 2018, 165, 465–472. [Google Scholar] [CrossRef]
- Huang, Y.S.; Wang, S.L.; Feng, Z.W.; Ouyang, Z.Y.; Wang, X.K.; Feng, Z.Z. Changes in soil quality due to introduction of broad-leaf trees into clear-felled Chinese fir forest in the mid-subtropics of China. Soil Use Manag. 2004, 20, 418–425. [Google Scholar] [CrossRef]
- Xiong, H.B. Study on variation of the properties of soil in multi-storied Chinese fir plantation. Subtrop. Agric. Res. 2007, 4, 283–286, (in Chinese with English abstract). [Google Scholar]
- Wang, H.Y.; Wu, F.; Li, M.; Zhu, X.K.; Shi, C.S.; Ding, G.J. Morphological and physiological responses of Pinus massoniana seedlings to different light gradients. Forests. 2021, 12, 523. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Huang, Z.J.; Wang, Z.N.; Chen, Y.F.; Wen, Z.M.; Liu, B.; Tigabu, M. Responses of leaf morphology, NSCs contents and C:N:P stoichiometry of Cunninghamia lanceolata and Schima superba to shading. BMC Plant Biol. 2020, 20, 354. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.I.; Khatun, S.; Azad, M.S.; Mollick, A.S. Leaf morphological and anatomical plasticity in Sundri (Heritiera fomes Buch.-Ham.) along different canopy light and salinity zones in the Sundarbans mangrove forest, Bangladesh. Glob. Ecol. Conserv. 2020, 23, e01127. [Google Scholar] [CrossRef]
- Samuelson, L.J.; Stokes, T.A. Leaf physiological and morphological responses to shade in grass-stage seedlings and young trees of longleaf pine. Forests. 2012, 3, 684–699. [Google Scholar] [CrossRef] [Green Version]
- Perrin, P.M.; Mitchell, F.J.G. Effects of shade on growth, biomass allocation and leaf morphology in European yew (Taxus baccata L.). Eur. J. Forest Res. 2013, 132, 211–218. [Google Scholar] [CrossRef]
- Poorter, L.; Werger, M.J.A. Light environment, sapling architecture, and leaf display in six rain forest tree species. Am. J. Bot. 1999, 86, 1464–1473. [Google Scholar] [CrossRef]
- Veenendaal, E.M.; Swaine, M.D.; Lecha, R.T.; Walsh, M.F.; Abebrese, I.K.; Owusu-afriyie, K. Responses of West African forest tree seedlings to irradiance and soil fertility. Funct. Ecol. 1996, 10, 501–511. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Wang, G.G.; Bauerle, W.L.; Mudder, B.T. Effects of light acclimation on the photosynthesis, growth, and biomass allocation in American chestnut (Castanea dentata) seedlings. For. Ecol. Manag. 2006, 226, 173–180. [Google Scholar] [CrossRef]
- Kennedy, S.; Black, K.; Reilly, C.O.; Dhubháin, A.N. The impact of shade on morphology, growth and biomass allocation in Picea sitchensis, Larix × eurolepis and Thuja plicata. New For. 2006, 33, 139–153. [Google Scholar] [CrossRef]
- Granata, M.U.; Bracco, F.; Catoni, R. Phenotypic plasticity of two invasive alien plant species inside a deciduous forest in a strict nature reserve in Italy. J. Sust. For. 2020, 39, 346–364. [Google Scholar] [CrossRef]
- Khan, M.N.I.; Kabir, M.E. Ecology of Kandelia obovata (S., L.) Yong: A fast-growing mangrove in Okinawa, Japan. In Participatory Mangrove Management in a Changing Climate: Perspectives from the Asia-Pacific; DasGupta, R., Shaw, R., Eds.; Springer: Japan, Tokyo, 2017; pp. 287–301. [Google Scholar]
- Dos Santos, V.A.H.F.; Ferreira, M.J. Are photosynthetic leaf traits related to the first-year growth of tropical tree seedlings? a light-induced plasticity test in a secondary forest enrichment planting. For. Ecol. Manag. 2020, 460, 117900. [Google Scholar] [CrossRef]
- Acosta, A.S.; Meave, J.A.; Sánchez-Velásquezc, L.R. Seedling biomass allocation and vital rates of cloud forest tree species: Responses to light in shade house conditions. For. Ecol. Manag. 2009, 258, 1650–1659. [Google Scholar] [CrossRef]
- Lusk, C.; Millaqueo, P.M.M.; Piper, F.I.; Saldaña, A. Ontogeny, understorey light interception and simulated carbon gain of juvenile rainforest evergreens differing in shade tolerance. Ann. Bot. 2011, 108, 419–428. [Google Scholar] [CrossRef]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Chen, F.S.; Zeng, D.H.; Fahey, T.J.; Yao, C.Y.; Yu, Z.Y. Response of leaf anatomy of Chenopodium acuminatum to soil resource availability in a semi-arid grassland. Plant Ecol. 2010, 209, 375–382. [Google Scholar] [CrossRef]
- Chmura, D.J.; Modrzyński, J.; Chmielarz, P.; Tjoelker, M.G. Plasticity in seedling morphology, biomass allocation and physiology among ten temperate tree species in response to shade is related to shade tolerance and not leaf habit. Plant Biol. 2016, 19, 172–182. [Google Scholar] [CrossRef]
- Tang, H.; Hu, Y.Y.; Yu, W.W.; Song, L.L.; Wu, J.S. Growth, photosynthetic and physiological responses of Torreya grandis seedlings to varied light environments. Trees 2015, 29, 1011–1022. [Google Scholar] [CrossRef]
- Poorter, A.H.; Nagel, O. The role of biomass allocation in the growth response to plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant. Physiol. 2000, 27, 595–607. [Google Scholar]
- Tackenberg, O. A new method for non-destructive measurement of biomass, growth rates, vertical biomass distribution and dry matter content based on digital image analysis. Ann. Bot. 2007, 99, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Portsmuth, A.; Niinemets, Ü. Structural and physiological plasticity to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance. Funct. Ecol. 2007, 21, 61–77. [Google Scholar] [CrossRef]
- Chen, J.F.; Ren, W.J.; Chou, Q.C.; Su, H.J.; Ni, L.Y.; Zhang, M.; Liu, Z.G.; Xie, P. Alterations in biomass allocation indicate the adaptation of submersed macrophytes to low-light stress. Ecol. Indic. 2020, 113, 106235. [Google Scholar] [CrossRef]
- Poorter, L. Resource capture and use by tropical forest tree seedlings and their consequences for competition. In Biotic Interactions in the Tropics: Their Role in the Maintenance of Species Diversity; Burslem, D., Pinard, M., Hartley, S., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 35–64. [Google Scholar]
- Sánchez-Gómez, D.; Valladares, F.; Zavala, M.A. Functional traits and plasticity in response to light in seedlings of four Iberian forest tree species. Tree Physiol. 2006, 26, 1425–1433. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.B.; Zhang, H.; Jing, Q.; Wang, J.X. Light pollution on the growth, physiology and chlorophyll fluorescence T response of landscape plant perennial ryegrass (Lolium perenne L.). Ecol. Indic. 2020, 115, 106448. [Google Scholar] [CrossRef]
- Bloor, J.M.G.; Grubb, P.J. Morphological plasticity of shade-tolerant tropical rainforest tree seedlings exposed to light changes. Funct. Ecol. 2004, 18, 337–348. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Laughlin, D.C. Root nutrient concentration and biomass allocation are more plastic than morphological traits in response to nutrient limitation. Plant Soil. 2017, 416, 539–550. [Google Scholar] [CrossRef]
- Huang, Y.X.; Fan, G.H.; Zhou, D.W.; Pang, J.Y. Phenotypic plasticity of four Chenopodiaceae species with contrasting saline-sodic tolerance in response to increased salinity–sodicity. Ecol. Evol. 2019, 9, 1545–1553. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Wright, I.J.; Bares, J.C.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The evolution of plant functional variation: Traits, spectra and strategies. Int. J. Plant Sci. 2003, 164, 143–164. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Shade | Species | ||||
|---|---|---|---|---|---|---|
| C. lanceolata | P. massoniana | S. superba | P. zhennan | L. formosana | ||
| Root biomass (g) | 0% | 10.6 ± 1.5 a | 4.8 ± 1.0 a | 14.5 ± 2.1 a | 7.0 ± 0.2 b | 14.5 ± 1.6 a |
| 40% | 10.1 ± 1.4 a | 1.7 ± 0.3 b | 11.7 ± 0.8 a | 9.9 ± 1.0 a | 13.6 ± 0.1 ab | |
| 60% | 8.2 ± 0.8 a | 0.8 ± 0.1 bc | 7.8 ± 1.1 b | 6.3 ± 0.1 b | 9.7 ± 2.3 bc | |
| 85% | 4.2 ± 1.0 b | 0.3 ± 0.1 bc | 4.4 ± 0.3 bc | 5.6 ± 1.4 b | 6.7 ± 0.6 c | |
| 95% | 1.2 ± 0.1 b | 0.2 ± 0.0 c | 1.7 ± 0.1 c | 1.9 ± 0.1 c | 1.8 ± 0.3 d | |
| Stem biomass (g) | 0% | 7.3 ± 0.7 a | 6.4 ± 1.3 a | 5.8 ± 1.0 a | 3.3 ± 0.2 bc | 11.7 ± 2.7 a |
| 40% | 5.4 ± 0.7 b | 4.6 ± 1.3 a | 6.8 ± 0.4 a | 5.3 ± 0.3 ab | 11.1 ± 2.1 a | |
| 60% | 6.2 ± 0.3 ab | 1.8 ± 0.3 b | 7.5 ± 0.7 a | 6.2 ± 0.4 a | 12.1 ± 2.6 a | |
| 85% | 5.6 ± 0.8 ab | 1.0 ± 0.1 b | 5.6 ± 0.2 a | 6.7 ± 1.7 a | 9.9 ± 1.2 ab | |
| 95% | 2.0 ± 0.0 c | 0.7 ± 0.1 b | 2.6 ± 0.3 b | 2.6 ± 0.3 c | 4.2 ± 0.3 b | |
| Leaf biomass (g) | 0% | 10.2 ± 1.5 a | 10.8 ± 1.8 a | 5.7 ± 0.2 b | 2.7 ± 0.1 c | 4.3 ± 0.9 a |
| 40% | 11.3 ± 1.1 a | 6.1 ± 1.6 b | 7.8 ± 0.6 a | 5.5 ± 0.3 ab | 6.5 ± 0.7 a | |
| 60% | 12.2 ± 0.6 a | 2.5 ± 0.3 c | 9.0 ± 0.4 a | 6.5 ± 0.8 ab | 6.3 ± 1.6 a | |
| 85% | 9.7 ± 1.2 a | 1.1 ± 0.2 c | 7.9 ± 0.7 a | 7.6 ± 1.6 a | 4.0 ± 0.6 a | |
| 95% | 3.5 ± 0.3 b | 0.8 ± 0.1 c | 6.2 ± 0.3 b | 4.5 ± 0.2 bc | 3.0 ± 0.5 b | |
| Total biomass (g) | 0% | 28.0 ± 3.5 a | 22.0 ± 3.9 a | 26.0 ± 2.9 a | 13.0 ± 0.4 ab | 30.4 ± 5.0 a |
| 40% | 26.7 ± 3.3 ab | 12.4 ± 3.1 b | 26.3 ± 0.4 a | 20.7 ± 1.5 a | 25.4 ± 2.6 a | |
| 60% | 26.6 ± 0.9 ab | 5.1 ± 0.7 c | 24.4 ± 1.4 a | 19.0 ± 1.0 a | 34.2 ± 6.4 a | |
| 85% | 19.4 ± 3.0 b | 2.4 ± 0.3 c | 17.9 ± 1.0 b | 19.8 ± 4.6 a | 17.9 ± 1.8 ab | |
| 95% | 6.7 ± 0.3 c | 1.7 ± 0.1 c | 10.6 ± 0.5 c | 9.0 ± 0.5 b | 10.5 ± 0.6 b | |
| Relative growth rate (RGR, g·g−1·month−1) | 0% | 0.19 ± 0.01 a | 0.24 ± 0.02 a | 0.21 ± 0.01 a | 0.12 ± 0.00 b | 0.20 ± 0.01 a |
| 40% | 0.19 ± 0.01 a | 0.19 ± 0.02 b | 0.21 ± 0.00 a | 0.15 ± 0.01 a | 0.18 ± 0.01 ab | |
| 60% | 0.19 ± 0.00 a | 0.12 ± 0.01 c | 0.20 ± 0.00 a | 0.15 ± 0.00 a | 0.21 ± 0.01 a | |
| 85% | 0.16 ± 0.01 b | 0.05 ± 0.01 d | 0.18 ± 0.00 b | 0.15 ± 0.02 a | 0.15 ± 0.02 bc | |
| 95% | 0.07 ± 0.00 c | 0.03 ± 0.01 d | 0.13 ± 0.00 c | 0.08 ± 0.00 c | 0.11 ± 0.02 c | |
| Net assimilation rate (NAR, g·cm−2·day−1) | 0% | 1.94 ± 0.11 a | 2.06 ± 0.18 a | 2.44 ± 0.28 a | 1.47 ± 0.03 a | 3.19 ± 0.21 a |
| 40% | 1.39 ± 0.10 b | 1.58 ± 0.18 b | 1.66 ± 0.08 b | 1.48 ± 0.09 a | 2.74 ± 0.40 a | |
| 60% | 1.26 ± 0.03 b | 0.83 ± 0.08 c | 1.29 ± 0.11 bc | 1.21 ± 0.05 b | 1.57 ± 0.42 b | |
| 85% | 0.84 ± 0.08 c | 0.34 ± 0.07 d | 0.92 ± 0.03 cd | 1.01 ± 0.14 b | 1.33 ± 0.11 b | |
| 95% | 0.33 ± 0.02 d | 0.21 ± 0.05 d | 0.50 ± 0.02 d | 0.44 ± 0.02 c | 0.70 ± 0.22 b | |
| Trait | Shade | Species | ||||
|---|---|---|---|---|---|---|
| C. lanceolata | P. massoniana | S. superba | P. zhennan | L. formosana | ||
| Height (cm) | 0% | 33.1 ± 1.3 c | 39.1 ± 0.9 a | 45.1 ± 1.5 c | 40.2 ± 1.8 bc | 64.5 ± 2.4 b |
| 40% | 32.5 ± 0.9 c | 43.0 ± 2.2 a | 55.5 ± 5.2 bc | 51.1 ± 3.3 ab | 68.4 ± 3.5 b | |
| 60% | 36.4 ± 0.9 b | 39.9 ± 1.1 a | 57.6 ± 1.3 b | 56.0 ± 5.3 a | 83.6 ± 4.1 a | |
| 85% | 42.4 ± 1.4 a | 33.7 ± 1.5 b | 70.3 ± 6.2 a | 56.4 ± 5.7 a | 84.7 ± 1.8 a | |
| 95% | 33.8 ± 0.6 bc | 28.5 ± 0.9 c | 50.9 ± 4.4 bc | 38.5 ± 1.8 c | 63.8 ± 3.0 b | |
| Basal diameter (cm) | 0% | 0.55 ± 0.05 a | 0.66 ± 0.05 a | 0.63 ± 0.02 a | 0.59 ± 0.04 b | 0.88 ± 0.05 a |
| 40% | 0.52 ± 0.04 ab | 0.53 ± 0.04 b | 0.68 ± 0.03 a | 0.67 ± 0.01 a | 0.74 ± 0.03 ab | |
| 60% | 0.54 ± 0.02 ab | 0.42 ± 0.03 c | 0.62 ± 0.02 a | 0.64 ± 0.02 ab | 0.74 ± 0.09 ab | |
| 85% | 0.45 ± 0.02 bc | 0.33 ± 0.01 cd | 0.60 ± 0.03 a | 0.67 ± 0.02 a | 0.66 ± 0.04 b | |
| 95% | 0.36 ± 0.01 c | 0.28 ± 0.01 d | 0.39 ± 0.02 b | 0.45 ± 0.02 c | 0.39 ± 0.02 c | |
| Slenderness quotient (SQ) | 0% | 61 ± 3 b | 61 ± 7 c | 72 ± 4 c | 70 ± 5 a | 74 ± 6 c |
| 40% | 63 ± 5 b | 82 ± 3 b | 83 ± 9bc | 76 ± 4 a | 93 ± 7 c | |
| 60% | 68 ± 4 b | 96 ± 6 ab | 94 ± 4 b | 87 ± 9 a | 119 ± 12 b | |
| 85% | 94 ± 4 a | 102 ± 4 a | 118 ± 10 a | 85 ± 9 a | 130 ± 5 b | |
| 95% | 94 ± 3 a | 103 ± 6 a | 130 ± 7 a | 88 ± 8 a | 165 ± 5 a | |
| Specific leaf area (SLA, cm−2·g−1) | 0% | 74 ± 8 d | 71 ± 1 c | 99 ± 7 d | 95 ± 2 d | 126 ± 3 c |
| 40% | 100 ± 2 c | 73 ± 2 c | 134 ± 5 c | 122 ± 6 c | 135 ± 4 c | |
| 60% | 109 ± 2 c | 96 ± 3 b | 148 ± 5 c | 122 ± 5 c | 164 ± 7 c | |
| 85% | 144 ± 4 b | 127 ± 8 a | 167 ± 4 b | 141 ± 4 b | 278 ± 17 b | |
| 95% | 174 ± 7 a | 127 ± 8 a | 208 ± 8 a | 171 ± 3 a | 341 ± 28 a | |
| Leaf area ratio (LAR, g·g−1) | 0% | 25 ± 1 e | 35 ± 1 c | 22 ± 2 e | 20 ± 0 e | 18 ± 1 b |
| 40% | 43 ± 1 d | 36 ± 1 c | 40 ± 3 d | 33 ± 2 d | 29 ± 3 ab | |
| 60% | 50 ± 1 c | 50 ± 1 ab | 55 ± 4 c | 42 ± 4 c | 38 ± 2 ab | |
| 85% | 73 ± 2 b | 56 ± 5 a | 73 ± 4 b | 55 ± 4 b | 66 ± 19 ab | |
| 95% | 93 ± 4 a | 47 ± 3 b | 123 ± 4 a | 86 ± 0 a | 79 ± 31 a | |
| Leaf area (LA, cm2) | 0% | 655 ± 66 b | 765 ± 124 a | 566 ± 19 c | 259 ± 13 c | 541 ± 111 c |
| 40% | 1134 ± 114 a | 444 ± 118 b | 1044 ± 81 b | 672 ± 36 b | 726 ± 61bc | |
| 60% | 1325 ± 63 a | 273 ± 21 bc | 1336 ± 52 a | 796 ± 99 ab | 1281 ± 59 a | |
| 85% | 1392 ± 165 a | 134 ± 22 c | 1313 ± 119 a | 1070 ± 225 a | 1042 ± 135 ab | |
| 95% | 603 ± 50 b | 81 ± 1 c | 1300 ± 51 a | 776 ± 40 ab | 714 ± 212 bc | |
| Trait | Shade | Species | ||||
|---|---|---|---|---|---|---|
| C. lanceolata | P. massoniana | S. superba | P. zhennan | L. formosana | ||
| Root mass ratio (RMR) | 0% | 0.38 ± 0.01 a | 0.22 ± 0.01 a | 0.55 ± 0.03 a | 0.54 ± 0.01 a | 0.48 ± 0.03 a |
| 40% | 0.37 ± 0.01 a | 0.15 ± 0.01 b | 0.44 ± 0.03 b | 0.48 ± 0.02 b | 0.44 ± 0.04 a | |
| 60% | 0.31 ± 0.02 b | 0.15 ± 0.01 b | 0.32 ± 0.03 c | 0.33 ± 0.01 c | 0.35 ± 0.00 b | |
| 85% | 0.21 ± 0.02 c | 0.13 ± 0.02 b | 0.25 ± 0.01 d | 0.28 ± 0.02 d | 0.33 ± 0.00 b | |
| 95% | 0.19 ± 0.01 c | 0.13 ± 0.01 b | 0.16 ± 0.01 e | 0.21 ± 0.02 e | 0.19 ± 0.02 c | |
| Stem mass ratio (SMR) | 0% | 0.26 ± 0.01 ab | 0.29 ± 0.02 b | 0.22 ± 0.02 b | 0.25 ± 0.01 c | 0.38 ± 0.02 ab |
| 40% | 0.20 ± 0.00 c | 0.37 ± 0.02 ab | 0.26 ± 0.02 ab | 0.26 ± 0.01 c | 0.35 ± 0.04 b | |
| 60% | 0.23 ± 0.02 bc | 0.36 ± 0.01 ab | 0.31 ± 0.02 a | 0.33 ± 0.02 ab | 0.43 ± 0.02 ab | |
| 85% | 0.29 ± 0.01 a | 0.43 ± 0.04 a | 0.31 ± 0.01 a | 0.33 ± 0.01 a | 0.48 ± 0.03 a | |
| 95% | 0.30 ± 0.01 a | 0.43 ± 0.03 a | 0.25 ± 0.02 b | 0.29 ± 0.02 bc | 0.47 ± 0.05 c | |
| Leaf mass ratio (LMR) | 0% | 0.36 ± 0.02 d | 0.49 ± 0.02 a | 0.22 ± 0.02 c | 0.21 ± 0.01 c | 0.14 ± 0.01 b |
| 40% | 0.42 ± 0.01 c | 0.49 ± 0.01 a | 0.30 ± 0.02 c | 0.27 ± 0.01 c | 0.21 ± 0.01 b | |
| 60% | 0.46 ± 0.01 bc | 0.49 ± 0.01 a | 0.37 ± 0.03 b | 0.34 ± 0.03 b | 0.22 ± 0.02 b | |
| 85% | 0.50 ± 0.02 ab | 0.44 ± 0.04 a | 0.44 ± 0.02 b | 0.39 ± 0.03 b | 0.19 ± 0.03 b | |
| 95% | 0.52 ± 0.02 a | 0.44 ± 0.03 a | 0.59 ± 0.02 a | 0.50 ± 0.00 a | 0.33 ± 0.05 a | |
| Root-to-shoot ratio (R∶S) | 0% | 0.61 ± 0.03 a | 0.28 ± 0.01 a | 1.26 ± 0.14 a | 1.16 ± 0.05 a | 0.95 ± 0.11 a |
| 40% | 0.60 ± 0.02 a | 0.17 ± 0.02 b | 0.81 ± 0.10 b | 0.91 ± 0.06 b | 0.81 ± 0.12 a | |
| 60% | 0.45 ± 0.05 b | 0.18 ± 0.02 b | 0.47 ± 0.06 c | 0.50 ± 0.03 c | 0.53 ± 0.01 b | |
| 85% | 0.27 ± 0.03 c | 0.15 ± 0.02 b | 0.33 ± 0.03 cd | 0.39 ± 0.05 cd | 0.48 ± 0.01 bc | |
| 95% | 0.23 ± 0.02 c | 0.15 ± 0.01 b | 0.19 ± 0.01 d | 0.26 ± 0.03 d | 0.24 ± 0.03 c | |
| Group | Trait | Phenotypic Plasticity Index(PI) | ||||
|---|---|---|---|---|---|---|
| C.lanceolata | P. massoniana | S. superba | P. zhennan | L. formosana | ||
| Growth | Root biomass | 0.88 | 0.95 | 0.88 | 0.81 | 0.88 |
| Stem biomass | 0.73 | 0.89 | 0.65 | 0.61 | 0.65 | |
| Leaf biomass | 0.71 | 0.93 | 0.37 | 0.64 | 0.53 | |
| Total biomass | 0.76 | 0.92 | 0.60 | 0.56 | 0.69 | |
| RGR | 0.62 | 0.88 | 0.36 | 0.45 | 0.48 | |
| NAR | 0.83 | 0.90 | 0.80 | 0.70 | 0.78 | |
| Morphology | Height | 0.23 | 0.34 | 0.36 | 0.32 | 0.25 |
| Basal diameter | 0.35 | 0.58 | 0.42 | 0.33 | 0.56 | |
| SQ | 0.35 | 0.41 | 0.44 | 0.21 | 0.55 | |
| SLA | 0.57 | 0.44 | 0.52 | 0.44 | 0.63 | |
| LAR | 0.73 | 0.37 | 0.82 | 0.77 | 0.78 | |
| LA | 0.57 | 0.89 | 0.58 | 0.76 | 0.58 | |
| Biomass allocation | SMR | 0.32 | 0.33 | 0.29 | 0.23 | 0.27 |
| LMR | 0.30 | 0.11 | 0.62 | 0.59 | 0.58 | |
| RMR | 0.51 | 0.40 | 0.71 | 0.61 | 0.60 | |
| R∶S | 0.62 | 0.46 | 0.85 | 0.77 | 0.74 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Z.; Liu, Q.; Tigabu, M.; Jin, S.; Ma, X.; Liu, B. Plastic Responses in Growth, Morphology, and Biomass Allocation of Five Subtropical Tree Species to Different Degrees of Shading. Forests 2022, 13, 996. https://doi.org/10.3390/f13070996
Huang Z, Liu Q, Tigabu M, Jin S, Ma X, Liu B. Plastic Responses in Growth, Morphology, and Biomass Allocation of Five Subtropical Tree Species to Different Degrees of Shading. Forests. 2022; 13(7):996. https://doi.org/10.3390/f13070996
Chicago/Turabian StyleHuang, Zhijun, Qingqing Liu, Mulualem Tigabu, Shaofei Jin, Xiangqing Ma, and Bo Liu. 2022. "Plastic Responses in Growth, Morphology, and Biomass Allocation of Five Subtropical Tree Species to Different Degrees of Shading" Forests 13, no. 7: 996. https://doi.org/10.3390/f13070996
APA StyleHuang, Z., Liu, Q., Tigabu, M., Jin, S., Ma, X., & Liu, B. (2022). Plastic Responses in Growth, Morphology, and Biomass Allocation of Five Subtropical Tree Species to Different Degrees of Shading. Forests, 13(7), 996. https://doi.org/10.3390/f13070996