
Creatine Supplementation Alleviates Fatigue after Exercise through Anti-Inflammatory Action in Skeletal Muscle and Brain


Abstract
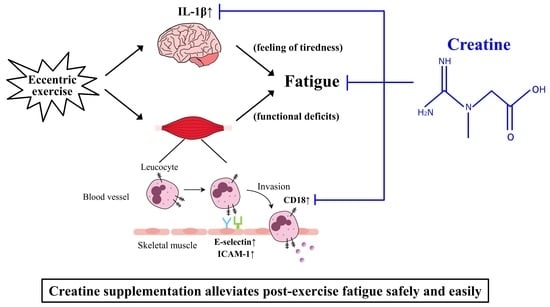
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Exercise Protocol
2.3. Dietary Intervention
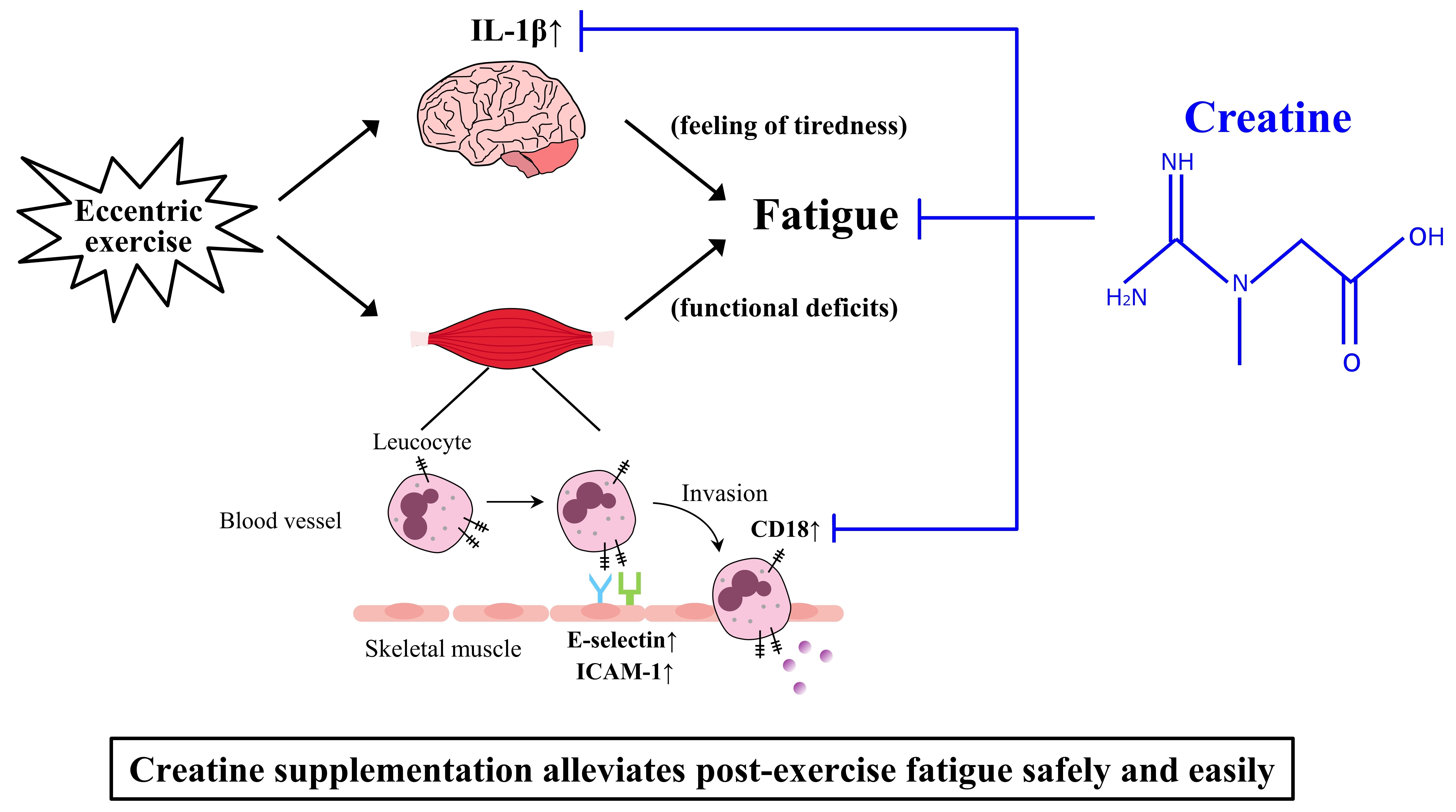
2.4. Experimental Design
2.4.1. Experiment 1-1
2.4.2. Experiment 1-2
2.4.3. Experiment 1-3
2.4.4. Experiment 2
2.5. Measurement of Total Creatine Concentration in Skeletal Muscle and Brain Tissue
2.6. Measurement of Urine Concentration of Titin N-Fragment
2.7. RNA Extraction and Quantitative Real Time PCR
2.8. Statistical Analyses
3. Results
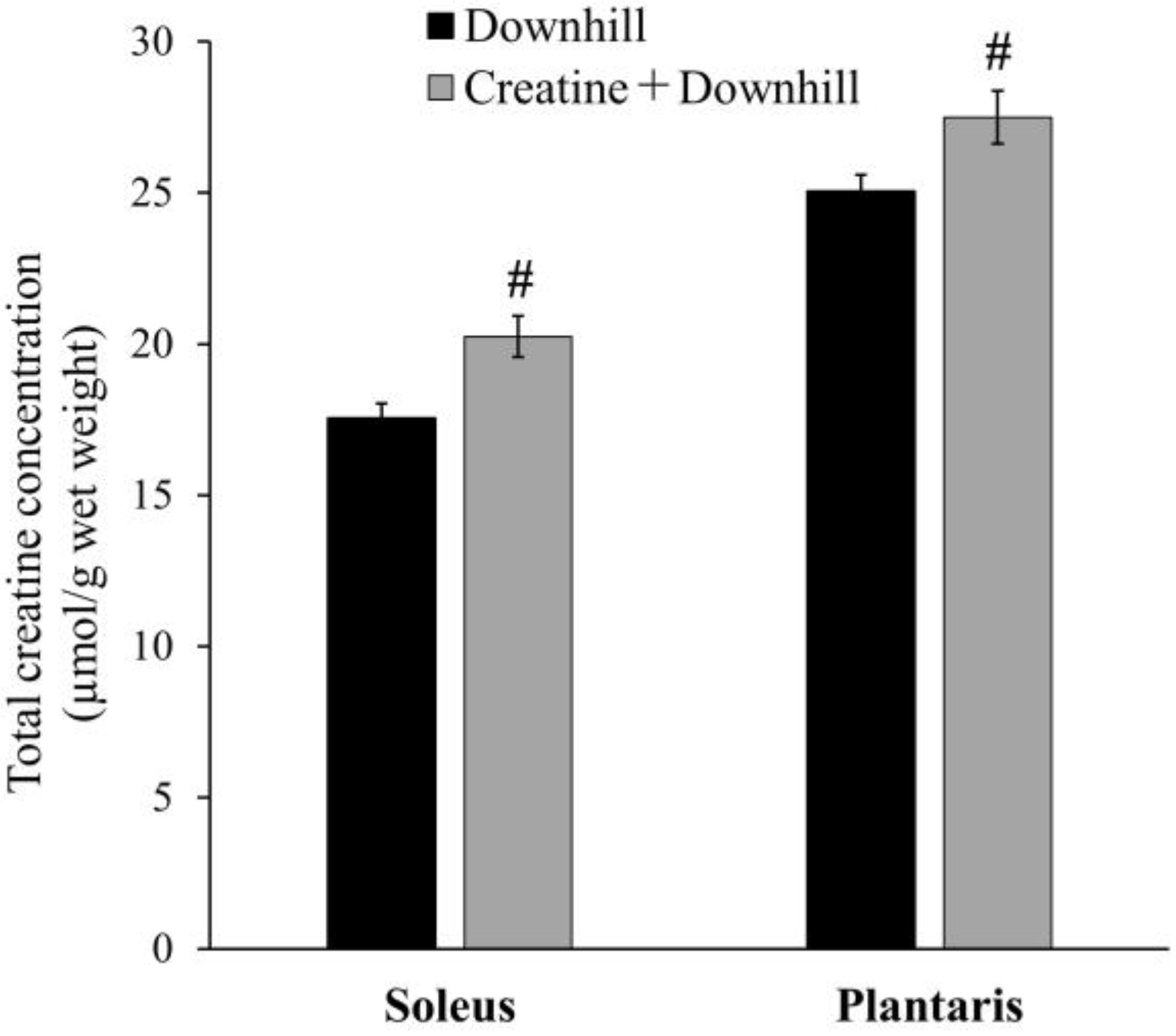
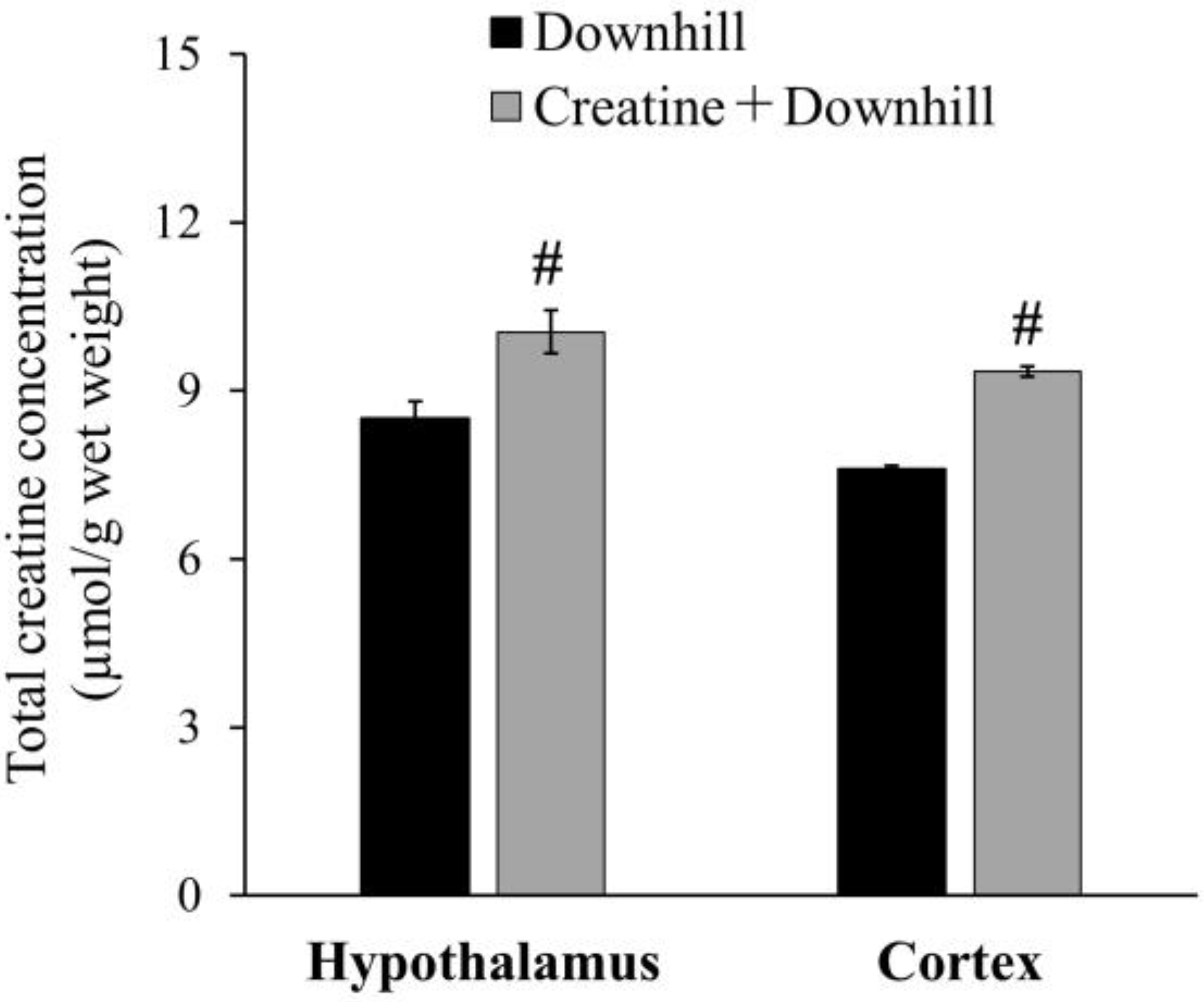
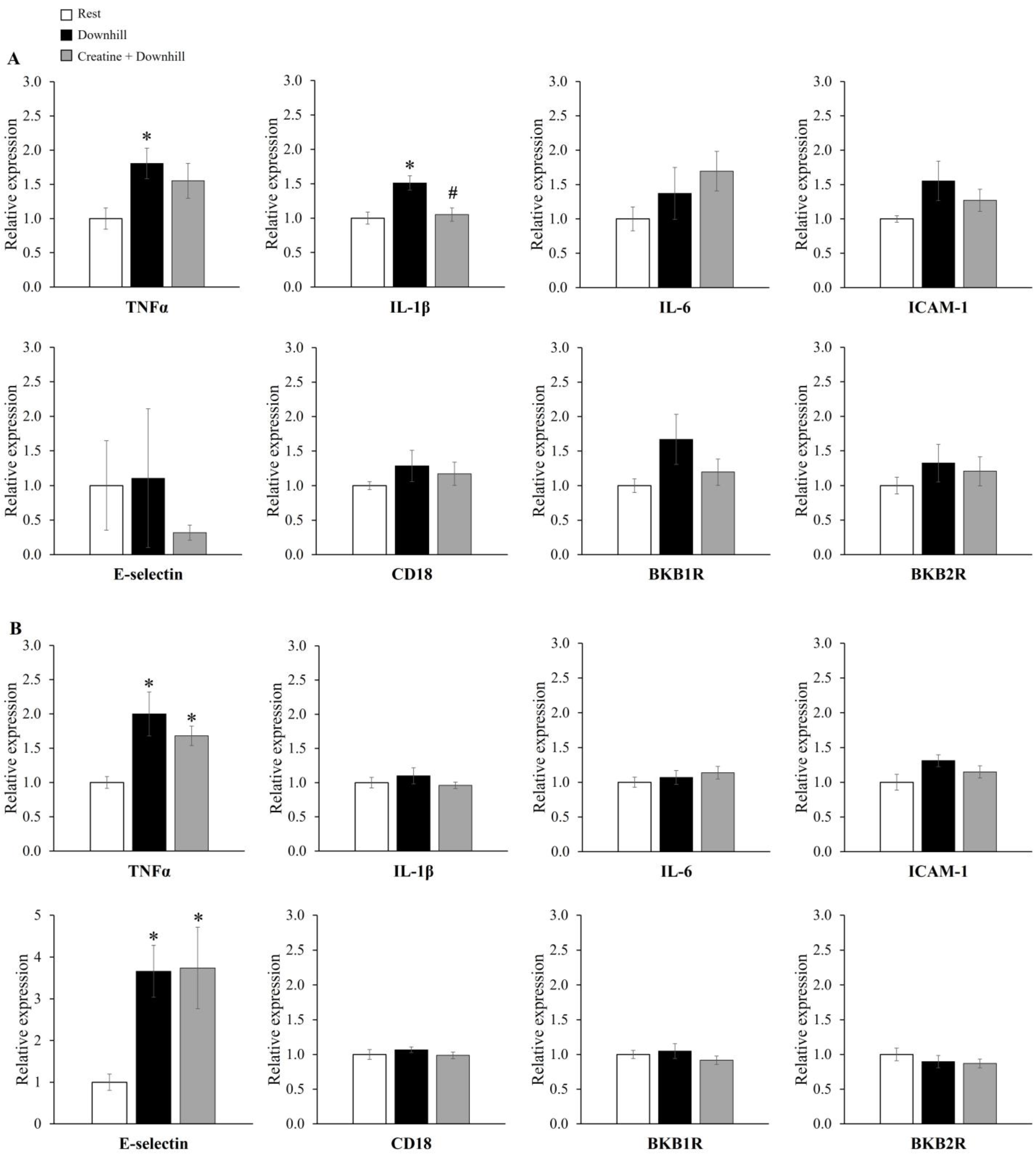
3.1. Experiment 1-1
3.2. Experiment 1-2
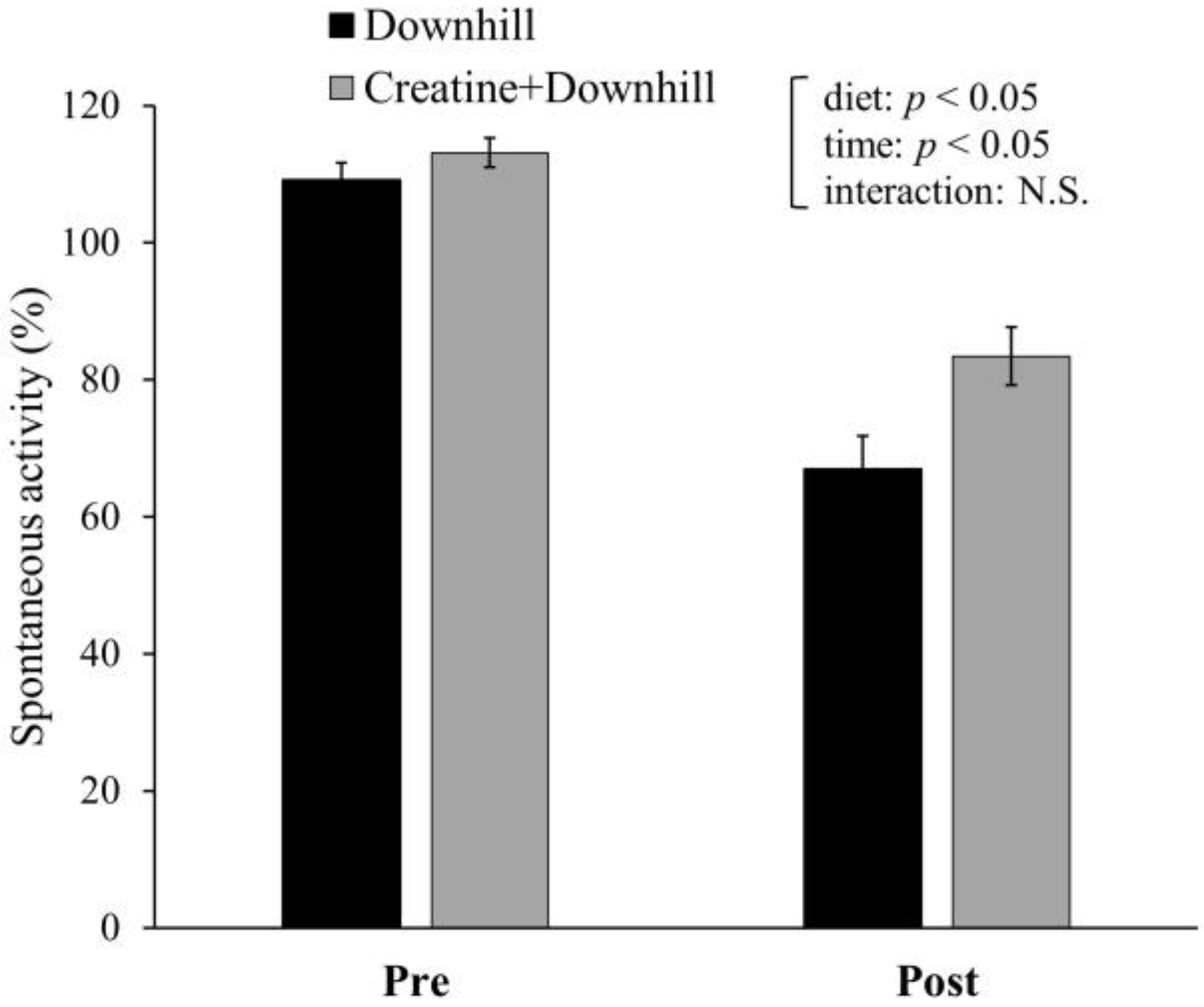
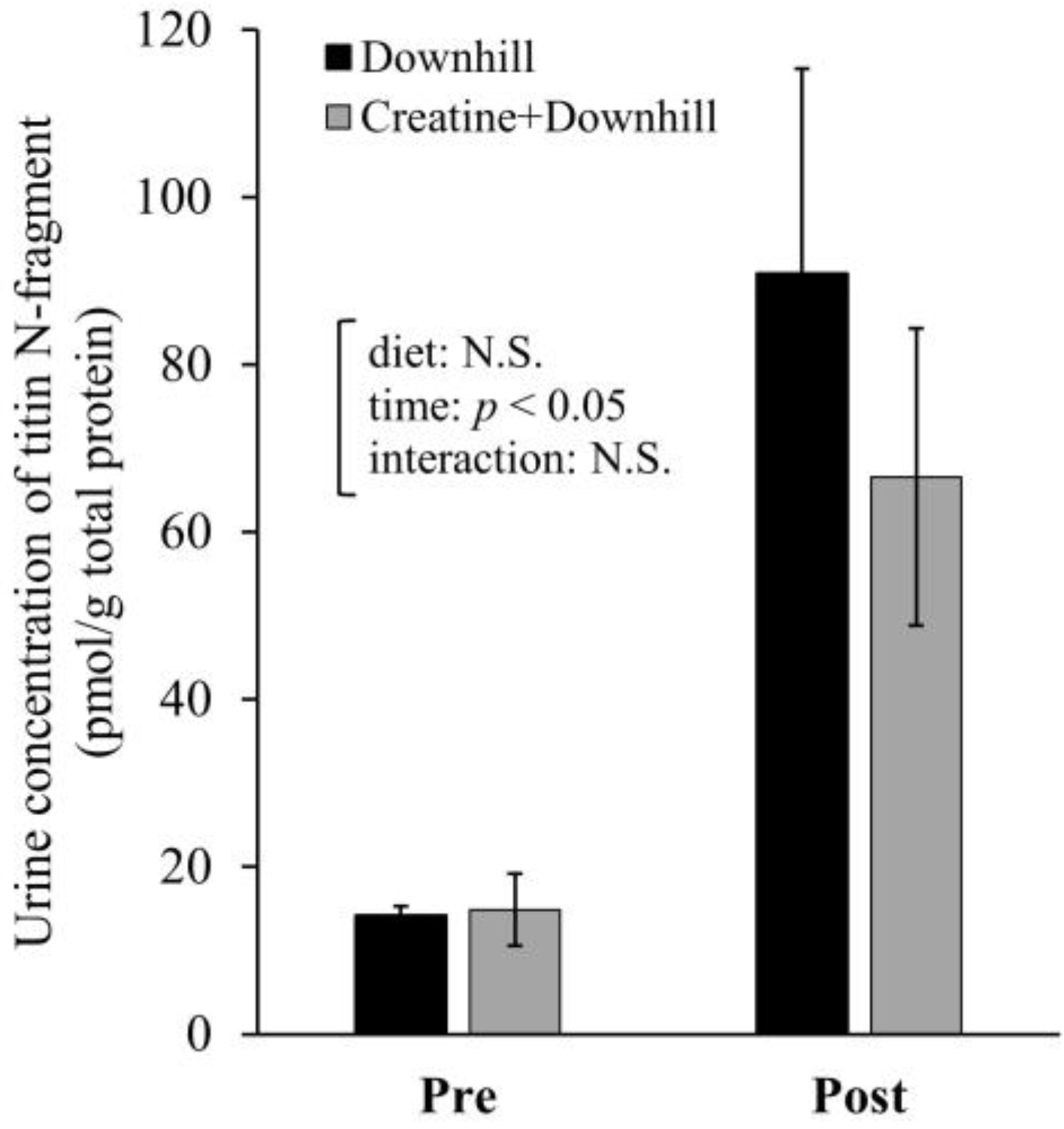
3.3. Experiment 1-3
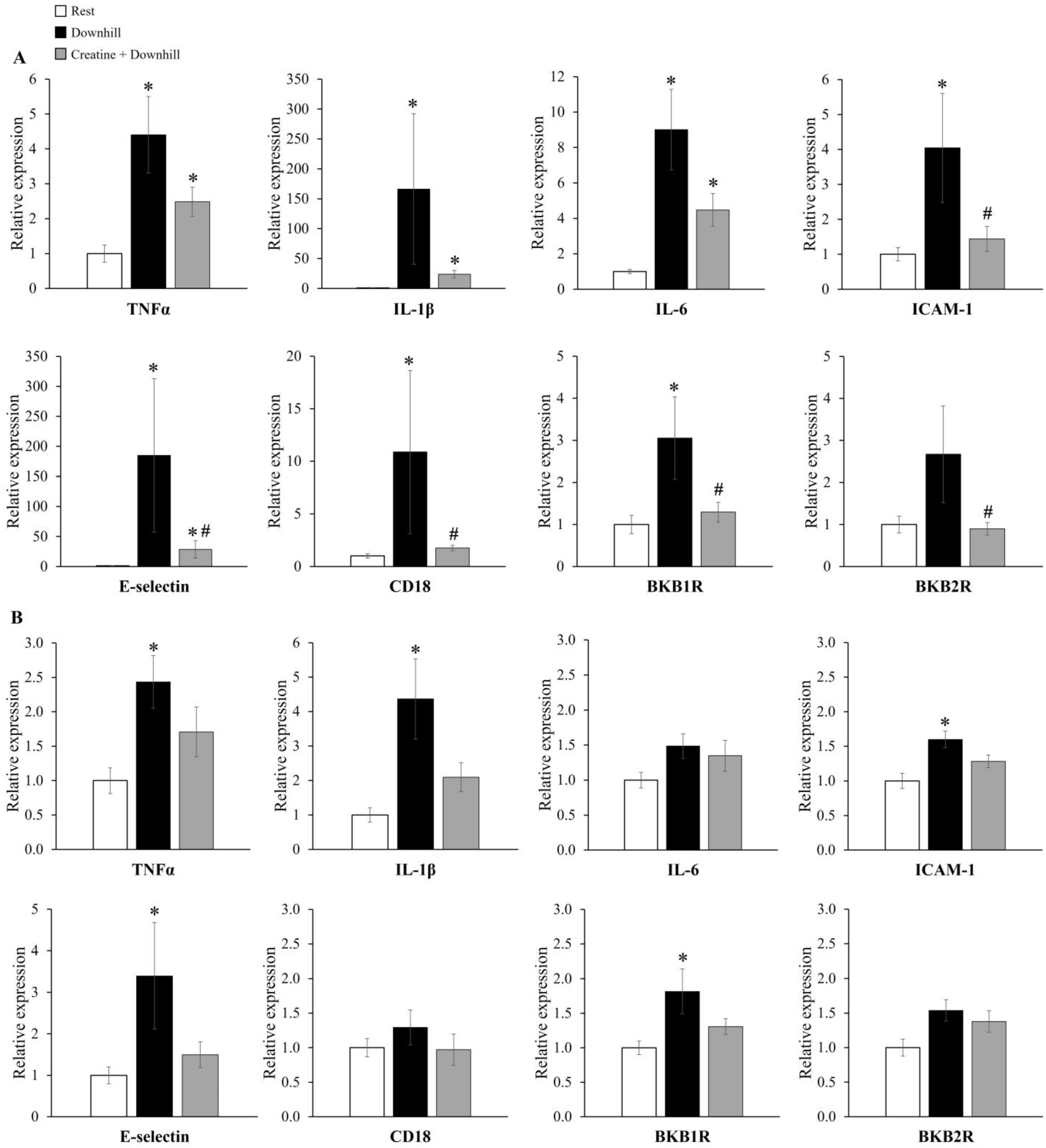
3.4. Experiment 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Byrne, C.; Twist, C.; Eston, R. Neuromuscular Function after Exercise-Induced Muscle Damage: Theoretical and Applied Implications. Sport. Med. 2004, 34, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Hody, S.; Croisier, J.L.; Bury, T.; Rogister, B.; Leprince, P. Eccentric Muscle Contractions: Risks and Benefits. Front. Physiol. 2019, 10, 536. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.C.; Fyfe, J.J. Post-Exercise Cold Water Immersion Effects on Physiological Adaptations to Resistance Training and the Underlying Mechanisms in Skeletal Muscle: A Narrative Review. Front. Sport. Act. Living 2021, 3, 660291. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, J.A.; Bishop, D.J. Effects of Dietary Supplements on Adaptations to Endurance Training. Sport. Med. 2020, 50, 25–53. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.; Teixeira, V.H.; Soares, J. Dietary Strategies to Recover from Exercise-Induced Muscle Damage. Int. J. Food Sci. Nutr. 2014, 65, 151–163. [Google Scholar] [CrossRef]
- Yamada, R.; Himori, K.; Tatebayashi, D.; Ashida, Y.; Ikezaki, K.; Miyata, H.; Kanzaki, K.; Wada, M.; Westerblad, H.; Yamada, T. Preconditioning Contractions Prevent the Delayed Onset of Myofibrillar Dysfunction after Damaging Eccentric Contractions. J. Physiol. 2018, 596, 4427–4442. [Google Scholar] [CrossRef]
- Tee, J.C.; Bosch, A.N.; Lambert, M.I. Metabolic Consequences of Exercise-Induced Muscle Damage. Sport. Med. 2007, 37, 827–836. [Google Scholar] [CrossRef]
- Kirwan, J.P.; del Aguila, L.F. Insulin Signalling, Exercise and Cellular Integrity. Biochem. Soc. Trans. 2003, 31, 1281–1285. [Google Scholar] [CrossRef]
- del Aguila, L.F.; Krishnan, R.K.; Ulbrecht, J.S.; Farrell, P.A.; Correll, P.H.; Lang, C.H.; Zierath, J.R.; Kirwan, J.P. Muscle Damage Impairs Insulin Stimulation of IRS-1, PI 3-Kinase, and Akt-Kinase in Human Skeletal Muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 279, 206–212. [Google Scholar] [CrossRef]
- Kirwan, J.P.; Hickner, R.C.; Yarasheski, K.E.; Kohrt, W.M.; Wiethop, B.V.; Holloszy, J.O. Eccentric Exercise Induces Transient Insulin Resistance in Healthy Individuals. J. Appl. Physiol. 1992, 72, 2197–2202. [Google Scholar] [CrossRef]
- Hickner, R.C.; Mehta, P.M.; Dyck, D.; Devita, P.; Houmard, J.A.; Koves, T.; Byrd, P. Relationship between Fat-to-Fat-Free Mass Ratio and Decrements in Leg Strength after Downhill Running. J. Appl. Physiol. 2001, 90, 1334–1341. [Google Scholar] [CrossRef]
- Zehnder, M.; Muelli, M.; Buchli, R.; Kuehne, G.; Boutellier, U. Further Glycogen Decrease during Early Recovery after Eccentric Exercise despite a High Carbohydrate Intake. Eur. J. Nutr. 2004, 43, 148–159. [Google Scholar] [CrossRef]
- Pizza, F.X.; Peterson, J.M.; Baas, J.H.; Koh, T.J. Neutrophils Contribute to Muscle Injury and Impair Its Resolution after Lengthening Contractions in Mice. J. Physiol. 2005, 562, 899–913. [Google Scholar] [CrossRef]
- Chen, T.C.; Nosaka, K.; Tu, J.H. Changes in Running Economy Following Downhill Running. J. Sport. Sci. 2007, 25, 55–63. [Google Scholar] [CrossRef]
- Chen, T.C.; Nosaka, K.; Lin, M.J.; Chen, H.L.; Wu, C.J. Changes in Running Economy at Different Intensities Following Downhill Running. J. Sport. Sci. 2009, 27, 1137–1144. [Google Scholar] [CrossRef]
- Burt, D.G.; Lamb, K.; Nicholas, C.; Twist, C. Effects of Exercise-Induced Muscle Damage on Resting Metabolic Rate, Sub-Maximal Running and Post-Exercise Oxygen Consumption. Eur. J. Sport. Sci. 2014, 14, 337–344. [Google Scholar] [CrossRef]
- Burt, D.; Lamb, K.; Nicholas, C.; Twist, C. Lower-Volume Muscle-Damaging Exercise Protects against High-Volume Muscle-Damaging Exercise and the Detrimental Effects on Endurance Performance. Eur. J. Appl. Physiol. 2015, 115, 1523–1532. [Google Scholar] [CrossRef]
- Proschinger, S.; Freese, J. Neuroimmunological and Neuroenergetic Aspects in Exercise-Induced Fatigue. Exerc. Immunol. Rev. 2019, 25, 8–19. [Google Scholar]
- Carmichael, M.D.; Davis, J.M.; Murphy, E.A.; Brown, A.S.; Carson, J.A.; Mayer, E.; Ghaffar, A. Recovery of Running Performance Following Muscle-Damaging Exercise: Relationship to Brain IL-1β. Brain Behav. Immun. 2005, 19, 445–452. [Google Scholar] [CrossRef]
- Carmichael, M.D.; Davis, J.M.; Murphy, E.A.; Brown, A.S.; Carson, J.A.; Mayer, E.P.; Ghaffar, A. Role of Brain IL-1β on Fatigue after Exercise-Induced Muscle Damage. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, 1344–1348. [Google Scholar] [CrossRef]
- Roerink, M.E.; van der Schaaf, M.E.; Dinarello, C.A.; Knoop, H.; van der Meer, J.W.M. Interleukin-1 as a Mediator of Fatigue in Disease: A Narrative Review. J. Neuroinflam. 2017, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Kreider, R.B.; Kalman, D.S.; Antonio, J.; Ziegenfuss, T.N.; Wildman, R.; Collins, R.; Candow, D.G.; Kleiner, S.M.; Almada, A.L.; Lopez, H.L. International Society of Sports Nutrition Position Stand: Safety and Efficacy of Creatine Supplementation in Exercise, Sport, and Medicine. J. Int. Soc. Sport. Nutr. 2017, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Wallimann, T.; Tokarska-Schlattner, M.; Schlattner, U. The Creatine Kinase System and Pleiotropic Effects of Creatine. Amino Acids 2011, 40, 1271–1296. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M.; Spriet, L.L. Skeletal Muscle Energy Metabolism during Exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Jäger, R.; Purpura, M.; Shao, A.; Inoue, T.; Kreider, R.B. Analysis of the Efficacy, Safety, and Regulatory Status of Novel Forms of Creatine. Amino Acids 2011, 40, 1369–1383. [Google Scholar] [CrossRef]
- Guzun, R.; Timohhina, N.; Tepp, K.; Gonzalez-Granillo, M.; Shevchuk, I.; Chekulayev, V.; Kuznetsov, A.V.; Kaambre, T.; Saks, V.A. Systems Bioenergetics of Creatine Kinase Networks: Physiological Roles of Creatine and Phosphocreatine in Regulation of Cardiac Cell Function. Amino Acids 2011, 40, 1333–1348. [Google Scholar] [CrossRef]
- Northeast, B.; Clifford, T. The Effect of Creatine Supplementation on Markers of Exercise-Induced Muscle Damage: A Systematic Review and Meta-Analysis of Human Intervention Trials. Int. J. Sport. Nutr. Exerc. Metab. 2021, 31, 276–291. [Google Scholar] [CrossRef]
- Cordingley, D.M.; Cornish, S.M.; Candow, D.G. Anti-Inflammatory and Anti-Catabolic Effects of Creatine Supplementation: A Brief Review. Nutrients 2022, 14, 544. [Google Scholar] [CrossRef]
- Nomura, A.; Zhang, M.; Sakamoto, T.; Ishii, Y.; Morishima, Y.; Mochizuki, M.; Kimura, T.; Uchida, Y.; Sekizawa, K. Anti-Inflammatory Activity of Creatine Supplementation in Endothelial Cells in Vitro. Br. J. Pharmacol. 2003, 139, 715–720. [Google Scholar] [CrossRef]
- Khanna, N.K.; Madan, B.R. Studies on the Anti-Inflammatory Activity of Creatine. Arch. Int. Pharmacodyn. Ther. 1978, 231, 340–350. [Google Scholar]
- Ohtsuki, S.; Tachikawa, M.; Takanaga, H.; Shimizu, H.; Watanabe, M.; Hosoya, K.; Terasaki, T. The Blood-Brain Barrier Creatine Transporter Is a Major Pathway for Supplying Creatine to the Brain. J. Cereb. Blood Flow. Metab. 2002, 22, 1327–1335. [Google Scholar] [CrossRef]
- Béard, E.; Braissant, O. Synthesis and Transport of Creatine in the CNS: Importance for Cerebral Functions. J. Neurochem. 2010, 115, 297–313. [Google Scholar] [CrossRef]
- Pazini, F.L.; Cunha, M.P.; Rodrigues, A.L.S. The Possible Beneficial Effects of Creatine for the Management of Depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 193–206. [Google Scholar] [CrossRef]
- Ainsley Dean, P.J.; Arikan, G.; Opitz, B.; Sterr, A. Potential for Use of Creatine Supplementation Following Mild Traumatic Brain Injury. Concussion 2017, 2, CNC34. [Google Scholar] [CrossRef]
- Schwane, J.A.; Armstrong, R.B. Effect of Training on Skeletal Muscle Injury from Downhill Running in Rats. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 969–975. [Google Scholar] [CrossRef]
- Hessel, A.L.; Lindstedt, S.L.; Nishikawa, K.C. Physiological Mechanisms of Eccentric Contraction and Its Applications: A Role for the Giant Titin Protein. Front. Physiol. 2017, 8, 70. [Google Scholar] [CrossRef]
- Rouillon, J.; Zocevic, A.; Leger, T.; Garcia, C.; Camadro, J.M.; Udd, B.; Wong, B.; Servais, L.; Voit, T.; Svinartchouk, F. Proteomics Profiling of Urine Reveals Specific Titin Fragments as Biomarkers of Duchenne Muscular Dystrophy. Neuromuscul. Disord. 2014, 24, 563–573. [Google Scholar] [CrossRef]
- Kanda, K.; Sakuma, J.; Akimoto, T.; Kawakami, Y.; Suzuki, K. Detection of Titin Fragments in Urine in Response to Exercise-Induced Muscle Damage. PLoS ONE 2017, 12, e0181623. [Google Scholar] [CrossRef]
- Inami, T.; Yamaguchi, S.; Ishida, H.; Kohtake, N.; Morito, A.; Yamada, S.; Shimomasuda, M.; Haramoto, M.; Nagata, N.; Murayama, M. Changes in Muscle Shear Modulus and Urinary Titin N-Terminal Fragment after Eccentric Exercise. J. Sport. Sci. Med. 2022, 21, 536–544. [Google Scholar] [CrossRef]
- Hultman, E.; Söderlund, K.; Timmons, J.A.; Cederblad, G.; Greenhaff, P.L. Muscle Creatine Loading in Men. J. Appl. Physiol. 1996, 81, 232–237. [Google Scholar] [CrossRef]
- Ou, Z.; Dolmatova, E.; Lassegue, B.; Griendling, K.K. B1- And B2-Integrins: Central Players in Regulating Vascular Permeability and Leukocyte Recruitment during Acute Inflammation. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk, M.; Stege, H.; Grabbe, S.; Bros, M. B2 Integrins—Multi-Functional Leukocyte Receptors in Health and Disease. Int. J. Mol. Sci. 2020, 21, 1402. [Google Scholar] [CrossRef] [PubMed]
- Chase, S.D.; Magnani, J.L.; Simon, S.I. E-Selectin Ligands as Mechanosensitive Receptors on Neutrophils in Health and Disease. Ann. Biomed. Eng. 2012, 40, 849–859. [Google Scholar] [CrossRef] [PubMed]
- Blythe, E.N.; Weaver, L.C.; Brown, A.; Dekaban, G.A. B2 Integrin CD11d/CD18: From Expression to an Emerging Role in Staged Leukocyte Migration. Front. Immunol. 2021, 12, 775447. [Google Scholar] [CrossRef]
- Hamada, K.; Vannier, E.; Sacheck, J.M.; Witsell, A.L.; Roubenoff, R. Senescence of Human Skeletal Muscle Impairs the Local Inflammatory Cytokine Response to Acute Eccentric Exercise. FASEB J. 2005, 19, 1–19. [Google Scholar] [CrossRef]
- Peake, J.M.; Neubauer, X.O.; della Gatta, P.A.; Nosaka, K. Muscle Damage and Inflammation during Recovery from Exercise. J. Appl. Physiol. 2017, 122, 559–570. [Google Scholar] [CrossRef]
- Pizza, F.X.; Mcloughlin, T.J.; Mcgregor, S.J.; Calomeni, E.P.; Gunning, W.T. Neutrophils Injure Cultured Skeletal Myotubes. Am. J. Physiol. Cell. Physiol. 2001, 281, 335–341. [Google Scholar] [CrossRef]
- Marceau, F.; Bachelard, H.; Bouthillier, J.; Fortin, J.P.; Morissette, G.; Bawolak, M.T.; Charest-Morin, X.; Gera, L. Bradykinin Receptors: Agonists, Antagonists, Expression, Signaling, and Adaptation to Sustained Stimulation. Int. Immunopharmacol. 2020, 82, 106305. [Google Scholar] [CrossRef]
- Ni, A.; Yin, H.; Agata, J.; Yang, Z.; Chao, L.; Chao, J. Overexpression of Kinin B1 Receptors Induces Hypertensive Response to Des-Arg9-Bradykinin and Susceptibility to Inflammation. J. Biol. Chem. 2003, 278, 219–225. [Google Scholar] [CrossRef]
- Todorov, A.G.; Andrade, D.; Pesquero, J.B.; Araujo, R.d.C.; Bader, M.; Stewart, J.; Gera, L.; Müller-Esterl, W.; Morandi, V.; Goldenberg, R.C.S.; et al. Trypanosoma Cruzi Induces Edematogenic Responses in Mice and Invades Cardiomyocytes and Endothelial Cells in Vitro by Activating Distinct Kinin Receptor (B1/B2) Subtypes. FASEB J. 2003, 17, 73–75. [Google Scholar] [CrossRef]
- Bortone, F.; Santos, H.A.; Albertini, R.; Pesquero, J.B.; Costa, M.S.; Silva, J.A. Low Level Laser Therapy Modulates Kinin Receptors mRNA Expression in the Subplantar Muscle of Rat Paw Subjected to Carrageenan-Induced Inflammation. Int. Immunopharmacol. 2008, 8, 206–210. [Google Scholar] [CrossRef]
- Mizumura, K.; Taguchi, T. Delayed Onset Muscle Soreness: Involvement of Neurotrophic Factors. J. Physiol. Sci. 2016, 66, 43–52. [Google Scholar] [CrossRef]
- Nagahisa, H.; Ikezaki, K.; Yamada, R.; Yamada, T.; Miyata, H. Preconditioning Contractions Suppress Muscle Pain Markers after Damaging Eccentric Contractions. Pain. Res. Manag. 2018, 2018, 3080715. [Google Scholar] [CrossRef]
- Carmichael, M.D.; Davis, J.M.; Murphy, E.A.; Carson, J.A.; van Rooijen, N.; Mayer, E.; Ghaffar, A. Role of Brain Macrophages on IL-1β and Fatigue Following Eccentric Exercise-Induced Muscle Damage. Brain Behav. Immun. 2010, 24, 564–568. [Google Scholar] [CrossRef]
- Serna-Rodríguez, M.F.; Bernal-Vega, S.; de la Barquera, J.A.O.S.; Camacho-Morales, A.; Pérez-Maya, A. The Role of Damage Associated Molecular Pattern Molecules (DAMPs) and Permeability of the Blood-Brain Barrier in Depression and Neuroinflammation. J. Neuroimmunol. 2022, 371, 577951. [Google Scholar] [CrossRef]
- Leland, K.M.; McDonald, T.L.; Drescher, K.M. Effect of Creatine, Creatinine, and Creatine Ethyl Ester on TLR Expression in Macrophages. Int. Immunopharmacol. 2011, 11, 1341–1347. [Google Scholar] [CrossRef]
- Damm, J.; Wiegand, F.; Harden, L.M.; Gerstberger, R.; Rummel, C.; Roth, J. Fever, Sickness Behavior, and Expression of Inflammatory Genes in the Hypothalamus after Systemic and Localized Subcutaneous Stimulation of Rats with the Toll-like Receptor 7 Agonist Imiquimod. Neuroscience 2012, 201, 166–183. [Google Scholar] [CrossRef]
- Harden, L.M.; du Plessis, I.; Roth, J.; Loram, L.C.; Poole, S.; Laburn, H.P. Differences in the Relative Involvement of Peripherally Released Interleukin (IL)-6, Brain IL-1β and Prostanoids in Mediating Lipopolysaccharide-Induced Fever and Sickness Behavior. Psychoneuroendocrinology 2011, 36, 608–622. [Google Scholar] [CrossRef]
- Pereira, B.C.; da Rocha, A.L.; Pauli, J.R.; Ropelle, E.R.; de Souza, C.T.; Cintra, D.E.; Sant’Ana, M.R.; da Silva, A.S.R. Excessive Eccentric Exercise Leads to Transitory Hypothalamic Inflammation, which May Contribute to the Low Body Weight Gain and Food Intake in Overtrained Mice. Neuroscience 2015, 311, 231–242. [Google Scholar] [CrossRef]
- Brancaccio, P.; Lippi, G.; Maffulli, N. Biochemical Markers of Muscular Damage. Clin. Chem. Lab. Med. 2010, 48, 757–767. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Diets (g/kg Diet) | |
|---|---|---|
| Control Diet | Creatine Diet | |
| Casein | 200 | 200 |
| L-Cystine | 3 | 3 |
| Cornstarch | 397.486 | 397.486 |
| α-Cornstarch | 132 | 132 |
| Sucrose | 100 | 100 |
| Cellulose | 50 | ― |
| Creatine monohydrate | ― | 50 |
| Soybean oil | 70 | 70 |
| t-butylhydroquinone | 0.014 | 0.014 |
| Mineral Mix (AIN93G) | 35 | 35 |
| Vitamin Mix (AIN93) | 10 | 10 |
| Choline bitartrate | 2.5 | 2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yokota, Y.; Yamada, S.; Yamamoto, D.; Kato, K.; Morito, A.; Takaoka, A. Creatine Supplementation Alleviates Fatigue after Exercise through Anti-Inflammatory Action in Skeletal Muscle and Brain. Nutraceuticals 2023, 3, 234-249. https://doi.org/10.3390/nutraceuticals3020019
Yokota Y, Yamada S, Yamamoto D, Kato K, Morito A, Takaoka A. Creatine Supplementation Alleviates Fatigue after Exercise through Anti-Inflammatory Action in Skeletal Muscle and Brain. Nutraceuticals. 2023; 3(2):234-249. https://doi.org/10.3390/nutraceuticals3020019
Chicago/Turabian StyleYokota, Yuma, Satoshi Yamada, Daisuke Yamamoto, Keita Kato, Akihisa Morito, and Akiko Takaoka. 2023. "Creatine Supplementation Alleviates Fatigue after Exercise through Anti-Inflammatory Action in Skeletal Muscle and Brain" Nutraceuticals 3, no. 2: 234-249. https://doi.org/10.3390/nutraceuticals3020019
APA StyleYokota, Y., Yamada, S., Yamamoto, D., Kato, K., Morito, A., & Takaoka, A. (2023). Creatine Supplementation Alleviates Fatigue after Exercise through Anti-Inflammatory Action in Skeletal Muscle and Brain. Nutraceuticals, 3(2), 234-249. https://doi.org/10.3390/nutraceuticals3020019