Quantitative RT-PCR Detection of Hepatitis A Virus, Rotaviruses and Enteroviruses in the Buffalo River and Source Water Dams in the Eastern Cape Province of South Africa


Abstract
:1. Introduction
2. Experimental Section
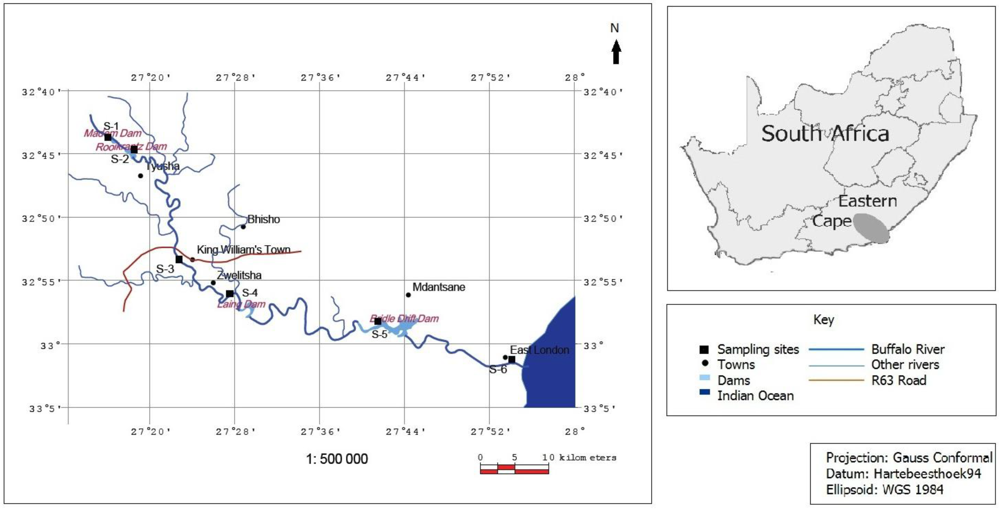
2.1. Study Area and Samples Collection
2.2. Concentration of Viruses in Water
2.3. Quantification of Viruses

{kind=link}
{kind=link}
{kind=link}
| Enteric virus | Primers and labelled TaqMan probe | Reference | Control virus |
|---|---|---|---|
| Hepatitis A virus | HAV68 (F): 5′-TCA CCG CCG TTT GCC TAG-3′ | [56] | ATCC VR-1357; Strain PA21 |
| HAV240 (R): 5′-GGA GAG CCC TGG AAG AAA G-3′ | |||
| HAV150 (P): 5′-FAM-CCT GAA CCT GCA GGA ATT AA-MGBNFQ-3′ | |||
| Rotaviruses | JVK (F): 5′-CAGTGGTTGATGCTCAAGATGGA-3′ | [59] | ATCC VR-2274; Strain 248 |
| JVK (R): 5′-TCATTGTAATCATATTGAATACCCA-3′ | |||
| JVK (P): 5′-FAM-ACAACTGCAGCTTCAAAAGAAGWGT-MGBNFQ-3′ | |||
| Enteroviruses | EV1 (F): 5′-CCCTGAATGCGGCTAAT-3′ | [57] | Coxsackie virus A2 (ATCC VR-1550; Strain FLEETWOOD) |
| EV1 (R): 5′-TGTCACCATA AGCAGCCA-3′ | |||
| EV-BHQ (P): 5′-FAM-ACGGACACCCAAAGTAGTCGGTTC-MGBNFQ-3′ |
2.3.1. RNA Extraction
2.3.2. Reverse-Transcription
2.3.3. Quantitative PCR (qPCR)
| Virus | Taq activation | 45 cycles of: | ||
|---|---|---|---|---|
| Denaturation | Annealing | Extension | ||
| Hepatitis A virus | 10 min at 95 °C | 15 s at 95 °C | 1 min at 60 °C | 1 min at 70 °C |
| Rotaviruses | 15 min at 95 °C | 15 s at 95 °C | 30 s at 55 °C | 30 s at 72 °C |
| Enteroviruses | 10 min at 95 °C | 15 s at 94 °C | 1 min at 58 °C | 20 s at 72 °C |
2.3.4. The Standard Curve
2.4. Quality Control
2.5. Statistical Analysis
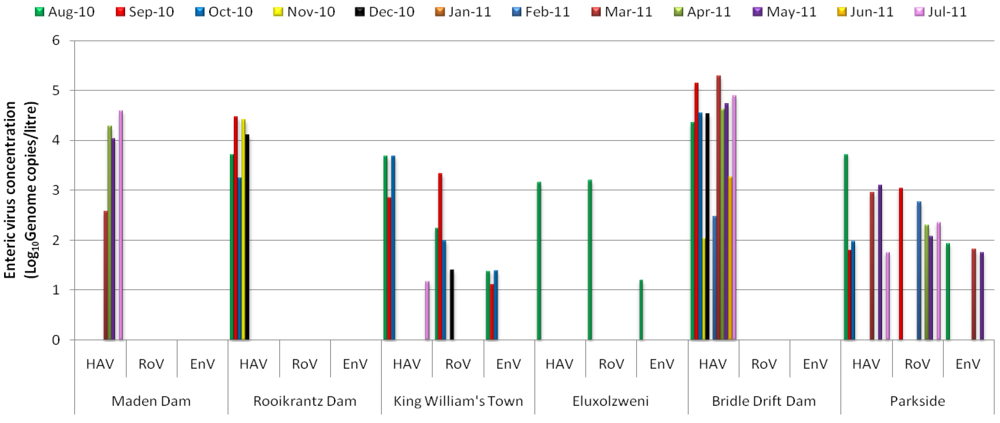
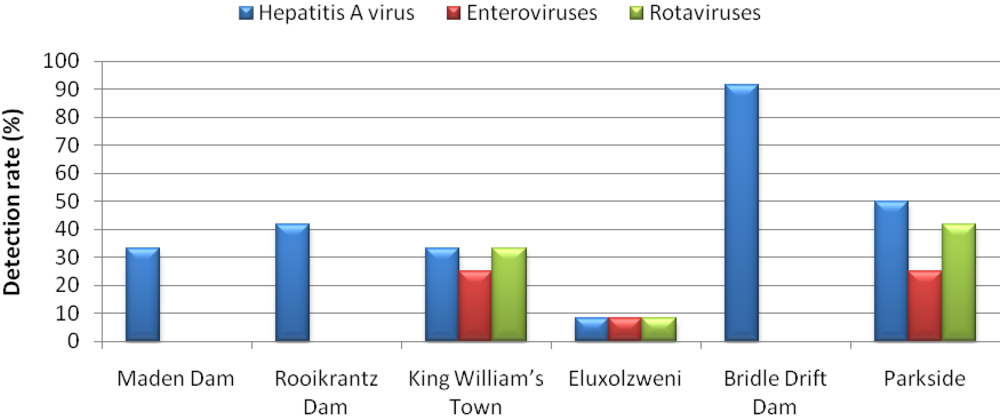
3. Results and Discussion

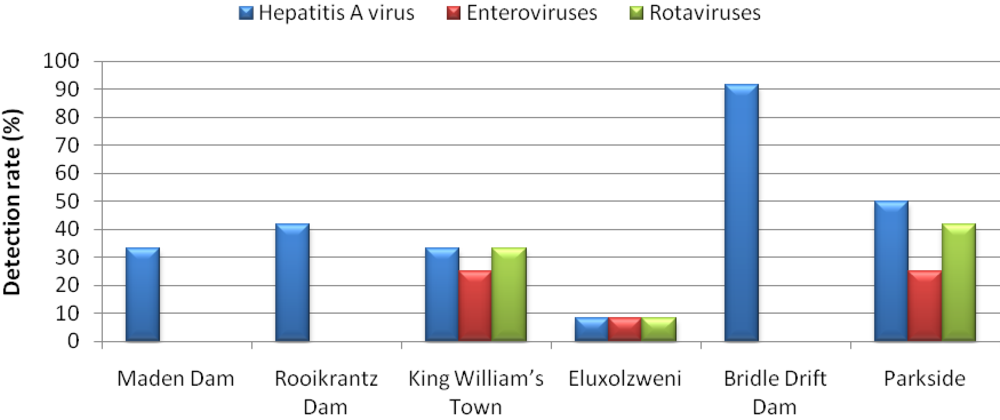
4. Conclusions
Acknowledgments
Conflict of Interest
References
- World Health Organization (WHO), World Health Statistics 2009; WHO Press: Geneva, Switzerland, 2009; pp. 1–149.
- Gibson, K.E.; Schwab, K.J. Detection of bacterial indicators and human and bovine enteric viruses in surface water and groundwater sources potentially impacted by animal and human wastes in Lower Yakima Valley, Washington. Appl. Environ. Microbiol. 2011, 77, 355–362. [Google Scholar] [CrossRef]
- Pruss, A.; Kay, D.; Fewtrell, L.; Bartram, J. Estimating the burden of disease from water sanitation and hygiene at a global level. Environ. Health Perspect. 2002, 110, 537–542. [Google Scholar] [CrossRef]
- Boschi-Pinto, C.; Velebit, L.; Shibuya, K. Estimating child mortality due to diarrhoea in developing countries. Bull. World Health Organ. 2008, 86, 710–717. [Google Scholar] [CrossRef]
- World Health Organization (WHO) and UNICEF, Progress on Sanitation and Drinking-Water: 2010 Update; WHO Press: Geneva, Switzerland, 2010; pp. 1–60.
- Muller, M.; Schreiner, B.; Smith, L.; van Koppen, B.; Sally, H.; Aliber, M.; Cousins, B.; Tapela, B.; van der Merwe-Botha, M.; Karar, E.; Pietersen, K. Water Security in South Africa; Development Planning Division DBSA: Midrand, South Africa, 2009. [Google Scholar]
- Shuval, H.I. Wastewater Irrigation in Developing Countries: Health Effects and Technical Solutions; The World Bank: Washinton, DC, USA, 1990; pp. 1–19. [Google Scholar]
- Okoh, A.I.; Sibanda, T.; Gusha, S.S. Inadequately treated wastewater as a source of human enteric viruses in the environment. Int. J. Environ. Res. Pub. Health 2010, 7, 2620–2637. [Google Scholar] [CrossRef]
- Chigor, V.N.; Umoh, V.J.; Okuofu, C.A.; Ameh, J.B.; Igbinosa, E.O.; Okoh, A.I. Water quality assessment: Surface water sources used for drinking and irrigation in Zaria, Nigeria are a public health hazard. Environ. Monit. Assess. 2012, 184, 3389–3400. [Google Scholar]
- Miagostovich, M.P.; Ferreira, F.F.M.; Guimaraes, F.R.; Fumian, T.M.; Diniz-Mendes, L.; Luz, S.L.B.; Silva, L.A.; Leite, J.P.G. Molecular detection and characterization of gastroenteritis viruses occurring naturally in the stream waters of Manaus, central Amazonia, Brazil. Appl. Environ. Microbiol. 2008, 74, 375–382. [Google Scholar] [CrossRef]
- Bosch, A.; Guix, S.; Sano, D.; Pinto, R.M. New tools for the study and direct surveillance of viral pathogens in water. Curr. Op. Biotechnol. 2008, 19, 1–7. [Google Scholar] [CrossRef]
- Fong, T.T.; Lipp, E.K. Enteric viruses of human and animals in aquatic environments: Heath risks, detection and potential water quality assessment tools. Appl. Environ. Microbiol. 2005, 69, 357–371. [Google Scholar]
- Haramoto, E.; Katayama, H.; Utagawa, E.; Ohgaki, S. Development of sample storage methods for detecting enteric viruses in environmental water. J. Virol. Methods 2008, 151, 1–6. [Google Scholar] [CrossRef]
- Bosch, A. Human enteric viruses in the water environment: A minireview. Int. Microbiol. 1998, 1, 191–196. [Google Scholar]
- Fauquet, C.M.; Mayo, M.A.; Maniloff, J.; Desselberger, U.; Ball, L.A. Virus Taxonomy. Eighth Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2005; pp. 853–857. [Google Scholar]
- Ward, R.L.; Bernstein, D.I.; Young, E.C.; Sherwood, J.R.; Knowlton, D.R.; Schiff, G.M. Human rotavirus studies in volunteers: Determination of infectious dose and serological response to infection. J. Infect. Dis. 1986, 154, 871–880. [Google Scholar] [CrossRef]
- Pusch, D.; Oh, D.Y.; Wolf, S.; Dumke, R.; Schroter-Bobsin, U.; Hohne, M.; Roske, I.; Schreier, E. Detection of enteric viruses and bacterial indicators in German environmental waters. Arch. Virol. 2005, 150, 929–947. [Google Scholar]
- Liu, J.; Kibiki, G.; Maro, V.; Maro, A.; Kumburu, H.; Swai, N.; Taniuchi, M.; Gratz, J.; Toney, D.; Kang, G.; Houpt, E. Multiplex reverse transcription PCR Luminex assay for detection and quantitation of viral agents of gastroenteritis. J. Clin. Virol. 2011, 50, 308–313. [Google Scholar]
- Kittigul, L.; Uthaisin, A.; Ekchaloemkiet, S.; Utrarachkij, F.; Luksamijarulkul, P. Detection and characterization of hepatitis A virus in water samples in Thailand. J. Appl. Microbiol. 2006, 100, 1318–1323. [Google Scholar] [CrossRef]
- Colbere-Garapin, F.; Martin-Latil, S.; Blondel, B.; Mousson, L.; Pelletier, I.; Autret, A.; Francois, A.; Niborski, V.; Grompone, G.; Catonnet, G.; van de Moer, A. Prevention and treatment of enteric viral infections: Possible benefits of probiotic bacteria. Microb. Infect. 2007, 9, 1623–1631. [Google Scholar] [CrossRef]
- Roque-Afonso, A.M.; Desbois, D.; Dussaix, E. Hepatitis A virus: Serology and molecular diagnostics. Future Virol. 2010, 5, 233–242. [Google Scholar] [CrossRef]
- Taylor, M.B.; Cox, N.; Very, M.A.; Grabow, W.O.K. The occurrence of hepatitis A and astroviruses in selected river and dam waters in South Africa. Water Res. 2001, 35, 2653–2660. [Google Scholar]
- Venter, J.M.E.; van Heerden, J.; Vivier, J.C.; Grabow, W.O.K.; Taylor, M.B. Hepatitis A virus in surface water in South Africa: What are the risks? J. Water Health 2007, 5, 229–240. [Google Scholar]
- MacIntyre, U.E.; de Villiers, F.P.R. The Economic burden of diarrheal disease in a tertiary level hospital, Gauteng, South Africa. J. Infect. Dis. 2010, 202, S116–S125. [Google Scholar] [CrossRef]
- Mwenda, J.M.; Ntoto, K.M.; Abebe, A.; Enweronu-Laryea, C.; Amina, I.; Mchomvu, J.; Kisakye, A.; Mpabalwani, E.M.; Pazvakavambwa, I.; Armah, G.E.; Seheri, L.M.; Kiulia, N.M.; Page, N.; Widdowson, M.A.; Steele, A.D. Burden and epidemiology of rotavirus diarrhea in selected African countries: preliminary results from the African Rotavirus Surveillance Network. J. Infect. Dis. 2010, 202, S5–S11. [Google Scholar]
- Patel, M.M.; Steele, D.; Gentsch, J.R.; Wecker, J.; Glass, R.I.; Parashar, U.D. Real-world impact of rotavirus vaccination. Pediat. Infect. Dis. J. 2011, 30, S1–S5. [Google Scholar]
- Ramachandran, M.; Gentsch, J.R.; Parashar, U.D.; Jin, S.; Woods, P.A.; Holmes, J.L.; Kirkwood, C.D.; Bishop, R.F.; Greenberg, H.B.; Urasawa, S.; Gerna, G.; Coulson, B.S.; Taniguchi, K.; Bresee, J.S.; Glass, R.I. Detection and characterization of novel rotavirus strains in the United States. J. Clin. Microbiol. 1998, 36, 3223–3229. [Google Scholar]
- Adah, M.; Wade, A.; Taniguchi, K. Molecular epidemiology of rotaviruses in Nigeria: Detection of unusual strains with G2P [6] and G8P [1] specificities. J. Clin. Microbiol. 2001, 39, 3969–3975. [Google Scholar] [CrossRef]
- Pinto, R.M.; Saiz, J.C. Enteric Hepatitis Viruses. In Human Viruses in Water; Bosch, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 39–67. [Google Scholar]
- Tallon, L.A.; Love, D.C.; Moore, Z.S.; Sobsey, M.D. Recovery and sequence analysis of hepatitis A virus from springwater implicated in an outbreak of acute viral hepatitis. Appl. Environ. Microbiol. 2008, 74, 6158–6160. [Google Scholar] [CrossRef]
- Taylor, M.B.; Schildhauer, C.I.; Parker, S.; Grabow, W.O.; Xi, J.; Estes, M.K. Two successive outbreaks of SRSV-associated gastroenteritis in South Africa. J. Med Virol. 1993, 41, 18–23. [Google Scholar] [CrossRef]
- Mans, J.; de Villiers, J.C.; du Plessis, N.M.; Avenant, T.; Taylor, M.B. Emerging norovirus GII.4 2008 variant detected in hospitalised paediatric patients in South Africa. J. Clin. Virol. 2010, 49, 258–264. [Google Scholar] [CrossRef]
- Rinaldi, M.; Brierley, E.; Bekker, A. Donor breastmilk saved infant lives during an outbreak of rotavirus in South Africa. Breastfeeding Medicine 2009, 4, 133–134. [Google Scholar] [CrossRef]
- Griffin, D.W.; Gibson, C.J.III; Lipp, E.K.; Riley, K.; Paul, J.H.III; Rose, J.B. Detection of viral pathogens by reverse transcriptase PCR and of microbial indicators by standard methods in the canals of the Florida Keys. Appl. Environ. Microbiol. 1999, 65, 4118–4125. [Google Scholar]
- Grabow, W.O.K.; Taylor, M.B.; de Villiers, J.C. New methods for the detection of viruses: Call for review of drinking water quality guidelines. Water Sci. Technol. 2001, 43, 1–8. [Google Scholar]
- Pina, S.; Buti, M.; Jardi, R.; Clemente-Casares, P.; Jofre, J.; Girones, R. Genetic analysis of hepatitis A virus strains recovered from the environment and from patients with acute hepatitis. J. Gen. Virol. 2001, 82, 2955–2963. [Google Scholar]
- Castello, A.A.; Arguelles, M.H.; Rota, R.P.; Olthoff, A.; Jiang, B.; Glass, R.I.; Gentsch, J.R.; Glikmann, G. Molecular epidemiology of Group A rotavirus diarrhea among children in Buenos Aires, Argentina, from 1999 to 2003 and emergence of the infrequent genotype G12. J. Clin. Microbiol. 2006, 44, 2046–2050. [Google Scholar]
- Gersberg, R.M.; Rose, M.A.; Robles-Sikisaka, R.; Dhar, A.K. Quantitative detection of hepatitis A virus and enteroviruses near the United States-Mexico border and correlation with levels of fecal indicator bacteria. Appl. Environ. Microbiol. 2006, 72, 7438–7444. [Google Scholar] [CrossRef]
- Meleg, E.; Banyai, K.; martella, V.; Jiang, B.; Kocsis, B.; Kisfali, P.; Melegh, B.; Szucs, G. Detection and quantification of group C rotaviruses in communal sewage. Appl. Environ. Microbiol. 2008, 74, 3394–3399. [Google Scholar]
- Cunliffe, N.A.; Ngwira, B.M.; Dove, W.; Nakagomi, O.; Nakagomi, T.; Perez, A.; Hart, C.A.; Kazembe, P.N.; Mwansambo, C.V. Serotype G12 rotaviruses, Lilongwe, Malawi. Emerg. Infect. Dis. 2009, 15, 87–90. [Google Scholar]
- Kiulia, N.M.; Netshikweta, R.N.A.; van Zyl, W.B.; Kiraithe, M.M.; Nyachieo, A.; Mwenda, J.M.; Taylor, M.B. The detection of enteric viruses in selected urban and rural river water and sewage in Kenya, with special reference to rotaviruses. J. Appl. Microbiol. 2010, 109, 818–828. [Google Scholar] [CrossRef]
- Grabow, W.O.K.; Taylor, M.B.; Wolfaardt, M. Research on Human Viruses in Diffuse Effluents and Related Water Environments; Water Research Commission: Pretoria, South Africa, 1996; pp. 1–25. [Google Scholar]
- Vivier, J.C.; Clay, C.G.; Grabow, W.O. Detection and rapid differentiation of human enteroviruses in water sources by restriction enzyme analysis. Water Sci. Technol. 2001, 4, 209–212. [Google Scholar]
- Pavlov, D.N. Poliovirus vaccine strains in sewage and river water in South Africa. Can. J. Microbiol. 2006, 52, 717–723. [Google Scholar] [CrossRef]
- Nadan, S.; Walter, J.E.; Grabow, W.O.K.; Mitchell, D.K.; Taylor, M.B. Molecular characterization of astroviruses by reverse transcriptase PCR and sequence analysis: Comparison of clinical and environmental isolates from South Africa. Appl. Environ. Microbiol. 2003, 69, 747–753. [Google Scholar] [CrossRef]
- Genthe, B.; Idema, G.K.; Kfir, R.; Grabow, W.O.K. Detection of rotavirus in South African waters: A comparison of a cytoimmunolabelling technique with commercially available immunoassays. Water Sci. Technol. 1991, 24, 241–244. [Google Scholar]
- van Zyl, W.B.; Williams, P.J.; Grabow, W.O.; Taylor, M.B. Application of a molecular method for the detection of group A rotaviruses in raw and treated water. Water Sci. Technol. 2004, 50, 223–228. [Google Scholar]
- Pourcher, A.M.; Picard-Bonnaud, F.P.B.; Agnieszka, V.F.; Vasilica, S.; Gerard, M. Survival of faecal indicators and enteroviruses in soil after land-spreading of municipal sewage sludge. Appl. Soil Ecol. 2007, 35, 473–479. [Google Scholar] [CrossRef]
- Muscillo, M.; Pourshaban, M.; Iaconelli, M.; Fontana, S.; Di Grazia, A.; Manzara, S.; Fadda, G.; Santangelo, R.; La Rosa, G. Detection and quantification of human adenoviruses in surface waters by nested PCR, TaqMan real-time PCR and cell culture assays. Water Air Soil Pollut. 2008, 191, 1–11. [Google Scholar] [CrossRef]
- Jurzik, L.; Hamza, I.A.; Puchert, W.; Überla, K.; Wilhelm, M. Chemical and microbiological parameters as possible indicators for human enteric viruses in surface water. Int. J. Hygi. Environ. Health 2010, 213, 210–216. [Google Scholar] [CrossRef]
- Dahling, D. Detection and enumeration of enteric viruses in cell culture. CRC Rev. Environ. Contamin. 1991, 21, 237–263. [Google Scholar]
- Schvoerer, E.; Bonneta, F.; Duboisa, V.; Cazauxb, G.; Serceauc, R.; Fleurya, H.J.A.; Lafona, M.E. PCR detection of human enteric viruses in bathing areas, waste waters and human stools in southwestern France. Res. Microbiol. 2000, 151, 693–701. [Google Scholar] [CrossRef]
- Griffin, D.W.; Donalson, K.A.; Paul, J.H.; Rose, J.B. Pathogenic human viruses in coastal waters. Clin.Microbiol. Rev. 2003, 16, 129–143. [Google Scholar] [CrossRef]
- de Paula, V.S.; Diniz-Mendes, L.; Villar, L.M.; Luz, S.L.B.; Silva, L.A.; Jesus, M.S.; da Silva, M.N.V.S.; Gaspar, A.S. Hepatitis A in environmental water samples from the Amazon Basin. Water Res. 2007, 41, 1169–1176. [Google Scholar] [CrossRef]
- Rodríguez, R.A.; Pepper, I.L.; Gerba, C.P. Application of PCR-based methods to assess the infectivity of enteric viruses in environmental samples. Appl. Environ. Microbiol. 2009, 75, 297–307. [Google Scholar] [CrossRef]
- Costafreda, M.I.; Bosch, A.; Pinto, R.M. Development, evaluation, and standardization of a real-time TaqMan reverse transcription-PCR assay for quantification of hepatitis A virus in clinical and shellfish samples. Appl. Environ. Microbiol. 2006, 72, 3846–3855. [Google Scholar] [CrossRef]
- Gregory, J.B.; Litaker, R.W.; Noble, R.T. Rapid one-step quantitative reverse transcriptase PCR Assay with competitive internal positive control for detection of enteroviruses in environmental samples. Appl. Environ. Microbiol. 2006, 72, 3960–3967. [Google Scholar] [CrossRef]
- Haramoto, E.; Katayama, H.; Oguma, K.; Ohgaki, S. Recovery of viral genomes in water by virus concentration methods. J. Virol. Methods 2007, 142, 169–173. [Google Scholar] [CrossRef]
- Jothikumar, N.; Kang, G.; Hill, V.R. Broadly reactive TaqMan assay for real-time RT-PCR detection of rotavirus in clinical and environmental samples. J. Virol. Methods 2009, 155, 126–131. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, H.; Yang, J.; Jiang, S.; Cai1, B. Detection and quantification of enteroviruses in coastal seawaters from Bohai Bay, Tianjin, China. J. Environ. Sci. 2010, 22, 150–154. [Google Scholar] [CrossRef]
- Hamza, I.A.; Jurzik, L.; Stang, A.; Sure, K.; Uberla, K.; Wilhelm, M. Detection of human viruses in rivers of a densly-populated area in Germany using a virus adsorption elution method optimized for PCR analyses. Water Res. 2009, 43, 2657–2668. [Google Scholar] [CrossRef]
- Bofill-Mas, S.; Calgua, B.; Clemente-Casares, P.; La Rosa, G.; Iaconelli, M.; Muscillo, M.; Rutjes, S.; de Roda Husman, A.M.; Grunert, A.; Graber, I.; Verani, M.; Carducci, A.; Calvo, M.; Wyn-Jones, P.; Girones, R. Quantification of human adenoviruses in European recreational waters. Food Environ. Virol. 2010, 2, 101–109. [Google Scholar] [CrossRef]
- Donia, D.; Bonanni, E.; Diaco, L.; Divizia, M. Statistical correlation between enterovirus genome copy numbers and infectious viral particles in wastewater samples. Lett. Appl. Microbiol. 2010, 50, 237–240. [Google Scholar] [CrossRef]
- Parshionikar, S.; Laseke, I.; Fout, G.S. Use of propidium monoazide in reverse transcriptase PCR to distinguish between infectious and noninfectious enteric viruses in water samples. Appl. Environ. Microbiol. 2010, 76, 4318–4326. [Google Scholar] [CrossRef]
- River Health Programme (RHP), State-of-Rivers Report: Buffalo River System; Department of Water Affairs and Forestry: Pretoria, South Africa, 2004; pp. 1–41.
- American Public Health Association (APHA), Standard Methods for the Examination of Water and Wastewater, 20t edAPHA: Washington DC, USA, 1998.
- Haramoto, E.; Katayama, H.; Oguma, K.; Ohgaki, S. Application of cation-coated filters method to detection of noroviruses, enteroviruses, adenoviruses and torque teno viruses in the Tamagawa River in Japan. Appl. Environ. Microbiol. 2005, 71, 2403–2411. [Google Scholar] [CrossRef]
- Department of Water Affairs (DWAF), 2012 Green Drop Progress Report; Department of Water Affairs: Pretoria, South Africa, 2012.
- Arankalle, V.A.; Ramakrishnan, J. Simian hepatitis A virus derived from a captive rhesus monkey in India is similar to the strain isolated from wild African green monkeys in Kenya. J. Viral Hepati. 2009, 16, 214–218. [Google Scholar] [CrossRef]
- Amado, L.A.; Marchevsky, R.S.; de Paula, V.S.; Hooper, C.; Freire, M.D.S.; Gaspar, A.M.C.; Pinto, M.A. Experimental hepatitis A virus (HAV) infection in cynomolgus monkeys Macaca fascicularis: Evidence of active extrahepatic site of HAV replication. Int. J. Exp. Pathol. 2010, 91, 87–97. [Google Scholar] [CrossRef]
- Global Alert Response (GAR): Hepatitis A, 2012. Available online: http://www.who.int/csr/disease/hepatitis/whocdscsredc2007/en/index3.html (accessed on 9 October 2012).
- Lodder, W.J.; van den Berg, H.H.J.L.; Rutjes, S.A.; de Roda Husman, A.M. Presence of enteric viruses in source waters for drinking water production in the Netherlands. Appl. Environ Microbiol. 2010, 76, 5965–5971. [Google Scholar] [CrossRef]
- Steele, A.D.; Glass, R. Rotavirus in South Africa: From discovery to vaccine introduction. South Afri. J. Epidemiol.Infect. 2011, 26, 184–190. [Google Scholar]
- Ehlers, M.M.; Grabow, W.O.K.; Pavlov, D.N. Detection of enteroviruses in untreated and treated drinking water supplies in South Africa. Water Res. 2005, 39, 2253–2258. [Google Scholar] [CrossRef]
- Harvala, H.; McIntyre, C.L.; Imai, N.; Clasper, L.; Djoko, C.F.; Lebreton, M.; Vermeulen, M.; Saville, A.; Mutapi, F.; Tamoufe, U.; et al. High seroprevalence of enterovirus infections in apes and old world monkeys. Emerg. Infect. Dis. 2012, 18, 283–286. [Google Scholar] [CrossRef]
- Chen, C.H.; Hsu, B.M.; Wan, M.T. Detection of enteroviruses within brackish water from the Damshui River watershed, Taiwan. J. Environ. Engineer. 2008, 134, 486. [Google Scholar] [CrossRef]
- Skraber, S.; Gassilloud, B.; Schwartzbrod, L.; Gantzer, C. Survival of infectious Poliovirus-1 in river water compared to the persistence of somatic coliphages, thermotolerant coliforms, and poliovirus-1 genome. Water Res. 2004, 38, 2927–2933. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chigor, V.N.; Okoh, A.I. Quantitative RT-PCR Detection of Hepatitis A Virus, Rotaviruses and Enteroviruses in the Buffalo River and Source Water Dams in the Eastern Cape Province of South Africa. Int. J. Environ. Res. Public Health 2012, 9, 4017-4032. https://doi.org/10.3390/ijerph9114017
Chigor VN, Okoh AI. Quantitative RT-PCR Detection of Hepatitis A Virus, Rotaviruses and Enteroviruses in the Buffalo River and Source Water Dams in the Eastern Cape Province of South Africa. International Journal of Environmental Research and Public Health. 2012; 9(11):4017-4032. https://doi.org/10.3390/ijerph9114017
Chicago/Turabian StyleChigor, Vincent Nnamdigadi, and Anthony Ifeanyi Okoh. 2012. "Quantitative RT-PCR Detection of Hepatitis A Virus, Rotaviruses and Enteroviruses in the Buffalo River and Source Water Dams in the Eastern Cape Province of South Africa" International Journal of Environmental Research and Public Health 9, no. 11: 4017-4032. https://doi.org/10.3390/ijerph9114017
APA StyleChigor, V. N., & Okoh, A. I. (2012). Quantitative RT-PCR Detection of Hepatitis A Virus, Rotaviruses and Enteroviruses in the Buffalo River and Source Water Dams in the Eastern Cape Province of South Africa. International Journal of Environmental Research and Public Health, 9(11), 4017-4032. https://doi.org/10.3390/ijerph9114017