Forage as a Primary Source of Mycotoxins in Animal Diets


Abstract
:1. Introduction
2. Experimental Section
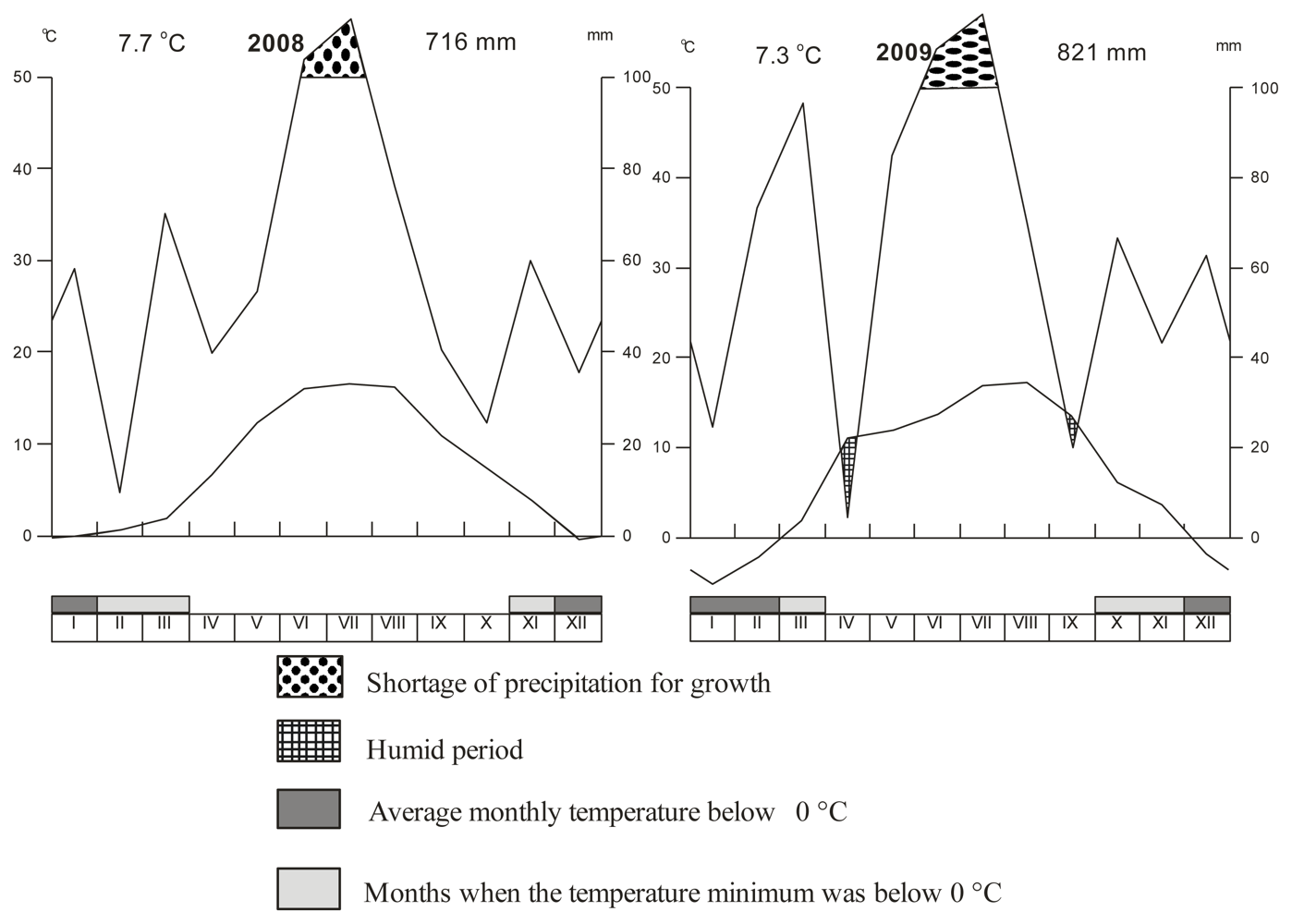
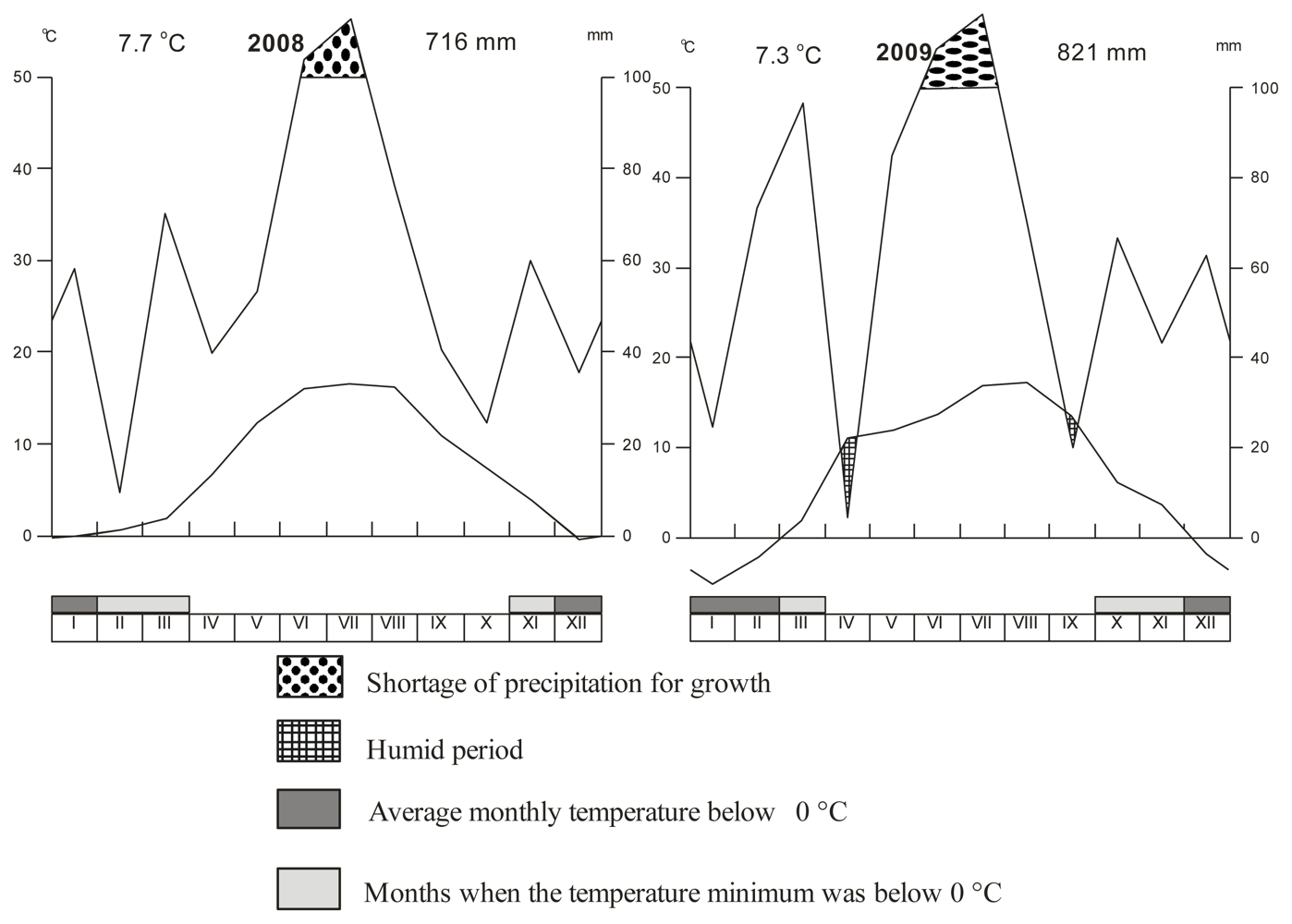
2.1. Experimental Localation
2.2. Experimental Design
2.3. Detected Parameters
2.4. Statistical Analyses
3. Results and Discussion
3.1. Characterization of Species Included in the Study
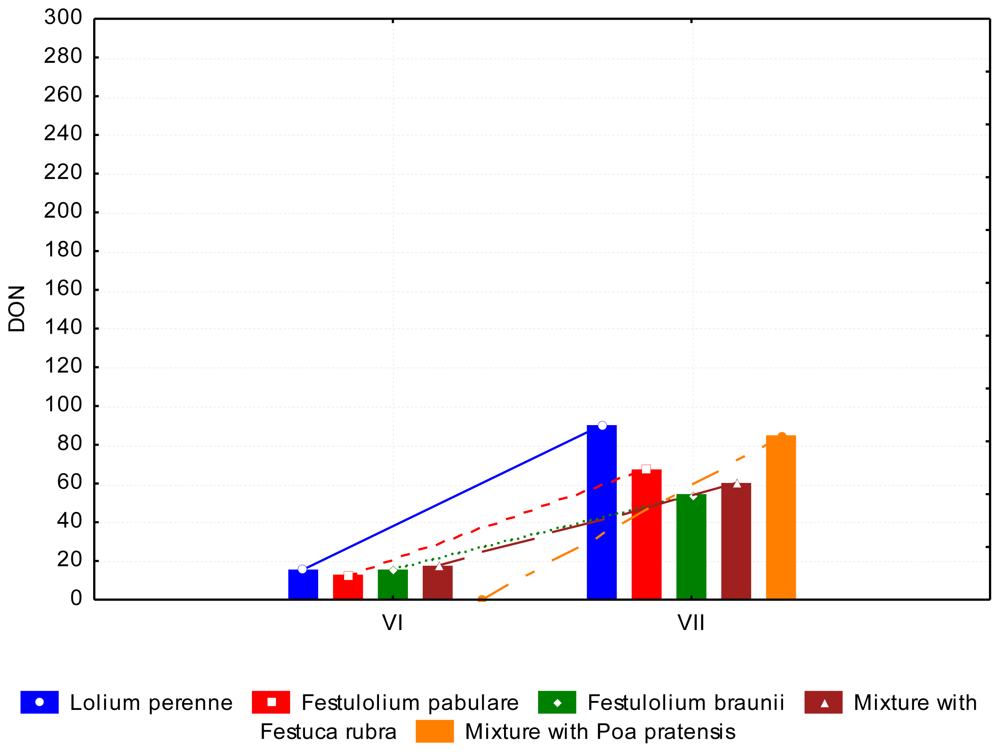
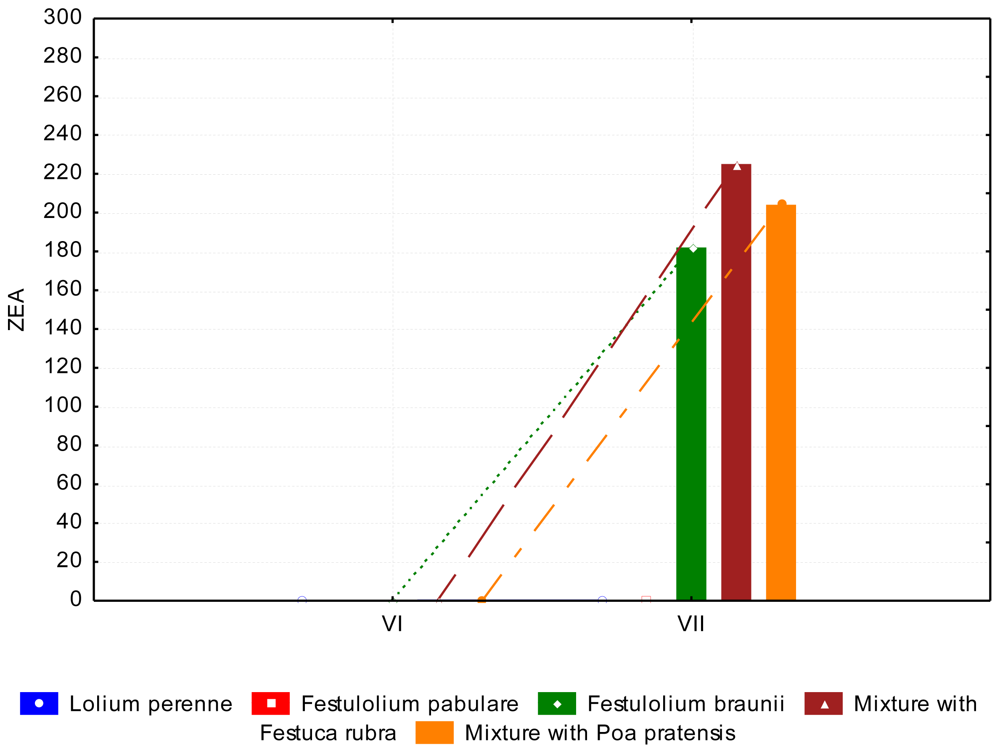
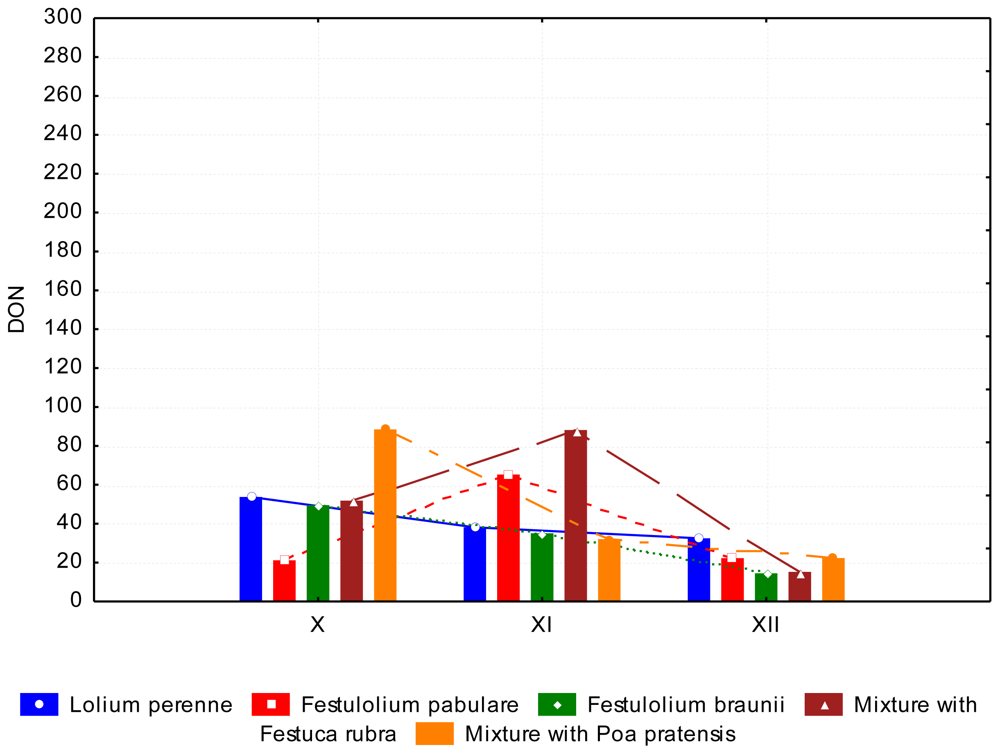
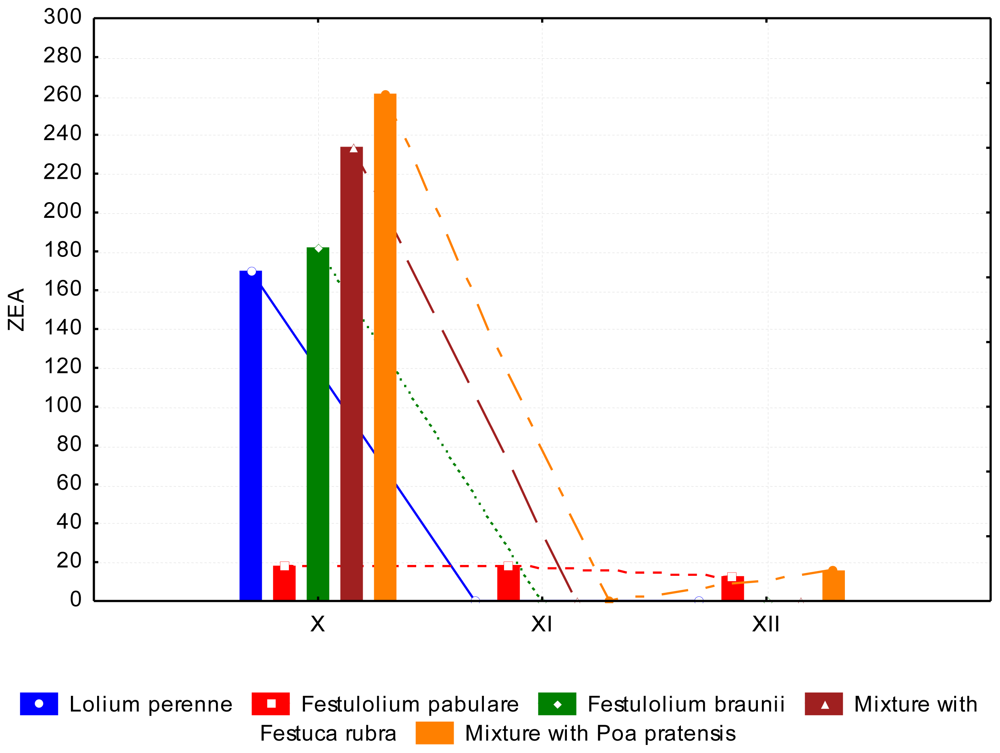
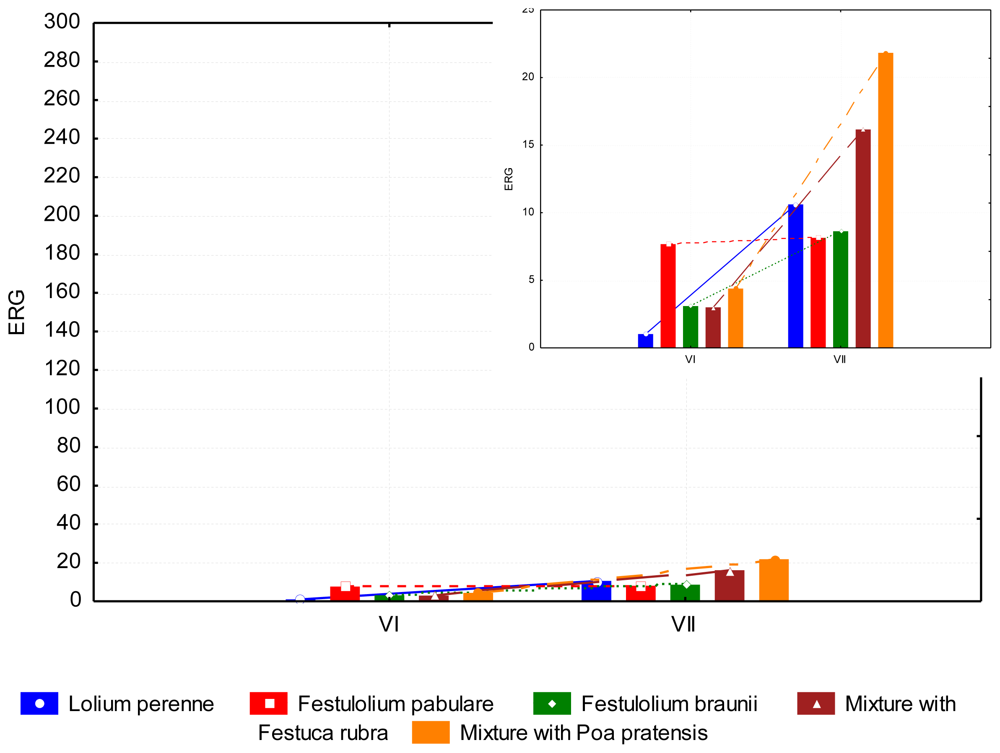
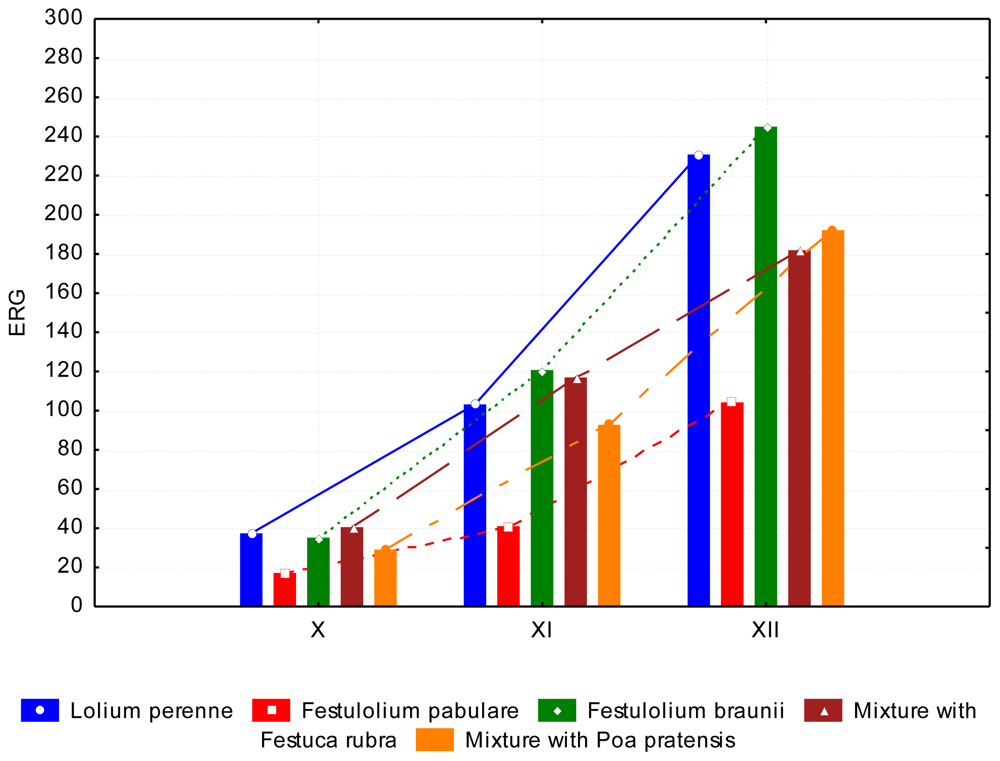
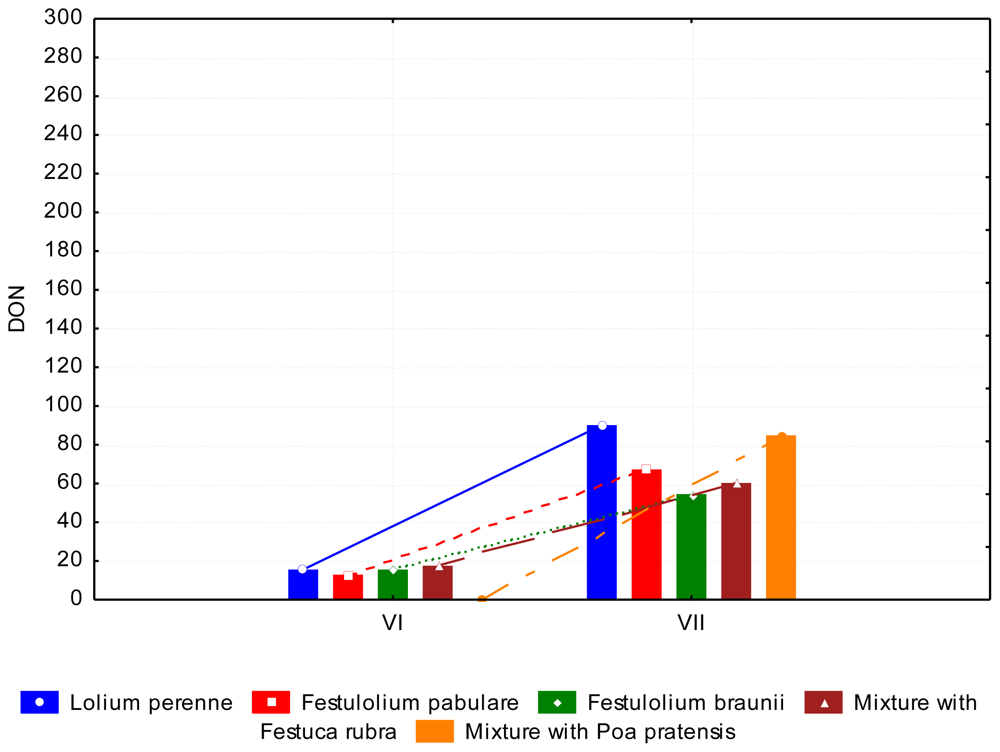
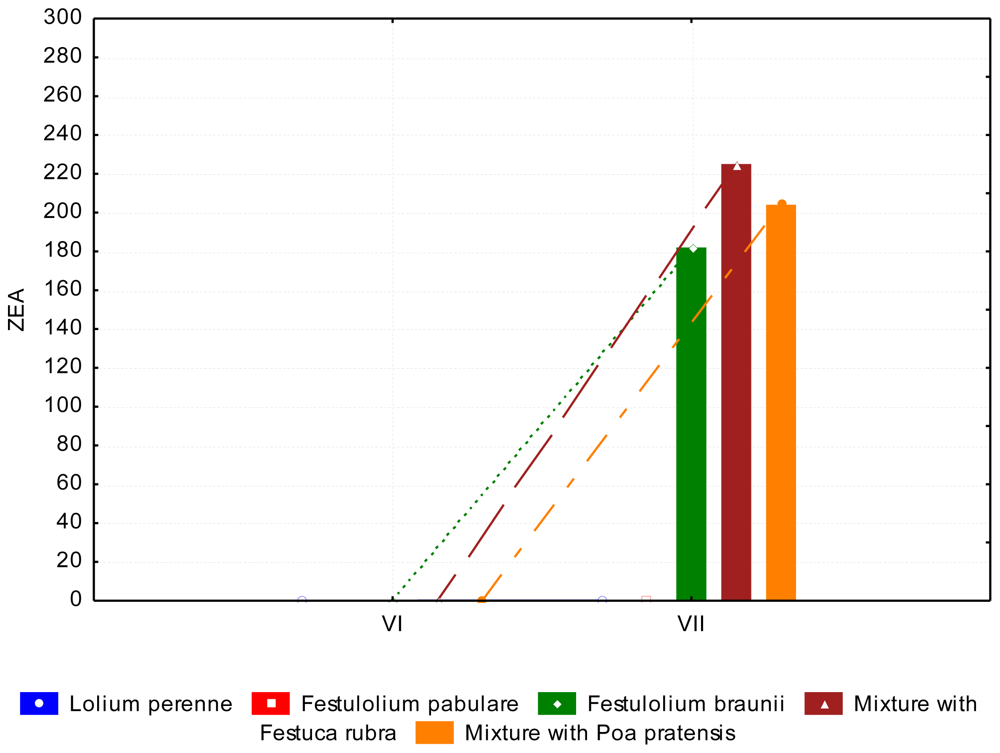
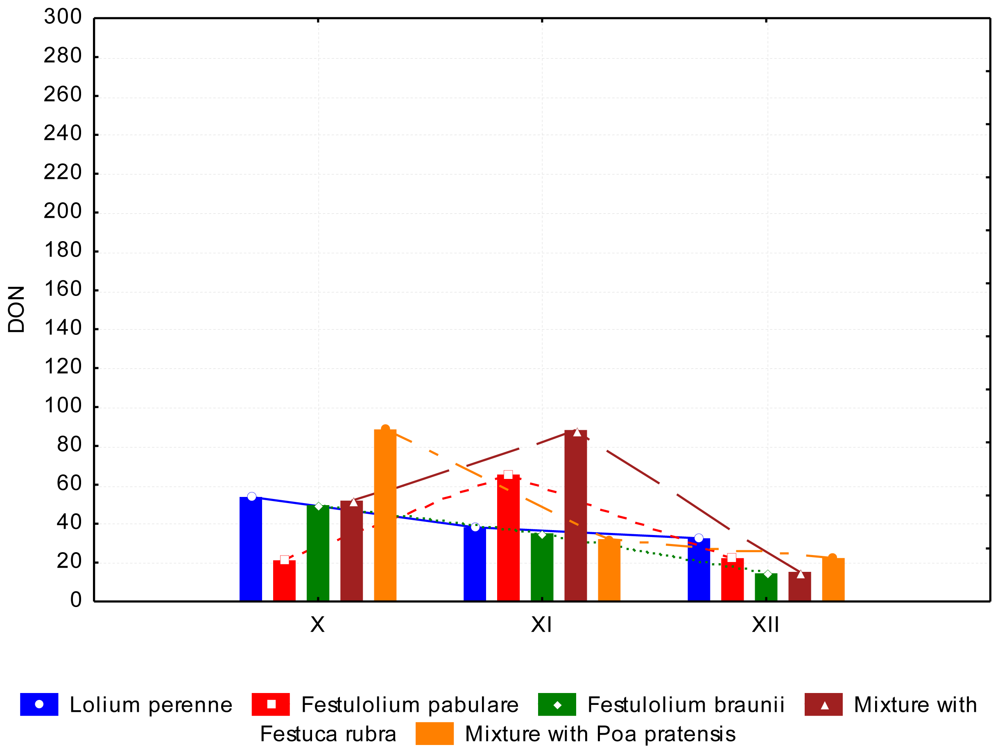
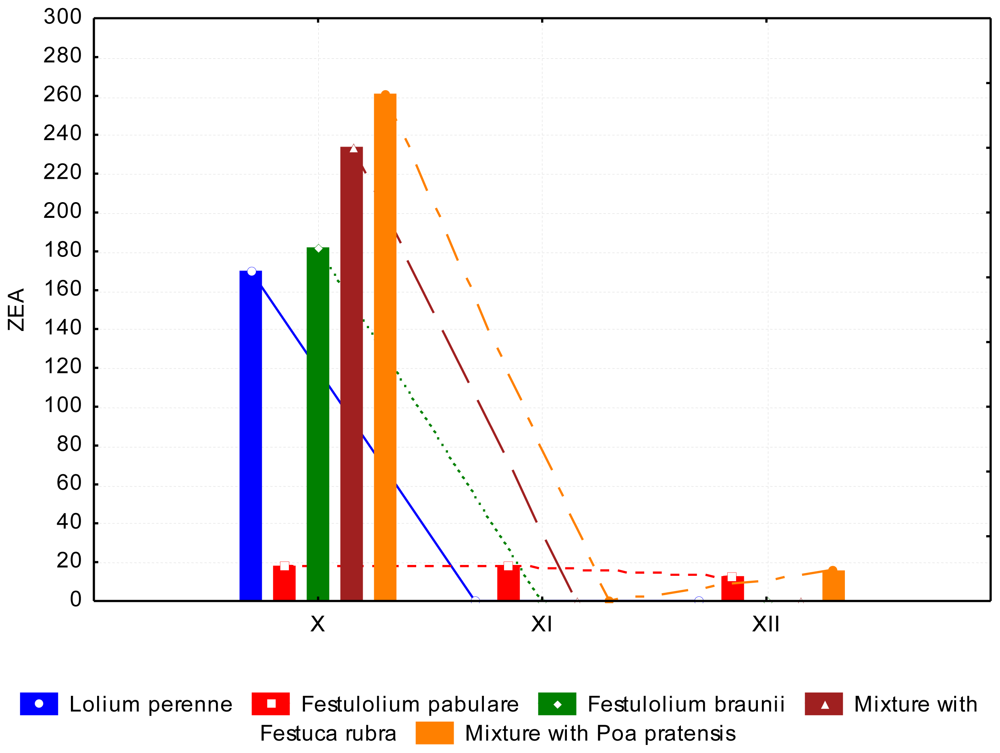
3.2. The Content of Mycotoxins and Statistical Evaluation of the Data
4. Conclusions
Acknowledgements
References and Notes
- Reverberi, M; Ricelli, A; Zjalic, S; Fabbri, AA; Fanelli, C. Natural functions of mycotoxins and control of their biosynthesis in fungi. Appl. Microbiol. Biotechnol 2010, 87, 899–911. [Google Scholar]
- Behrendt, U; Muller, T; Seyfarth, W. The influence of extensification in grassland management on the populations of micro-organisms in the phyllosphere of grasses. Microbiol. Res 1997, 152, 75–85. [Google Scholar]
- Holubek, R; Holubek, I; Stehlikova, B. The influence of various ways of regeneration and fertilization on economic profit from grass stand. Acta Fytotechnica et Zootechnica 2001, 5, 74–77. [Google Scholar]
- Giesler, LJ; Yuen, GY; Horst, GL. The microclimate in tall fescue turf as affected by canopy density and its influence on brown patch disease. Plant Dis 1996, 80, 389–394. [Google Scholar]
- Skladanka, J; Adam, V; Ryant, P; Dolezal, P; Havlicek, Z. Can be Festulolium, Dactylis glomerata and Arrhenatherum elatius used for extending autumn grazing season in Central Europe? Plant Soil Environ 2010, 56, 488–498. [Google Scholar]
- von Boberfeld, WO; Banzhaf, K; Hrabe, F; Skladanka, J; Kozlowski, S; Golinski, P; Szeman, L; Tasi, J. Effect of different agronomical measures on yield and quality of autumn saved herbage during winter grazing—2(nd) communication: Crude protein, energy and ergosterol concentration. Czech J. Anim. Sci 2006, 51, 271–277. [Google Scholar]
- von Boberfeld, WO. Changes of the quality including mycotoxin problems of the primary growth of a hay meadow—. Arrhenatherion elatioris. Agribiol. Res.-Z. Agrarbiol. Agrik.chem. Okol 1996, 49, 52–62. [Google Scholar]
- Leitao, AL. Potential of penicillium species in the bioremediation field. Int. J. Environ. Res. Public Health 2009, 6, 1393–1417. [Google Scholar]
- Sammon, NB; Harrower, KM; Fabbro, LD; Reed, RH. Microfungi in drinking water: The role of the frog. Litoria caerulea. Int. J. Environ. Res. Public Health 2010, 7, 3225–3234. [Google Scholar]
- von Boberfeld, WO; Banzhaf, K. Yield and forage quality of different xFestulolium cultivars in winter. J. Agron. Crop Sci 2006, 192, 239–247. [Google Scholar]
- Skladanka, J; Dohnal, V; Dolezal, P; Jezkova, A; Zeman, L. Factors affecting the content of ergosterol and zearalenone in selected grass species at the end of the growing season. Acta Vet. BRNO 2009, 78, 353–360. [Google Scholar]
- Marin, S; Vinaixa, M; Brezmes, J; Llobet, E; Vilanova, X; Correig, X; Ramos, AJ; Sanchis, V. Use of a MS-electronic nose for prediction of early fungal spoilage of bakery products. Int. J. Food Microbiol 2007, 114, 10–16. [Google Scholar]
- Vasatkova, A; Krizova, S; Adam, V; Zeman, L; Kizek, R. Changes in metallothionein level in rat hepatic tissue after administration of natural mouldy wheat. Int. J. Mol. Sci 2009, 10, 1138–1160. [Google Scholar]
- Vasatkova, A; Krizova, S; Krystofova, O; Adam, V; Zeman, L; Beklova, M; Kizek, R. Effect of naturally mouldy wheat or fungi administration on metallothioneins-3 level in brain tissues of rats. Neuroendocrinol. Lett 2010, 30, 163–168. [Google Scholar]
- Dohnal, V; Jezkova, A; Jun, D; Kuca, K. Metabolic pathways of T-2 toxin. Curr. Drug Metab 2008, 9, 77–82. [Google Scholar]
- Wu, QH; Dohnal, V; Huang, LL; Kuca, K; Yuan, ZH. Metabolic pathways of trichothecenes. Drug Metab. Rev 2010, 42, 250–267. [Google Scholar]
- Kushiro, M. Effects of milling and cooking processes on the deoxynivalenol content in wheat. Int. J. Mol. Sci 2008, 9, 2127–2145. [Google Scholar]
- Ueno, Y. The toxicology of mycotoxins. CRC Crit. Rev. Toxicol 1985, 14, 99–132. [Google Scholar]
- Ueno, Y. Toxicology of trichothecene mycotoxins. ISI Atlas Sci.-Pharm 1988, 2, 121–124. [Google Scholar]
- Sobrova, P; Adam, V; Vasatkova, A; Beklova, M; Zeman, L; Kizek, R. Deoxynivalenol and its toxicity. Interdisc. Toxicol 2010, 3, 101–106. [Google Scholar]
- Kumar, V; Basu, MS; Rajendran, TP. Mycotoxin research and mycoflora in some commercially important agricultural commodities. Crop Prot 2008, 27, 891–905. [Google Scholar]
- Mngadi, PT; Govinden, R; Odhav, B. Co-occurring mycotoxins in animal feeds. Afr. J. Biotechnol 2008, 7, 2239–2243. [Google Scholar]
- Niessen, L. PCR-based diagnosis and quantification of mycotoxin producing fungi. Int. J. Food Microbiol 2007, 119, 38–46. [Google Scholar]
- Sarmadha, MK; Balachandran, C. Serum electrolyte changes in Penicillic acid toxicosis. Indian Vet. J 2008, 85, 248–250. [Google Scholar]
- Wagacha, JM; Muthomi, JW. Mycotoxin problem in Africa: Current status, implications to food safety and health and possible management strategies. Int. J. Food Microbiol 2008, 124, 1–12. [Google Scholar]
- Annison, EF; Bryden, WL. Perspectives on ruminant nutrition and metabolism I. Metabolism in the rumen. Nutr. Res. Rev 1998, 11, 173–198. [Google Scholar]
- Schatzmayr, G; Zehner, F; Taubel, M; Schatzmayr, D; Klimitsch, A; Loibner, AP; Binder, EM. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res 2006, 50, 543–551. [Google Scholar]
- Gutzwiller, A; Czegledi, L; Stoll, P; Bruckner, L. Effects of Fusarium toxins on growth, humoral immune response and internal organs in weaner pigs, and the efficacy of apple pomace as an antidote. J. Anim. Physiol. Anim. Nutr 2007, 91, 432–438. [Google Scholar]
- Leung, MCK; Smith, TK; Karrow, NA; Boermans, HJ. Effects of feedborne Fusarium mycotoxins with and without a polymeric glucomannan mycotoxin adsorbent on body weight, feed intake, serum chemistry, and nutrient digestibility of mature beagles. Poult. Sci 2007, 86, 267–268. [Google Scholar]
- Kurtbay, HM; Bekci, Z; Merdivan, M; Yurdakoc, K. Reduction of ochratoxin A levels in red wine by bentonite, modified bentonites, and chitosan. J. Agric. Food Chem 2008, 56, 2541–2545. [Google Scholar]
- Worrell, NR; Mallett, AK; Cook, WM; Baldwin, NCP; Shepherd, MJ. The Role of Gut Microorganisms in the Metabolism of Deoxynivalenol Administered to Rats. Xenobiotica 1989, 19, 25–32. [Google Scholar]
- Baird, R; Abbas, HK; Windham, G; Williams, P; Baird, S; Ma, P; Kelley, R; Hawkins, L; Scruggs, M. Identification of select fumonisin forming Fusarium species using PCR applications of the polyketide synthase gene and its relationship to fumonisin production in vitro. Int. J. Mol. Sci 2008, 9, 554–570. [Google Scholar]
- Guan, S; Ji, C; Zhou, T; Li, JX; Ma, QG; Niu, TG. Aflatoxin B-1 degradation by Stenotrophomonas maltophilia and other microbes selected using coumarin medium. Int. J. Mol. Sci 2008, 9, 1489–1503. [Google Scholar]
- Mantle, PG; Nagy, JM. Binding of ochratoxin a to a urinary globulin: A new concept to account for gender difference in rat nephrocarcinogenic responses. Int. J. Mol. Sci 2008, 9, 719–735. [Google Scholar]
- Martins, HM; Almeida, I; Marques, M; Bernardo, F. Interaction of wild strains of Aspergilla with Aspergillus parasiticus ATCC15517 on aflatoxins production. Int. J. Mol. Sci 2008, 9, 394–400. [Google Scholar]
- Lancova, K; Hajslova, J; Kostelanska, M; Kohoutkova, J; Nedelnik, J; Moravcova, H; Vanova, M. Fate of trichothecene mycotoxins during the processing: Milling and baking. Food Addit. Contam 2008, 25, 650–659. [Google Scholar]
- Dohnal, V; Jezkova, A; Pavlikova, L; Musilek, K; Jun, D; Kuca, K. Fluctuation in the ergosterol and deoxynivalenol content in barley and malt during malting process. Anal. Bioanal. Chem 2010, 397, 109–114. [Google Scholar]
- Skladanka, J; Dohnal, V; Jezkova, A. Fibre and ergosterol contents in forage of Arrhenatherum elatius, Dactylis glomerata and Festulolium at the end of the growing season. Czech J. Anim. Sci 2008, 53, 320–329. [Google Scholar]
- de Nijs, M; Soentoro, P; Asch, EDV; Kamphuis, H; Rombouts, FM; Notermans, SHW. Fungal infection and presence of deoxynivalenol and zearalenone in cereals grown in the Netherlands. J. Food Prot 1996, 59, 772–777. [Google Scholar]
- Engels, R; Kramer, J. Incidence of fusaria and occurence of selected Fusarium mycotoxins on Lolium ssp. in Germany. Mycotoxin. Res 1996, 12, 31–40. [Google Scholar]
- Dmello, JPF; Macdonald, AMC. Mycotoxins. Anim. Feed Sci. Technol 1997, 69, 155–166. [Google Scholar]
- Marasas, WFO; Vanrensburg, SJ; Mirocha, CJ. Incidence of Fusarium species and the mycotoxins, deoxynivalenol and zearalenone, in corn produced in esophageal cancer areas in transkei. J. Agric. Food Chem 1979, 27, 1108–1112. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Deoxynivalenol | Zearalenone | Ergosterol | |||
|---|---|---|---|---|---|---|
| x (ppb) | s.e. | x (ppb) | s.e. | x (mg kg−1) | s.e. | |
| Specie | ||||||
| Lolium perenne | 46.02 | 10.83 | 34.06 | 30.87 | 76.61 ab | 29.80 |
| Festulolium pabulare | 37.78 | 9.61 | 9.82 | 5.05 | 35.65 a | 13.12 |
| Festulolium braunii | 33.83 | 9.06 | 72.85 | 48.53 | 82.53 b | 32.55 |
| Mixture with Festuca rubra | 46.60 | 11.54 | 91.81 | 61.18 | 71.71 ab | 24.30 |
| Mixture with Poa pratensis | 45.55 | 13.25 | 96.24 | 59.50 | 68.05 ab | 23.74 |
| Date of cut | ||||||
| June | 12.34 a | 5.07 | 0.0 a | 0.01 | 3.84 a | 0.92 |
| July | 71.43 b | 7.67 | 122.3 ab | 62.54 | 13.07 a | 3.72 |
| October | 52.95 bc | 9.16 | 173.0 b | 66.17 | 31.87 a | 3.91 |
| November | 51.71 bc | 12.04 | 3.7 a | 3.59 | 94.94 b | 11.31 |
| December | 21.35 ac | 6.07 | 5.8 a | 3.82 | 190.82 c | 23.89 |
| Year | ||||||
| 2008 | 37.63 | 7.66 | 115.76 a | 37.82 | 68.50 | 19.18 |
| 2009 | 46.28 | 5.67 | 6.15b | 2.50 | 65.31 | 12.05 |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Skládanka, J.; Nedělník, J.; Adam, V.; Doležal, P.; Moravcová, H.; Dohnal, V. Forage as a Primary Source of Mycotoxins in Animal Diets. Int. J. Environ. Res. Public Health 2011, 8, 37-50. https://doi.org/10.3390/ijerph8010037
Skládanka J, Nedělník J, Adam V, Doležal P, Moravcová H, Dohnal V. Forage as a Primary Source of Mycotoxins in Animal Diets. International Journal of Environmental Research and Public Health. 2011; 8(1):37-50. https://doi.org/10.3390/ijerph8010037
Chicago/Turabian StyleSkládanka, Jiří, Jan Nedělník, Vojtěch Adam, Petr Doležal, Hana Moravcová, and Vlastimil Dohnal. 2011. "Forage as a Primary Source of Mycotoxins in Animal Diets" International Journal of Environmental Research and Public Health 8, no. 1: 37-50. https://doi.org/10.3390/ijerph8010037
APA StyleSkládanka, J., Nedělník, J., Adam, V., Doležal, P., Moravcová, H., & Dohnal, V. (2011). Forage as a Primary Source of Mycotoxins in Animal Diets. International Journal of Environmental Research and Public Health, 8(1), 37-50. https://doi.org/10.3390/ijerph8010037