
Global DNA Methylation Profiles in Peripheral Blood of WTC-Exposed Community Members with Breast Cancer



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.1.1. World Trade Center Environmental Health Center (WTC EHC)
2.1.2. New York University Women’s Health Study (NYUWHS)
2.1.3. Study Design
2.2. Blood Sample Collection, Nucleic Acid Isolation, DNA Processing, and Differential Methylation Analysis
2.3. Statistical Analysis and Processing of Methylation Data
2.4. Functional Genomic Pathway Analysis
3. Results
3.1. WTC EHC Breast Cancer Cases vs. NYUWHS Pre-Diagnostic Breast Cancer Cases
3.1.1. Characteristics of WTC EHC Breast Cancer Cases vs. NYUWHS Pre-Diagnostic Breast Cancer Cases
3.1.2. Methylation Profiles of WTC EHC vs. NYUWHS Pre-Diagnostic Breast Cancer Cases
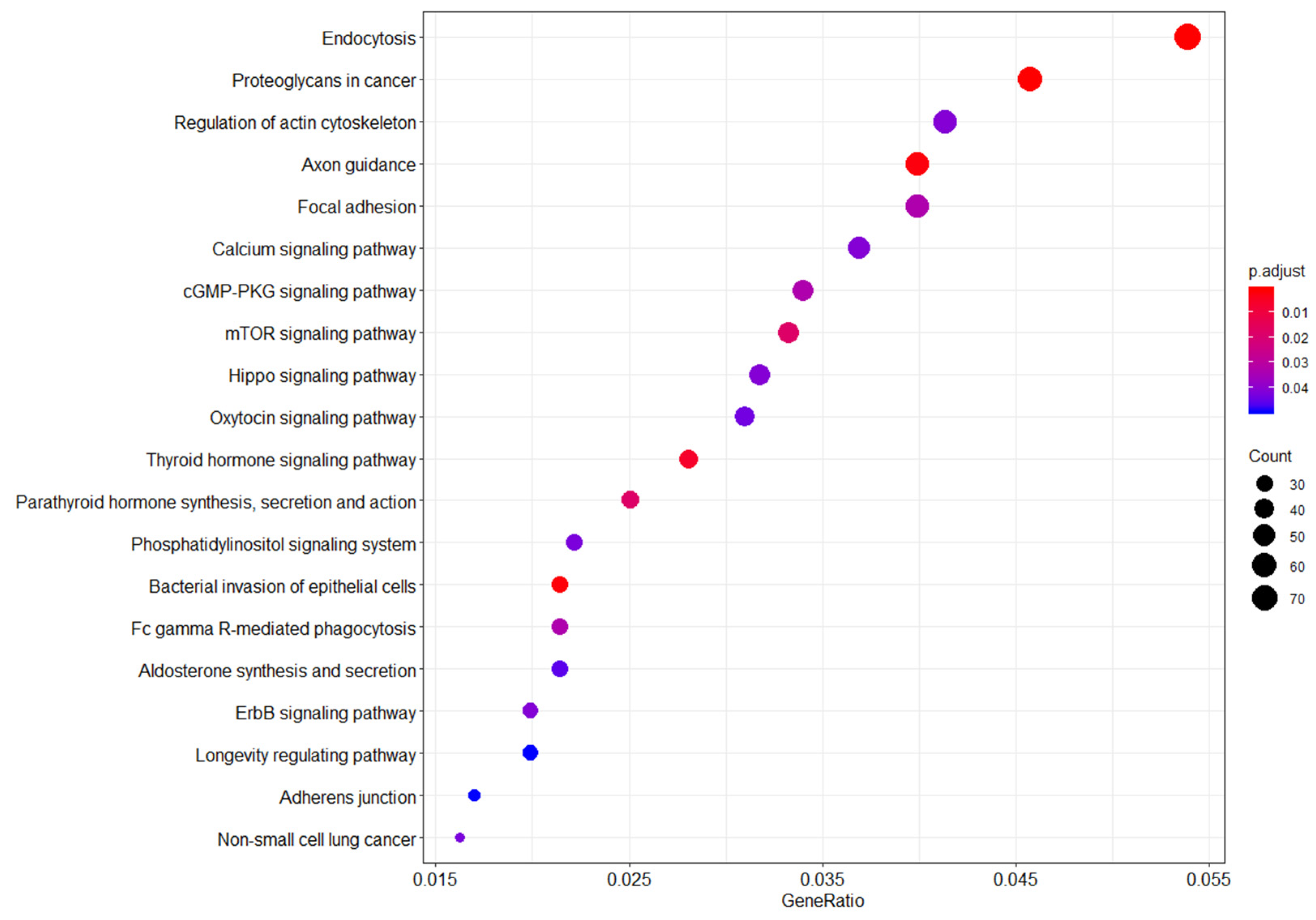
3.1.3. Functional Pathway Enrichment Analysis for WTC EHC Breast Cancer Cases vs. NYUWHS Pre-Diagnostic Breast Cancer Cases
3.2. WTC EHC Breast Cancer Cases vs. WTC EHC Cancer-Free Group
3.2.1. Characteristics of WTC EHC Breast Cancer Cases vs. WTC-Exposed Cancer-Free Group
3.2.2. Methylation Profiles of WTC EHC Breast Cancer Cases vs. Cancer-Free Women
3.2.3. Functional Pathway Enrichment Analysis for WTC EHC Breast Cancer Cases vs. Cancer-Free Women
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Claudio, L. Environmental Aftermath. Environ. Health Perspect. 2001, 109, A528–A536. [Google Scholar] [CrossRef] [PubMed]
- Lioy, P.J.; Georgopoulos, P. The Anatomy of the Exposures That Occurred around the World Trade Center Site: 9/11 and Beyond. Ann. N. Y. Acad. Sci. 2006, 1076, 54–79. [Google Scholar] [CrossRef] [PubMed]
- Yiin, L.-M.; Millette, J.R.; Vette, A.; Ilacqua, V.; Quan, C.; Gorczynski, J.; Kendall, M.; Chen, L.C.; Weisel, C.P.; Buckley, B.; et al. Comparisons of the Dust/Smoke Particulate That Settled inside the Surrounding Buildings and Outside on the Streets of Southern New York City after the Collapse of the World Trade Center, September 11, 2001. J. Air Waste Manag. Assoc. 2004, 54, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Zeig-Owens, R.; Webber, M.P.; Hall, C.B.; Schwartz, T.; Jaber, N.; Weakley, J.; Rohan, T.E.; Cohen, H.W.; Derman, O.; Aldrich, T.K.; et al. Early Assessment of Cancer Outcomes in New York City Firefighters after the 9/11 Attacks: An Observational Cohort Study. Lancet 2011, 378, 898–905. [Google Scholar] [CrossRef]
- Solan, S.; Wallenstein, S.; Shapiro, M.; Teitelbaum, S.L.; Stevenson, L.; Kochman, A.; Kaplan, J.; Dellenbaugh, C.; Kahn, A.; Biro, F.N.; et al. Cancer Incidence in World Trade Center Rescue and Recovery Workers, 2001–2008. Environ. Health Perspect. 2013, 121, 699–704. [Google Scholar] [CrossRef]
- Li, J.; Cone, J.E.; Kahn, A.R.; Brackbill, R.M.; Farfel, M.R.; Greene, C.M.; Hadler, J.L.; Stayner, L.T.; Stellman, S.D. Association between World Trade Center Exposure and Excess Cancer Risk. JAMA 2012, 308, 2479–2488. [Google Scholar] [CrossRef] [PubMed]
- Landrigan, P.J.; Lioy, P.J.; Thurston, G.; Berkowitz, G.; Chen, L.C.; Chillrud, S.N.; Gavett, S.H.; Georgopoulos, P.G.; Geyh, A.S.; Levin, S.; et al. Health and Environmental Consequences of the World Trade Center Disaster. Environ. Health Perspect. 2004, 112, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Durmus, N.; Shao, Y.; Arslan, A.A.; Zhang, Y.; Pehlivan, S.; Fernandez-Beros, M.-E.; Umana, L.; Corona, R.; Smyth-Giambanco, S.; Abbott, S.A.; et al. Characteristics of Cancer Patients in the World Trade Center Environmental Health Center. Int. J. Environ. Res. Public Health 2020, 17, 7190. [Google Scholar] [CrossRef]
- Arslan, A.; Zhang, Y.; Durmus, N.; Pehlivan, S.; Addessi, A.; Schnabel, F.; Shao, Y.; Reibman, J. Breast Cancer Characteristics in the Population of Survivors Participating in the World Trade Center Environmental Health Center Program 2002–2019. Int. J. Environ. Res. Public Health 2021, 18, 7555. [Google Scholar] [CrossRef]
- Mazzio, E.A.; Soliman, K.F. Basic Concepts of Epigenetics. Epigenetics 2012, 7, 119–130. [Google Scholar] [CrossRef]
- Baccarelli, A.; Bollati, V. Epigenetics and Environmental Chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Arslan, A.A.; Tuminello, S.; Yang, L.; Zhang, Y.; Durmus, N.; Snuderl, M.; Heguy, A.; Zeleniuch-Jacquotte, A.; Shao, Y.; Reibman, J. Genome-Wide DNA Methylation Profiles in Community Members Exposed to the World Trade Center Disaster. Int. J. Environ. Res. Public Health 2020, 17, 5493. [Google Scholar] [CrossRef] [PubMed]
- Kuan, P.-F.; Mi, Z.; Georgopoulos, P.; Hashim, D.; Luft, B.J.; Boffetta, P. Enhanced Exposure Assessment and Genome-Wide DNA Methylation in World Trade Center Disaster Responders. Eur. J. Cancer Prev. 2019, 28, 225–233. [Google Scholar] [CrossRef]
- Gong, Y.; Wang, L.; Yu, H.; Alpert, N.; Cohen, M.D.; Prophete, C.; Horton, L.; Sisco, M.; Park, S.-H.; Lee, H.-W.; et al. Prostate Cancer in World Trade Center Responders Demonstrates Evidence of an Inflammatory Cascade. Mol. Cancer Res. 2019, 17, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Reibman, J.; Levy-Carrick, N.; Miles, T.; Flynn, K.; Hughes, C.; Crane, M.; Lucchini, R.G. Destruction of the World Trade Center Towers. Lessons Learned from an Environmental Health Disaster. Ann. Am. Thorac. Soc. 2016, 13, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Durmus, N.; Zhang, Y.; Pehlivan, S.; Fernandez-Beros, M.-E.; Umana, L.; Corona, R.; Addessi, A.; Abbott, S.A.; Smyth-Giambanco, S.; et al. The Development of a WTC Environmental Health Center Pan-Cancer Database. Int. J. Environ. Res. Public Health 2021, 18, 1646. [Google Scholar] [CrossRef] [PubMed]
- Dagur, P.K.; McCoy, J.P. Collection, Storage, and Preparation of Human Blood Cells. Curr. Protoc. Cytom. 2015, 73, 5.1.1–5.1.16. [Google Scholar] [CrossRef]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A Flexible and Comprehensive Bioconductor Package for the Analysis of Infinium DNA Methylation Microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef]
- Walker, E.J.; Zhang, C.; Castelo-Branco, P.; Hawkins, C.; Wilson, W.; Zhukova, N.; Alon, N.; Novokmet, A.; Baskin, B.; Ray, P.; et al. Monoallelic Expression Determines Oncogenic Progression and Outcome in Benign and Malignant Brain Tumors. Cancer Res. 2012, 72, 636–644. [Google Scholar] [CrossRef]
- Catana, A.; Apostu, A.P.; Antemie, R.-G. Multi Gene Panel Testing for Hereditary Breast Cancer—Is It Ready to Be Used? Med. Pharm. Rep. 2019, 92, 220–225. [Google Scholar] [CrossRef]
- Shiovitz, S.; Korde, L.A. Genetics of Breast Cancer: A Topic in Evolution. Ann. Oncol. 2015, 26, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Raney, B.J.; Dreszer, T.R.; Barber, G.P.; Clawson, H.; Fujita, P.A.; Wang, T.; Nguyen, N.; Paten, B.; Zweig, A.S.; Karolchik, D.; et al. Track Data Hubs Enable Visualization of User-Defined Genome-Wide Annotations on the UCSC Genome Browser. Bioinformatics 2014, 30, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics Enrichment Tools: Paths toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Beissbarth, T.; Hyde, L.; Smyth, G.K.; Job, C.; Boon, W.-M.; Tan, S.-S.; Scott, H.S.; Speed, T.P. Statistical Modeling of Sequencing Errors in SAGE Libraries. Bioinformatics 2004, 20, i31–i39. [Google Scholar] [CrossRef][Green Version]
- Martin, E.M.; Fry, R.C. Environmental Influences on the Epigenome: Exposure- Associated DNA Methylation in Human Populations. Annu. Rev. Public Health 2018, 39, 309–333. [Google Scholar] [CrossRef]
- Ali, A.H.K.; Kondo, K.; Namura, T.; Senba, Y.; Takizawa, H.; Nakagawa, Y.; Toba, H.; Kenzaki, K.; Sakiyama, S.; Tangoku, A. Aberrant DNA Methylation of Some Tumor Suppressor Genes in Lung Cancers from Workers with Chromate Exposure. Mol. Carcinog. 2011, 50, 89–99. [Google Scholar] [CrossRef]
- Majumdar, S.; Chanda, S.; Ganguli, B.; Mazumder, D.N.G.; Lahiri, S.; Dasgupta, U.B. Arsenic Exposure Induces Genomic Hypermethylation. Environ. Toxicol. 2010, 25, 315–318. [Google Scholar] [CrossRef]
- Ruiz-Hernandez, A.; Kuo, C.-C.; Rentero-Garrido, P.; Tang, W.-Y.; Redon, J.; Ordovas, J.M.; Navas-Acien, A.; Tellez-Plaza, M. Environmental Chemicals and DNA Methylation in Adults: A Systematic Review of the Epidemiologic Evidence. Clin. Epigenetics 2015, 7, 55. [Google Scholar] [CrossRef]
- Hou, L.; Zhang, X.; Wang, D.; Baccarelli, A. Environmental Chemical Exposures and Human Epigenetics. Int. J. Epidemiol. 2012, 41, 79–105. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Marshall, W.F. Ciliogenesis: Building the Cell’s Antenna. Nat. Rev. Mol. Cell Biol. 2011, 12, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Steib, E.; Laporte, M.H.; Gambarotto, D.; Olieric, N.; Zheng, C.; Borgers, S.; Olieric, V.; Le Guennec, M.; Koll, F.; Tassin, A.-M.; et al. WDR90 Is a Centriolar Microtubule Wall Protein Important for Centriole Architecture Integrity. eLife 2020, 9, e57205. [Google Scholar] [CrossRef] [PubMed]
- OCM Oncomodulin [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=DetailsSearch&Term=654231 (accessed on 14 January 2022).
- Qu, X.; Zhao, B.; Hu, M.; Ji, Z.; Xu, J.; Xia, W.; Qu, Y. Downregulation of TBC1 Domain Family Member 24 (BC1D24) Inhibits Breast Carcinoma Growth via IGF1R/PI3K/AKT Pathway. Med. Sci. Monit. 2018, 24, 3987–3996. [Google Scholar] [CrossRef]
- Xue, J.; Chi, Y.; Chen, Y.; Huang, S.; Ye, X.; Niu, J.; Wang, W.; Pfeffer, L.M.; Shao, Z.-M.; Wu, Z.-H.; et al. MiRNA-621 Sensitizes Breast Cancer to Chemotherapy by Suppressing FBXO11 and Enhancing P53 Activity. Oncogene 2016, 35, 448–458. [Google Scholar] [CrossRef] [PubMed]
- You, E.; Huh, Y.H.; Kwon, A.; Kim, S.H.; Chae, I.H.; Lee, O.-J.; Ryu, J.-H.; Park, M.H.; Kim, G.-E.; Lee, J.S.; et al. SPIN90 Depletion and Microtubule Acetylation Mediate Stromal Fibroblast Activation in Breast Cancer Progression. Cancer Res. 2017, 77, 4710–4722. [Google Scholar] [CrossRef]
- Yasuoka, H.; Kodama, R.; Hirokawa, M.; Takamura, Y.; Miyauchi, A.; Inagaki, M.; Sanke, T.; Nakamura, Y. Neuropilin-2 Expression in Papillary Thyroid Carcinoma: Correlation with VEGF-D Expression, Lymph Node Metastasis, and VEGF-D-Induced Aggressive Cancer Cell Phenotype. J. Clin. Endocrinol. Metab. 2011, 96, E1857–E1861. [Google Scholar] [CrossRef]
- Rainero, E.; Cianflone, C.; Porporato, P.E.; Chianale, F.; Malacarne, V.; Bettio, V.; Ruffo, E.; Ferrara, M.; Benecchia, F.; Capello, D.; et al. The Diacylglycerol Kinase α/Atypical PKC/Β1 Integrin Pathway in SDF-1α Mammary Carcinoma Invasiveness. PLoS ONE 2014, 9, e97144. [Google Scholar] [CrossRef]
- De Smet, C.; Lurquin, C.; Lethé, B.; Martelange, V.; Boon, T. DNA Methylation Is the Primary Silencing Mechanism for a Set of Germ Line- and Tumor-Specific Genes with a CpG-Rich Promoter. Mol. Cell. Biol. 1999, 19, 7327–7335. [Google Scholar] [CrossRef]
- Luo, S.; Chen, J.; Mo, X. The Association of PTEN Hypermethylation and Breast Cancer: A Meta-Analysis. Onco Targets Ther. 2016, 9, 5643–5650. [Google Scholar] [CrossRef]
- Ma, N.; Li, X.; Wei, H.; Zhang, H.; Zhang, S. Circular RNA CircNFATC3 Acts as a MiR-9-5p Sponge to Promote Cervical Cancer Development by Upregulating SDC2. Cell Oncol. 2021, 44, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Seol, M.-A.; Park, J.-H.; Jeong, J.H.; Lyu, J.; Han, S.Y.; Oh, S.-M. Role of TOPK in Lipopolysaccharide-Induced Breast Cancer Cell Migration and Invasion. Oncotarget 2017, 8, 40190–40203. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Park, A.K.; Lee, K.-M.; Park, S.K.; Han, S.; Han, W.; Noh, D.-Y.; Yoo, K.-Y.; Kim, H.; Chanock, S.J.; et al. Candidate Gene Approach Evaluates Association between Innate Immunity Genes and Breast Cancer Risk in Korean Women. Carcinogenesis 2009, 30, 1528–1531. [Google Scholar] [CrossRef] [PubMed]
- Sgambato, A.; Caredda, E.; Leocata, P.; Rossi, G.; Boninsegna, A.; Vitale, A.; Grandi, T.; Cittadini, A.; Migaldi, M. Expression of Alpha-Dystroglycan Correlates with Tumour Grade and Predicts Survival in Oral Squamous Cell Carcinoma. Pathology 2010, 42, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xiao, Z.; Zhou, J.; Yang, M.; Feng, S.; Huang, Q.; Zou, J.; Zeng, T.; Li, Y.; Peng, L.; et al. ZNF382: A Transcription Inhibitor down-Regulated in Multiple Tumors Due to Promoter Methylation. Clin. Chim. Acta 2020, 500, 220–225. [Google Scholar] [CrossRef]
- Zabkiewicz, C.; Resaul, J.; Hargest, R.; Jiang, W.G.; Ye, L. Increased Expression of Follistatin in Breast Cancer Reduces Invasiveness and Clinically Correlates with Better Survival. Cancer Genom. Proteom. 2017, 14, 241–251. [Google Scholar] [CrossRef]
- Locke, W.J.; Guanzon, D.; Ma, C.; Liew, Y.J.; Duesing, K.R.; Fung, K.Y.C.; Ross, J.P. DNA Methylation Cancer Biomarkers: Translation to the Clinic. Front. Genet. 2019, 10, 1150. [Google Scholar] [CrossRef]
- Sunil, V.R.; Vayas, K.N.; Fang, M.; Zarbl, H.; Massa, C.; Gow, A.J.; Cervelli, J.A.; Kipen, H.; Laumbach, R.J.; Lioy, P.J.; et al. World Trade Center (WTC) Dust Exposure in Mice Is Associated with Inflammation, Oxidative Stress and Epigenetic Changes in the Lung. Exp. Mol. Pathol. 2017, 102, 50–58. [Google Scholar] [CrossRef]
- Reibman, J.; Liu, M.; Cheng, Q.; Liautaud, S.; Rogers, L.; Lau, S.; Berger, K.I.; Goldring, R.M.; Marmor, M.; Fernandez-Beros, M.E.; et al. Characteristics of a Residential and Working Community with Diverse Exposure to World Trade Center Dust, Gas, and Fumes. J. Occup. Environ. Med. 2009, 51, 534–541. [Google Scholar] [CrossRef]
- Bassett, L.W.; Liu, T.H.; Giuliano, A.E.; Gold, R.H. The prevalence of carcinoma in palpable vs impalpable, mammographically detected lesions. Am. J. Roentgenol. 1991, 157, 21–24. [Google Scholar] [CrossRef]
- Li, Y.; Pan, X.; Roberts, M.L.; Liu, P.; Kotchen, T.A.; Cowley, A.W., Jr.; Mattson, D.L.; Liu, Y.; Liang, M.; Kidambi, S. Stability of Global Methylation Profiles of Whole Blood and Extracted DNA under Different Storage Durations and Conditions. Epigenomics 2018, 10, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Saha, D.; Niemann, H.; Gryshkov, O.; Glasmacher, B.; Hofmann, N. Effects of Cryopreservation on the Epigenetic Profile of Cells. Cryobiology 2017, 74, 1–7. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Breast Cancer Cases, WTC EHC | Pre-Diagnostic Breast Cancer Cases, NYUWHS | Cancer-Free Controls, WTC EHC | |
|---|---|---|---|
| Characteristic | n = 28 | n = 24 | n = 18 |
| Age at blood donation, mean (SD) | 60.4 (8.9) | 52.1 (8.1) | 57.4 (8.4) |
| Race/ethnicity, n (%) | - | - | - |
| Caucasian | 16 (57.1) | 21 (87.5) | 8 (44.4) |
| Hispanic | 4 (14.3) | 1 (4.2) | 8 (44.4) |
| African-American | 7 (25.0) | 1 (4.2) | 1 (5.6) |
| Asian | 1 (3.6) | 1 (4.2) | 1 (5.6) |
| Body mass index, n (%) | - | - | - |
| Normal weight (<25) | 9 (32.1) | 10 (41.7) | 7 (39) |
| Overweight (25–30) | 11 (39.3) | 10 (41.7) | 6 (33) |
| Obese (≥30) | 8 (28.6) | 4 (16.7) | 5 (28) |
| Smoking, (n%) | - | - | - |
| Never | 17 (60.7) | 12 (50.0) | 12 (67) |
| Former | 10 (35.7) | 11 (45.8) | 5 (28) |
| Current | 1 (3.6) | 1 (4.2) | 1 (5) |
| Community member status, n (%) | - | - | - |
| Resident | 8 (28.6) | 24 (100) | 5 (27) |
| Local worker | 17 (60.7) | - | 11 (61) |
| Clean-up worker | 3 (10.7) | - | 2 (11) |
| WTC dust cloud exposure, n (%) | |||
| Acute, on 9/11 | 20 (71.4) | - | 12 (67) |
| Chronic, post 9/11 | 8 (28.6) | - | 6 (33) |
| Gene(s) | Probe ID | WTC EHC Breast Cancer Cases Methylation Value, Mean n = 28 | NYUWHS Breast Cancer Methylation Value, Mean n = 24 | p Value |
|---|---|---|---|---|
| IFT74 | cg00877966 | 0.056 | 0.089 | 8.28 × 10−17 |
| WDR90 | cg00320059 | 0.785 | 0.737 | 1.49 × 10−16 |
| OCM | cg20061873 | 0.369 | 0.443 | 3.66 × 10−16 |
| NA | cg14471455 | 0.582 | 0.661 | 3.78 × 10−16 |
| TBC1D24 | cg21791024 | 0.681 | 0.613 | 5.78 × 10−16 |
| RIN3 | cg11815205 | 0.142 | 0.203 | 1.06 × 10−15 |
| FBXO11 | cg16602504 | 0.143 | 0.110 | 1.19 × 10−15 |
| SEMA4G; MRPL43 | cg19672644 | 0.737 | 0.677 | 1.67 × 10−15 |
| GPRIN3 | cg21998046 | 0.317 | 0.464 | 1.68 × 10−15 |
| ODZ4 | cg00299230 | 0.823 | 0.778 | 1.83 × 10−15 |
| LINC00426 | cg18855351 | 0.203 | 0.294 | 2.31 × 10−15 |
| NCKIPSD | cg18849300 | 0.171 | 0.238 | 2.35 × 10−15 |
| TIMM23B | cg09227337 | 0.798 | 0.740 | 2.48 × 10−15 |
| ACVR1B | cg17781357 | 0.562 | 0.600 | 2.63 × 10−15 |
| NRP2 | cg01323148 | 0.201 | 0.269 | 2.73 × 10−15 |
| DGKA | cg13634319 | 0.239 | 0.347 | 3.32 × 10−15 |
| Gene | Probe ID | WTC EHC Methylation Value, Mean | NYUWHS Methylation Value, Mean | Methylation Status * | Regulatory Feature | Breast Cancer Penetrance [21] |
|---|---|---|---|---|---|---|
| ATM | cg18457775 | 0.055 | 0.041 | Hypermethylated | Promoter | Moderate |
| BRCA1 | cg16630982 | 0.096 | 0.077 | Hypermethylated | Promoter | High |
| CARS | cg25637226 | 0.749 | 0.702 | Hypermethylated | NA | Poorly characterized |
| CDKN2C | cg07013994 | 0.052 | 0.040 | Hypermethylated | Promoter | Poorly characterized |
| CREBBP | cg01963870 | 0.900 | 0.871 | Hypermethylated | Gene | Poorly characterized |
| EXT1 | cg14850625 | 0.066 | 0.045 | Hypermethylated | Promoter | Poorly characterized |
| NBN | cg22881279 | 0.062 | 0.046 | Hypermethylated | Promoter | Poorly characterized |
| NOTCH1 | cg23457546 | 0.735 | 0.680 | Hypermethylated | NA | Poorly characterized |
| NUP98 | cg23511374 | 0.180 | 0.144 | Hypermethylated | Promoter | Poorly characterized |
| PALB2 | cg07627390 | 0.093 | 0.064 | Hypermethylated | Promoter | Moderate |
| PLM | cg26861525 | 0.055 | 0.082 | Hypomethylated | Promoter | Poorly characterized |
| PMS1 | cg09481946 | 0.064 | 0.046 | Hypermethylated | Promoter | Poorly characterized |
| PTEN | cg20849549 | 0.075 | 0.093 | Hypomethylated | Promoter | High |
| RUNX1 | cg21358438 | 0.736 | 0.694 | Hypermethylated | NA | Poorly characterized |
| TP53 | cg18198734 | 0.847 | 0.817 | Hypermethylated | NA | High |
| TSC2 | cg02504384 | 0.651 | 0.596 | Hypermethylated | Unclassified | Poorly characterized |
| WRN | cg03410815 | 0.106 | 0.076 | Hypermethylated | Promoter | Poorly characterized |
| Gene(s) | Probe ID | WTC EHC Breast Cancer Cases Methylation Value, Mean n = 28 | WTC EHC Controls Methylation Value, Mean n = 18 | p Value |
|---|---|---|---|---|
| SDC2 | cg10292139 | 0.085 | 0.112 | 2.33 × 10−6 |
| LINC00578 | cg15710610 | 0.573 | 0.653 | 3.93 × 10−6 |
| PBK | cg17973773 | 0.697 | 0.630 | 4.11 × 10−6 |
| NA | cg18281243 | 0.059 | 0.036 | 5.38 × 10−6 |
| ALOX12B | cg11868553 | 0.806 | 0.840 | 6.98 × 10−6 |
| TMEM222 | cg26528255 | 0.523 | 0.555 | 9.14 × 10−6 |
| NA | cg21166544 | 0.301 | 0.335 | 9.24 × 10−6 |
| NA | cg25992645 | 0.095 | 0.066 | 9.67 × 10−6 |
| NA | cg00187322 | 0.839 | 0.862 | 9.70 × 10−6 |
| ZSWIM6 | cg15221192 | 0.892 | 0.910 | 9.77 × 10−6 |
| DAG1 | cg11048959 | 0.067 | 0.075 | 1.27 × 10−5 |
| ZNF529; ZNF382 | cg19406736 | 0.096 | 0.115 | 1.40 × 10−5 |
| NA | cg09562797 | 0.768 | 0.728 | 1.54 × 10−5 |
| NA | cg05526438 | 0.623 | 0.650 | 1.56 × 10−5 |
| HIST1H4A; HIST1H3A | cg07109238 | 0.055 | 0.062 | 1.59 × 10−5 |
| CNBD1 | cg20630812 | 0.542 | 0.569 | 1.64 × 10−5 |
| NA | cg19366463 | 0.688 | 0.717 | 1.73 × 10−5 |
| MYCBPAP | cg09079356 | 0.789 | 0.810 | 1.75 × 10−5 |
| NARS | cg21911276 | 0.462 | 0.381 | 1.81 × 10−5 |
| ZNF607 | cg25563044 | 0.058 | 0.067 | 1.86 × 10−5 |
| FST | cg22288251 | 0.111 | 0.132 | 1.90 × 10−5 |
| ZCCHC8 | cg02708659 | 0.143 | 0.161 | 1.90 × 10−5 |
| Gene | Probe ID | Breast Cancer Methylation Value, Mean | Cancer-Free Methylation Value, Mean | Methylation Status * | Regulatory Feature | Breast Cancer Penetrance [21] |
|---|---|---|---|---|---|---|
| BCL11B | cg23580725 | 0.392 | 0.461 | Hypomethylated | NA | Poorly characterized |
| CBFA2T3 | cg05540133 | 0.914 | 0.935 | Hypomethylated | NA | Poorly characterized |
| EXT1 | cg01483826 | 0.573 | 0.277 | Hypermethylated | NA | Poorly characterized |
| EXT2 | cg19330452 | 0.812 | 0.794 | Hypermethylated | NA | Poorly characterized |
| MAP2K4 | cg26019016 | 0.893 | 0.908 | Hypomethylated | NA | Poorly characterized |
| MEN1 | cg00603409 | 0.723 | 0.743 | Hypomethylated | NA | Poorly characterized |
| NF1 | cg07917842 | 0.546 | 0.574 | Hypomethylated | NA | Poorly characterized |
| NOTCH1 | cg04271687 | 0.054 | 0.059 | Hypomethylated | Promoter | Poorly characterized |
| NPM1 | cg17872779 | 0.119 | 0.131 | Hypomethylated | Unclassified | Poorly characterized |
| NR4A3 | cg13412395 | 0.054 | 0.058 | Hypomethylated | NA | Poorly characterized |
| NUP98 | cg20457962 | 0.138 | 0.154 | Hypomethylated | Promoter | Poorly characterized |
| PMS2 | cg10310847 | 0.095 | 0.114 | Hypomethylated | NA | Poorly characterized |
| RB1 | cg13389575 | 0.842 | 0.863 | Hypomethylated | NA | Poorly characterized |
| RUNX1 | cg13521940 | 0.137 | 0.110 | Hypermethylated | NA | Poorly characterized |
| SMARCA4 | cg17094383 | 0.909 | 0.923 | Hypomethylated | NA | Poorly characterized |
| STK11 | cg02671671 | 0.064 | 0.047 | Hypermethylated | Promoter | High |
| SUFU | cg07636870 | 0.045 | 0.048 | Hypomethylated | Promoter | Poorly characterized |
| TSC2 | cg25446438 | 0.515 | 0.547 | Hypomethylated | Promoter | Poorly characterized |
| WT1 | cg22533573 | 0.129 | 0.111 | Hypermethylated | NA | Poorly characterized |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuminello, S.; Zhang, Y.; Yang, L.; Durmus, N.; Snuderl, M.; Heguy, A.; Zeleniuch-Jacquotte, A.; Chen, Y.; Shao, Y.; Reibman, J.; et al. Global DNA Methylation Profiles in Peripheral Blood of WTC-Exposed Community Members with Breast Cancer. Int. J. Environ. Res. Public Health 2022, 19, 5104. https://doi.org/10.3390/ijerph19095104
Tuminello S, Zhang Y, Yang L, Durmus N, Snuderl M, Heguy A, Zeleniuch-Jacquotte A, Chen Y, Shao Y, Reibman J, et al. Global DNA Methylation Profiles in Peripheral Blood of WTC-Exposed Community Members with Breast Cancer. International Journal of Environmental Research and Public Health. 2022; 19(9):5104. https://doi.org/10.3390/ijerph19095104
Chicago/Turabian StyleTuminello, Stephanie, Yian Zhang, Lei Yang, Nedim Durmus, Matija Snuderl, Adriana Heguy, Anne Zeleniuch-Jacquotte, Yu Chen, Yongzhao Shao, Joan Reibman, and et al. 2022. "Global DNA Methylation Profiles in Peripheral Blood of WTC-Exposed Community Members with Breast Cancer" International Journal of Environmental Research and Public Health 19, no. 9: 5104. https://doi.org/10.3390/ijerph19095104
APA StyleTuminello, S., Zhang, Y., Yang, L., Durmus, N., Snuderl, M., Heguy, A., Zeleniuch-Jacquotte, A., Chen, Y., Shao, Y., Reibman, J., & Arslan, A. A. (2022). Global DNA Methylation Profiles in Peripheral Blood of WTC-Exposed Community Members with Breast Cancer. International Journal of Environmental Research and Public Health, 19(9), 5104. https://doi.org/10.3390/ijerph19095104