
Flurochloridone Induced Cell Apoptosis via ER Stress and eIF2α-ATF4/ATF6-CHOP-Bim/Bax Signaling Pathways in Mouse TM4 Sertoli Cells


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Cell Viability Assay
2.3. Cytotoxicity Assay
2.4. Determination of Apoptotic Cells by Flow Cytometry
2.5. Determination of ER Stress, UPR Signaling Pathways and Apoptosis Related Proteins by Western Blot
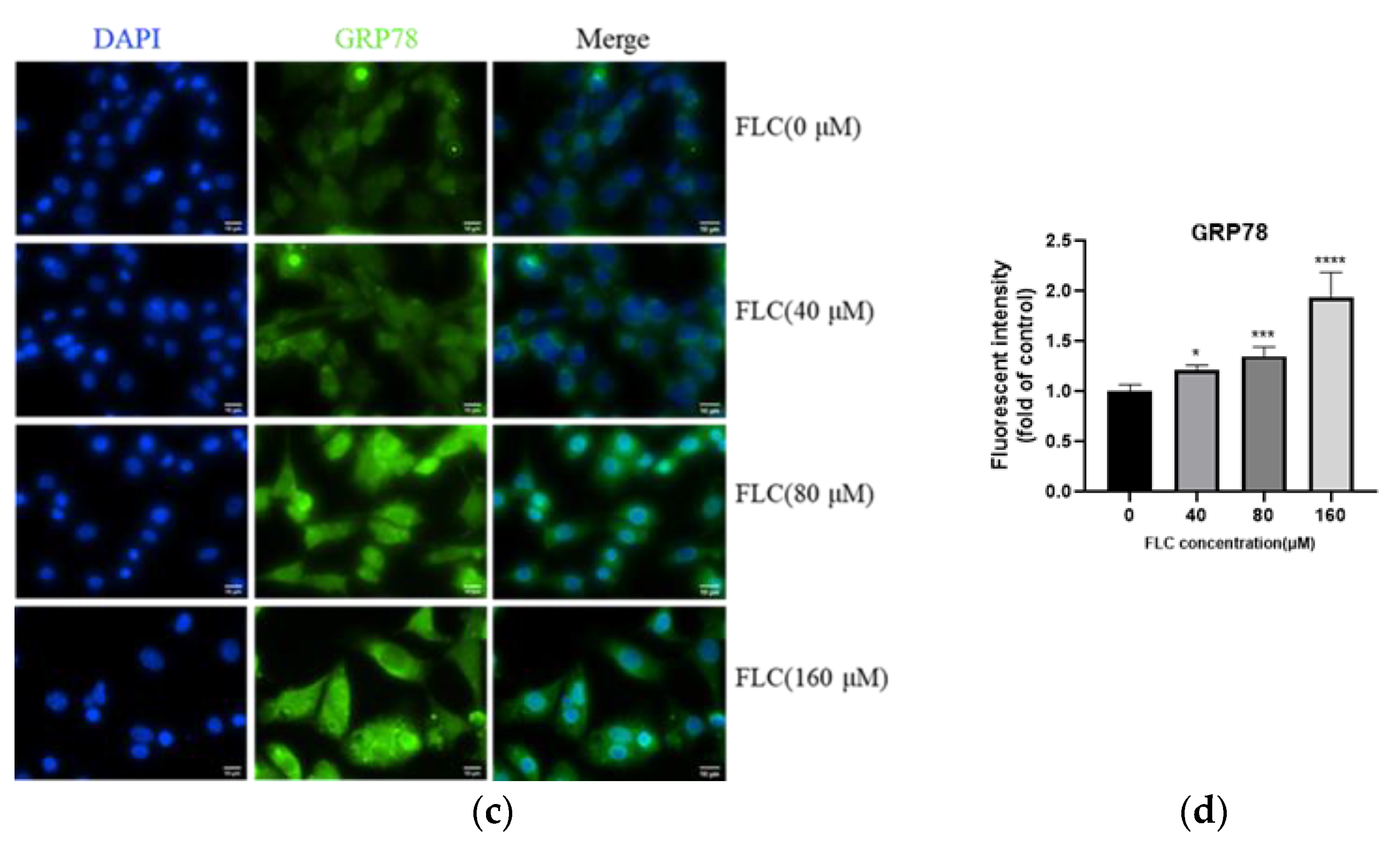
2.6. Determination of GRP78 by Immunofluorescence Staining
2.7. Statistical Analysis
3. Results
3.1. FLC Inhibited Cell Viability and Induced Cytotoxicity in TM4 Cells
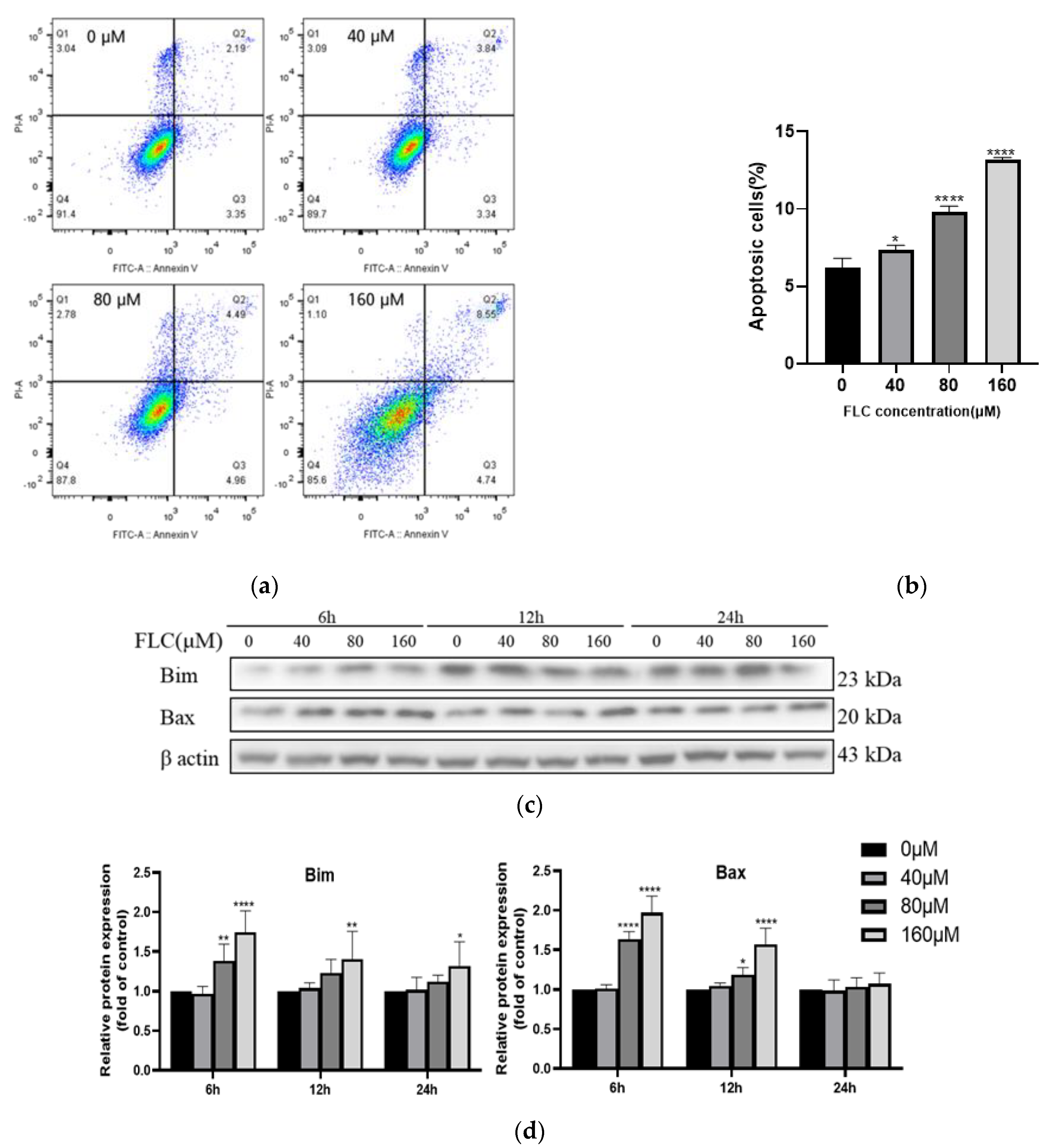
3.2. FLC Induced Apoptosis and Up-Regulated the Level of Pro-Apoptotic Proteins
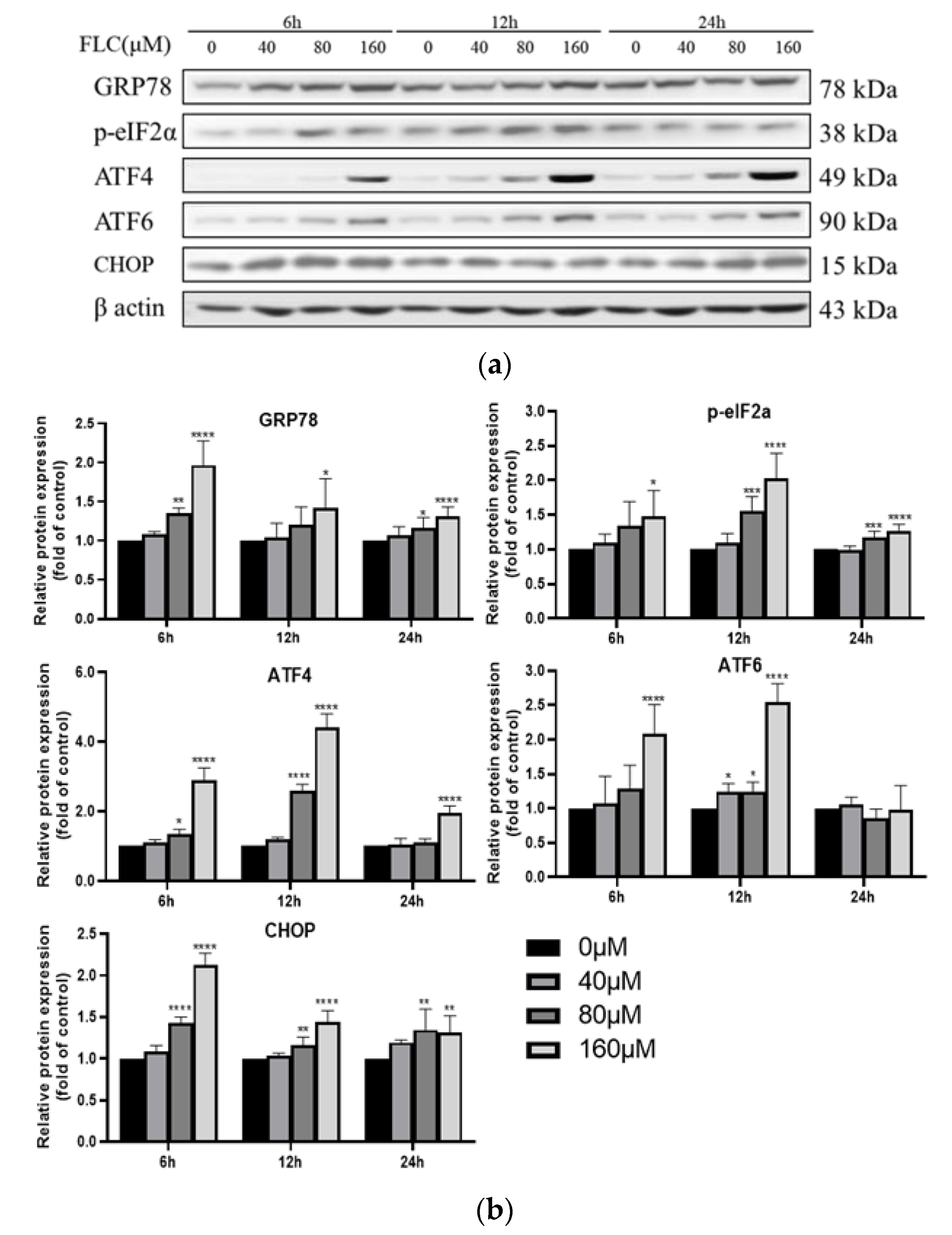
3.3. FLC Induced Endoplasmic Reticulum (ER) Stress and Activated Unfolded Protein Response (UPR) Signaling Pathways
3.4. FLC Upregulated the Protein Level of CHOP
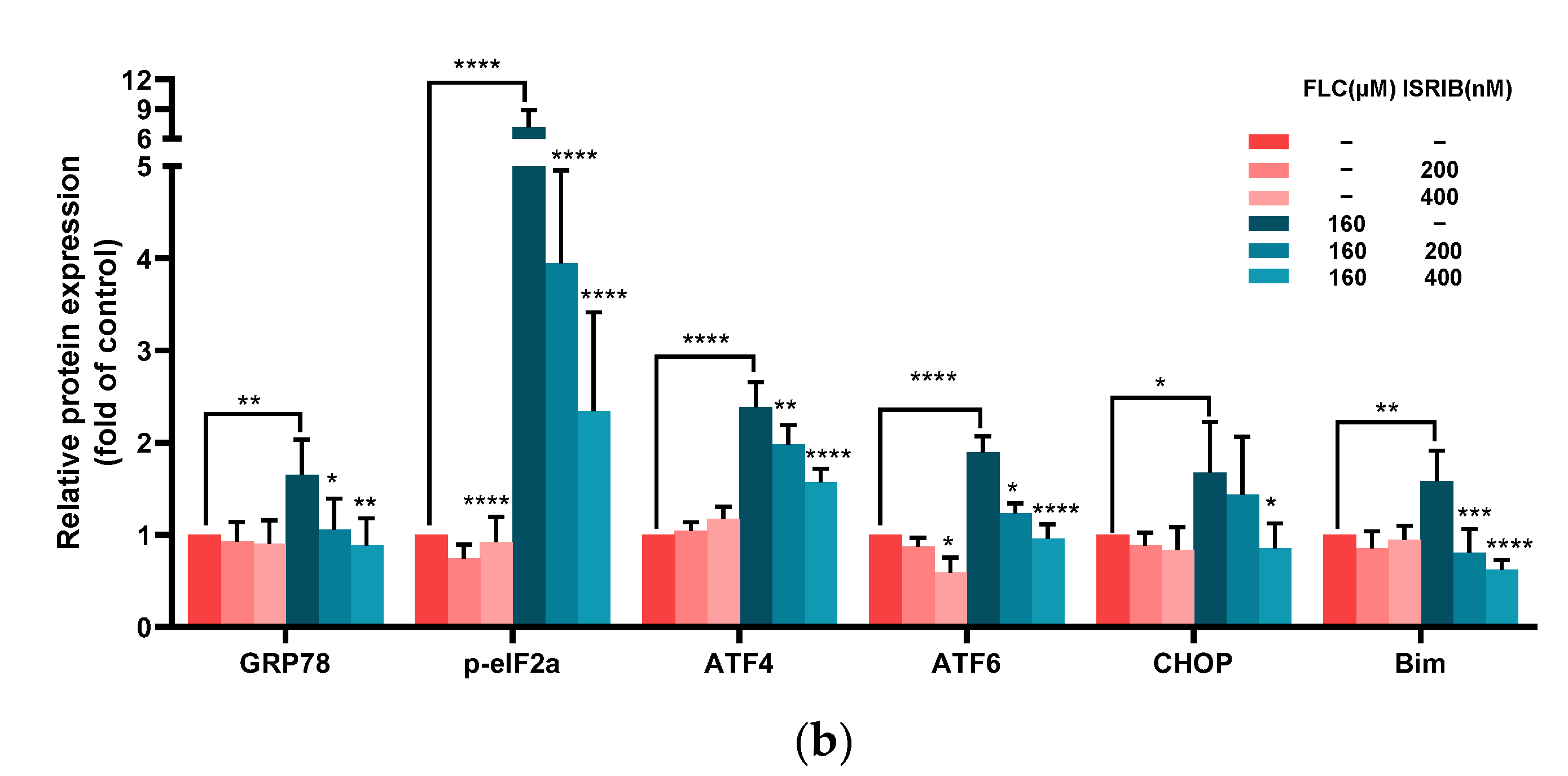
3.5. ISRIB Inhibited UPR Signaling Pathways and Alleviated ER Stress
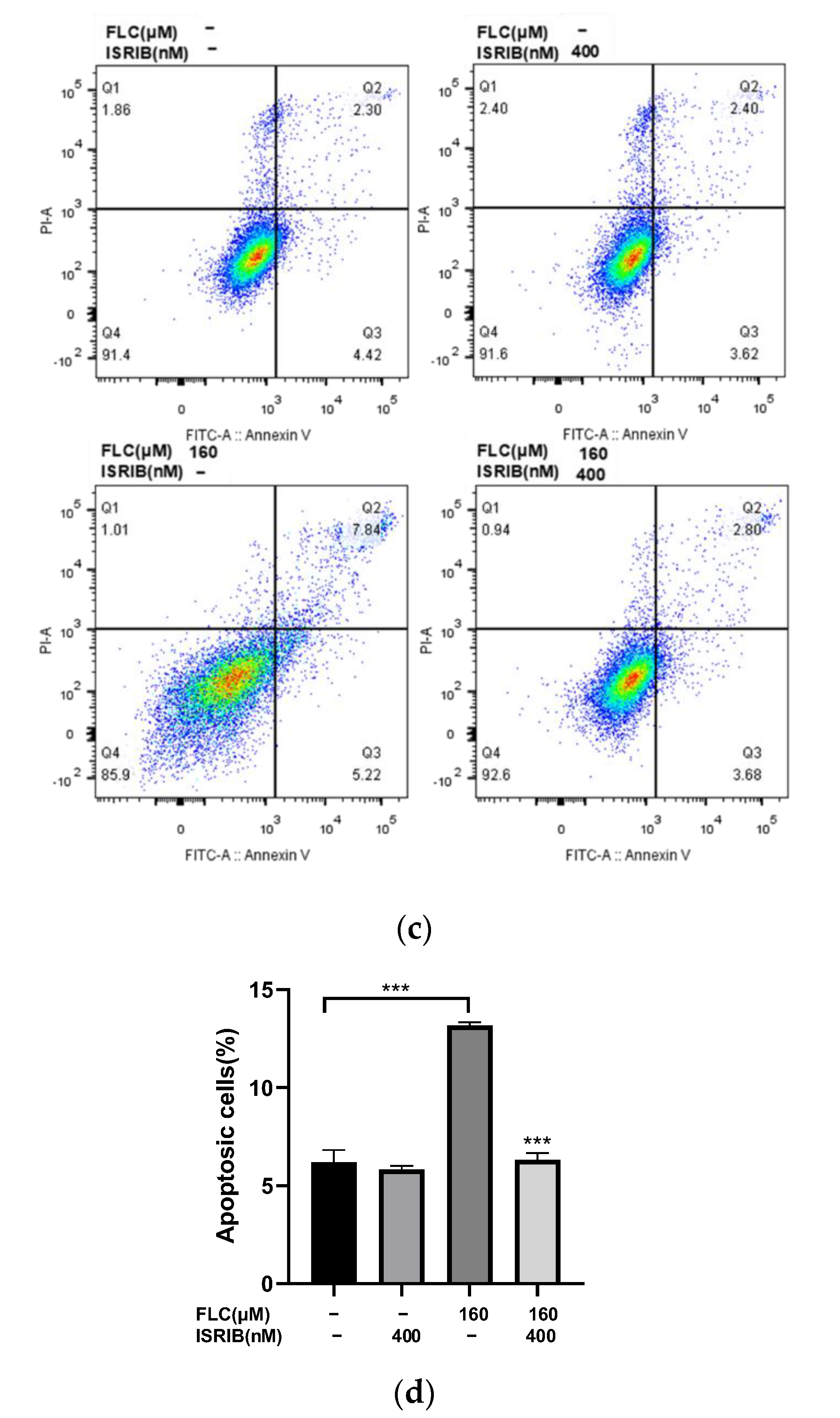
3.6. ISRIB Alleviated FLC-Induced Cytotoxicity and Apoptosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaya, A.; Yigit, E. The physiological and biochemical effects of salicylic acid on sunflowers (Helianthus annuus) exposed to flurochloridone. Ecotoxicol. Environ. Saf. 2014, 106, 232–238. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Conclusion on the peer review of the pesticide risk assessment of the active substance Flurochloridone (notified active substance). EFSA J. 2010, 8, 1869. [Google Scholar] [CrossRef] [Green Version]
- Nikoloff, N.; Natale, G.S.; Marino, D.; Soloneski, S.; Larramendy, M.L. Flurochloridone-based herbicides induced genotoxicity effects on Rhinella arenarum tadpoles (Anura: Bufonidae). Ecotoxicol. Environ. Saf. 2014, 100, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Nikoloff, N.; Soloneski, S.; Larramendy, M.L. Genotoxic and cytotoxic evaluation of the herbicide flurochloridone on Chinese hamster ovary (CHO-K1) cells. Toxicol. In Vitro 2012, 26, 157–163. [Google Scholar] [CrossRef]
- Nikoloff, N.; Larramendy, M.L.; Soloneski, S. Assessment of DNA damage, cytotoxicity, and apoptosis in human hepatoma (HepG2) cells after flurochloridone herbicide exposure. Food Chem. Toxicol. 2014, 65, 233–241. [Google Scholar] [CrossRef]
- Zhang, S.; Cheng, X.; Wang, Y.; Fan, J.; Li, R.; Zhou, S.; Liu, S.; Shi, J.; Sun, J.; Hu, Y.; et al. Ninety day toxicity and toxicokinetics of fluorochloridone after oral administration in rats. Int. J. Environ. Res. Public Health 2015, 12, 4942–4966. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Li, R.; Zhou, S.; Zhang, S.; Wang, Y.; Liu, S.; Song, Q.; Chang, X.; Zhang, Y.; Liu, L.; et al. The Oral NOAEL of Flurochloridone in Male Wistar Rats in Ninety-Day Subchronic Toxicity Test Was 3 mg/kg/day. Int. J. Environ. Res. Public Health 2019, 16, 533. [Google Scholar] [CrossRef] [Green Version]
- Mruk, D.D.; Cheng, C.Y. The Mammalian Blood-Testis Barrier: Its Biology and Regulation. Endocr. Rev. 2015, 36, 564–591. [Google Scholar] [CrossRef]
- Larose, H.; Kent, T.; Ma, Q.; Shami, A.N.; Harerimana, N.; Li, J.Z.; Hammoud, S.S.; Handel, M.A. Regulation of meiotic progression by Sertoli-cell androgen signaling. Mol. Biol. Cell 2020, 31, 2841–2862. [Google Scholar] [CrossRef]
- Wu, S.; Yan, M.; Ge, R.; Cheng, C.Y. Crosstalk between Sertoli and Germ Cells in Male Fertility. Trends Mol. Med. 2020, 26, 215–231. [Google Scholar] [CrossRef]
- Alves, M.G.; Neuhaus-Oliveira, A.; Moreira, P.I.; Socorro, S.; Oliveira, P.F. Exposure to 2,4-dichlorophenoxyacetic acid alters glucose metabolism in immature rat Sertoli cells. Reprod. Toxicol. 2013, 38, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Gorga, A.; Rindone, G.M.; Centola, C.L.; Sobarzo, C.; Pellizzari, E.H.; Camberos, M.D.C.; Cigorraga, S.B.; Riera, M.F.; Galardo, M.N.; Meroni, S.B. In vitro effects of glyphosate and Roundup on Sertoli cell physiology. Toxicol. In Vitro 2020, 62, 104682. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Mruk, D.; Lian, Q.; Ge, R.; Li, C.; Silvestrini, B.; Cheng, C.Y. Mechanistic Insights into PFOS-Mediated Sertoli Cell Injury. Trends Mol. Med. 2018, 24, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, X.; Liu, T.; Mo, M.; Ao, L.; Liu, J.; Cao, J.; Cui, Z. ZnSO4 rescued vimentin from collapse in DBP-exposed Sertoli cells by attenuating ER stress and apoptosis. Toxicol. In Vitro 2018, 48, 195–204. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, B.; Si, M.; Zou, H.; Song, R.; Gu, J.; Yuan, Y.; Liu, X.; Zhu, G.; Bai, J.; et al. Zearalenone altered the cytoskeletal structure via ER stress- autophagy- oxidative stress pathway in mouse TM4 Sertoli cells. Sci. Rep. 2018, 8, 3320. [Google Scholar] [CrossRef]
- Liu, L.; Chang, X.; Zhang, Y.; Wu, C.; Li, R.; Tang, L.; Zhou, Z. Fluorochloridone induces primary cultured Sertoli cells apoptosis: Involvement of ROS and intracellular calcium ions-mediated ERK1/2 activation. Toxicol. In Vitro 2018, 47, 228–237. [Google Scholar] [CrossRef]
- Sun, W.; Ni, Z.; Li, R.; Chang, X.; Li, W.; Yang, M.; Zhou, Z. Flurochloridone induces Sertoli cell apoptosis through ROS-dependent mitochondrial pathway. Ecotoxicol. Environ. Saf. 2021, 216, 112183. [Google Scholar] [CrossRef]
- Ni, Z.; Sun, W.; Li, R.; Yang, M.; Zhang, F.; Chang, X.; Li, W.; Zhou, Z. Fluorochloridone induces autophagy in TM4 Sertoli cells: Involvement of ROS-mediated AKT-mTOR signaling pathway. Reprod. Biol. Endocrinol. 2021, 19, 64. [Google Scholar] [CrossRef]
- Lebeaupin, C.; Proics, E.; de Bieville, C.H.; Rousseau, D.; Bonnafous, S.; Patouraux, S.; Adam, G.; Lavallard, V.J.; Rovere, C.; Le Thuc, O.; et al. ER stress induces NLRP3 inflammasome activation and hepatocyte death. Cell Death Dis. 2015, 6, e1879. [Google Scholar] [CrossRef] [Green Version]
- Coleman, O.I.; Haller, D. ER Stress and the UPR in Shaping Intestinal Tissue Homeostasis and Immunity. Front. Immunol. 2019, 10, 2825. [Google Scholar] [CrossRef]
- Hetz, C.; Saxena, S. ER stress and the unfolded protein response in neurodegeneration. Nat. Rev. Neurol. 2017, 13, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, P.; Jiang, R.; Xia, F.; Li, M.; Guo, F.J. Retraction Note to: Explore on the effect of ATF6 on cell growth and apoptosis in cartilage development. Histochem. Cell Biol. 2020, 154, 245. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, X.; Hao, J.; Zhu, Y.; Wang, Y.; Wang, L.; Guo, S.; Yi, H.; Liu, Y.; Liu, J. The role of ATF6 in Cr(VI)-induced apoptosis in DF-1 cells. J. Hazard Mater. 2021, 410, 124607. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Sun, H.; Li, J.; Shan, Y.; Zhao, Y.; Liao, F.; Yang, Y.; Cui, X.; Liu, Z. Involvement of CHOP in activin Ainduced myeloma NS1 cell apoptosis. Oncol. Rep. 2019, 42, 2644–2654. [Google Scholar] [CrossRef]
- Iurlaro, R.; Munoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puthalakath, H.; O’Reilly, L.A.; Gunn, P.; Lee, L.; Kelly, P.N.; Huntington, N.D.; Hughes, P.D.; Michalak, E.M.; McKimm-Breschkin, J.; Motoyama, N.; et al. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 2007, 129, 1337–1349. [Google Scholar] [CrossRef] [Green Version]
- Vanlaeys, A.; Dubuisson, F.; Seralini, G.E.; Travert, C. Formulants of glyphosate-based herbicides have more deleterious impact than glyphosate on TM4 Sertoli cells. Toxicol In Vitro 2018, 52, 14–22. [Google Scholar] [CrossRef]
- Li, Z.; Wang, H.; Huang, S.; Zhou, L.; Wang, L.; Du, C.; Wang, C. Establishment of stable MRP1 knockdown by lentivirus-delivered shRNA in the mouse testis Sertoli TM4 cell line. Toxicol. Mech. Methods 2015, 25, 81–90. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Q.; Wang, S.; Huang, T.; Li, X.; Ni, C.; Fang, Y.; Li, L.; Lian, Q.; Ge, R.S. Paraquat exposure delays stem/progenitor Leydig cell regeneration in the adult rat testis. Chemosphere 2019, 231, 60–71. [Google Scholar] [CrossRef]
- Sai, L.; Qu, B.; Zhang, J.; Liu, J.; Jia, Q.; Bo, C.; Zhang, Y.; Yu, G.; Han, R.; Peng, C. Analysis of long non-coding RNA involved in atrazine-induced testicular degeneration of Xenopus laevis. Environ. Toxicol. 2019, 34, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Krzastek, S.C.; Farhi, J.; Gray, M.; Smith, R.P. Impact of environmental toxin exposure on male fertility potential. Transl. Androl. Urol. 2020, 9, 2797–2813. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Li, R.; Hou, W.; Wang, Y.; Zhang, S.; Yu, Y.; Zhang, L.; Zhu, H.; Zhang, Z.; Fang, J.; et al. RNA-seq analysis of testes from flurochloridone-treated rats. Toxicol. Mech. Methods 2020, 30, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Martucciello, S.; Masullo, M.; Cerulli, A.; Piacente, S. Natural Products Targeting ER Stress, and the Functional Link to Mitochondria. Int. J. Mol. Sci. 2020, 21, 1905. [Google Scholar] [CrossRef] [Green Version]
- Shigemi, Z.; Manabe, K.; Hara, N.; Baba, Y.; Hosokawa, K.; Kagawa, H.; Watanabe, T.; Fujimuro, M. Methylseleninic acid and sodium selenite induce severe ER stress and subsequent apoptosis through UPR activation in PEL cells. Chem. Biol. Interact 2017, 266, 28–37. [Google Scholar] [CrossRef]
- Akazawa, Y.; Cazanave, S.; Mott, J.L.; Elmi, N.; Bronk, S.F.; Kohno, S.; Charlton, M.R.; Gores, G.J. Palmitoleate attenuates palmitate-induced Bim and PUMA up-regulation and hepatocyte lipoapoptosis. J. Hepatol. 2010, 52, 586–593. [Google Scholar] [CrossRef] [Green Version]
- Panganiban, R.A.; Park, H.R.; Sun, M.; Shumyatcher, M.; Himes, B.E.; Lu, Q. Genome-wide CRISPR screen identifies suppressors of endoplasmic reticulum stress-induced apoptosis. Proc. Natl. Acad. Sci. USA 2019, 116, 13384–13393. [Google Scholar] [CrossRef] [Green Version]
- Mota, M.; Banini, B.A.; Cazanave, S.C.; Sanyal, A.J. Molecular mechanisms of lipotoxicity and glucotoxicity in nonalcoholic fatty liver disease. Metabolism 2016, 65, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Kopp, M.C.; Larburu, N.; Durairaj, V.; Adams, C.J.; Ali, M. UPR proteins IRE1 and PERK switch BiP from chaperone to ER stress sensor. Nat. Struct. Mol. Biol. 2019, 26, 1053–1062. [Google Scholar] [CrossRef]
- Moncan, M.; Mnich, K.; Blomme, A.; Almanza, A.; Samali, A.; Gorman, A.M. Regulation of lipid metabolism by the unfolded protein response. J. Cell Mol. Med. 2021, 25, 1359–1370. [Google Scholar] [CrossRef]
- Read, A.; Schroder, M. The Unfolded Protein Response: An Overview. Biology (Basel) 2021, 10, 384. [Google Scholar] [CrossRef] [PubMed]
- Zyryanova, A.F.; Kashiwagi, K.; Rato, C.; Harding, H.P.; Crespillo-Casado, A.; Perera, L.A.; Sakamoto, A.; Nishimoto, M.; Yonemochi, M.; Shirouzu, M.; et al. ISRIB Blunts the Integrated Stress Response by Allosterically Antagonising the Inhibitory Effect of Phosphorylated eIF2 on eIF2B. Mol. Cell 2021, 81, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Rabouw, H.H.; Langereis, M.A.; Anand, A.A.; Visser, L.J.; de Groot, R.J.; Walter, P.; van Kuppeveld, F. Small molecule ISRIB suppresses the integrated stress response within a defined window of activation. Proc. Natl. Acad. Sci. USA 2019, 116, 2097–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugallo, R.; Marlin, E.; Baltanas, A.; Toledo, E.; Ferrero, R.; Vinueza-Gavilanes, R.; Larrea, L.; Arrasate, M.; Aragon, T. Fine tuning of the unfolded protein response by ISRIB improves neuronal survival in a model of amyotrophic lateral sclerosis. Cell Death Dis. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Teske, B.F.; Wek, S.A.; Bunpo, P.; Cundiff, J.K.; McClintick, J.N.; Anthony, T.G.; Wek, R.C. The eIF2 kinase PERK and the integrated stress response facilitate activation of ATF6 during endoplasmic reticulum stress. Mol. Biol. Cell 2011, 22, 4390–4405. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Ni, Z.; Zhao, S.; Wang, Y.; Chang, X.; Zhou, Z. Flurochloridone Induced Cell Apoptosis via ER Stress and eIF2α-ATF4/ATF6-CHOP-Bim/Bax Signaling Pathways in Mouse TM4 Sertoli Cells. Int. J. Environ. Res. Public Health 2022, 19, 4564. https://doi.org/10.3390/ijerph19084564
Zhang F, Ni Z, Zhao S, Wang Y, Chang X, Zhou Z. Flurochloridone Induced Cell Apoptosis via ER Stress and eIF2α-ATF4/ATF6-CHOP-Bim/Bax Signaling Pathways in Mouse TM4 Sertoli Cells. International Journal of Environmental Research and Public Health. 2022; 19(8):4564. https://doi.org/10.3390/ijerph19084564
Chicago/Turabian StyleZhang, Fen, Zhijing Ni, Shuqi Zhao, Yanna Wang, Xiuli Chang, and Zhijun Zhou. 2022. "Flurochloridone Induced Cell Apoptosis via ER Stress and eIF2α-ATF4/ATF6-CHOP-Bim/Bax Signaling Pathways in Mouse TM4 Sertoli Cells" International Journal of Environmental Research and Public Health 19, no. 8: 4564. https://doi.org/10.3390/ijerph19084564
APA StyleZhang, F., Ni, Z., Zhao, S., Wang, Y., Chang, X., & Zhou, Z. (2022). Flurochloridone Induced Cell Apoptosis via ER Stress and eIF2α-ATF4/ATF6-CHOP-Bim/Bax Signaling Pathways in Mouse TM4 Sertoli Cells. International Journal of Environmental Research and Public Health, 19(8), 4564. https://doi.org/10.3390/ijerph19084564