
Effects of Perch Essence Supplementation on Improving Exercise Performance and Anti-Fatigue in Mice


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals, and Experiment Design
2.3. Grip Strength
2.4. Swimming Exercise Endurance Test
2.5. Determination of Fatigue-Associated Biochemical Variables
2.6. Clinical Biochemical Profiles
2.7. Serum Myoglobin
2.8. Body Composition, Histopathology, and Glycogen Content
2.9. Statistical Analysis
3. Results
3.1. General Characteristics of Mice after SPSPE Supplementation for Four Weeks
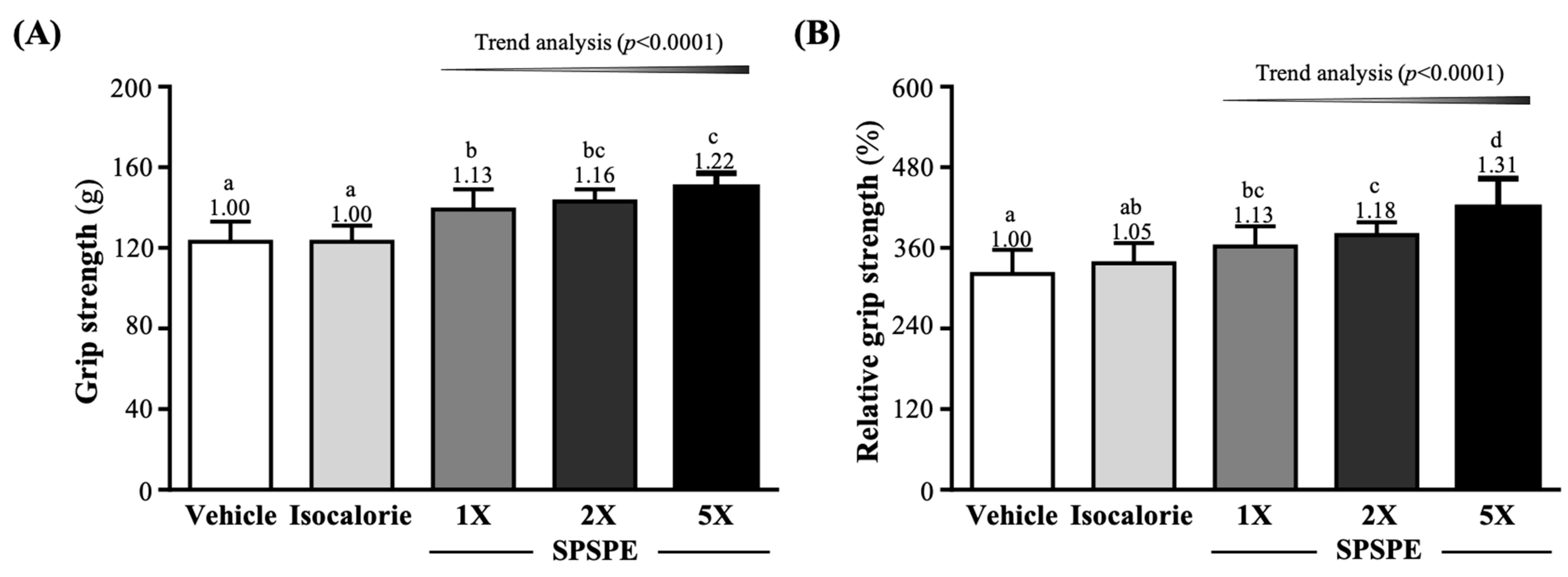
3.2. Effect of SPSPE Supplementation on Grip Strength
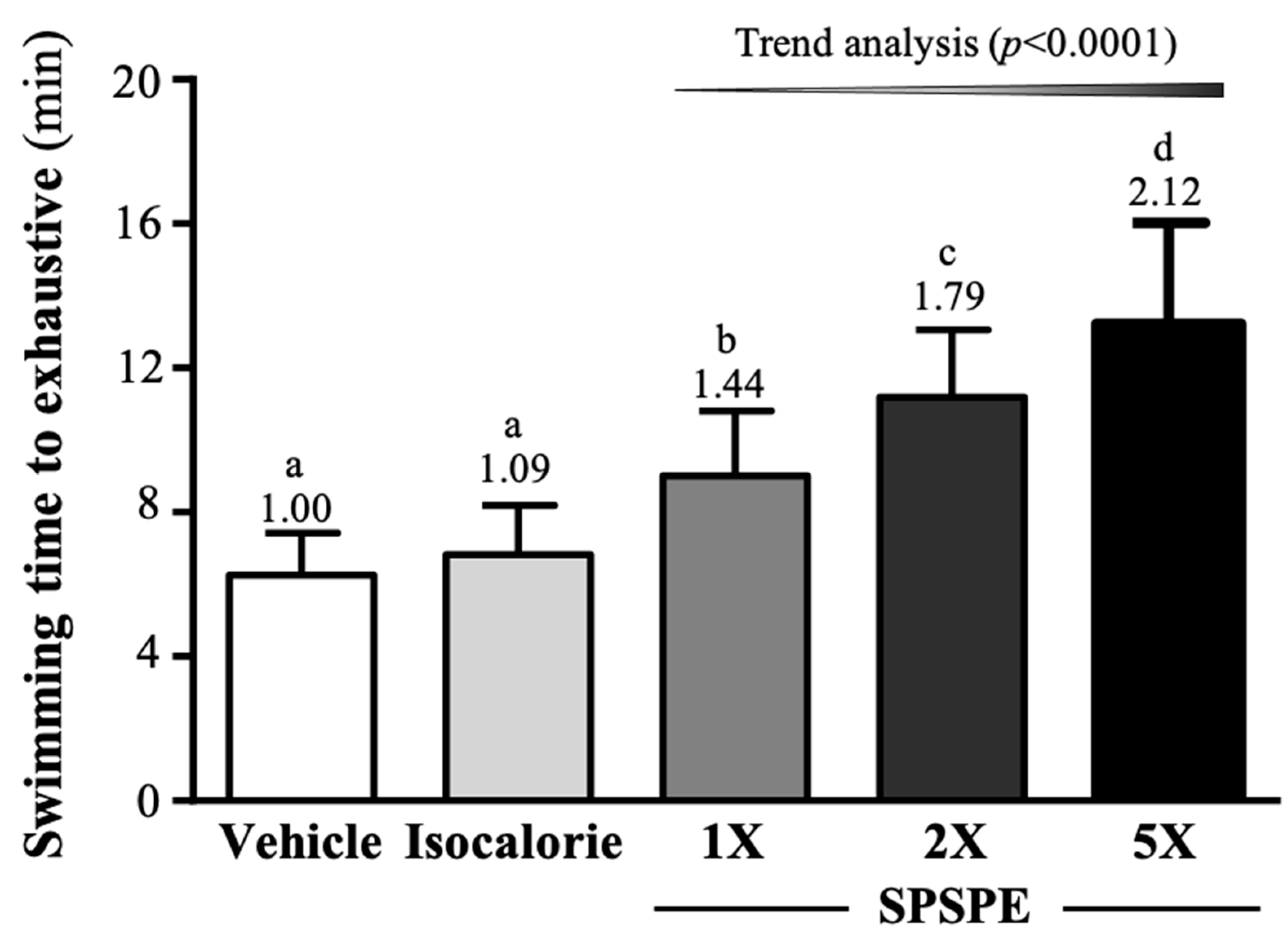
3.3. Effect of SPSPE Supplementation in the Exhaustive Swimming Test
3.4. Effect of SPSPE Supplementation on Serum Lactate Levels after the 10-min Swim Test
3.5. Effect of SPSPE Supplementation on Fatigue-Related Biochemical Indicators after the 10-min Swim Test or a 90-min Swim Test and 60-min Rest
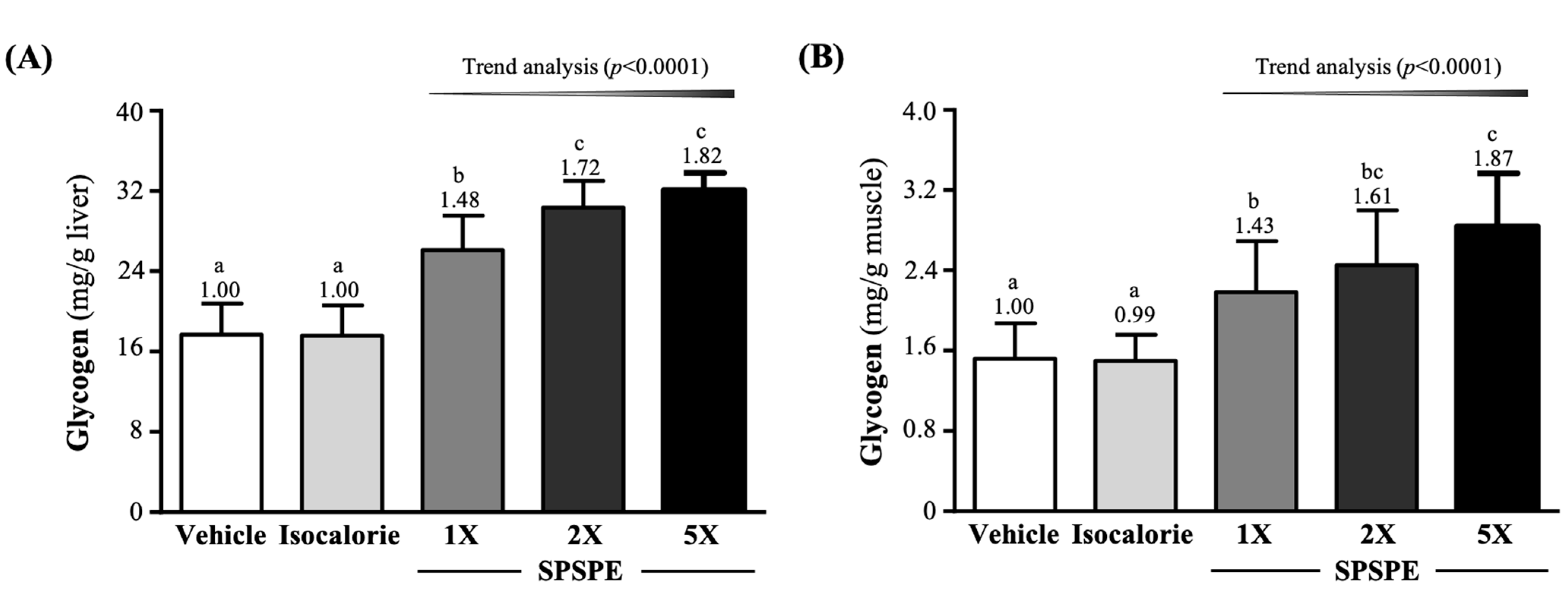
3.6. Effect of SPSPE Supplementation on Liver and Muscle Glycogen Levels
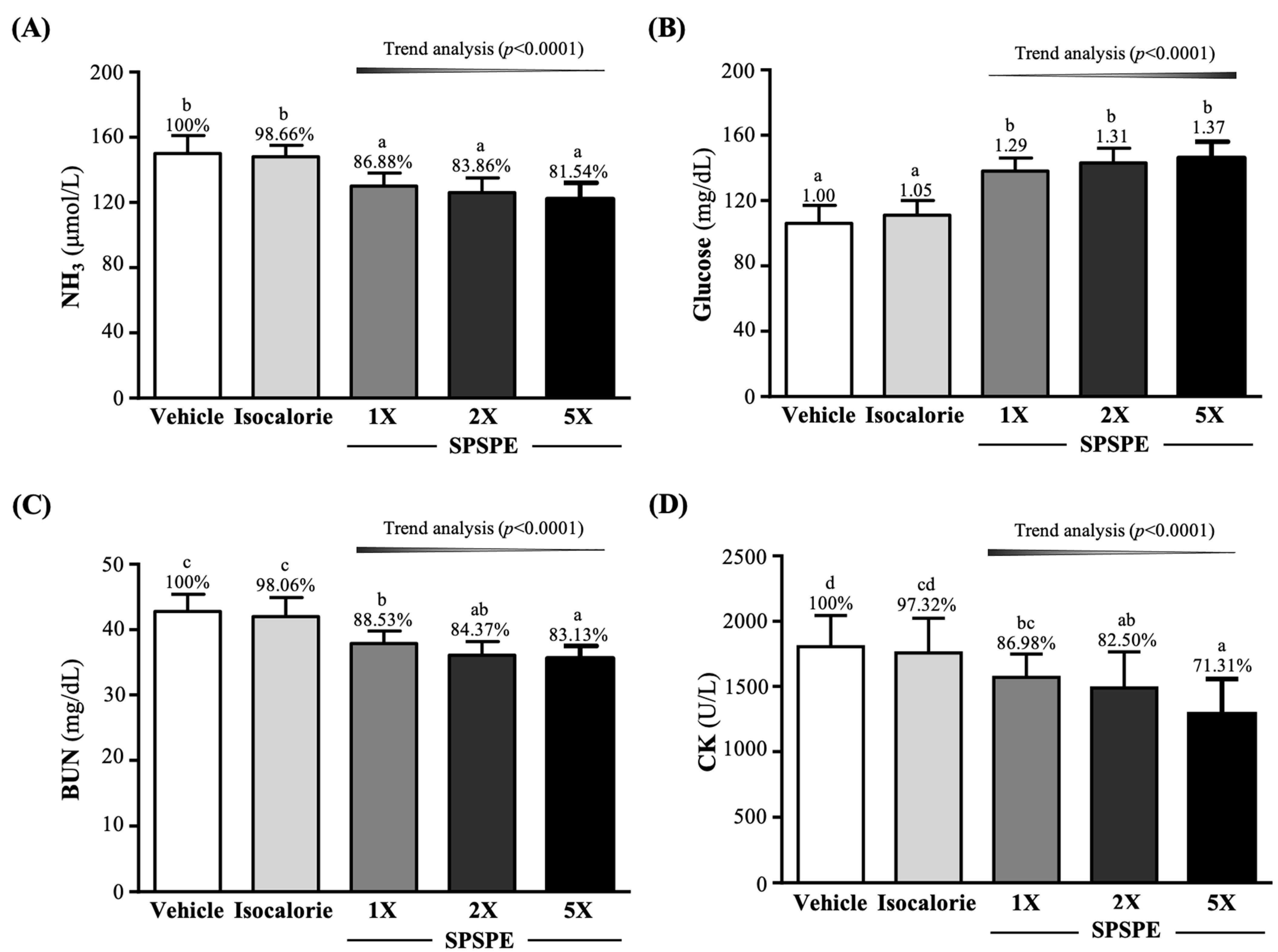
3.7. Effect of SPSPE Supplementation on Biochemical Variables at the End of the Experiment
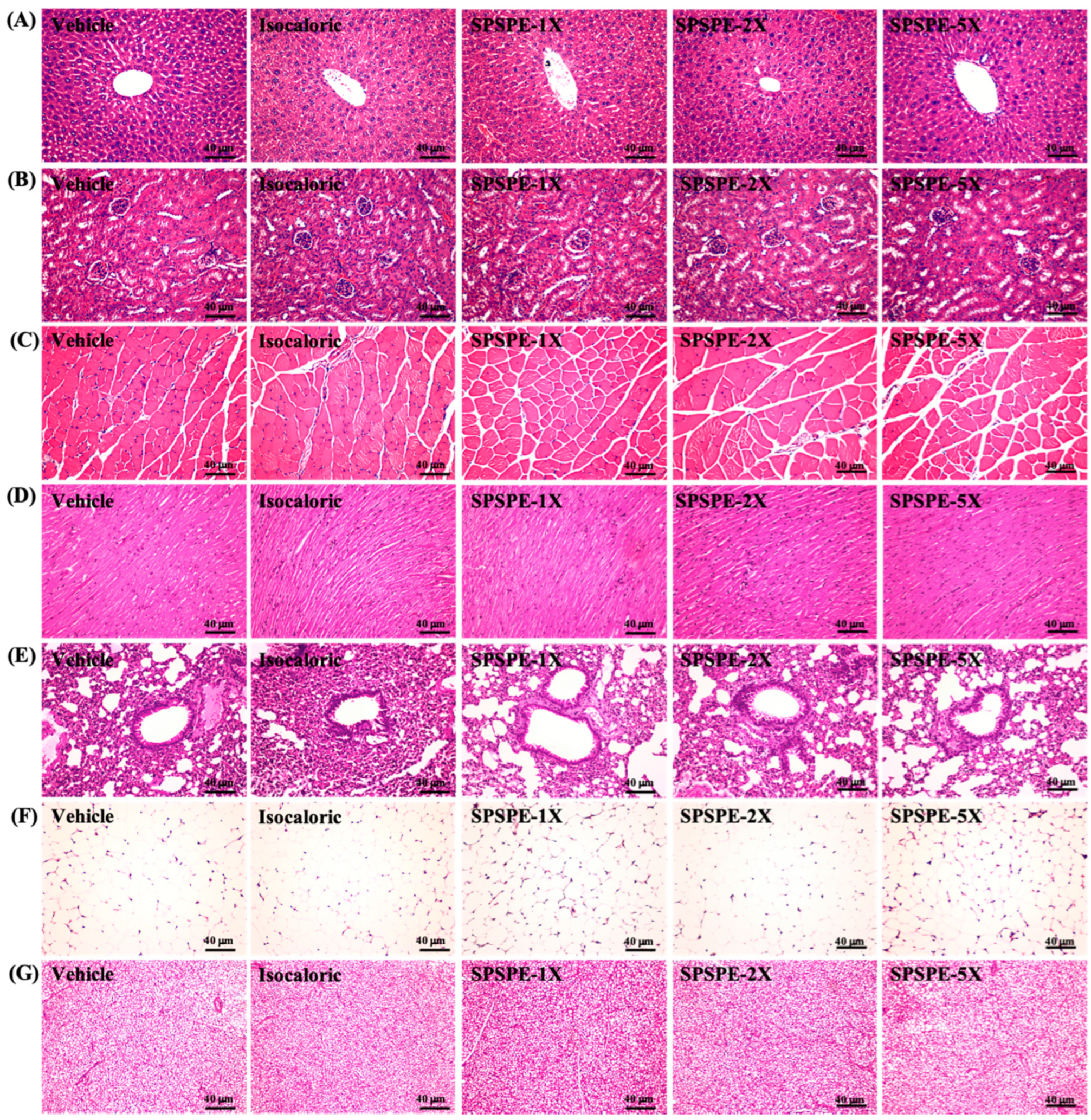
3.8. Effect of SPSPE Supplementation on Tissue Histology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weichselbaum, E.; Coe, S.; Buttriss, J.; Stanner, S. Fish in the diet: A review. Nutr. Bull. 2013, 38, 128–177. [Google Scholar] [CrossRef]
- Erkan, N.; Özden, Ö. Gutted and un-gutted sea bass (Dicentrarchus labrax) stored in ice: Influence on fish quality and shelf-life. Int. J. Food Prop. 2006, 9, 331–345. [Google Scholar] [CrossRef]
- Manninen, A.H. Protein hydrolysates in sports and exercise: A brief review. J. Sports Sci. Med. 2004, 3, 60–63. [Google Scholar]
- Cordeiro, E.M.; de Oliveira, G.V.; Volino-Souza, M.; Velozo, O.D.C.; Alvares, T.S. Effects of fish protein hydrolysate ingestion on postexercise aminoacidemia compared with whey protein hydrolysate in young individuals. J. Food Sci. 2020, 85, 21–27. [Google Scholar] [CrossRef]
- Oliveira, G.V.; Volino-Souza, M.; Cordeiro, E.M.; Conte-Junior, C.A.; Alvares, T.S. Effects of fish protein hydrolysate ingestion on endothelial function compared to whey protein hydrolysate in humans. Int. J. Food Sci. Nutr. 2020, 71, 242–248. [Google Scholar] [CrossRef]
- Lees, M.J.; Carson, B.P. The Potential Role of Fish-Derived Protein Hydrolysates on Metabolic Health, Skeletal Muscle Mass and Function in Ageing. Nutrients 2020, 12, 2434. [Google Scholar] [CrossRef]
- Lin, L.; Tao, N.; Su, H.; Zhang, J.; Zhong, J. Migration of nutrients and formation of micro/nano-sized particles in Atlantic salmon (Salmo salar) and bighead carp (Aristichthys nobilis) head soups. Food Biosci. 2020, 36, 100646. [Google Scholar] [CrossRef]
- Lekjing, S.; Venkatachalam, K.; Wangbenmad, C. Biochemical evaluation of novel seabass (Lates calcarifer) fish essence soup prepared by prolonged boiling process. Arab. J. Chem. 2021, 14, 103365. [Google Scholar] [CrossRef]
- Ament, W.; Verkerke, G.J. Exercise and fatigue. Sports Med. 2009, 39, 389–422. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Chiu, W.C.; Chuang, H.L.; Tang, D.W.; Lee, Z.M.; Wei, L.; Chen, F.A.; Huang, C.C. Effect of curcumin supplementation on physiological fatigue and physical performance in mice. Nutrients 2015, 7, 905–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.M.; Bailey, S.P. Possible mechanisms of central nervous system fatigue during exercise. Med. Sci. Sports Exerc. 1997, 29, 45–57. [Google Scholar] [CrossRef]
- Wan, J.J.; Qin, Z.; Wang, P.Y.; Sun, Y.; Liu, X. Muscle fatigue: General understanding and treatment. Exp. Mol. Med. 2017, 49, e384. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.L.; Lu, R.; Zhou, Y.J.; Ma, R.; Lv, J.Q.; Li, X.L.; Chen, L.J.; Yao, Z. The decapeptide CMS001 enhances swimming endurance in mice. Peptides 2008, 29, 1176–1182. [Google Scholar] [CrossRef]
- Burnley, M.; Jones, A.M. Power-duration relationship: Physiology, fatigue, and the limits of human performance. Eur. J. Sport Sci. 2018, 18, 1–12. [Google Scholar] [CrossRef]
- Huang, S.W.; Hsu, Y.J.; Lee, M.C.; Li, H.S.; Yeo, P.C.W.; Lim, A.L.; Huang, C.C. In Vitro and In Vivo Functional Characterization of Essence of Chicken as an Ergogenic Aid. Nutrients 2018, 10, 1943. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.C.; Lin, C.I.; Chiu, C.C.; Lin, Y.T.; Huang, W.K.; Huang, H.Y.; Huang, C.C. Chicken essence improves exercise performance and ameliorates physical fatigue. Nutrients 2014, 6, 2681–2696. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.C.; Hsu, Y.J.; Ho, H.H.; Kuo, Y.W.; Lin, W.Y.; Tsai, S.Y.; Chen, W.L.; Lin, C.L.; Huang, C.C. Effectiveness of human-origin Lactobacillus plantarum PL-02 in improving muscle mass, exercise performance and anti-fatigue. Sci. Rep. 2021, 11, 19469. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.C.; Hsu, Y.J.; Ho, H.H.; Hsieh, S.H.; Kuo, Y.W.; Sung, H.C.; Huang, C.C. Lactobacillus salivarius Subspecies salicinius SA-03 is a New Probiotic Capable of Enhancing Exercise Performance and Decreasing Fatigue. Microorganisms 2020, 8, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.C.; Hsu, Y.J.; Chuang, H.L.; Hsieh, P.S.; Ho, H.H.; Chen, W.L.; Chiu, Y.S.; Huang, C.C. In Vivo Ergogenic Properties of the Bifidobacterium longum OLP-01 Isolated from a Weightlifting Gold Medalist. Nutrients 2019, 11, 2003. [Google Scholar] [CrossRef] [Green Version]
- Gondal, A.Z.; Foris, L.A.; Richards, J.R. Serum Myoglobin; [Internet]; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Floros, J.D.; Newsome, R.; Fisher, W.; Barbosa-Cánovas, G.V.; Chen, H.; Dunne, C.P.; German, J.B.; Hall, R.L.; Heldman, D.R.; Karwe, M.V.; et al. Feeding the World Today and Tomorrow: The Importance of Food Science and Technology: An IFT Scientific Review. Compr. Rev. Food Sci. Food Saf. 2010, 9, 572–599. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2009, 37, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; López-Maside, L.; Donapetry-García, C.; Fernández-Fernández, C.; Sixto-Leal, C. Enzymes involved in branched-chain amino acid metabolism in humans. Amino Acids 2017, 49, 1005–1028. [Google Scholar] [CrossRef] [PubMed]
- Manoli, I.; Venditti, C.P. Disorders of branched chain amino acid metabolism. Transl. Sci. Rare Dis. 2016, 1, 91–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holeček, M. Branched-Chain Amino Acids and Branched-Chain Keto Acids in Hyperammonemic States: Metabolism and as Supplements. Metabolites 2020, 10, 324. [Google Scholar] [CrossRef]
- de Campos-Ferraz, P.L.; Ribeiro, S.M.; Luz Sdos, S.; Lancha, A.H., Jr.; Tirapegui, J. Exercise × BCAA Supplementation in Young Trained Rats: What Are Their Effects on Body Growth? J. Sports Sci. Med. 2011, 10, 483–490. [Google Scholar]
- Peyrollier, K.; Hajduch, E.; Blair, A.S.; Hyde, R.; Hundal, H.S. L-leucine availability regulates phosphatidylinositol 3-kinase, p70 S6 kinase and glycogen synthase kinase-3 activity in L6 muscle cells: Evidence for the involvement of the mammalian target of rapamycin (mTOR) pathway in the L-leucine-induced up-regulation of system A amino acid transport. Biochem. J. 2000, 350 Pt 2, 361–368. [Google Scholar]
- Monirujjaman, M.; Ferdouse, A. Metabolic and physiological roles of branched-chain amino acids. Adv. Mol. Biol. 2014, 2014, 364976. [Google Scholar] [CrossRef] [Green Version]
- Falavigna, G.; de Araújo, J.A., Jr.; Rogero, M.M.; Pires, I.S.; Pedrosa, R.G.; Martins, E., Jr.; Alves de Castro, I.; Tirapegui, J. Effects of diets supplemented with branched-chain amino acids on the performance and fatigue mechanisms of rats submitted to prolonged physical exercise. Nutrients 2012, 4, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualano, A.B.; Bozza, T.; Lopes De Campos, P.; Roschel, H.; Dos Santos Costa, A.; Luiz Marquezi, M.; Benatti, F.; Herbert Lancha Junior, A. Branched-chain amino acids supplementation enhances exercise capacity and lipid oxidation during endurance exercise after muscle glycogen depletion. J. Sports Med. Phys. Fit. 2011, 51, 82–88. [Google Scholar]
- Litwack, G. Chapter 8—Glycolysis and Gluconeogenesis; Academic Press: Boston, MA, USA, 2018; pp. 183–198. [Google Scholar]
- Williams, M. Dietary supplements and sports performance: Amino acids. J. Int. Soc. Sports Nutr. 2005, 2, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.C.; Hsu, Y.J.; Huang, C.C.; Liu, H.C.; Lee, M.C. Exercise Training Combined with Bifidobacterium longum OLP-01 Supplementation Improves Exercise Physiological Adaption and Performance. Nutrients 2020, 12, 1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, S.P. Lactic acid and exercise performance: Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, S.H.; Jeong, W.S.; Lee, H.Y. Effect of BCAA intake during endurance exercises on fatigue substances, muscle damage substances, and energy metabolism substances. J. Exerc. Nutr. Biochem. 2013, 17, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaoka, Y. McArdle Disease and Exercise Physiology. Biology 2014, 3, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Korzeniewski, B. AMP deamination delays muscle acidification during heavy exercise and hypoxia. J. Biol. Chem. 2006, 281, 3057–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.L.; Lee, M.C.; Hsu, Y.J.; Huang, W.C.; Huang, C.C.; Huang, S.W. Isolated Soy Protein Supplementation and Exercise Improve Fatigue-Related Biomarker Levels and Bone Strength in Ovariectomized Mice. Nutrients 2018, 10, 1792. [Google Scholar] [CrossRef] [Green Version]
- Coombes, J.S.; McNaughton, L.R. Effects of branched-chain amino acid supplementation on serum creatine kinase and lactate dehydrogenase after prolonged exercise. J. Sports Med. Phys. Fit. 2000, 40, 240–246. [Google Scholar]
- Wei, L.; Wen, Y.T.; Lee, M.C.; Ho, H.M.; Huang, C.C.; Hsu, Y.J. Effects of isolated soy protein and strength exercise training on exercise performance and biochemical profile in postpartum mice. Metabolism 2019, 94, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Jackman, S.R.; Witard, O.C.; Jeukendrup, A.E.; Tipton, K.D. Branched-chain amino acid ingestion can ameliorate soreness from eccentric exercise. Med. Sci. Sports Exerc. 2010, 42, 962–970. [Google Scholar] [CrossRef]
- Leenders, M.; Verdijk, L.B.; Van der Hoeven, L.; Van Kranenburg, J.; Nilwik, R.; Wodzig, W.K.; Senden, J.M.; Keizer, H.A.; Van Loon, L.J. Protein supplementation during resistance-type exercise training in the elderly. Med. Sci. Sports Exerc. 2013, 45, 542–552. [Google Scholar] [CrossRef]
- Uojima, H.; Sakurai, S.; Hidaka, H.; Kinbara, T.; Sung, J.H.; Ichita, C.; Tokoro, S.; Masuda, S.; Sasaki, A.; Koizumi, K.; et al. Effect of branched-chain amino acid supplements on muscle strength and muscle mass in patients with liver cirrhosis. Eur. J. Gastroenterol. Hepatol. 2017, 29, 1402–1407. [Google Scholar] [CrossRef] [PubMed]
- Kohno, K.; Hayakawa, F.; Xichang, W.; Shunsheng, C.; Yokoyama, M.; Kasai, M.; Takeutchi, F.; Hatae, K. Comparative study on flavor preference between Japanese and Chinese for dried bonito stock and chicken bouillon. J. Food Sci. 2005, 70, S193–S198. [Google Scholar] [CrossRef]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle sources: Meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampels, S. Towards a more sustainable production of fish as an important source for human nutrition. J. Fish. Livest. Prod. 2014, 2, 119. [Google Scholar] [CrossRef]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future Protein Supply and Demand: Strategies and Factors Influencing a Sustainable Equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrition Facts | /100 g SPSPE |
| Total calories | 376 kcal |
| Protein | 90.8 |
| Fat | 0.1 |
| Saturated fat | - |
| Trans fat | - |
| Carbohydrate | 2.9 |
| Sodium | 376 mg |
| Hydrolyzed Amino Acid Profiles | g/100 g |
| Leucine | 2.98 |
| Isoleucine | 1.33 |
| Valine | 2.04 |
| Cystine | 1.43 |
| tryptophan | 0.11 |
| methionine | 1.75 |
| threonine | 2.96 |
| Histidine | 1.18 |
| Tyrosine | 0.89 |
| Alanine | 8.45 |
| Glycine | 15.54 |
| Serine | 2.45 |
| Proline | 11.31 |
| Phenylalanine | 2.24 |
| Lysine | 3.43 |
| Arginine | 6.18 |
| Aspartic acid | 5.66 |
| Glutamic acid | 10.87 |
| Total BCAA (leucine, isoleucine, and valine) | 6.35 |
| Characteristics | Vehicle | Isocaloric | SPSPE-1X | SPSPE-2X | SPSPE-5X |
|---|---|---|---|---|---|
| Initial BW (g) | 29.6 ± 0.7 a | 29.7 ± 0.7 a | 29.8 ± 0.5 a | 30.0 ± 0.7 a | 30.0 ± 0.6 a |
| 1st week BW | 31.3 ± 0.8 a | 31.3 ± 1.0 a | 31.2 ± 1.0 a | 31.4 ± 0.9 a | 31.5 ± 1.3 a |
| 2nd week BW | 33.7 ± 0.9 a | 33.5 ± 0.9 a | 33.7 ± 1.2 a | 33.5 ± 1.5 a | 33.7 ± 1.3 a |
| 3rd week BW | 35.2 ± 0.7 a | 35.0 ± 1.2 a | 35.0 ± 1.2 a | 35.1 ± 1.5 a | 35.1 ± 1.5 a |
| 4th week BW | 36.0 ± 0.7 a | 36.1 ± 1.1a | 36.0 ± 1.6 a | 36.1 ± 1.8 a | 36.0 ± 1.6 a |
| 5th week BW | 37.0 ± 0.7 a | 37.0 ± 0.9 a | 37.0 ± 1.6 a | 37.2 ± 2.0 a | 37.0 ± 1.4 a |
| Final BW (g) | 37.5 ± 0.7 a | 37.7 ± 0.6 a | 37.7 ± 2.0 a | 37.8 ± 2.0 a | 37.6 ± 1.2 a |
| Water intake (mL/mouse/day) | 7.1 ± 0.3 a | 7.1 ± 0.3 a | 7.1 ± 0.4 a | 7.0 ± 0.2 a | 7.0 ± 0.3 a |
| Diet (g/mouse/day) | 6.6 ± 0.8 a | 6.6 ± 0.9 a | 6.6 ± 1.0 a | 6.6 ± 0.8 a | 6.6 ± 1.0 a |
| Calorie intake from diet (Chow 5001) (Kcal/mouse/day) (A) | 22.3 ± 2.9 a | 22.0 ± 3.0 a | 22.3 ± 3.4 a | 22.3 ± 2.8 a | 22.2 ± 3.3 a |
| Calorie intake from supplements (Kcal/mouse/day) (B) | 0.00 | 0.13 | 0.13 | 0.27 | 0.66 |
| Total daily calorie intake (Kcal/mouse/day) (A) + (B) | 22.3 ± 2.6 a | 22.2 ± 2.7 a | 22.5 ± 3.1 a | 22.6 ± 2.6 a | 22.9 ± 3.0 a |
| Liver (g) | 2.17 ± 0.18 a | 2.18 ± 0.17 a | 2.18 ± 0.14 a | 2.18 ± 0.07 a | 2.18 ± 0.08 a |
| Kidney (g) | 0.61 ± 0.04 a | 0.61 ± 0.07 a | 0.61 ± 0.04 a | 0.61 ± 0.05 a | 0.61 ± 0.07 a |
| Muscle (g) | 0.37 ± 0.02 a | 0.37 ± 0.03 a | 0.37 ± 0.02 a | 0.37 ± 0.02 a | 0.37 ± 0.04 a |
| Heart (g) | 0.22 ± 0.04 a | 0.21 ± 0.03 a | 0.21 ± 0.03 a | 0.22 ± 0.02 a | 0.21 ± 0.02 a |
| Lung (g) | 0.26 ± 0.03 a | 0.27 ± 0.02 a | 0.27 ± 0.03 a | 0.27 ± 0.03 a | 0.26 ± 0.02 a |
| EFP (g) | 0.35 ± 0.06 a | 0.35 ± 0.09 a | 0.36 ± 0.02 a | 0.36 ± 0.02 a | 0.35 ± 0.04 a |
| BAT (g) | 0.09 ± 0.02 a | 0.09 ± 0.01 a | 0.09 ± 0.02 a | 0.09 ± 0.01 a | 0.09 ± 0.02 a |
| Relative liver weight (%) | 5.79 ± 0.40 a | 5.79 ± 0.36 a | 5.78 ± 0.24 a | 5.78 ± 0.27 a | 5.79 ± 0.05 a |
| Relative kidney weight (%) | 1.62 ± 0.08 a | 1.63 ± 0.16 a | 1.62 ± 0.04 a | 1.61 ± 0.05 a | 1.62 ± 0.14 a |
| Relative muscle weight (%) | 0.99 ± 0.03 a | 0.98 ± 0.07 a | 0.97 ± 0.02 a | 0.98 ± 0.03 a | 0.97 ± 0.09 a |
| Relative heart weight (%) | 0.58 ± 0.09 a | 0.57 ± 0.06 a | 0.57 ± 0.05 a | 0.57 ± 0.03 a | 0.56 ± 0.04 a |
| Relative lung weight (%) | 0.69 ± 0.09 a | 0.71 ± 0.06 a | 0.71 ± 0.05 a | 0.70 ± 0.05 a | 0.70 ± 0.04 a |
| Relative EFP weight (%) | 0.92 ± 0.13 a | 0.93 ± 0.21 a | 0.95 ± 0.02 a | 0.95 ± 0.03 a | 0.93 ± 0.08 a |
| Relative BAT weight (%) | 0.24 ± 0.04 a | 0.24 ± 0.03 a | 0.23 ± 0.03 a | 0.24 ± 0.02 a | 0.25 ± 0.04 a |
| Time Point | Vehicle | Isocaloric | SPSPE-1X | SPSPE-2X | SPSPE-5X |
|---|---|---|---|---|---|
| Lactate (mmol/L) | |||||
| Before swimming (A) | 3.34 ± 0.38 a | 3.32 ± 0.32 a | 3.25 ± 0.37 a | 3.45 ± 0.29 a | 3.33 ± 0.36 a |
| After swimming (B) | 7.65 ± 0.57 c | 7.26 ± 0.50 c | 6.18 ± 0.71 b | 6.00 ± 0.72 ab | 5.54 ± 0.74 a |
| After a 20 min rest (C) | 6.39 ± 0.47 c | 6.09 ± 0.54 c | 4.92 ± 0.52 b | 4.76 ± 0.42 b | 4.27 ± 0.52 a |
| Rates of lactate production and clearance | |||||
| Production rate = B/A | 2.30 ± 0.10 e | 2.19 ± 0.09 d | 1.90 ± 0.06 c | 1.74 ± 0.08 b | 1.66 ± 0.06 a |
| Clearance rate = (B − C)/B | 0.16 ± 0.01 a | 0.16 ± 0.03 a | 0.20 ± 0.04 b | 0.21 ± 0.03 b | 0.23 ± 0.04 b |
| Vehicle | Isocaloric | SPSPE-1X | SPSPE-2X | SPSPE-5X | |
|---|---|---|---|---|---|
| AST (U/L) | 110 ± 18 a | 111 ± 15 a | 111 ± 14 a | 110 ± 12 a | 110 ± 16 a |
| ALT (U/L) | 58 ± 13 a | 57 ± 6 a | 57 ± 10 a | 58 ± 7 a | 58 ± 9 a |
| ALB (mg/dL) | 3.17 ± 0.13 a | 3.17 ± 0.08 a | 3.17 ± 0.12 a | 3.17 ± 0.14 a | 3.17 ± 0.24 a |
| BUN (mg/dL) | 23.4 ± 3.2 a | 23.8 ± 1.1 a | 23.7 ± 1.8 a | 23.3 ± 1.7 a | 23.2 ± 1.3 a |
| CREA (mg/dL) | 0.42 ± 0.04 a | 0.41 ± 0.04 a | 0.41 ± 0.03 a | 0.41 ± 0.03 a | 0.42 ± 0.05 a |
| UA (mg/dL) | 1.8 ± 0.9 a | 1.7 ± 0.5 a | 1.7 ± 0.5 a | 1.7 ± 0.3 a | 1.8 ± 0.3 a |
| TP (mg/dL) | 5.2 ± 0.5 a | 5.3 ± 0.1 a | 5.3 ± 0.2 a | 5.3 ± 0.3 a | 5.2 ± 0.3 a |
| TC (mg/dL) | 130 ± 14 a | 130 ± 13 a | 129 ± 9 a | 130 ± 12 a | 128 ± 20 a |
| TG (mg/dL) | 161 ± 16 a | 162 ± 22 a | 162 ± 25 a | 161 ± 11 a | 161 ± 28 a |
| CK (U/L) | 275 ± 34 a | 276 ± 40 a | 281 ± 21 a | 278 ± 33 a | 281 ± 37 a |
| Myoglobin (ng/mL) | 3.59 ± 0.47 a | 3.60 ± 0.82 a | 3.57 ± 0.59 a | 3.60 ± 0.66 a | 3.62 ± 0.46 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-C.; Hsu, Y.-J.; Lin, Y.-Q.; Chen, L.-N.; Chen, M.-T.; Huang, C.-C. Effects of Perch Essence Supplementation on Improving Exercise Performance and Anti-Fatigue in Mice. Int. J. Environ. Res. Public Health 2022, 19, 1155. https://doi.org/10.3390/ijerph19031155
Lee M-C, Hsu Y-J, Lin Y-Q, Chen L-N, Chen M-T, Huang C-C. Effects of Perch Essence Supplementation on Improving Exercise Performance and Anti-Fatigue in Mice. International Journal of Environmental Research and Public Health. 2022; 19(3):1155. https://doi.org/10.3390/ijerph19031155
Chicago/Turabian StyleLee, Mon-Chien, Yi-Ju Hsu, Yu-Qian Lin, Ling-Ni Chen, Mu-Tsung Chen, and Chi-Chang Huang. 2022. "Effects of Perch Essence Supplementation on Improving Exercise Performance and Anti-Fatigue in Mice" International Journal of Environmental Research and Public Health 19, no. 3: 1155. https://doi.org/10.3390/ijerph19031155
APA StyleLee, M.-C., Hsu, Y.-J., Lin, Y.-Q., Chen, L.-N., Chen, M.-T., & Huang, C.-C. (2022). Effects of Perch Essence Supplementation on Improving Exercise Performance and Anti-Fatigue in Mice. International Journal of Environmental Research and Public Health, 19(3), 1155. https://doi.org/10.3390/ijerph19031155