


5-Methyltetrahydrofolate Alleviates Memory Impairment in a Rat Model of Alzheimer’s Disease Induced by D-Galactose and Aluminum Chloride

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Treatments
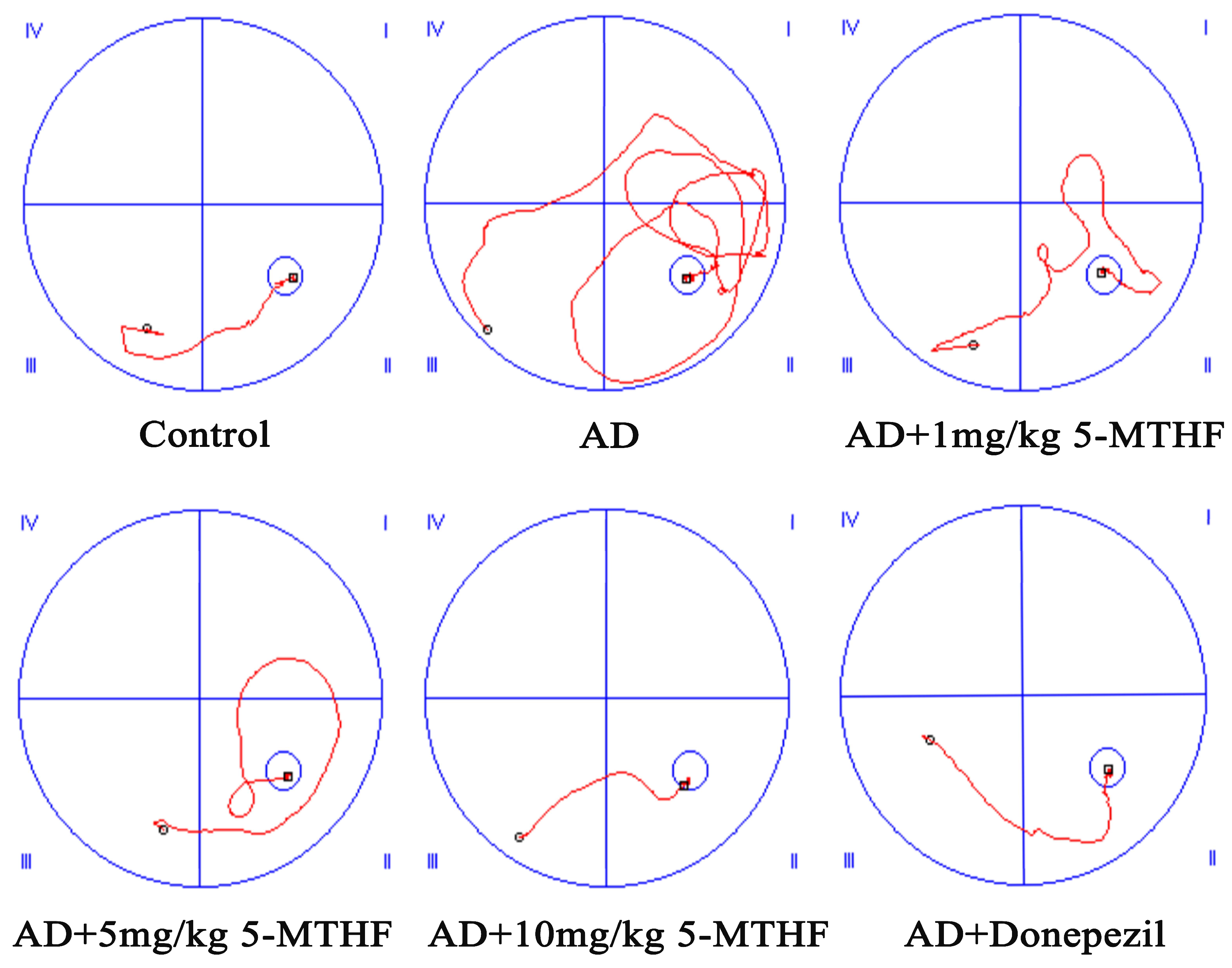
2.3. Morris Water Maze
2.4. Tissue Processing
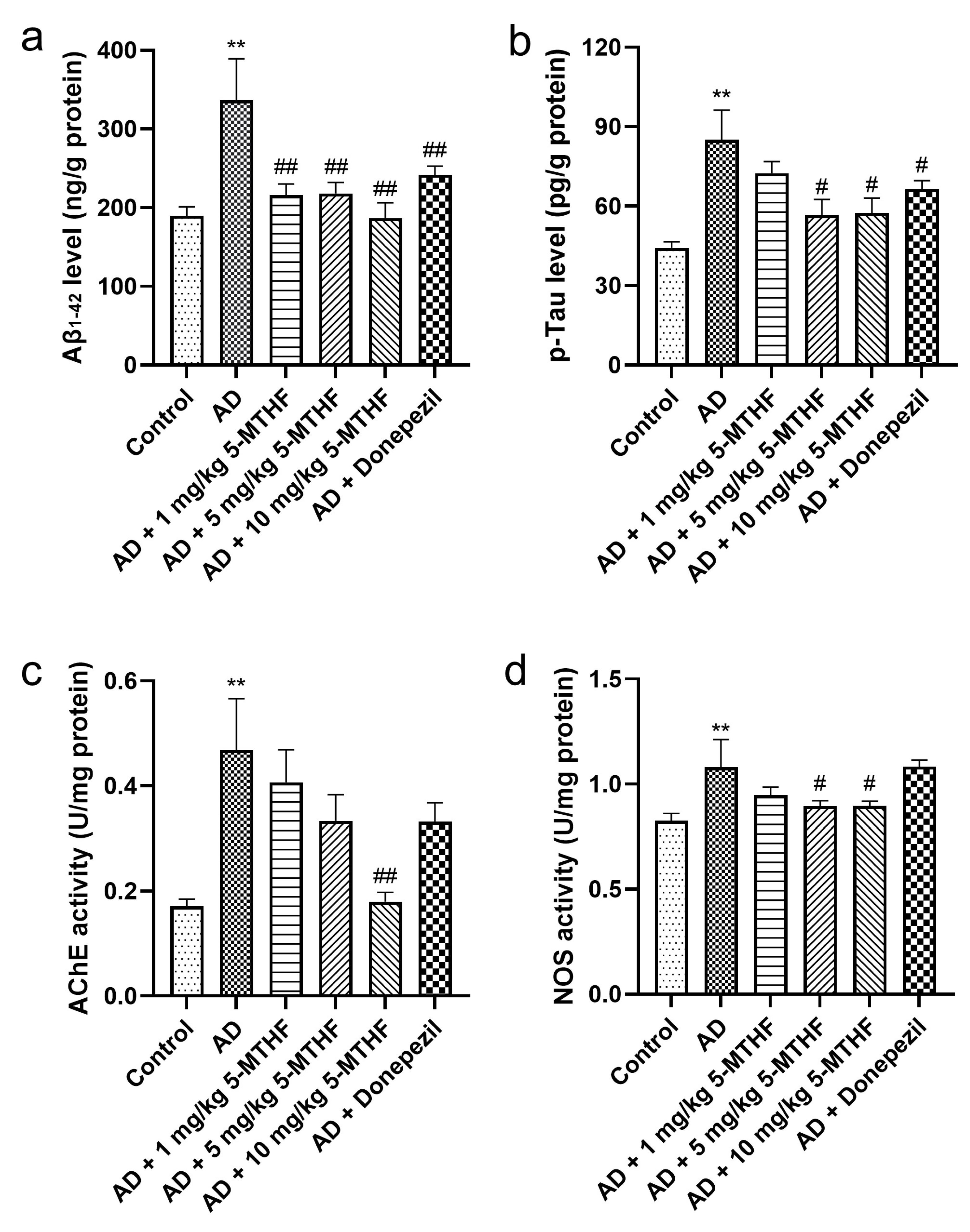
2.5. Determination of Aβ1-42, p-Tau, AChE and NOS
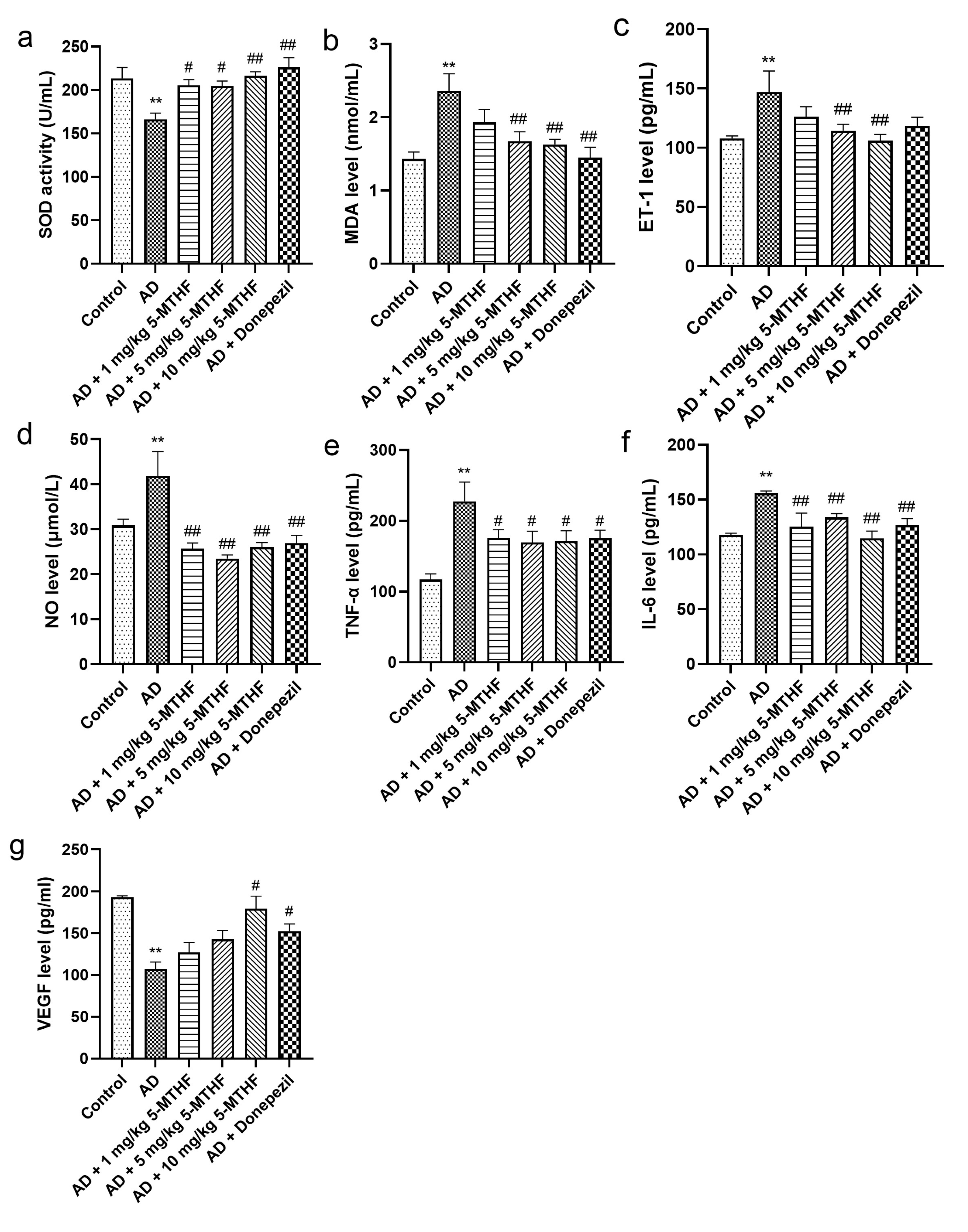
2.6. Determination of Serum Antioxidant and Cytokine Indices
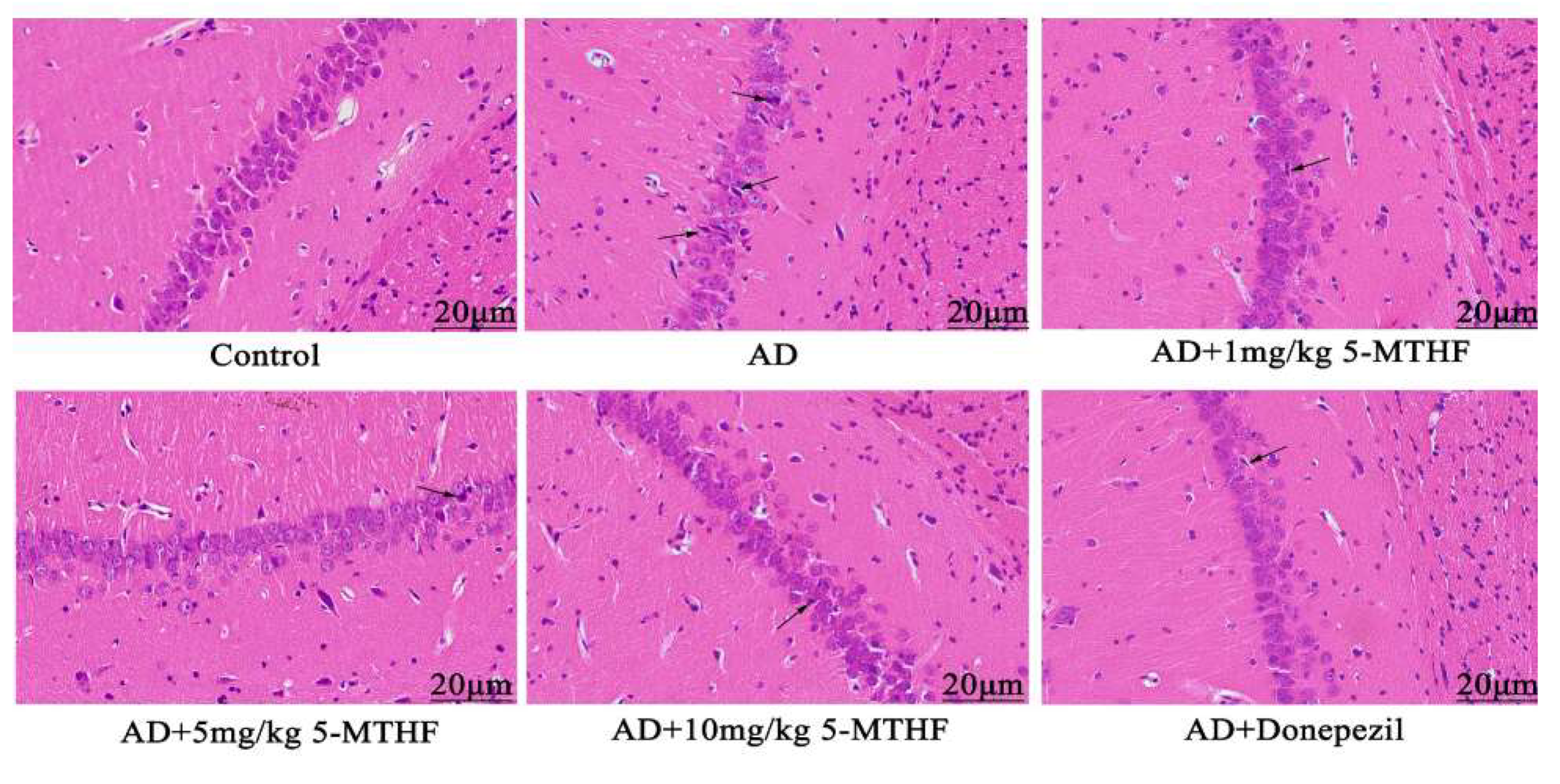
2.7. HE Staining of Hippocampus
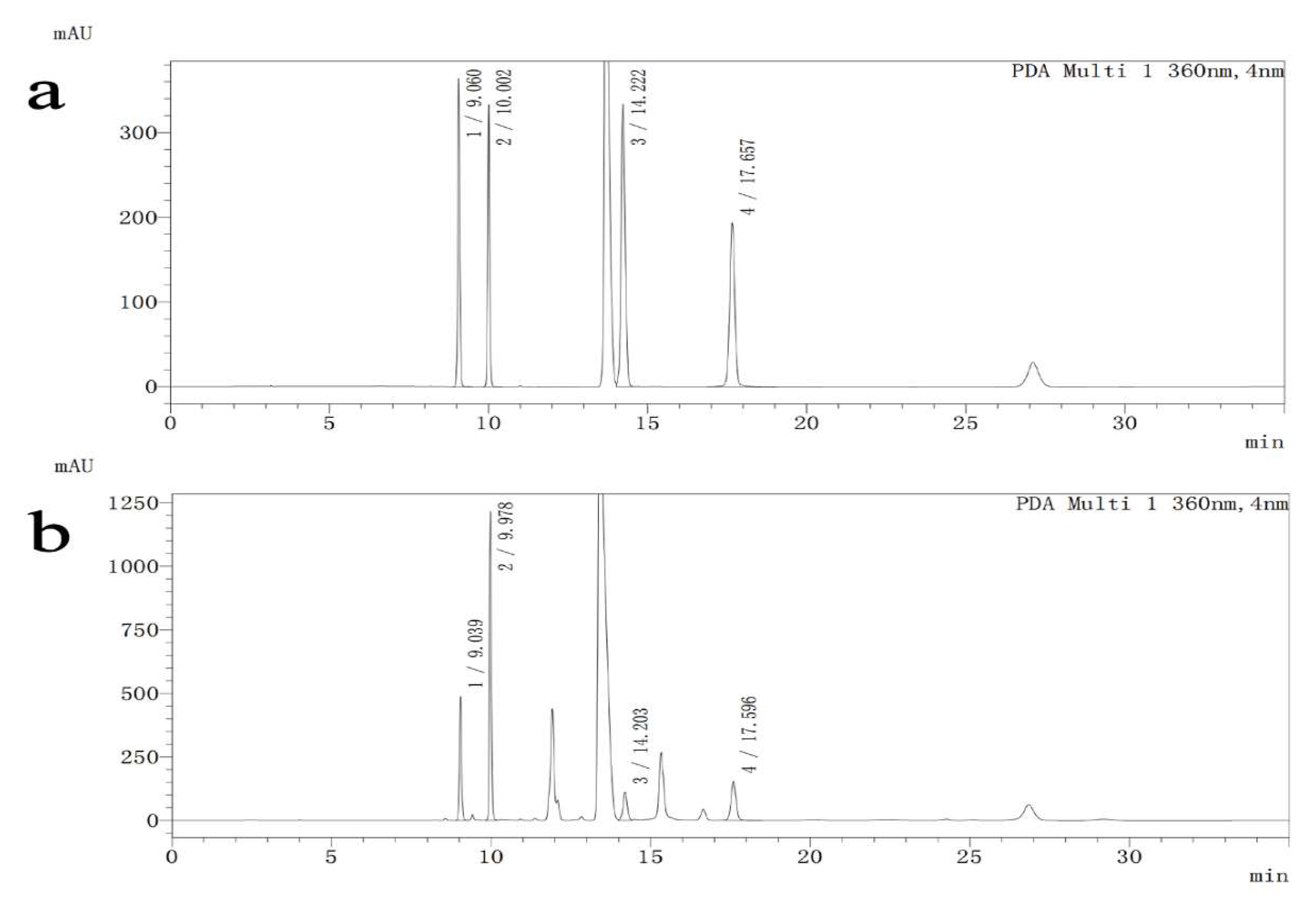
2.8. Determination of Brain Amino Acid Neurotransmitters
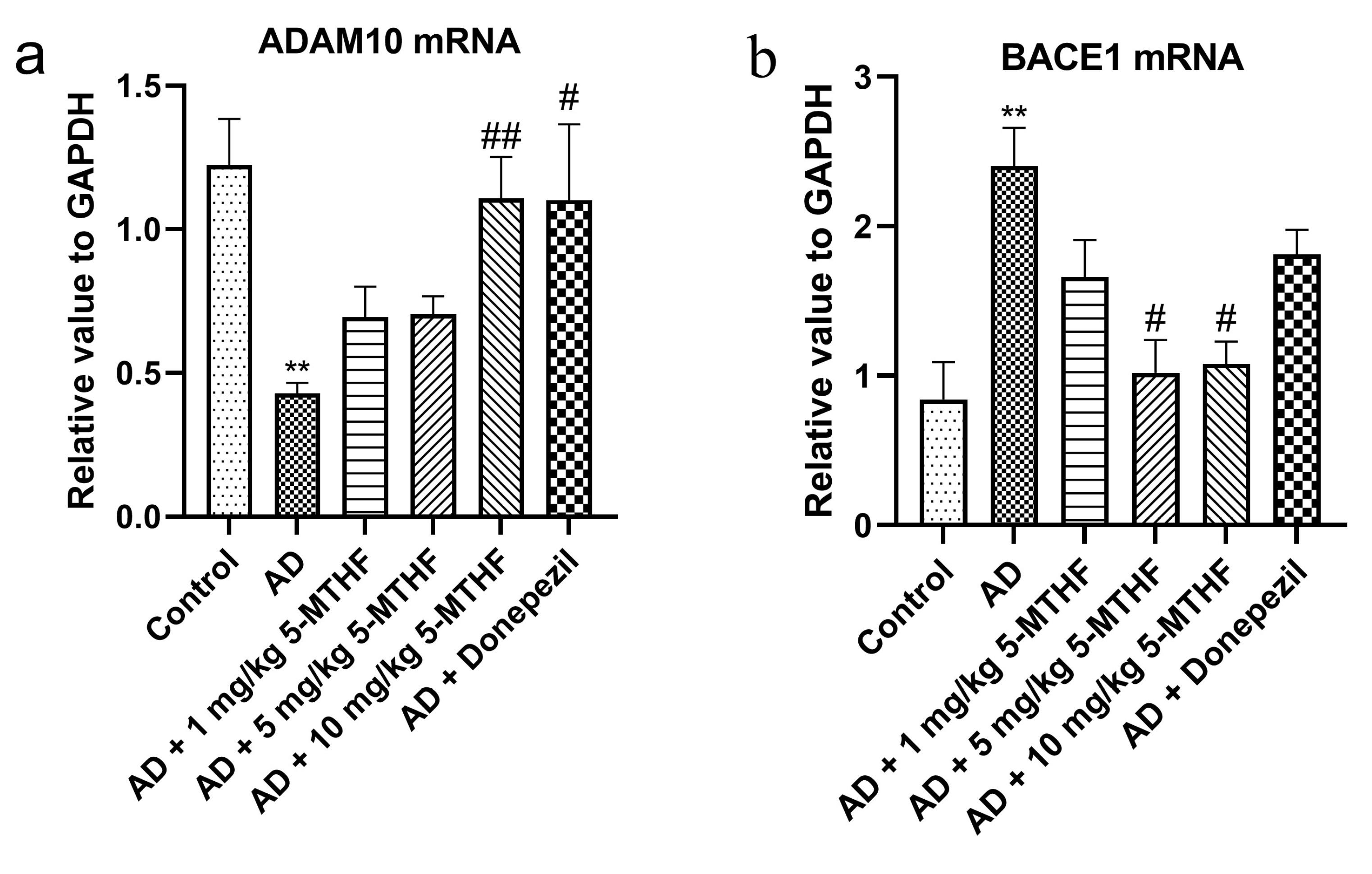
2.9. RT-PCR Determination of the Hippocampal Expression of ADAM10 and BACE1 mRNA
2.10. Statistical Analysis
3. Results
3.1. Effects of 5-MTHF on Memory Dysfunction in a Rat Model of AD
3.2. Effects of 5-MTHF on Brain Aβ1-42, p-Tau, AChE and NOS
3.3. Effects of 5-MTHF on Serum Antioxidant and Cytokine Indices
3.4. Effects of 5-MTHF on Hippocampal Neuronal Morphology
3.5. Effects of 5-MTHF on Neurotransmitter Levels
3.6. Effects of 5-MTHF on Hippocampal ADAM10 and BACE1 mRNA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Leszek, J.; Mikhaylenko, E.V.; Belousov, D.M.; Koutsouraki, E.; Szczechowiak, K.; Kobusiak-Prokopowicz, M.; Mysiak, A.; Diniz, B.S.; Somasundaram, S.G.; Kirkland, C.E.; et al. The links between cardiovascular diseases and Alzheimer’s disease. Curr. Neuropharmacol. 2021, 19, 152–169. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.A. Risk factors for Alzheimer’s disease. Folia Neuropathol. 2019, 57, 87–105. [Google Scholar] [CrossRef]
- Uddin, M.S.; Tewari, D.; Mamun, A.A.; Kabir, M.T.; Niaz, K.; Wahed, M.I.I.; Barreto, G.E.; Ashraf, G.M. Circadian and sleep dysfunction in Alzheimer’s disease. Ageing Res. Rev. 2020, 60, 101046. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, A.; Samra, Y.A.; Elsherbiny, N.M.; Al-Shabrawey, M. Implication of hyperhomocysteinemia in blood retinal barrier (BRB) dysfunction. Biomolecules 2020, 10, 1119. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.; Kim, Y.S.; Park, S.H. Identifying the relationship between leisure walking and prevalence of Alzheimer’s disease and other dementias. Int. J. Environ Res. Public Health 2022, 19, 8076. [Google Scholar] [CrossRef]
- Reitz, C.; Mayeux, R. Alzheimer disease: Epidemiology, diagnostic criteria, risk factors and biomarkers. Biochem. Pharmacol. 2014, 88, 640–651. [Google Scholar] [CrossRef]
- Vaz, M.; Silvestre, S. Alzheimer’s disease: Recent treatment strategies. Eur. J. Pharmacol. 2020, 887, 173554. [Google Scholar] [CrossRef]
- Shulpekova, Y.; Nechaev, V.; Kardasheva, S.; Sedova, A.; Kurbatova, A.; Bueverova, E.; Kopylov, A.; Malsagova, K.; Dlamini, J.C.; Ivashkin, V. The concept of folic acid in health and disease. Molecules 2021, 26, 3731. [Google Scholar] [CrossRef]
- Chen, H.; Liu, S.; Ji, L.; Wu, T.; Ji, Y.; Zhou, Y.; Zheng, M.; Zhang, M.; Xu, W.; Huang, G. Folic acid supplementation mitigates Alzheimer’s disease by reducing inflammation: A randomized controlled trial. Mediat. Inflamm. 2016, 2016, 5912146. [Google Scholar] [CrossRef]
- Tian, T.; Bai, D.; Li, W.; Huang, G.W.; Liu, H. Effects of folic acid on secretases involved in Aβ deposition in APP/PS1 mice. Nutrients 2016, 8, 556. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, J.; Chang, H.; Liu, X.; Zhu, R. Homocysteine and folic acid: Risk factors for Alzheimer’s disease-an updated meta-analysis. Front. Aging Neurosci. 2021, 13, 665114. [Google Scholar] [CrossRef]
- Lv, X.; Zhou, D.; Ge, B.; Chen, H.; Du, Y.; Liu, S.; Ji, Y.; Sun, C.; Wang, G.; Gao, Y.; et al. Association of folate metabolites and mitochondrial function in peripheral blood cells in Alzheimer’s disease: A matched case-control study. J. Alzheimer’s Dis. 2019, 70, 1133–1142. [Google Scholar] [CrossRef]
- McEneny, J.; Couston, C.; McKibben, B.; Young, I.S.; Woodside, J.V. Folate: In vitro and in vivo effects on VLDL and LDL oxidation. Int. J. Vitam. Nutr. Res. 2007, 77, 66–72. [Google Scholar] [CrossRef]
- Willems, F.F.; Boers, G.H.; Blom, H.J.; Aengevaeren, W.R.; Verheugt, F.W. Pharmacokinetic study on the utilisation of 5-methyltetrahydrofolate and folic acid in patients with coronary artery disease. Br. J. Pharmacol. 2004, 141, 825–830. [Google Scholar] [CrossRef]
- Papakostas, G.I.; Shelton, R.C.; Zajecka, J.M.; Etemad, B.; Rickels, K.; Clain, A.; Baer, L.; Dalton, E.D.; Sacco, G.R.; Schoenfeld, D.; et al. L-methylfolate as adjunctive therapy for SSRI-resistant major depression: Results of two randomized, double-blind, parallel-sequential trials. Am. J. Psychiatry 2012, 169, 1267–1274. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, G.; Qian, S.; Yang, Z.; Chen, X.; Chen, J.; Cai, C.; Liang, X.; Guo, J. Cerebrovascular protection of β-asarone in Alzheimer’s disease rats: A behavioral, cerebral blood flow, biochemical and genic study. J. Ethnopharmacol. 2012, 144, 305–312. [Google Scholar] [CrossRef]
- Abulfadl, Y.S.; El-Maraghy, N.N.; Ahmed, A.A.E.; Nofal, S.; Badary, O.A. Protective effects of thymoquinone on D-galactose and aluminum chloride induced neurotoxicity in rats: Biochemical, histological and behavioral changes. Neurol. Res. 2018, 40, 324–333. [Google Scholar] [CrossRef]
- Song, X.; Liu, B.; Cui, L.; Zhou, B.; Liu, W.; Xu, F.; Hayashi, T.; Hattori, S.; Ushiki-Kaku, Y.; Tashiro, S.I.; et al. Silibinin ameliorates anxiety/depression-like behaviors in amyloid β-treated rats by upregulating BDNF/TrkB pathway and attenuating autophagy in hippocampus. Physiol. Behav. 2017, 179, 487–493. [Google Scholar] [CrossRef]
- Wang, Y.J.; Wang, X.Y.; Hao, X.Y.; Yan, Y.M.; Hong, M.; Wei, S.F.; Zhou, Y.L.; Wang, Q.; Cheng, Y.X.; Liu, Y.Q. Ethanol extract of centipeda minima exerts antioxidant and neuroprotective effects via activation of the Nrf2 signaling pathway. Oxid. Med. Cell. Longev. 2019, 2019, 9421037. [Google Scholar] [CrossRef]
- Ali, A.; Shah, S.A.; Zaman, N.; Uddin, M.N.; Khan, W.; Ali, A.; Riaz, M.; Kamil, A. Vitamin D exerts neuroprotection via SIRT1/nrf-2/NF-kB signaling pathways against D-galactose-induced memory impairment in adult mice. Neurochem. Int. 2021, 142, 104893. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Liu, Y.; Wang, X.; Ma, H.; He, J.; Zhang, Y.; Yu, C.; Guan, W.; Ma, Y. The effects of amyloid-β42 oligomer on the proliferation and activation of astrocytes in vitro. In Vitro Cell. Dev. Biol. Anim. 2011, 47, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wu, H.; Yu, Y.; Qin, Y.; Han, X.; Hou, Y. Progesterone inhibits neuronal damage induced by Aβ-induced astrocyte activation. Chin. Pharmacol. Bull. 2014, 30, 1539–1543. [Google Scholar]
- Zhang, L.; Jin, C.; Lu, X.; Yang, J.; Wu, S.; Liu, Q.; Chen, R.; Bai, C.; Zhang, D.; Zheng, L.; et al. Aluminium chloride impairs long-term memory and downregulates cAMP-PKA-CREB signalling in rats. Toxicology 2014, 323, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, X.; Li, G.; Zhang, Y.; Wu, Y.; Song, W. Modifications and trafficking of APP in the pathogenesis of Alzheimer’s disease. Front. Mol. Neurosci. 2017, 10, 294. [Google Scholar] [CrossRef]
- Yu, H.; Yuan, B.; Chu, Q.; Wang, C.; Bi, H. Protective roles of isoastilbin against Alzheimer’s disease via Nrf2-mediated antioxidation and anti-apoptosis. Int. J. Mol. Med. 2019, 43, 1406–1416. [Google Scholar] [CrossRef]
- Yang, X.; Yuan, Y.; Niu, Q. Effects of aluminium chloride on the methylation of app in hippocampal of rats. J. Hyg. Res. 2016, 45, 345–349+355. [Google Scholar]
- Blurton-Jones, M.; Laferla, F.M. Pathways by which Abeta facilitates tau pathology. Curr. Alzheimer Res. 2006, 3, 437–448. [Google Scholar] [CrossRef]
- Nordberg, A. Mechanisms behind the neuroprotective actions of cholinesterase inhibitors in Alzheimer disease. Alzheimer Dis. Assoc. Disord. 2006, 20, S12–S18. [Google Scholar] [CrossRef]
- Peng, X.M.; Gao, L.; Huo, S.X.; Liu, X.M.; Yan, M. The mechanism of memory enhancement of acteoside (verbascoside) in the senescent mouse model induced by a combination of D-gal and AlCl3. Phytother. Res. 2015, 29, 1137–1144. [Google Scholar] [CrossRef]
- Akama, K.T.; Albanese, C.; Pestell, R.G.; Van Eldik, L.J. Amyloid beta-peptide stimulates nitric oxide production in astrocytes through an NFkappaB-dependent mechanism. Proc. Natl. Acad. Sci. USA 1998, 95, 5795–5800. [Google Scholar] [CrossRef]
- Tönnies, E.; Trushina, E. Oxidative stress, synaptic dysfunction, and Alzheimer’s disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Y.Q.; Fang, J.Y.; Li, P.; Li, F. Establishment of the concurrent experimental model of osteoporosis combined with Alzheimer’s disease in rat and the dual-effects of echinacoside and acteoside from Cistanche tubulosa. J. Ethnopharmacol. 2020, 257, 112834. [Google Scholar] [CrossRef]
- Grammas, P. Neurovascular dysfunction, inflammation and endothelial activation: Implications for the pathogenesis of Alzheimer’s disease. J. Neuroinflamm. 2011, 25, 26. [Google Scholar] [CrossRef]
- Nortley, R.; Korte, N.; Izquierdo, P.; Hirunpattarasilp, C.; Mishra, A.; Jaunmuktane, Z.; Kyrargyri, V.; Pfeiffer, T.; Khennouf, L.; Madry, C.; et al. Amyloid β oligomers constrict human capillaries in Alzheimer’s disease via signaling to pericytes. Science 2019, 365, eaav9518. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Fan, Y.C.; Wang, M.; Wang, D.; Li, X.H. Atorvastatin attenuates the production of IL-1β, IL-6, and TNF-α in the hippocampus of an amyloid β1-42-induced rat model of Alzheimer’s disease. Clin. Interv. Aging 2013, 8, 103–110. [Google Scholar]
- Zarezadehmehrizi, A.; Hong, J.; Lee, J.; Rajabi, H.; Gharakhanlu, R.; Naghdi, N.; Azimi, M.; Park, Y. Exercise training ameliorates cognitive dysfunction in amyloid beta-injected rat model: Possible mechanisms of Angiostatin/VEGF signaling. Metab. Brain. Dis. 2021, 36, 2263–2271. [Google Scholar] [CrossRef]
- Saha, P.; Guha, S.; Biswas, S.C. P38K and JNK pathways are induced by amyloid-β in astrocyte: Implication of MAPK pathways in astrogliosis in Alzheimer’s disease. Mol. Cell. Neurosci. 2020, 108, 103551. [Google Scholar] [CrossRef]
- Karunakaran, S.; Ravindranath, V. Activation of p38 MAPK in the substantia nigra leads to nuclear translocation of NF-kappaB in MPTP-treated mice: Implication in Parkinson’s disease. J. Neurochem. 2009, 109, 1791–1799. [Google Scholar] [CrossRef]
- Patel, N.S.; Mathura, V.S.; Bachmeier, C.; Beaulieu-Abdelahad, D.; Laporte, V.; Weeks, O.; Mullan, M.; Paris, D. Alzheimer’s beta-amyloid peptide blocks vascular endothelial growth factor mediated signaling via direct interaction with VEGFR-2. J. Neurochem. 2010, 112, 66–76. [Google Scholar] [CrossRef]
- Martin, L.; Bouvet, P.; Chounlamountri, N.; Watrin, C.; Besançon, R.; Pinatel, D.; Meyronet, D.; Honnorat, J.; Buisson, A.; Salin, P.A.; et al. VEGF counteracts amyloid-β-induced synaptic dysfunction. Cell. Rep. 2021, 35, 109121. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.L.; Ganaraja, B.; Murlimanju, B.V.; Joy, T.; Krishnamurthy, A.; Agrawal, A. Hippocampus and its involvement in Alzheimer’s disease: A review. 3 Biotech. 2022, 12, 55. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Hayes, L.; Soteriades, S.; Williams, G.; Brain, S.A.E.; Firbank, M.J.; Longoni, G.; Arnold, R.J.; Rowe, J.B.; O’Brien, J.T. Hippocampal stratum radiatum, lacunosum, and moleculare sparing in mild cognitive impairment. J. Alzheimer’s Dis. 2018, 61, 415–424. [Google Scholar] [CrossRef]
- Amber, S.; Sumera; Mirza, F.J.; Asif, M.; Hassan, D.; Ahmed, T.; Zahid, S. Amyloid-beta induced neurotoxicity impairs cognition and adult hippocampal neurogenesis in a mouse model for Alzheimer’s disease. Curr. Alzheimer Res. 2020, 17, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J. Hippocampal neurogenesis and pro-neurogenic therapies for Alzheimer’s disease. Animal Model. Exp. Med. 2022, 5, 3–14. [Google Scholar] [CrossRef]
- Walgrave, H.; Balusu, S.; Snoeck, S.; Vanden Eynden, E.; Craessaerts, K.; Thrupp, N.; Wolfs, L.; Horré, K.; Fourne, Y.; Ronisz, A.; et al. Restoring miR-132 expression rescues adult hippocampal neurogenesis and memory deficits in Alzheimer’s disease. Cell Stem Cell 2021, 28, 1805–1821.e8. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, D. Research progress of amino acid neurotransmitters and nervous system diseases and their therapeutic drugs. Acta Neuropharmacol. 2018, 8, 60. [Google Scholar]
- Patel, A.B.; Tiwari, V.; Veeraiah, P.; Saba, K. Increased astroglial activity and reduced neuronal function across brain in AβPP-PS1 mouse model of Alzheimer’s disease. J. Cereb. Blood Flow Metab. 2018, 38, 1213–1226. [Google Scholar] [CrossRef]
- Silva, M.V.F.; Loures, C.M.G.; Alves, L.C.V.; de Souza, L.C.; Borges, K.B.G.; Carvalho, M.D.G. Alzheimer’s disease: Risk factors and potentially protective measures. J. Biomed. Sci. 2019, 26, 33. [Google Scholar] [CrossRef]
- Wang, L.; Hu, J.; Zhao, Y.; Lu, X.; Zhang, Q.; Niu, Q. Effects of aluminium on β-amyloid (1-42) and secretases (APP-cleaving enzymes) in rat brain. Neurochem. Res. 2014, 39, 1338–1345. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, L.; Zhang, Z. ADAM10 and ADAM17 are degraded by lysosomal pathway via asparagine endopeptidase. Biochem. Biophys. Res. Commun. 2021, 537, 15–21. [Google Scholar] [CrossRef]
- Chen, L.; Na, R.; Gu, M.; Richardson, A.; Ran, Q. Lipid peroxidation up-regulates BACE1 expression in vivo: A possible early event of amyloidogenesis in Alzheimer’s disease. J. Neurochem. 2008, 107, 197–207. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Regression Equation | Correlation Coefficient |
|---|---|---|
| Asp | A = 24,412C − 18,411 | 0.9992 |
| Glu | A = 23,381C − 11,067 | 0.9991 |
| Gly | A = 65,655C − 107,277 | 0.9998 |
| γ-GABA | A = 46,910C − 91,513 | 0.9992 |
| Group | Asp | Glu | Gly | γ-GABA |
|---|---|---|---|---|
| Control | 71.27 ± 2.04 | 235.50 ± 6.86 | 21.62 ± 1.51 | 53.97 ± 2.73 |
| AD | 86.71 ± 1.30 ** | 188.18 ± 6.38 ** | 15.76 ± 0.43 * | 31.80 ± 1.46 ** |
| AD + 1 mg/kg 5-MTHF | 78.08 ± 2.76 | 214.19 ± 6.76 | 16.67 ± 0.73 | 40.40 ± 2.64 |
| AD + 5 mg/kg 5-MTHF | 71.32 ± 2.45 ## | 211.58 ± 8.70 | 16.00 ± 0.66 | 41.21 ± 2.59 |
| AD + 10 mg/kg 5-MTHF | 80.88 ± 3.28 | 240.80 ± 14.94 ## | 17.91 ± 1.39 | 41.30 ± 5.54 |
| AD + Donepezil | 74.67 ± 2.90 # | 204.52 ± 8.13 | 16.78 ± 0.47 | 42.21 ± 2.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wu, H.; Qi, S.; Tang, Y.; Qin, C.; Liu, R.; Zhang, J.; Cao, Y.; Gao, X. 5-Methyltetrahydrofolate Alleviates Memory Impairment in a Rat Model of Alzheimer’s Disease Induced by D-Galactose and Aluminum Chloride. Int. J. Environ. Res. Public Health 2022, 19, 16426. https://doi.org/10.3390/ijerph192416426
Zhang Z, Wu H, Qi S, Tang Y, Qin C, Liu R, Zhang J, Cao Y, Gao X. 5-Methyltetrahydrofolate Alleviates Memory Impairment in a Rat Model of Alzheimer’s Disease Induced by D-Galactose and Aluminum Chloride. International Journal of Environmental Research and Public Health. 2022; 19(24):16426. https://doi.org/10.3390/ijerph192416426
Chicago/Turabian StyleZhang, Zhengduo, Hong Wu, Shaojun Qi, Yanjin Tang, Chuan Qin, Rui Liu, Jiacheng Zhang, Yiyao Cao, and Xibao Gao. 2022. "5-Methyltetrahydrofolate Alleviates Memory Impairment in a Rat Model of Alzheimer’s Disease Induced by D-Galactose and Aluminum Chloride" International Journal of Environmental Research and Public Health 19, no. 24: 16426. https://doi.org/10.3390/ijerph192416426
APA StyleZhang, Z., Wu, H., Qi, S., Tang, Y., Qin, C., Liu, R., Zhang, J., Cao, Y., & Gao, X. (2022). 5-Methyltetrahydrofolate Alleviates Memory Impairment in a Rat Model of Alzheimer’s Disease Induced by D-Galactose and Aluminum Chloride. International Journal of Environmental Research and Public Health, 19(24), 16426. https://doi.org/10.3390/ijerph192416426