Author Contributions
A.S.R. assembled the data: designed and conducted the analyses, and wrote the first manuscript draft. G.K.H. provided technical and logistic support, co-wrote the paper, assisted with gaining ethical approval, provided advice on manuscript preparation and general guidance to study conduct. A.S.R. had the idea for the article, performed the literature search, wrote the first draft and is the guarantor for the article. All authors have read and agreed to the published version of the manuscript.
Figure 1.
Map-graph of log(testicular cancer incidence rates) across USA by state and year.
Figure 1.
Map-graph of log(testicular cancer incidence rates) across USA by state and year.
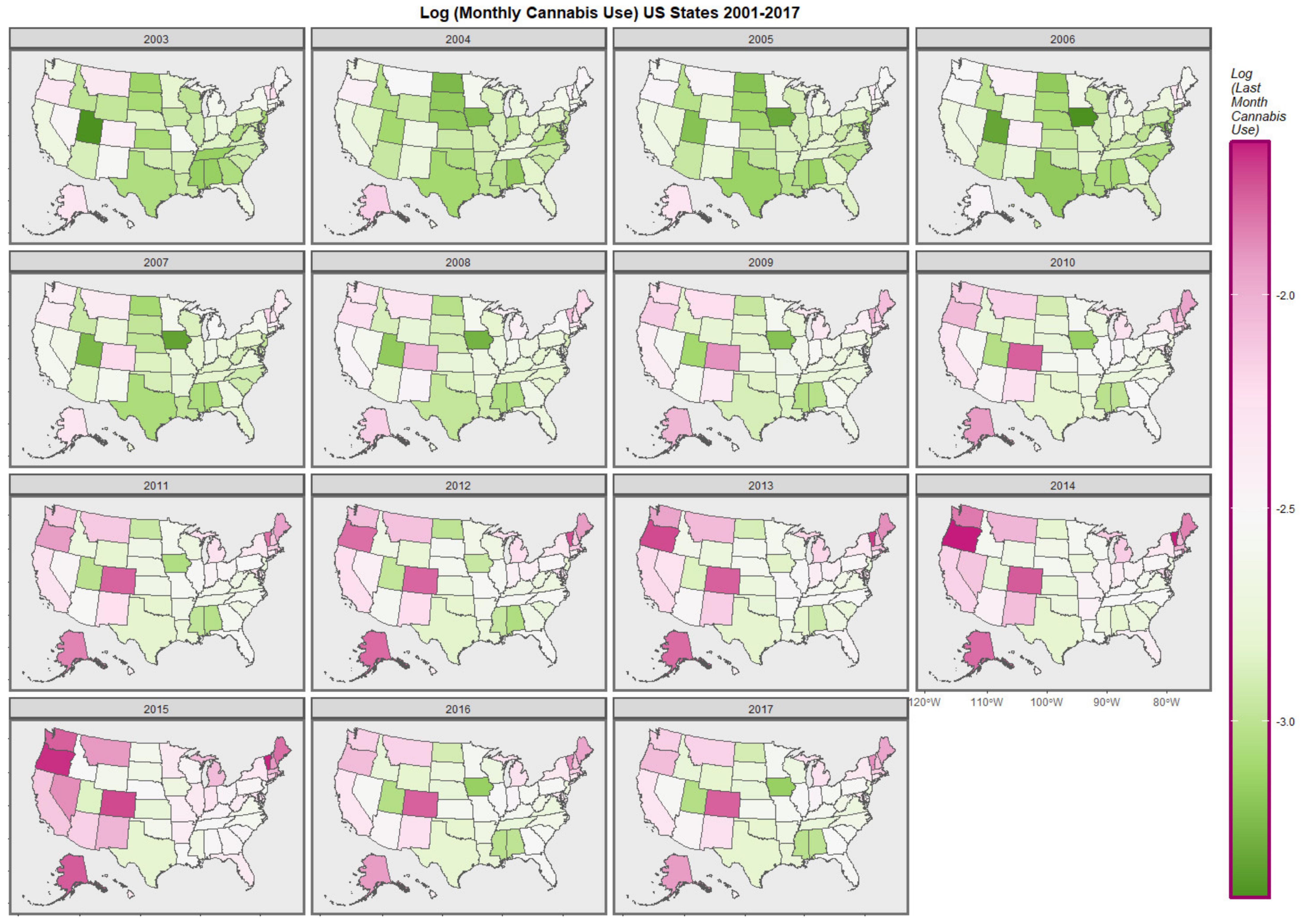
Figure 2.
Map-graph of log(last month cannabis use rates) across USA by state and year.
Figure 2.
Map-graph of log(last month cannabis use rates) across USA by state and year.
Figure 3.
Testicular cancer incidence rates by substance exposure. Substance exposure is listed as the fraction of the population reporting the applicable exposures. Median household income is reported as annual income in US dollars.
Figure 3.
Testicular cancer incidence rates by substance exposure. Substance exposure is listed as the fraction of the population reporting the applicable exposures. Median household income is reported as annual income in US dollars.
Figure 4.
Testicular cancer incidence rates by cannabinoid exposure.
Figure 4.
Testicular cancer incidence rates by cannabinoid exposure.
Figure 5.
Time course of drug use across USA.
Figure 5.
Time course of drug use across USA.
Figure 6.
Time course of drug use across USA by cannabis legal status—scatterplots.
Figure 6.
Time course of drug use across USA by cannabis legal status—scatterplots.
Figure 7.
Substance use across USA by cannabis legal status—boxplots with aggregated time. Note that where the notches on the boxplots do not overlap this signifies a statistically significant difference.
Figure 7.
Substance use across USA by cannabis legal status—boxplots with aggregated time. Note that where the notches on the boxplots do not overlap this signifies a statistically significant difference.
Figure 8.
Cannabis use and testicular cancer incidence rates by quintiles of cannabis use. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) boxplots. (C,D) scatterplots with regression lines. Note that where the notches on the boxplots do not overlap this signifies a statistically significant difference.
Figure 8.
Cannabis use and testicular cancer incidence rates by quintiles of cannabis use. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) boxplots. (C,D) scatterplots with regression lines. Note that where the notches on the boxplots do not overlap this signifies a statistically significant difference.
Figure 9.
Cannabis use and testicular cancer incidence rates by dichotomized quintiles of cannabis use. Dichotomy contrasts highest cannabis use quintile with the lower four quintiles. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) scatterplots with regression lines. (C,D) boxplots.
Figure 9.
Cannabis use and testicular cancer incidence rates by dichotomized quintiles of cannabis use. Dichotomy contrasts highest cannabis use quintile with the lower four quintiles. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) scatterplots with regression lines. (C,D) boxplots.
Figure 10.
Heatmap of testicular cancer rates by state. Note Hawaii near the top which is an obvious standout hotspot.
Figure 10.
Heatmap of testicular cancer rates by state. Note Hawaii near the top which is an obvious standout hotspot.
Figure 11.
Map of geospatial neighbour links use din spatial regressions (A) edited and (B) final.
Figure 11.
Map of geospatial neighbour links use din spatial regressions (A) edited and (B) final.
Figure 12.
Cannabis use and testicular cancer incidence rates by cannabis legal status. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) boxplots. (C,D) scatterplots with regression lines.
Figure 12.
Cannabis use and testicular cancer incidence rates by cannabis legal status. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) boxplots. (C,D) scatterplots with regression lines.
Figure 13.
Cannabis use and testicular cancer incidence rates by dichotomized cannabis legal status. Dichotomy contrasts states where cannabis is illegal vs. others. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) scatterplots with regression lines. (C,D) boxplots with notches.
Figure 13.
Cannabis use and testicular cancer incidence rates by dichotomized cannabis legal status. Dichotomy contrasts states where cannabis is illegal vs. others. (A,C) cannabis use. (B,D) testicular cancer rates. (A,B) scatterplots with regression lines. (C,D) boxplots with notches.
Table 1.
Introductory Linear Regressions.
Table 1.
Introductory Linear Regressions.
| Parameter Estimates | Model Parameters |
|---|
| Term and Model | Estimate (C.I.) | p-Value | S.D. | R-Squared | F | dF | p |
|---|
| lm(Rate~Time) | | | | | | | |
| Year—Caucasian-Americans | 0.08 (0.07, 0.09) | 7.80 × 10−23 | 0.3452 | 0.8988 | 374 | 1.41 | 3.20 × 10−22 |
| Year—African Americans | 0.011 (−0.001, 0.024) | 0.0945 | 0.2184 | 0.0820 | 3.054 | 1.22 | 0.0945 |
| lm(Rate~mrjmon) | | | | | | | |
| Cannabis | 0.47 (0.34, 0.59) | 7.50 × 10−13 | 0.5929 | 0.0652 | 53.25 | 1.748 | 7.49 × 10−13 |
| lm(Rate~Time * mrjmon) | | | | | |
| Cannabis | 0.47 (0.34, 0.59) | 7.50 × 10−13 | 0.5928 | 0.0652 | 53.25 | 1.748 | 7.50 × 10−13 |
| Additive model | | | | | | | |
| lm(Rate~Cigarettes + AUD + Cannabis + Analgesics + Cocaine) |
| AUD | 14.82 (12.14, 17.51) | 6.46 × 10−44 | 0.5336 | 0.2428 | 49.04 | 5.744 | 6.46 × 10−44 |
| Cannabis | 0.45 (0.32, 0.57) | 7.24 × 10−12 | | | | | |
| Cocaine | −14.05 (−20.36, −7.74) | 1.45 × 10−5 | | | | | |
| Analgesics | −10.22 (−14.69, −5.76) | 8.43 × 10−6 | | | | | |
| Cigarettes | −3.19 (−4.21, −2.17) | 1.64 × 10−9 | | | | | |
| Quintiles | | | | | | | |
| lm(Rate~Quintile) | | | | | | |
| Quintile 5 | 0.17 (0.13, 0.21) | 5.2 × 10−14 | 0.1927 | 0.1145 | 25.22 | 4.745 | 1.29 × 10−19 |
| lm(Rate~Year * Quintile) | | | | | | |
| Year | 0.0038 (0.0007, 0.007) | 0.0184 | 0.1922 | 0.1199 | 21.42 | 5.744 | 4.66 × 10−20 |
| Year: Quintile 5 | 0.17 (0.13, 0.21) | 4.21 × 10−14 | | | | | |
| lm(Rate~Year * Quintiles_Dichotomized) | | | | | |
| Year | 0.0038 (0.0006, 0.007) | 0.0202 | 0.1925 | 0.1165 | 50.4 | 2.747 | 2.94 × 10−21 |
| Upper Quintiles | 0.17 (0.14, 0.21) | 1.37 × 10−21 | | | | | |
| Substances | | | | | | | |
| lm(Rate~Substances) | | | | | | |
| Cigarettes | −3.56 (−4.57, −2.55) | 1.04 × 10−11 | 0.5949 | 0.0587 | 47.74 | 1.748 | 1.04 × 10−11 |
| AUD | 10.61 (7.88, 13.34) | 8.51 × 10−14 | 0.5911 | 0.0706 | 57.85 | 1.748 | 8.51 × 10−14 |
| Cannabis | 0.47 (0.34, 0.59) | 7.50 × 10−13 | 0.5928 | 0.0652 | 53.25 | 1.748 | 7.49 × 10−13 |
| Analgesics | −0.62 (−0.83, −0.41) | 5.42 × 10−9 | 0.5998 | 0.0432 | 34.84 | 1.748 | 5.43 × 10−9 |
| Cocaine | −0.46 (−6.82, 5.9) | 8.87 × 10−1 | 0.6136 | −0.0013 | 0.0202 | 1.748 | 8.87 × 10−1 |
| Cannabinoids | | | | | | | |
| lm(Rate~Cannabinoids) | | | | | | |
| THC | 0.25 (0.17, 0.34) | 6.75 × 10−9 | 0.5999 | 0.0427 | 34.39 | 1.748 | 6.75 × 10−9 |
| Cannabigerol | 0.37 (0.26, 0.48) | 3.55 × 10−11 | 0.5958 | 0.0557 | 45.19 | 1.748 | 3.55 × 10−11 |
| Cannabichromene | 0.45 (0.32, 0.57) | 1.83 × 10−12 | 0.5935 | 0.0630 | 51.38 | 1.748 | 1.83 × 10−142 |
| Cannabinol | 0.24 (0.16, 0.32) | 1.91 × 10−9 | 0.5989 | 0.0458 | 36.97 | 1.748 | 1.91 × 10−9 |
| Cannabidiol | 0.16 (0.06, 0.25) | 2.25 × 10−3 | 0.6098 | 0.0111 | 9.397 | 1.748 | 2.30 × 10−3 |
| Cannabis x THC Potency | 0.25 (0.17, 0.34) | 6.75 × 10−9 | 0.6098 | 0.0427 | 34.39 | 1.748 | 6.75 × 10−9 |
Table 2.
Mixed Effects Regressions.
Table 2.
Mixed Effects Regressions.
| Parameter Estimates | Model Parameters |
|---|
| Parameter | Estimate (C.I.) | p-Value | S.D. | AIC | BIC | Loglik |
|---|
| Cannabis Alone—Race as Random Effects | | |
| lme(Testicular_Cancer_Rate~Cannabis) | | | | | |
| Cannabis | 0.16 (0.15, 0.18) | 1.70 × 10−75 | 0.1972 | −1816.237 | −1790.591 | 912.1183 |
| Additive Model | | | | | | |
| lme(Testicular_Cancer_Rate~Cigarettes + AUD + Cannabis + Analgesics + Cocaine) |
| AUD | 4.96 (4.59, 5.32) | 3.01 × 10−146 | 0.1764 | −2802.042 | −2750.758 | 1409.021 |
| Cannabis | 0.15 (0.14, 0.17) | 1.14 × 10−68 | | | | |
| Cocaine | −4.23 (−5.09, −3.38) | 3.72 × 10−22 | | | | |
| Analgesics | −0.16 (−0.19, −0.13) | 9.53 × 10−31 | | | | |
| Cigarettes | −1.07 (−1.21, −0.93) | 2.96 × 10−50 | | | | |
| 3-Way Interactive Model | | | | | |
| lme(Testicular_Cancer_Rate~Cigarettes * AUD * Cannabis + Analgesics + Cocaine) |
| Cigarettes: Cannabis | 11.07 (9.32, 12.82) | 1.03 × 10−34 | 0.1695 | −3172.38 | −3095.465 | 1598.19 |
| Cigarettes | 27.64 (22.84, 32.44) | 3.84 × 10−29 | | | | |
| AUD | 64.33 (48.44, 80.22) | 2.64 × 10−15 | | | | |
| Cannabis: AUD | 23.66 (17.76, 29.55) | 4.40 × 10−15 | | | | |
| Cocaine | −2.75 (−3.6, −1.89) | 3.43 × 10−10 | | | | |
| Cigarettes: AUD | −293.19 (−361.6, −224.77) | 5.95 × 10−17 | | | | |
| Cigarettes: Cannabis: AUD | −115.24 (−140.38, −90.09) | 3.82 × 10−19 | | | | |
| Cannabis | −2.14 (−2.54, −1.74) | 1.57 × 10−25 | | | | |
| Analgesics | −0.15 (−0.18, −0.12) | 3.76 × 10−29 | | | | |
| 4-Way Interactive Model | | | | |
| lme(Testicular_Cancer_Rate~Cigarettes * AUD * Cannabis * Analgesics + Cocaine) |
| Cannabis: Analgesics | 2.08 (1.64, 2.51) | 9.32 × 10−21 | 0.1684 | −3212.364 | −3116.23 | 1621.182 |
| Analgesics | 5.44 (4.3, 6.58) | 1.12 × 10−20 | | | | |
| Cigarettes: Cannabis: AUD: Analgesics | 33.83 (25.81, 41.85) | 1.78 × 10−16 | | | | |
| Cigarettes: AUD: Analgesics | 85.86 (63.96, 107.76) | 1.87 × 10−14 | | | | |
| Cannabis | 4.4 (3.09, 5.71) | 4.51 × 10−11 | | | | |
| Cocaine | −1.94 (−2.8, −1.07) | 1.15 × 10−05 | | | | |
| Cigarettes: Cannabis | −20.18 (−26.02, −14.34) | 1.41 × 10−11 | | | | |
| Cannabis: AUD: Analgesics | −6.68 (−8.56, −4.79) | 4.30 × 10−12 | | | | |
| Cigarettes | −55.44 (−71.02, −39.86) | 3.54 × 10−12 | | | | |
| AUD: Analgesics | −18.32 (−23.42, −13.22) | 2.14 × 10−12 | | | | |
| Cigarettes: Analgesics | −26.55 (−31.79, −21.31) | 4.98 × 10−23 | | | | |
| Cigarettes: Cannabis: Analgesics | −9.99 (−11.95, −8.02) | 3.75 × 10−23 | | | | |
| 4-Way Interactive Model with Income | | | | | |
| lme(Testicular_Cancer_Rate~Cigarettes * AUD * Cannabis * Analgesics + Cocaine + Income) |
| Analgesics | 5.46 (4.33, 6.59) | 5.04 × 10−21 | 0.1674 | −3258.286 | −3155.747 | 1645.143 |
| Cannabis: Analgesics | 2.04 (1.61, 2.47) | 3.17 × 10−20 | | | | |
| Cigarettes: Cannabis: AUD: Analgesics | 34.58 (26.61, 42.56) | 2.57 × 10−17 | | | | |
| Cigarettes: AUD: Analgesics | 87.71 (65.94, 109.49) | 3.63 × 10−15 | | | | |
| log(MHY) | 0.16 (0.12, 0.21) | 2.38 × 10−13 | | | | |
| Cannabis | 4.25 (2.95, 5.54) | 1.70 × 10−10 | | | | |
| Cocaine | −2.15 (−3.01, −1.29) | 1.08 × 10−6 | | | | |
| Cigarettes: Cannabis | −20.22 (−26.03, −14.42) | 9.74 × 10−12 | | | | |
| Cigarettes | −56.6 (−72.1, −41.11) | 9.34 × 10−13 | | | | |
| Cannabis: AUD: Analgesics | −6.91 (−8.78, −5.04) | 5.83 × 10−13 | | | | |
| AUD: Analgesics | −18.86 (−23.93, −13.79) | 3.56 × 10−13 | | | | |
| Cigarettes: Cannabis: Analgesics | −10 (−11.95, −8.04) | 1.96 × 10−23 | | | | |
| Cigarettes: Analgesics | −27.04 (−32.25, −21.83) | 4.60 × 10−24 | | | | |
Table 3.
Robust Inverse Probability Weighted Regressions.
Table 3.
Robust Inverse Probability Weighted Regressions.
| Parameter | Estimate (C.I.) | p-Value |
|---|
| Additive Model with State Cannabis | |
| svyglm(TestCaRt~Cigarettes + Cannabis + Race + AUD + Analgesics + Cocaine) |
| AUD | 64.55 (56.89, 72.22) | <2.2 × 10−16 |
| Analgesics | 11.01 (6.24, 15.77) | 5.7 × 10−5 |
| NHWhite | 5.44 (2.36, 8.53) | 0.0013 |
| Hispanic | 4.63 (1.54, 7.71) | 0.0055 |
| Cannabis | −9.38 (−12.76, −5.99) | 3.4 × 10−6 |
| Cocaine | −131.14 (−138.96, −123.33) | <2.2 × 10−16 |
| Additive Model with Ethnic THC Exposure | |
| svyglm(TestCaRt~Cigarettes * EthnicTHCExposure * Race + AUD + Analgesics + Cocaine) |
| Cigarettes | 28.96 (27.87, 30.05) | <2 × 10−16 |
| Hispanic | 2.55 (2.05, 3.04) | 1.8 × 10−12 |
| Asian | 4.89 (1.08, 8.69) | 0.0160 |
| EthnicTHCExposure | 2.9 (0.41, 5.38) | 0.0281 |
| Cocaine | −56.03 (−110.14, −1.93) | 0.0492 |
| Interactive Model with Ethnic_THC_Exposure | |
| svyglm(TestCaRt~Cigarettes * EthnicTHCExposure * Race + AUD + Analgesics + Cocaine + Income) |
| EthnicTHCExposure | 4.72 (2.04, 7.41) | 0.0018 |
| Asian | 3.93 (1.64, 6.22) | 0.0022 |
| Cigarettes | 11.09 (3.77, 18.42) | 0.0060 |
| Cigarettes: NHWhite | 109.87 (24.24, 195.5) | 0.0177 |
| Cigarettes: EthnicTHCExposure: Asian | 17.32 (1.64, 33.01) | 0.0388 |
| NHWhite | −19.53 (−39.11, 0.05) | 0.0603 |
| EthnicTHCExposure: Asian | −4.45 (−8.41, −0.5) | 0.0352 |
| Cocaine | −88.11 (−161.51, −14.7) | 0.0257 |
| Cigarettes: EthnicTHCExposure | −20.11 (−32.44, −7.78) | 0.0034 |
| Cigarettes: Asian | −16.35 (−26.11, −6.6) | 0.0027 |
| Interactive Model with State Cannabis | |
| svyglm(TestCaRt~Cigarettes * Cannabis * Race + AUD + Analgesics + Cocaine + Income) |
| AUD | 13.58 (7.69, 19.47) | 0.0001 |
| Cigarettes: Cannabis: NHWhite | 398.68 (214.58, 582.79) | 0.0002 |
| Cigarettes: NHWhite | 933.24 (493.05, 1373.44) | 0.0003 |
| Cigarettes: Hispanic | 1372.45 (672.78, 2072.11) | 0.0007 |
| Cigarettes: Cannabis: Hispanic | 635.8 (305.36, 966.24) | 0.0008 |
| Analgesics | 19.88 (9.19, 30.57) | 0.0011 |
| Cannabis | 42.63 (18.65, 66.61) | 0.0017 |
| Cigarettes: Cannabis | −199.68 (−315.91, −83.44) | 0.0023 |
| Cigarettes | −578.67 (−907.11, −250.23) | 0.0018 |
| Cocaine | −75.51 (−117.23, −33.78) | 0.0015 |
| Cannabis: Hispanic | −144.06 (−218.53, −69.59) | 0.0008 |
| Hispanic | −308.36 (−466.63, −150.09) | 0.0007 |
| NHWhite | −214 (−315.14, −112.87) | 0.0003 |
| Cannabis: NHWhite | −93.06 (−135.37, −50.75) | 0.0002 |
| Interactive Model with State Cannabinoids | |
| svyglm(TestCaRt~Cigarettes * THC * Cannabigerol * Race + AUD + Analgesics + Cocaine + Income) |
| Cigarettes: NHWhite | 127.24 (92.37, 162.1) | 4.7 × 10−7 |
| Cigarettes: THC: Cannabigerol: NHBlack | 13.87 (6.33, 21.41) | 0.0017 |
| Cigarettes: NHBlack | 53.77 (18.14, 89.41) | 0.0075 |
| Cigarettes: THC: NHBlack | 49.95 (14.93, 84.96) | 0.0108 |
| Cigarettes: Cannabigerol: NHBlack | 14.62 (3.67, 25.57) | 0.0161 |
| Cigarettes: THC: Cannabigerol: NHWhite | 35.97 (7.97, 63.97) | 0.0200 |
| Cigarettes: THC: Hispanic | 214.68 (31.83, 397.53) | 0.0317 |
| Cigarettes: Hispanic | −310.38 (−540.45, −80.31) | 0.0152 |
| Cigarettes: Cannabigerol: Hispanic | −89.37 (−155.46, −23.28) | 0.0150 |
| AUD | −43.07 (−68.76, −17.38) | 0.0035 |
| THC: Cannabigerol: NHWhite | −8.18 (−11.39, −4.96) | 6.2 × 10−5 |
| NHWhite | −16.98 (−23.16, −10.81) | 2.4 × 10−5 |
Table 4.
Geospatiotemporal Regressions.
Table 4.
Geospatiotemporal Regressions.
| Lagged Variables | Parameter | Model |
|---|
| Parameter | Estimate (C.I.) | p-Value | LogLik | S.D. | Model Parameter | Estimate | p-Value |
|---|
| | spreml(Rate~Cannabis) | | | | phi | 1.2910 | 1.4 × 10−5 |
| | Cannabis | 0.19 (0.1, 0.28) | 3.4 × 10−5 | −390.8963 | 0.3939 | psi | −0.1114 | 0.0055 |
| | | | | | | rho | −0.3795 | 0.0055 |
| | | | | | | lambda | 0.4298 | 2.1 × 10−5 |
| | spreml(Rate~Cigarettes + AUD + Cannabis + Analgesics + Cocaine)
| | phi | 1.2910 | 1.4 × 10−5 |
| | Cannabis | 0.19 (0.1, 0.28) | 3.4 × 10−5 | −390.8693 | 0.5316 | psi | −0.1114 | 0.0055 |
| | | | | | | rho | −0.3795 | 0.0055 |
| | | | | | | lambda | 0.4298 | 2.1 × 10−5 |
| | spreml(Rate~Cigarettes * Cannabis * AUD + Analgesics + Cocaine)
| | phi | 1.2650 | 2.6 × 10−5 |
| | Cigarettes: Cannabis | 0.36 (0.19, 0.53) | 4.6 × 10−5 | −391.1668 | 0.5300 | psi | −0.1101 | 0.0062 |
| | | | | | | rho | −0.3696 | 0.0088 |
| | | | | | | lambda | 0.4242 | 5.2 × 10−5 |
| | spreml(Rate~Cigarettes * Cannabis * AUD + Analgesics + Cocaine + Income + 5_Races) |
| | CaucAsian-Am. | 1.59 (1.26, 1.93) | <2.2 × 10−16 | −353.3539 | 0.1584 | phi | 0.1846 | 0.0004 |
| | Hispanic-Am. | 0.1 (0.03, 0.16) | 6.0 × 10−3 | | | psi | −0.0891 | 0.0284 |
| | Asian-Am. | 0.13 (0.07, 0.2) | 8.3 × 10−5 | | | rho | −0.1766 | 0.2595 |
| | African-Am. | −0.23 (−0.27, −0.19) | <2.2 × 10−16 | | | lambda | 0.2183 | 0.0847 |
| | spreml(Rate~Cigarettes * THC * Cannabigerol * AUD + Analgesics + Cocaine + Income + 5_Races) |
| | CaucAsian-Am. | 1.6 (1.16, 2.03) | 8.0 × 10−13 | −348.3428 | 0.3929 | phi | 0.1572 | 0.0009 |
| | Hispanic-Am. | 0.11 (0.04, 0.18) | 0.0028 | | | psi | −0.0955 | 0.0197 |
| | Asian-Am. | 0.11 (0.03, 0.19) | 0.0074 | | | rho | −0.1642 | 0.1982 |
| | Cigarettes: Cannabigerol | 1.39 (0.24, 2.53) | 0.0177 | | | lambda | 0.1945 | 0.0531 |
| | THC: Cannabigerol | 0.07 (0.01, 0.12) | 0.0187 | | | | | |
| | Cigarettes | 4.45 (0.34, 8.56) | 0.0340 | | | | | |
| | Analgesics | −0.18 (−0.36, 0) | 0.0457 | | | | | |
| | African-Am. | −0.22 (−0.26, −0.17) | <2.2 × 10−16 | | | | | |
| | 2 Spatial Lags | | | | | | | |
| | spreml(Rate~Cigarettes * THC * Cannabigerol * AUD + Analgesics + Cocaine + Income + 5_Races) |
| CBG, 2 | Caucasian-Am. | 1.63 (1.18, 2.08) | 1.2 × 10−12 | −351.162 | 0.3970 | phi | 0.1882 | 0.0004 |
| | Asian-Am. | 0.14 (0.06, 0.22) | 0.0007 | | | psi | −0.0976 | 0.0167 |
| | Hispanic-Am. | 0.1 (0.03, 0.17) | 0.0054 | | | rho | −0.1825 | 0.2500 |
| | Cigarettes: THC: Cannabigerol | 0.71 (0.05, 1.37) | 0.0350 | | | lambda | 0.2199 | 0.0845 |
| | Cigarettes: THC | 2.58 (0.17, 4.98) | 0.0356 | | | | | |
| | African-Am. | −0.23 (−0.27, −0.19) | <2.2 × 10−16 | | | | | |
| | 2 Temporal Lags | | | | | | | |
| | spreml(Rate~Cigarettes * THC * Cannabigerol * AUD + Analgesics + Cocaine + Income + 5_Races) |
| CBG, 2 | Caucasian-Am. | 1.8 (1.33, 2.26) | 4.6 × 10−14 | −294.7663 | 0.3863 | phi | 0.1487 | 0.0012 |
| | Asian-Am. | 0.16 (0.08, 0.23) | 5.2 × 10−5 | | | psi | −0.0972 | 0.0289 |
| | Hispanic-Am. | 0.15 (0.07, 0.22) | 7.3 × 10−5 | | | rho | −0.1770 | 0.2840 |
| | Cigarettes: THC | 23.6 (11.92, 35.29) | 7.5 × 10−5 | | | lambda | 0.2025 | 0.1205 |
| | Cigarettes: THC: Cannabigerol | 6.22 (3.07, 9.37) | 0.0001 | | | | | |
| | Cannabigerol | 0.18 (0.04, 0.32) | 0.0146 | | | | | |
| | Analgesics | −0.27 (−0.44, −0.09) | 0.0031 | | | | | |
| | THC: Cannabigerol | −1.26 (−1.94, −0.57) | 0.0003 | | | | | |
| | THC | −4.86 (−7.36, −2.37) | 0.0001 | | | | | |
| | African-Am. | −0.22 (−0.26, −0.18) | <2.2 × 10−16 | | | | | |
| | Full Model with Ethnic THC Exposure
| | | | | |
| | spreml(Rate~Cigarettes * AUD + Analgesics + Cocaine + MHY + NHCaucasian-Am._THC_Exposure * NHAfrican-Am._THC_Exposure * Hispanic-Am._THC_Exposure + Asian-Am._THC_Exposure + AIAN_THC_Exposure) |
| | NHAfrican-Am._THC_Exposure | 0.15 (0.06, 0.25) | 0.0009 | −380.0512 | 0.4856 | phi | 1.0087 | 1.1 × 10−5 |
| | Asian-Am._THC_Exposure | −0.1 (−0.19, −0.02) | 0.0173 | | | psi | −0.1142 | 0.0044 |
| | | | | | | rho | −0.5161 | 3.9 × 10−8 |
| | | | | | | lambda | 0.4720 | 4.4 × 10−13 |
Table 5.
Selected e-Values.
Table 5.
Selected e-Values.
| Parameter | Estimate (C.I.) | R.R. (C.I.) | E-Values |
|---|
| LINEAR MODELS | | | |
|---|
| Testicular_Cancer~Cannabis | | |
| Cannabis | 0.47 (0.34, 0.59) | 2.04 (1.69, 2.47) | 3.50, 2.76 |
| Testicular_Cancer~Time * Cannabis | | |
| Cannabis | 0.47 (0.34, 0.59) | 2.04 (1.68, 2.47) | 3.50, 2.76 |
| Additive Drug Model | | | |
| Cannabis | 0.45 (0.32, 0.57) | 2.14 (1.73, 2.65) | 3.70, 2.85 |
| Testicular_Cancer~Cannabis Quintiles | | |
| Quintile 5 | 0.17 (0.13, 0.21) | 2.13 (1.75, 2.58) | 3.68, 2.91 |
| Testicular_Cancer~Time * Cannabis_Quintiles | |
| Year: Quintile 5 | 0.17 (0.13, 0.21) | 2.23 (1.82, 2.73) | 3.88, 3.04 |
| Testicular_Cancer~Time * Dichotomized_Cannabis_Quintiles |
| Upper Quintiles | 0.17 (0.14, 0.21) | 2.24 (1.91, 2.64) | 3.91, 3.22 |
| Substances | | |
| Cannabis | 0.47 (0.34, 0.59) | 2.04 (122.69, 2.43) | 3.50, 2.76 |
| THC | 0.25 (0.17, 0.34) | 1.47 (1.29, 1.67) | 2.30, 1.91 |
| Cannabigerol | 0.37 (0.26, 0.48) | 1.76 (1.49, 2.08) | 2.92, 2.35 |
| Cannabichromene | 0.45 (0.32, 0.57) | 1.98 (1.64, 2.39) | 3.38, 2.68 |
| Cannabinol | 0.24 (0.16, 0.32) | 1.43 (1.27, 1.61) | 2.23, 1.88 |
| Cannabidiol | 0.16 (0.06, 0.25) | 1.26 (1.08, 1.46) | 1.83, 1.40 |
| Cannabis x THC Potency | 0.25 (0.17, 0.34) | 1.46 (1.28, 1.66) | 2.28, 1.90 |
| MIXED EFFECTS | | | |
| Cannabis Alone—Race as Random Effects | |
| Cannabis | 0.16 (0.15, 0.18) | 3.51 (3.08, 4.00) | 6.48, 5.61 |
| Additive Model | | | |
| Cannabis | 0.15 (0.14, 0.17) | 3.29 (2.89, 3.76) | 6.05, 5.23 |
| 3-Way Interactive Model | | |
| Cigarettes: Cannabis | 11.07 (9.32, 12.82) | 9.67 × 1025 (7.66 × 1021, 1.22 × 1030) | 1.95 × 1026, 1.54 × 1022 |
| Cannabis: AUD | 23.66 (17.76, 29.55) | 3.23 × 1055 (5.17 × 1041, 2.02 × 1069) | 6.45 × 1055, 1.03 × 1042 |
| 4-Way Interactive Model | | |
| Cannabis: Analgesics | 2.08 (1.64, 2.51) | 7.48 × 104 (7.22 × 103, 7.75 × 105) | 1.49 × 105, 1.44 × 104 |
| Cigarettes: Cannabis: AUD: Analgesics | 33.83 (25.81, 41.85) | 2.41 × 1079 (4.00 × 1060, 1.46 × 1098) | 4.84 × 1079, 8.01 × 1060 |
| Cannabis | 4.4 (3.09, 5.71) | 2.13 × 1010 (1.85 × 108, 2.45 × 1013) | 4.26 × 1010, 3.71 × 108 |
| 4-Way Interactive Model with Income | |
| Cannabis: Analgesics | 2.04 (1.61, 2.47) | 6.41 × 104 (6.18 × 103, 6.64 × 105) | 1.28 × 105, 1.24 × 104 |
| Cigarettes: Cannabis: AUD: Analgesics | 34.58 (26.61, 42.56) | 4.31 × 1081 (7.03 × 1062, 2.64 × 10100) | 8.61 × 1081, 1.40 × 1063 |
| Cannabis | 4.25 (2.95, 5.54) | 1.05 × 1010 (9.08 × 106, 1.21 × 1013) | 2.09 × 1010, 1.82 × 107 |
| GEOSPATIAL MODELS | | |
| spreml(Rate~Cannabis) | | | |
| Cannabis | 0.19 (0.1, 0.28) | 1.55 (1.18, 2.05) | 2.48, 1.64 |
| spreml(Rate~Cigarettes + AUD + Cannabis + Analgesics + Cocaine) | | |
| Cannabis | 0.19 (0.1, 0.28) | 1.39 (1.19, 1.62) | 2.12, 1.66 |
| spreml(Rate~Cigarettes * Cannabis * AUD + Analgesics + Cocaine) |
| Cigarettes: Cannabis | 0.36 (0.19, 0.53) | 1.85 (1.38, 2.49) | 3.11, 2.10 |
| spreml(Rate~Cigarettes * THC * Cannabigerol * AUD + Analgesics + Cocaine + Income + 5_Races) |
| Cigarettes: Cannabigerol | 1.39 (0.24, 2.53) | 24.77 (1.76, 349.61) | 49.05, 2.91 |
| THC: Cannabigerol | 0.07 (0.01, 0.12) | 1.16 (1.03, 1.32) | 1.60, 1.19 |
| 2 Spatial Lags | | | |
| Cigarettes: THC: Cannabigerol | 0.71 (0.05, 1.37) | 5.09 (1.12, 23.10) | 9.66, 1.50 |
| Cigarettes: THC | 2.58 (0.17, 4.98) | 368.56 (1.51, 9.02 × 104) | 736.63, 2.38 |
| 2 Temporal Lags | | | |
| Cigarettes: THC | 23.6 (11.92, 35.29) | 1.40 × 1024 (1.64 × 1012, 1.20 × 1036) | 2.81 × 1024, 3.29 × 1012 |
| Cigarettes: THC: Cannabigerol | 6.22 (3.07, 9.37) | 2.31 × 106 (1.39 × 103, 3.82 × 109) | 4.62 × 106, 2.79 × 103 |
| Cannabigerol | 0.18 (0.04, 0.32) | 1.52 (1.09, 2.13) | 2.41, 1.39 |
| spreml(Rate~Cigarettes * AUD + Analgesics + Cocaine + MHY + NHCaucasian-Am._THC_Exposure * NHAfrican-Am._THC_Exposure * Hispanic-Am._THC_Exposure + Asian-Am._THC_Exposure + AIAN_THC_Exposure) |
| NHAfrican-Am._THC_Exposure | 0.15 (0.06, 0.25) | 1.33 (1.13, 1.58) | 2.00, 1.51 |
| LEGAL STATUS | | | |
| Legal | 0.14 (0.06, 0.22) | 1.88, (1.33, 2.66) | 3.16, 1.98 |
| Medical | 0.05 (0.01, 0.09) | 1.25 (1.06, 1.48) | 1.82, 1.31 |
| Legal Status Over Time | | | |
| Legal | 0.11 (0.03, 0.19) | 1.63 (1.12, 2.34) | 2.63, 1.50 |
| Dichotomized Status | | | |
| Liberal | 0.05 (0.02, 0.08) | 1.25 (1.10, 1.43) | 1.81, 1.43 |
| Dichotomized Status Over Time | | |
| Liberal | 0.05 (0.02, 0.08) | 1.25 (1.10, 1.43) | 1.81, 1.43 |
| Quintiles | | | |
| Quintile 5 | 0.20 (0.13, 0.21) | 2.13 (1.75, 2.58) | 3.68, 2.91 |
| Dichotomized Quintiles | | | |
| Upper_2_Quintiles | 0.20 (0.06, 0.35) | 2.24 (1.91, 2.64) | 3.91, 3.22 |
Table 6.
Ordered e-Value Lists.
Table 6.
Ordered e-Value Lists.
| No. | E-Value Estimates | Minimum E-Value |
|---|
| 1 | 8.61 × 1081 | 1.40 × 1063 |
| 2 | 4.84 × 1079 | 8.01 × 1060 |
| 3 | 6.45 × 1055 | 1.03 × 1042 |
| 4 | 1.95 × 1026 | 1.54 × 1022 |
| 5 | 2.81 × 1024 | 3.29 × 1012 |
| 6 | 4.26 × 1010 | 3.71 × 108 |
| 7 | 2.09 × 1010 | 1.82 × 107 |
| 8 | 4.62 × 106 | 14,400.00 |
| 9 | 1.49 × 105 | 12,400.00 |
| 10 | 1.28 × 105 | 2790.00 |
| 11 | 736.63 | 5.61 |
| 12 | 49.05 | 5.23 |
| 13 | 9.66 | 3.22 |
| 14 | 6.48 | 3.22 |
| 15 | 6.05 | 3.04 |
| 16 | 3.91 | 2.91 |
| 17 | 3.91 | 2.91 |
| 18 | 3.88 | 2.91 |
| 19 | 3.70 | 2.85 |
| 20 | 3.68 | 2.76 |
| 21 | 3.68 | 2.76 |
| 22 | 3.50 | 2.76 |
| 23 | 3.50 | 2.68 |
| 24 | 3.50 | 2.38 |
| 25 | 3.38 | 2.35 |
| 26 | 3.16 | 2.10 |
| 27 | 3.11 | 1.98 |
| 28 | 2.92 | 1.91 |
| 29 | 2.63 | 1.9 |
| 30 | 2.48 | 1.88 |
| 31 | 2.41 | 1.66 |
| 32 | 2.30 | 1.64 |
| 33 | 2.28 | 1.51 |
| 34 | 2.23 | 1.50 |
| 35 | 2.12 | 1.50 |
| 36 | 2.00 | 1.43 |
| 37 | 1.83 | 1.43 |
| 38 | 1.82 | 1.40 |
| 39 | 1.81 | 1.39 |
| 40 | 1.81 | 1.31 |
| 41 | 1.60 | 1.19 |
Table 7.
Effect of Cannabis Legalization.
Table 7.
Effect of Cannabis Legalization.
| Linear Models |
|---|
| Parameter Estimates | Model Parameters |
|---|
| Parameter | Estimate (C.I.) | p-Value | S.D. | R-Squared | F | dF | p |
|---|
| Legal Status | | | | | | | |
| (lm(Rate~Status) | | | | | | | |
| Decriminalized | 0.03 (−0.01, 0.07) | 0.0979 | 0.2029 | 0.0196 | 5.979 | 3.746 | 0.0005 |
| Legal | 0.14 (0.06, 0.22) | 0.0004 | | | | | |
| Medical | 0.05 (0.01, 0.09) | 0.0086 | | | | | |
| (lm(Rate~Year * Status) | | | | | | |
| Legal | 0.11 (0.03, 0.19) | 0.0102 | 0.2029 | 1.96 × 10−2 | 5.979 | 3.746 | 0.0005 |
| (lm(Rate~Dichotomized Status) | | | | | | |
| Liberal | 0.05 (0.02, 0.08) | 0.0008 | 0.2035 | 0.0135 | 11.24 | 1.748 | 0.0008 |
| (lm(Rate~Year * Dichotomized Status) | | | | | |
| Liberal | 0.05 (0.02, 0.08) | 0.0008 | 0.2035 | 0.0135 | 11.24 | 1.748 | 0.0008 |
| Linear Models from Imputed Dataset |
| Parameters | Model |
| Parameter | Estimate (C.I.) | p-Value | No. Imputations | SD | lambda | FMI |
| From Imputed Dataset | | | | | | |
| Dichotomized Quintiles | | | | | | |
| lm(TestCaRt~Dichotomized_Quintiles) | | | | | |
| Upper_2_Quintiles | 0.20 (0.06, 0.35) | 0.0058 | 256 | 1.8375 | 0.0309 | 0.0316 |


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}