
Molecular Alterations and Severe Abnormalities in Spermatozoa of Young Men Living in the “Valley of Sacco River” (Latium, Italy): A Preliminary Study

 ,
,  ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Preface
1.2. The Issue
1.3. Pollution and Reproduction
1.4. The Aim of Work
2. Materials and Methods
2.1. Reagents
2.2. Ethical Statements
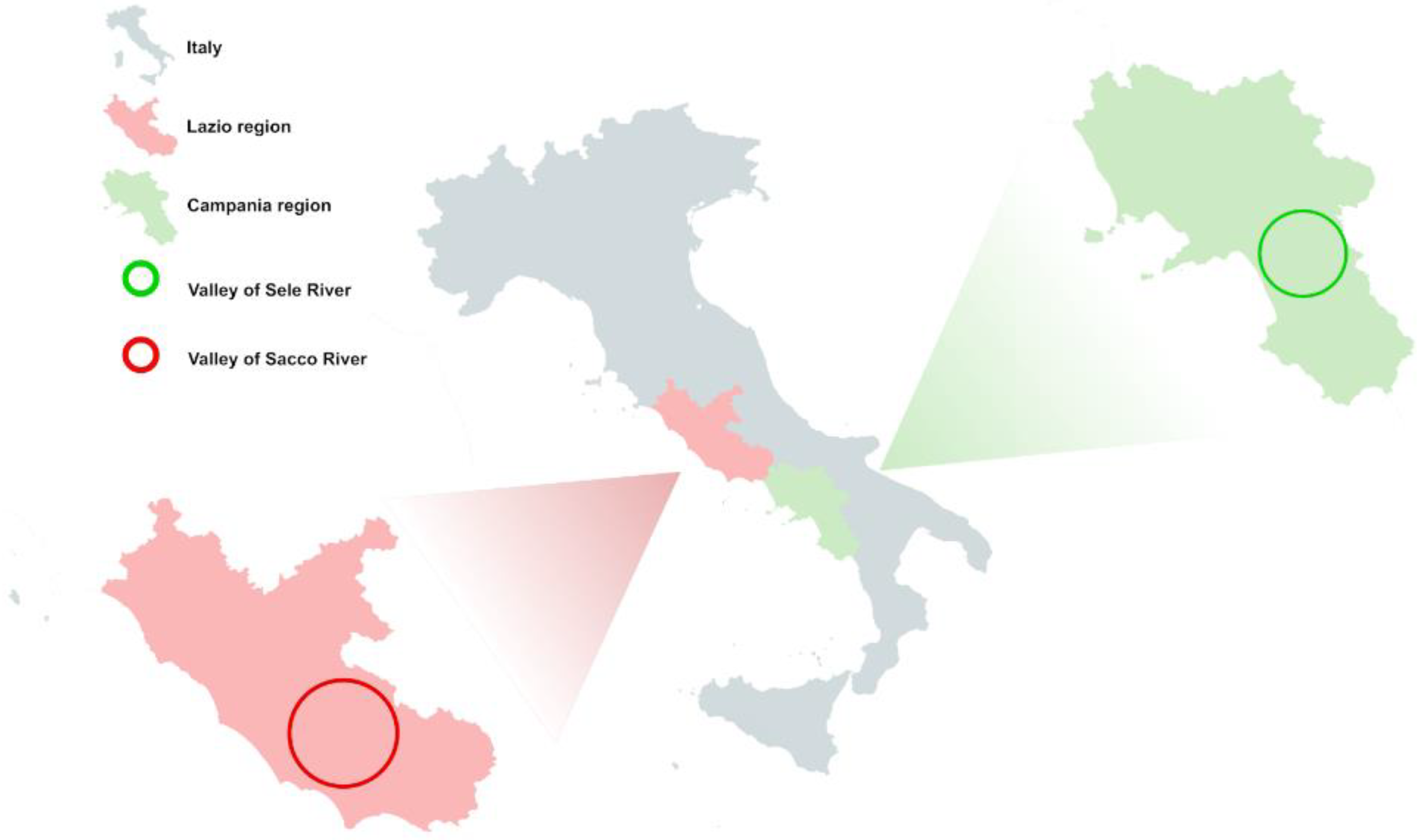
2.3. Recruitment
2.4. Sample Collection
2.5. SNBP Extraction from Spermatozoa
2.6. Electrophoretic Analysis of SNBPs
2.7. Plasmid DNA Preparation
2.8. Analysis of SNBP/DNA Binding
2.9. VOC Analysis
2.10. Spermatozoa Staining
2.11. Statistical Analysis
3. Results
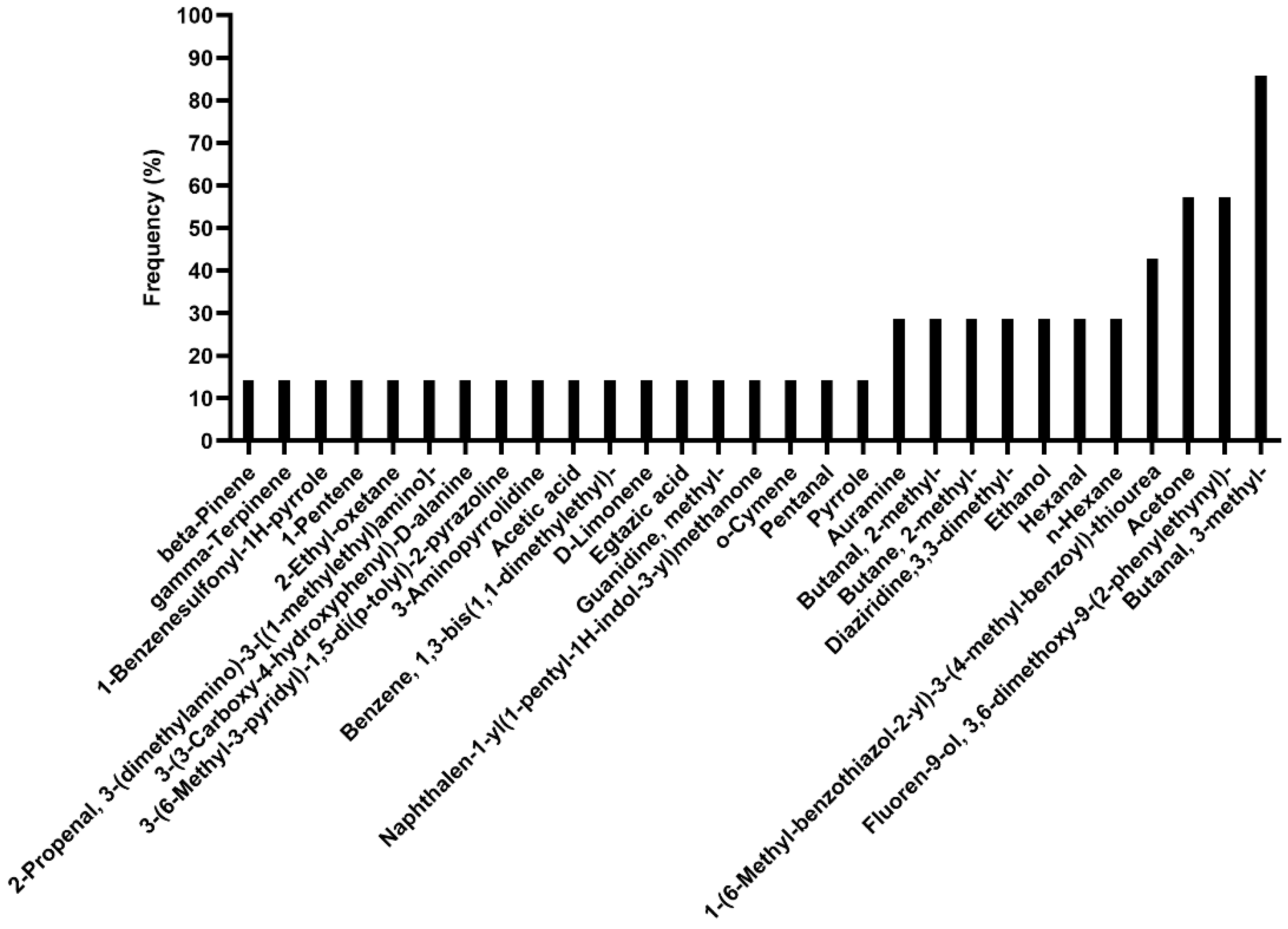
3.1. Qualitative and Quantitative Analysis of VOCs
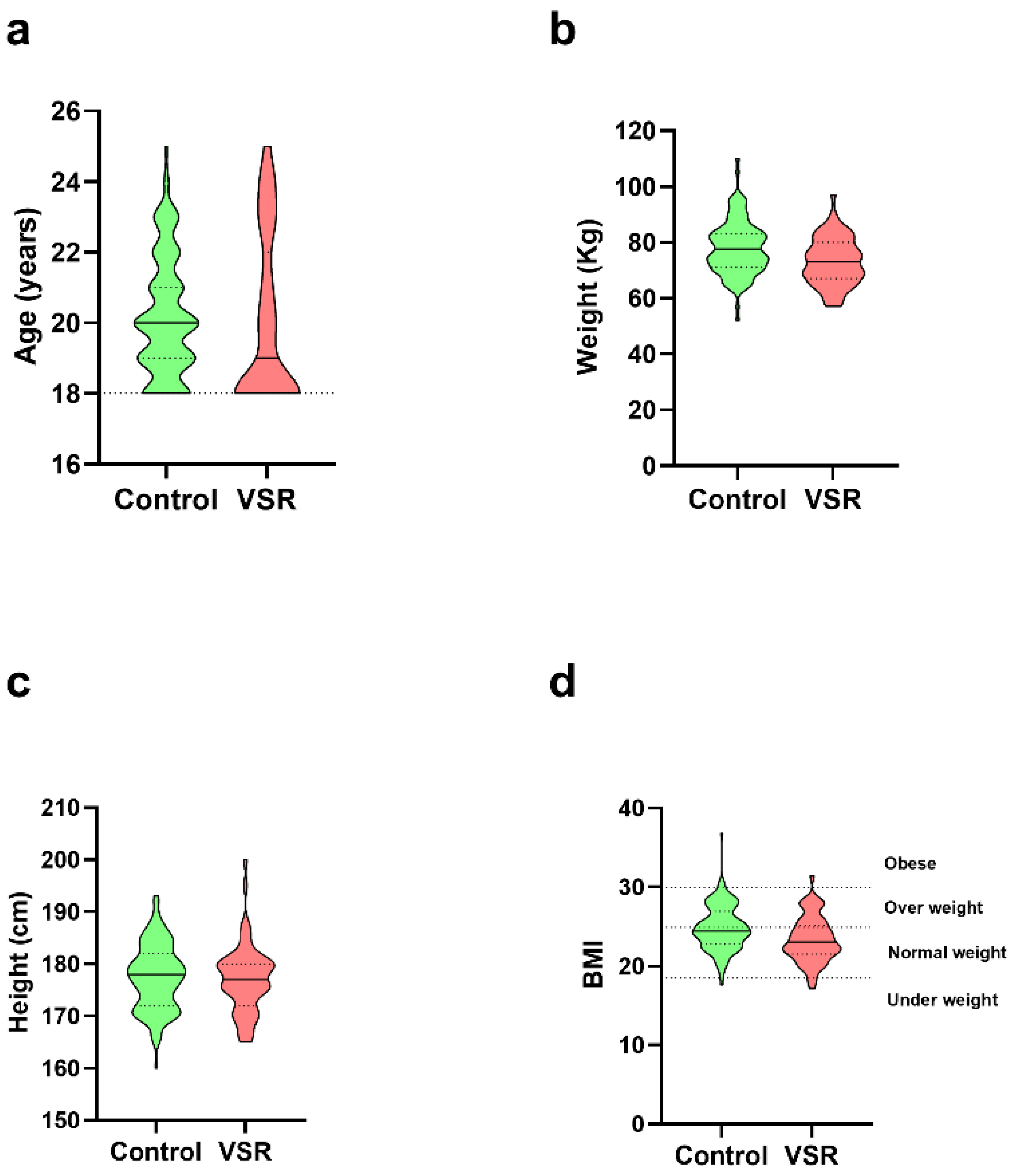
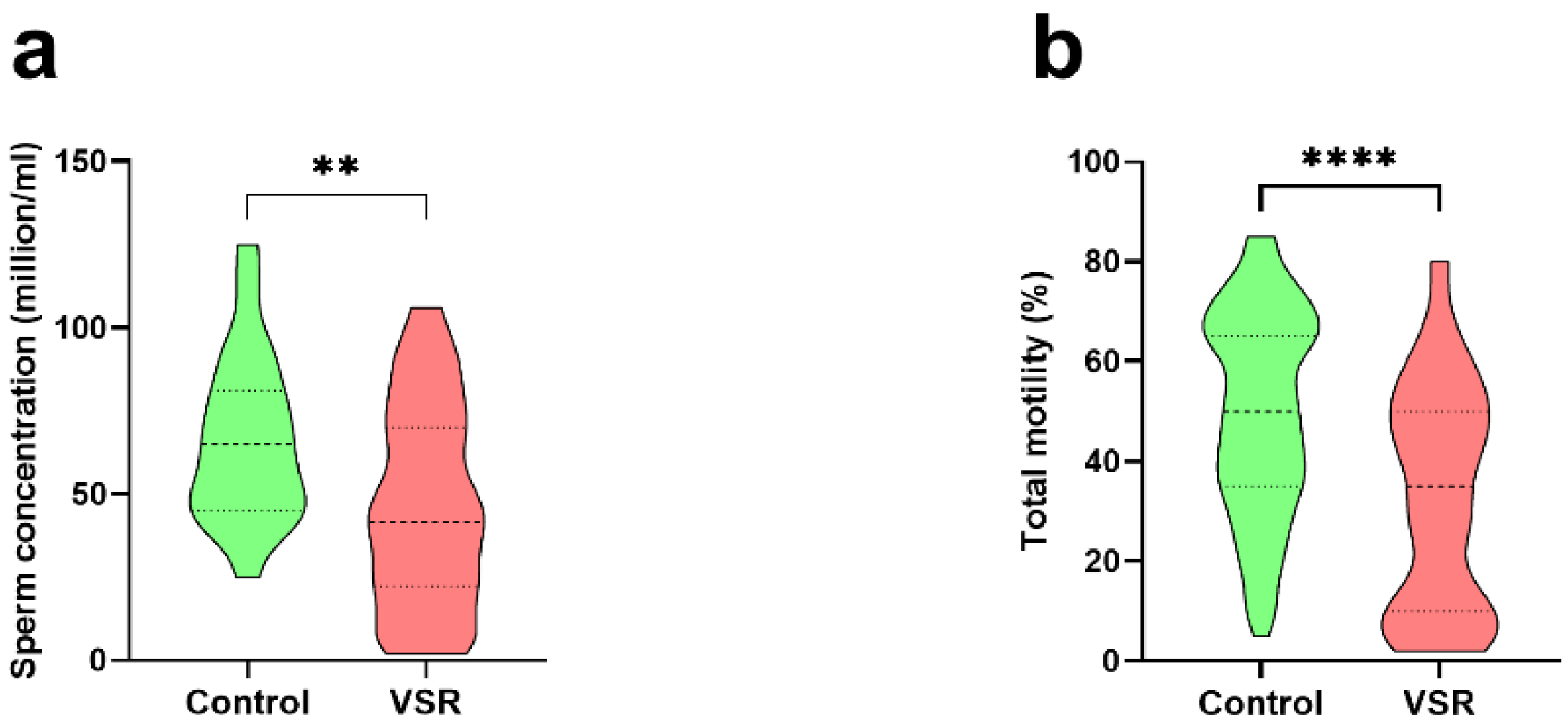
3.2. Anthropometric Data of Individuals and Semen Analyses
3.3. Semen Analysis
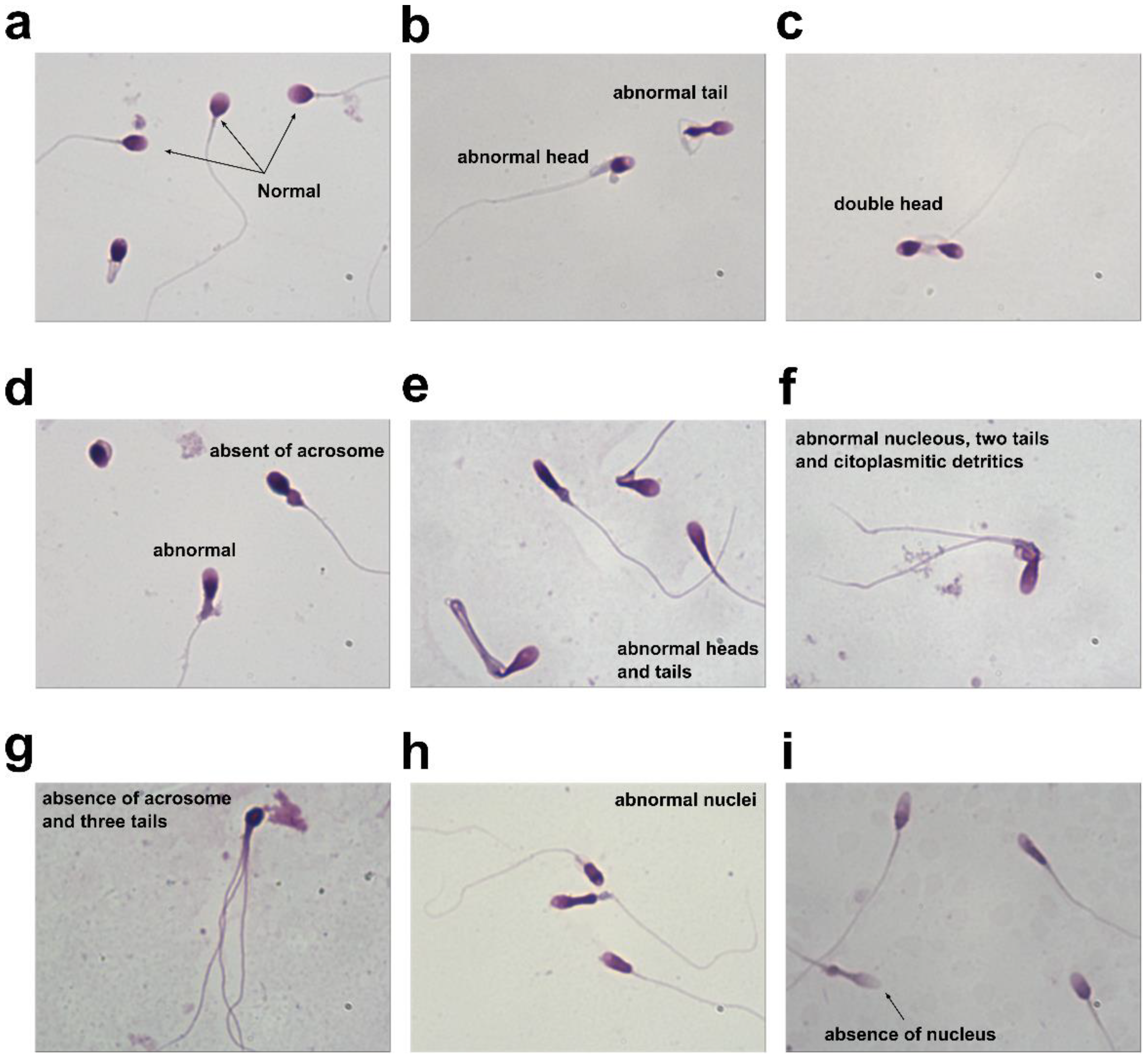
3.4. Evaluation of Sperm Morphology of “Valle del Sacco” Samples
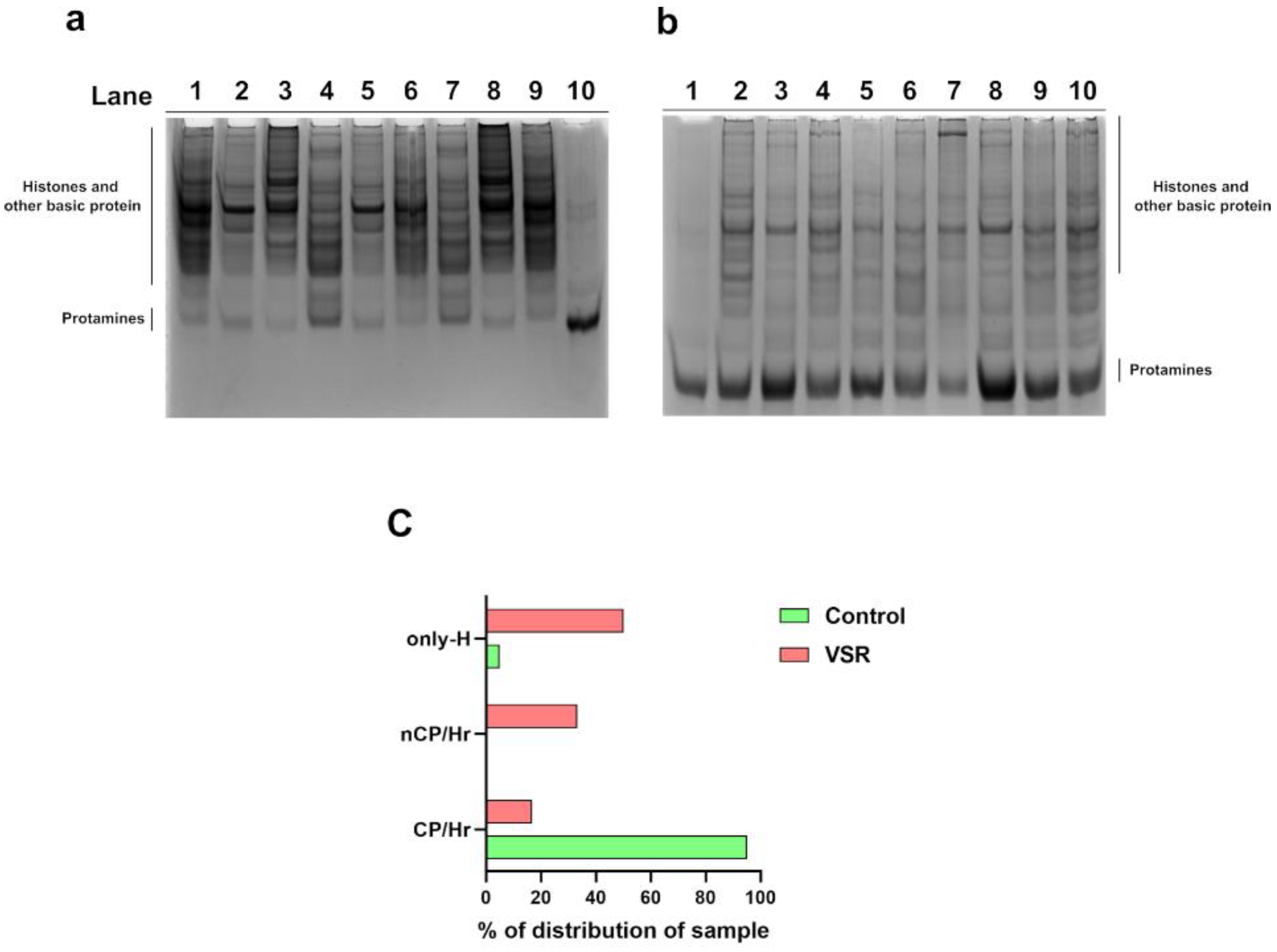
3.5. Analysis of SNBPs
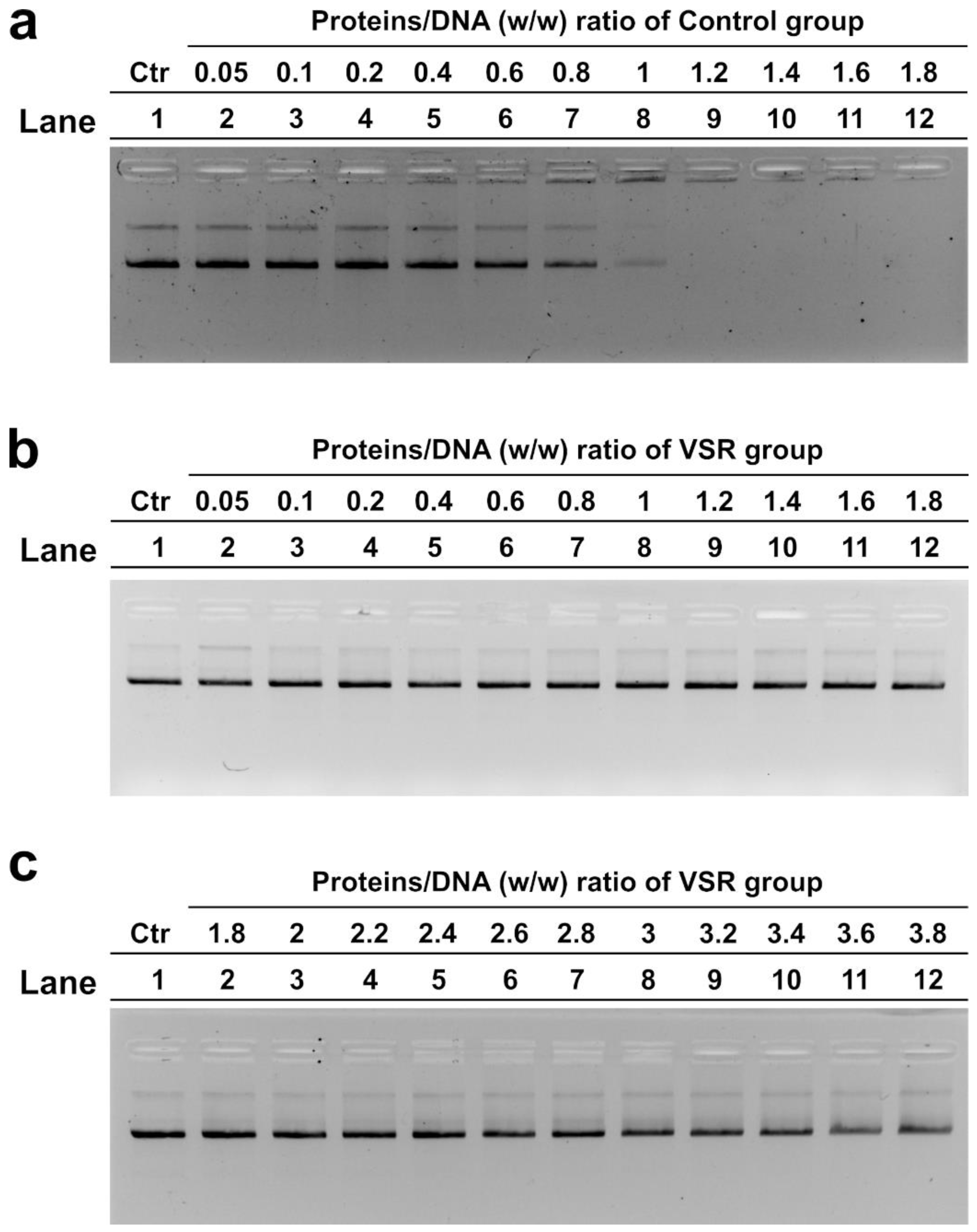
3.6. DNA Binding Ability of SNBPs Analyzed by EMSA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| VSR | Valley Of Sacco River |
| EMSA | Electrophoretic Mobility Shift Assays |
| EDTA | Ethylenediaminetetraacetic Acid |
| VOCs | Volatile Organic Compounds |
| SNBP | Sperm Nuclear Basic Protein |
| CP/Hr | Canonical Protamines/Histones Ratio |
| nCP/Hr | Not Canonical Protamines/Histones Ratio |
| Only-H | Only histones and other basic proteins |
| BMI | Body Mass Index |
| AU-PAGE | Acetic Acid-Urea Gel Electrophoresis |
| PMSF | Phenylmethylsulfonyl Fluoride |
| DTT | Dithiothreitol |
| TCA | Trichloro Acetic Acid |
| TBE | Tris-Borate-EDTA |
References
- Fantini, F.; Porta, D.; Fano, V.; De Felip, E.; Senofonte, O.; Abballe, A.; D’Ilio, S.; Ingelido, A.M.; Mataloni, F.; Narduzzi, S.; et al. Epidemiologic Studies on the Health Status of the Population Living in the Sacco River Valley. Epidemiol. Prev. 2012, 36, 44–52. [Google Scholar] [PubMed]
- Sala, M.; Caminiti, A.; Rombolà, P.; Volpe, A.; Roffi, C.; Caperna, O.; Miceli, M.; Ubaldi, A.; Battisti, A.; Scaramozzino, P. Beta-hexachlorocyclohexane contamination in dairy farms of the Sacco River Valley, Latium, Italy, 2005. A retrospective cohort study. Epidemiol. Prev. 2012, 36, 34–43. [Google Scholar] [PubMed]
- Narduzzi, S.; Fantini, F.; Blasetti, F.; Rantakokko, P.; Kiviranta, H.; Forastiere, F.; Michelozzi, P.; Porta, D. Predictors of Beta-Hexachlorocyclohexane Blood Levels among People Living Close to a Chemical Plant and an Illegal Dumping Site. Environ. Health 2020, 19, 9. [Google Scholar] [CrossRef]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular Effects of Copper on the Reproductive System of Mytilus Galloprovincialis. Mol. Reprod. Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Marra, F.; Moriello, C.; Prisco, M.; Notari, T.; Trifuoggi, M.; Giarra, A.; Bosco, L.; Montano, L.; Piscopo, M. Molecular Alterations in Spermatozoa of a Family Case Living in the Land of Fires. A First Look at Possible Transgenerational Effects of Pollutants. Int. J. Mol. Sci. 2020, 21, 6710. [Google Scholar] [CrossRef]
- Piscopo, M. Seasonal Dependence of Cadmium Molecular Effects on Mytilus Galloprovincialis (Lamarck, 1819) Protamine-like Protein Properties. Mol. Reprod. Dev. 2019, 86, 1418–1429. [Google Scholar] [CrossRef]
- Lettieri, G.; D’Agostino, G.; Mele, E.; Cardito, C.; Esposito, R.; Cimmino, A.; Giarra, A.; Trifuoggi, M.; Raimondo, S.; Notari, T.; et al. Discovery of the Involvement in DNA Oxidative Damage of Human Sperm Nuclear Basic Proteins of Healthy Young Men Living in Polluted Areas. Int J. Mol. Sci 2020, 21, 4198. [Google Scholar] [CrossRef]
- Piscopo, M.; Notariale, R.; Rabbito, D.; Ausió, J.; Olanrewaju, O.S.; Guerriero, G. Mytilus Galloprovincialis (Lamarck, 1819) Spermatozoa: Hsp70 Expression and Protamine-like Protein Property Studies. Environ. Sci. Pollut. Res. Int. 2018, 25, 12957–12966. [Google Scholar] [CrossRef]
- De Guglielmo, V.; Puoti, R.; Notariale, R.; Maresca, V.; Ausió, J.; Troisi, J.; Verrillo, M.; Basile, A.; Febbraio, F.; Piscopo, M. Alterations in the Properties of Sperm Protamine-like II Protein after Exposure of Mytilus Galloprovincialis (Lamarck 1819) to Sub-Toxic Doses of Cadmium. Ecotoxicol. Environ. Saf. 2019, 169, 600–606. [Google Scholar] [CrossRef]
- Piscopo, M.; Trifuoggi, M.; Notariale, R.; Labar, S.; Troisi, J.; Giarra, A.; Rabbito, D.; Puoti, R.; de Benedictis, D.; Brundo, M.V.; et al. Protamine-like Proteins’ Analysis as an Emerging Biotechnique for Cadmium Impact Assessment on Male Mollusk Mytilus Galloprovincialis (Lamarck 1819). Acta Biochim. Pol. 2018, 65, 259–267. [Google Scholar] [CrossRef]
- Lettieri, G.; Notariale, R.; Ambrosino, A.; Di Bonito, A.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus Galloprovincialis to Mercury. Int J. Mol. Sci. 2021, 22, 1618. [Google Scholar] [CrossRef] [PubMed]
- Montano, L.; Pironti, C.; Pinto, G.; Ricciardi, M.; Buono, A.; Brogna, C.; Venier, M.; Piscopo, M.; Amoresano, A.; Motta, O. Polychlorinated Biphenyls (PCBs) in the Environment: Occupational and Exposure Events, Effects on Human Health and Fertility. Toxics 2022, 10, 365. [Google Scholar] [CrossRef]
- Bergamo, P.; Volpe, M.G.; Lorenzetti, S.; Mantovani, A.; Notari, T.; Cocca, E.; Cerullo, S.; Di Stasio, M.; Cerino, P.; Montano, L. Human Semen as an Early, Sensitive Biomarker of Highly Polluted Living Environment in Healthy Men: A Pilot Biomonitoring Study on Trace Elements in Blood and Semen and Their Relationship with Sperm Quality and RedOx Status. Reprod. Toxicol. 2016, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bosco, L.; Notari, T.; Ruvolo, G.; Roccheri, M.C.; Martino, C.; Chiappetta, R.; Carone, D.; Lo Bosco, G.; Carrillo, L.; Raimondo, S.; et al. Sperm DNA Fragmentation: An Early and Reliable Marker of Air Pollution. Environ. Toxicol. Pharmacol. 2018, 58, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Vecoli, C.; Montano, L.; Andreassi, M.G. Environmental Pollutants: Genetic Damage and Epigenetic Changes in Male Germ Cells. Environ. Sci. Pollut. Res. Int. 2016, 23, 23339–23348. [Google Scholar] [CrossRef] [PubMed]
- Vecoli, C.; Montano, L.; Borghini, A.; Notari, T.; Guglielmino, A.; Mercuri, A.; Turchi, S.; Andreassi, M.G. Effects of Highly Polluted Environment on Sperm Telomere Length: A Pilot Study. Int. J. Mol. Sci. 2017, 18, 1703. [Google Scholar] [CrossRef]
- Basile, A.; Loppi, S.; Piscopo, M.; Paoli, L.; Vannini, A.; Monaci, F.; Sorbo, S.; Lentini, M.; Esposito, S. The Biological Response Chain to Pollution: A Case Study from the “Italian Triangle of Death” Assessed with the Liverwort Lunularia Cruciata. Environ. Sci. Pollut. Res. Int. 2017, 24, 26185–26193. [Google Scholar] [CrossRef]
- Piscopo, M.; Trifuoggi, M.; Scarano, C.; Gori, C.; Giarra, A.; Febbraio, F. Relevance of Arginine Residues in Cu(II)-Induced DNA Breakage and Proteinase K Resistance of H1 Histones. Sci. Rep. 2018, 8, 7414. [Google Scholar] [CrossRef]
- Tortora, F.; Notariale, R.; Maresca, V.; Good, K.V.; Sorbo, S.; Basile, A.; Piscopo, M.; Manna, C. Phenol-Rich Feijoa Sellowiana (Pineapple Guava) Extracts Protect Human Red Blood Cells from Mercury-Induced Cellular Toxicity. Antioxidants 2019, 8, 220. [Google Scholar] [CrossRef]
- Maresca, V.; Lettieri, G.; Sorbo, S.; Piscopo, M.; Basile, A. Biological Responses to Cadmium Stress in Liverwort Conocephalum Conicum (Marchantiales). Int. J. Mol. Sci. 2020, 21, 6485. [Google Scholar] [CrossRef]
- Montano, L.; Donato, F.; Bianco, P.M.; Lettieri, G.; Guglielmino, A.; Motta, O.; Bonapace, I.M.; Piscopo, M. Semen Quality as a Potential Susceptibility Indicator to SARS-CoV-2 Insults in Polluted Areas. Environ. Sci. Pollut. Res. Int. 2021, 28, 37031–37040. [Google Scholar] [CrossRef] [PubMed]
- Montano, L.; Donato, F.; Bianco, P.M.; Lettieri, G.; Guglielmino, A.; Motta, O.; Bonapace, I.M.; Piscopo, M. Air Pollution and COVID-19: A Possible Dangerous Synergy for Male Fertility. Int. J. Environ. Res. Public Health 2021, 18, 6846. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, S.; Gentile, M.; Esposito, G.; Gentile, T.; Ferrara, I.; Crescenzo, C.; Palmieri, M.; Cuomo, F.; De Filippo, S.; Lettieri, G.; et al. Could Kallikrein-Related Serine Peptidase 3 Be an Early Biomarker of Environmental Exposure in Young Women? Int. J. Environ. Res. Public Health 2021, 18, 8833. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Forleo, A.; Radogna, A.V.; Siciliano, P.; Notari, T.; Pappalardo, S.; Piscopo, M.; Montano, L.; Capone, S. A Novel Human Biomonitoring Study by Semiconductor Gas Sensors in Exposomics: Investigation of Health Risk in Contaminated Sites. Environ. Pollut. 2022, 304, 119119. [Google Scholar] [CrossRef] [PubMed]
- Marić, T.; Fučić, A.; Aghayanian, A. Environmental and Occupational Exposures Associated with Male Infertility. Arh. Hig. Rada Toksikol. 2021, 72, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Montano, L.; Maugeri, A.; Volpe, M.G.; Micali, S.; Mirone, V.; Mantovani, A.; Navarra, M.; Piscopo, M. Mediterranean Diet as a Shield against Male Infertility and Cancer Risk Induced by Environmental Pollutants: A Focus on Flavonoids. Int. J. Mol. Sci. 2022, 23, 1568. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and Consequences of Oxidative Stress in Spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Nowicka-Bauer, K.; Nixon, B. Molecular Changes Induced by Oxidative Stress That Impair Human Sperm Motility. Antioxidants 2020, 9, 134. [Google Scholar] [CrossRef]
- Rathke, C.; Baarends, W.M.; Awe, S.; Renkawitz-Pohl, R. Chromatin Dynamics during Spermiogenesis. Biochim. Biophys. Acta 2014, 1839, 155–168. [Google Scholar] [CrossRef]
- Bao, J.; Bedford, M.T. Epigenetic Regulation of the Histone-to-Protamine Transition during Spermiogenesis. Reproduction 2016, 151, R55-70. [Google Scholar] [CrossRef] [Green Version]
- Oliva, R. Protamines and Male Infertility. Hum. Reprod. Update 2006, 12, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Forleo, A.; Ferramosca, A.; Notari, T.; Pappalardo, S.; Siciliano, P.; Capone, S.; Montano, L. Blood, Urine and Semen Volatile Organic Compound (VOC) Pattern Analysis for Assessing Health Environmental Impact in Highly Polluted Areas in Italy. Environ. Pollut. 2021, 286, 117410. [Google Scholar] [CrossRef] [PubMed]
- FASE-3-Rapporto-Ambientale-Preliminare. Available online: https://www.comune.capaccio.sa.it/files/2018/12/FASE-3-Rapporto-ambientale-preliminare.pdf (accessed on 30 August 2022).
- Cooper, T.G.; Noonan, E.; von Eckardstein, S.; Auger, J.; Baker, H.W.G.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T.; et al. World Health Organization Reference Values for Human Semen Characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Lettieri, G.; Notariale, R.; Carusone, N.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. New Insights into Alterations in PL Proteins Affecting Their Binding to DNA after Exposure of Mytilus Galloprovincialis to Mercury—A Possible Risk to Sperm Chromatin Structure? Int. J. Mol. Sci. 2021, 22, 5893. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, Q.A.; Caccavale, F.; Avagnano, S.; Murolo, A.; Guerriero, G.; Fucci, L.; Ausió, J.; Piscopo, M. New Insights into Protamine-like Component Organization in Mytilus Galloprovincialis’ Sperm Chromatin. DNA Cell. Biol. 2015, 34, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Fioretti, F.M.; Fucci, L.; Ausió, J.; Piscopo, M. High Efficiency Method to Obtain Supercoiled DNA with a Commercial Plasmid Purification Kit. Acta Biochim. Pol. 2012, 59, 275–278. [Google Scholar] [CrossRef]
- Lettieri, G.; Maione, M.; Ranauda, M.A.; Mele, E.; Piscopo, M. Molecular Effects on Spermatozoa of Mytilus Galloprovincialis Exposed to Hyposaline Conditions. Mol. Reprod. Dev. 2019, 86, 650–660. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Edwards, K.W.; Chan, P.J.; Wei, A.K.; Corselli, J.U.; Jacobson, J.D. Further Evidence Supporting the Diff-Quik Stain for Sperm Chromatin Integrity Testing. J. Reprod. Med. 2015, 60, 6–12. [Google Scholar]
- Soler-Ventura, A.; Castillo, J.; de la Iglesia, A.; Jodar, M.; Barrachina, F.; Ballesca, J.L.; Oliva, R. Mammalian Sperm Protamine Extraction and Analysis: A Step-By-Step Detailed Protocol and Brief Review of Protamine Alterations. Protein Pept. Lett. 2018, 25, 424–433. [Google Scholar] [CrossRef]
- Montano, L.; Longo, V.; Forleo, A.; Pappalardo, S.; Ferramosca, A.; Siciliano, P.; Capone, S. SC299—Human Semen: Excellent Bioaccumulator of Volatile Organic Compounds (VOCs). (EcoFoodFertility Project). Eur. Urol. Suppl. 2019, 18, e3334. [Google Scholar] [CrossRef]
- Piscopo, M.; Conte, M.; Di Paola, F.; Conforti, S.; Rana, G.; De Petrocellis, L.; Fucci, L.; Geraci, G. Relevance of Arginines in the Mode of Binding of H1 Histones to DNA. DNA Cell Biol. 2010, 29, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Fioretti, F.M.; Febbraio, F.; Carbone, A.; Branno, M.; Carratore, V.; Fucci, L.; Ausió, J.; Piscopo, M. A Sperm Nuclear Basic Protein from the Sperm of the Marine Worm Chaetopterus Variopedatus with Sequence Similarity to the Arginine-Rich C-Termini of Chordate Protamine-Likes. DNA Cell Biol. 2012, 31, 1392–1402. [Google Scholar] [CrossRef]
- Sikka, S.C.; Ayaz, A. Standardized Semen Analysis and Quality Control Management for Multicenter Male Reproductive Toxicology Clinical Trials. In Bioenvironmental Issues Affecting Men’s Reproductive and Sexual Health; Elsevier: Amsterdam, The Netherlands, 2018; pp. 371–386. ISBN 978-0-12-801299-4. [Google Scholar]
- De Lacy Costello, B.; Amann, A.; Al-Kateb, H.; Flynn, C.; Filipiak, W.; Khalid, T.; Osborne, D.; Ratcliffe, N.M. A Review of the Volatiles from the Healthy Human Body. J. Breath Res. 2014, 8, 014001. [Google Scholar] [CrossRef] [PubMed]
- Jung, R.; Beermann, D.; Glatt, H.R.; Oesch, F. Mutagenicity of Structurally Related Oxiranes: Derivatives of Benzene and Its Hydrogenated Congeners. Mutat. Res. 1981, 81, 11–19. [Google Scholar] [CrossRef]
- Xing, C.; Marchetti, F.; Li, G.; Weldon, R.H.; Kurtovich, E.; Young, S.; Schmid, T.E.; Zhang, L.; Rappaport, S.; Waidyanatha, S.; et al. Benzene Exposure near the U.S. Permissible Limit Is Associated with Sperm Aneuploidy. Environ. Health Perspect. 2010, 118, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, F.; Eskenazi, B.; Weldon, R.H.; Li, G.; Zhang, L.; Rappaport, S.M.; Schmid, T.E.; Xing, C.; Kurtovich, E.; Wyrobek, A.J. Occupational Exposure to Benzene and Chromosomal Structural Aberrations in the Sperm of Chinese Men. Environ. Health Perspect. 2012, 120, 229–234. [Google Scholar] [CrossRef]
- Yu, C.-H.; Li, Y.; Zhao, X.; Yang, S.-Q.; Li, L.; Cui, N.-X.; Rong, L.; Yi, Z.-C. Benzene Metabolite 1,2,4-Benzenetriol Changes DNA Methylation and Histone Acetylation of Erythroid-Specific Genes in K562 Cells. Arch. Toxicol. 2019, 93, 137–147. [Google Scholar] [CrossRef]
- Williams, K.E.; Carver, T.A.; Miranda, J.J.L.; Kautiainen, A.; Vogel, J.S.; Dingley, K.; Baldwin, M.A.; Turteltaub, K.W.; Burlingame, A.L. Attomole Detection of in Vivo Protein Targets of Benzene in Mice: Evidence for a Highly Reactive Metabolite. Mol. Cell Proteom. 2002, 1, 885–895. [Google Scholar] [CrossRef]
- Kumar, K.; Woo, S.M.; Siu, T.; Cortopassi, W.A.; Duarte, F.; Paton, R.S. Cation-π Interactions in Protein-Ligand Binding: Theory and Data-Mining Reveal Different Roles for Lysine and Arginine. Chem. Sci. 2018, 9, 2655–2665. [Google Scholar] [CrossRef]
- Chow, P.Y.; Holland, M.K.; Suter, D.A.; White, I.G. Evaluation of Ten Potential Organic Spermicides. Int J. Fertil 1980, 25, 281–286. [Google Scholar] [PubMed]
- Zocolo, G.J.; Pilon dos Santos, G.; Vendemiatti, J.; Vacchi, F.I.; de Umbuzeiro, G.A.; Zanoni, M.V.B. Using SPE-LC-ESI-MS/MS Analysis to Assess Disperse Dyes in Environmental Water Samples. J. Chromatogr. Sci. 2015, 53, 1257–1264. [Google Scholar] [CrossRef]
- Vacchi, F.I.; de Vendemiatti, J.A.S.; da Silva, B.F.; Zanoni, M.V.B.; de Aragão Umbuzeiro, G. Quantifying the Contribution of Dyes to the Mutagenicity of Waters under the Influence of Textile Activities. Sci. Total Environ. 2017, 601–602, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, P.; Somanathan, R. Toxicity of Imine-Iminium Dyes and Pigments: Electron Transfer, Radicals, Oxidative Stress and Other Physiological Effects. J. Appl. Toxicol. 2014, 34, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Douteau-Guével, N.; Coleman, A.W.; Morel, J.-P.; Morel-Desrosiers, N. Complexation of the Basic Amino Acids Lysine and Arginine by Three Sulfonatocalix[n]Arenes (n = 4, 6 and 8) in Water: Microcalorimetric Determination of the Gibbs Energies, Enthalpies and Entropies of Complexation. J. Chem. Soc. Perkin Trans. 2 1999, 3, 629–634. [Google Scholar] [CrossRef]
- Sarwar, A.H.M.G.; Nirala, R.K.; Arif, M.; Khillare, B.; Thakur, S.C. Spermicidal Activity of the Hexane Extract of Piper Longum: An in Vitro Study. Nat. Prod. Res. 2015, 29, 1166–1169. [Google Scholar] [CrossRef]
- Nylén, P.; Ebendal, T.; Eriksdotter-Nilsson, M.; Hansson, T.; Henschen, A.; Johnson, A.C.; Kronevi, T.; Kvist, U.; Sjöstrand, N.O.; Höglund, G. Testicular Atrophy and Loss of Nerve Growth Factor-Immunoreactive Germ Cell Line in Rats Exposed to n-Hexane and a Protective Effect of Simultaneous Exposure to Toluene or Xylene. Arch. Toxicol. 1989, 63, 296–307. [Google Scholar] [CrossRef]
- Dcunha, R.; Hussein, R.S.; Ananda, H.; Kumari, S.; Adiga, S.K.; Kannan, N.; Zhao, Y.; Kalthur, G. Current Insights and Latest Updates in Sperm Motility and Associated Applications in Assisted Reproduction. Reprod. Sci. 2022, 29, 7–25. [Google Scholar] [CrossRef]
- Dai, J.-B.; Wang, Z.-X.; Qiao, Z.-D. The Hazardous Effects of Tobacco Smoking on Male Fertility. Asian J. Androl. 2015, 17, 954–960. [Google Scholar] [CrossRef]
- Longo, V.; Forleo, A.; Provenzano, S.P.; Coppola, L.; Zara, V.; Ferramosca, A.; Siciliano, P.; Capone, S. HS-SPME-GC-MS Metabolomics Approach for Sperm Quality Evaluation by Semen Volatile Organic Compounds (VOCs) Analysis. Biomed. Phys. Eng. Express 2018, 5, 015006. [Google Scholar] [CrossRef]
- Raimondi, M.V.; Cascioferro, S.; Schillaci, D.; Petruso, S. Synthesis and Antimicrobial Activity of New Bromine-Rich Pyrrole Derivatives Related to Monodeoxypyoluteorin. Eur. J. Med. Chem. 2006, 41, 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xia, Q.; Fu, P.P.; Lin, G. Pyrrole-Protein Adducts—A Biomarker of Pyrrolizidine Alkaloid-Induced Hepatotoxicity. J. Food Drug Anal. 2018, 26, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Estep, J.E.; Lamé, M.W.; Jones, A.D.; Segall, H.J. N-Acetylcysteine-Conjugated Pyrrole Identified in Rat Urine Following Administration of Two Pyrrolizidine Alkaloids, Monocrotaline and Senecionine. Toxicol. Lett. 1990, 54, 61–69. [Google Scholar] [CrossRef]
- He, X.; Ma, L.; Xia, Q.; Fu, P.P. 7-N-Acetylcysteine-Pyrrole Conjugate—A Potent DNA Reactive Metabolite of Pyrrolizidine Alkaloids. J. Food Drug Anal. 2016, 24, 682–694. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Chan, E.; Duan, W.; Huang, M.; Chen, Y.-Z. Drug Bioactivation, Covalent Binding to Target Proteins and Toxicity Relevance. Drug Metab. Rev. 2005, 37, 41–213. [Google Scholar] [CrossRef] [PubMed]
- Pumford, N.R.; Halmes, N.C. Protein Targets of Xenobiotic Reactive Intermediates. Annu Rev. Pharmacol Toxicol 1997, 37, 91–117. [Google Scholar] [CrossRef]
- Anderson, R.A.; Willis, B.R.; Oswald, C.; Zaneveld, L.J. Ethanol-Induced Male Infertility: Impairment of Spermatozoa. J. Pharmacol. Exp. Ther. 1983, 225, 479–486. [Google Scholar]
- Alvarez, J.G.; Lee, M.A.; Iozzo, R.V.; Lopez, I.; Touchstone, J.C.; Storey, B.T. Ethanol Accelerates Acrosomal Loss in Human Spermatozoa. J. Androl. 1988, 9, 357–366. [Google Scholar] [CrossRef]
- Tangsrisakda, N.; Iamsaard, S. Effect of Ethanol on the Changes in Testicular Protein Expression in Adult Male Rats. Andrologia 2020, 52, e13784. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Hada, M.; Fukuda, Y.; Inoue, E.; Makino, Y.; Katou, Y.; Shirahige, K.; Okada, Y. Re-Evaluating the Localization of Sperm-Retained Histones Revealed the Modification-Dependent Accumulation in Specific Genome Regions. Cell Rep. 2018, 23, 3920–3932. [Google Scholar] [CrossRef]
- Brykczynska, U.; Hisano, M.; Erkek, S.; Ramos, L.; Oakeley, E.J.; Roloff, T.C.; Beisel, C.; Schübeler, D.; Stadler, M.B.; Peters, A.H.F.M. Repressive and Active Histone Methylation Mark Distinct Promoters in Human and Mouse Spermatozoa. Nat. Struct. Mol. Biol. 2010, 17, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.H.; Sauria, M.E.G.; Lyu, X.; Cheema, M.S.; Ausio, J.; Taylor, J.; Corces, V.G. Chromatin States in Mouse Sperm Correlate with Embryonic and Adult Regulatory Landscapes. Cell Rep. 2017, 18, 1366–1382. [Google Scholar] [CrossRef] [PubMed]
- Hammoud, S.S.; Nix, D.A.; Zhang, H.; Purwar, J.; Carrell, D.T.; Cairns, B.R. Distinctive Chromatin in Human Sperm Packages Genes for Embryo Development. Nature 2009, 460, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Martuzzi, M.; Mitis, F.; Bianchi, F.; Minichilli, F.; Comba, P.; Fazzo, L. Cancer Mortality and Congenital Anomalies in a Region of Italy with Intense Environmental Pressure Due to Waste. Occup. Environ. Med. 2009, 66, 725–732. [Google Scholar] [CrossRef]
- Montano, L.; Ceretti, E.; Donato, F.; Bergamo, P.; Zani, C.; Viola, G.C.V.; Notari, T.; Pappalardo, S.; Zani, D.; Ubaldi, S.; et al. Effects of a Lifestyle Change Intervention on Semen Quality in Healthy Young Men Living in Highly Polluted Areas in Italy: The FASt Randomized Controlled Trial. Eur. Urol. Focus 2022, 8, 351–359. [Google Scholar] [CrossRef]
- Choy, J.T.; Eisenberg, M.L. Male Infertility as a Window to Health. Fertil. Steril. 2018, 110, 810–814. [Google Scholar] [CrossRef]
- Glazer, C.H.; Bonde, J.P.; Eisenberg, M.L.; Giwercman, A.; Hærvig, K.K.; Rimborg, S.; Vassard, D.; Pinborg, A.; Schmidt, L.; Bräuner, E.V. Male Infertility and Risk of Nonmalignant Chronic Diseases: A Systematic Review of the Epidemiological Evidence. Semin. Reprod. Med. 2017, 35, 282–290. [Google Scholar] [CrossRef]
- Pisarska, M. Fertility Status and Overall Health. Semin. Reprod. Med. 2017, 35, 203–204. [Google Scholar] [CrossRef]
- Montano, L.; Bergamo, P.; Andreassi, M.G.; Lorenzetti, S. The Role of Human Semen as an Early and Reliable Tool of Environmental Impact Assessment on Human Health. In Spermatozoa—Facts and Perspectives; Meccariello, R., Chianese, R., Eds.; InTech: London, UK, 2018; ISBN 978-1-78923-170-0. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrone, P.; Lettieri, G.; Marinaro, C.; Longo, V.; Capone, S.; Forleo, A.; Pappalardo, S.; Montano, L.; Piscopo, M. Molecular Alterations and Severe Abnormalities in Spermatozoa of Young Men Living in the “Valley of Sacco River” (Latium, Italy): A Preliminary Study. Int. J. Environ. Res. Public Health 2022, 19, 11023. https://doi.org/10.3390/ijerph191711023
Perrone P, Lettieri G, Marinaro C, Longo V, Capone S, Forleo A, Pappalardo S, Montano L, Piscopo M. Molecular Alterations and Severe Abnormalities in Spermatozoa of Young Men Living in the “Valley of Sacco River” (Latium, Italy): A Preliminary Study. International Journal of Environmental Research and Public Health. 2022; 19(17):11023. https://doi.org/10.3390/ijerph191711023
Chicago/Turabian StylePerrone, Pasquale, Gennaro Lettieri, Carmela Marinaro, Valentina Longo, Simonetta Capone, Angiola Forleo, Sebastiana Pappalardo, Luigi Montano, and Marina Piscopo. 2022. "Molecular Alterations and Severe Abnormalities in Spermatozoa of Young Men Living in the “Valley of Sacco River” (Latium, Italy): A Preliminary Study" International Journal of Environmental Research and Public Health 19, no. 17: 11023. https://doi.org/10.3390/ijerph191711023
APA StylePerrone, P., Lettieri, G., Marinaro, C., Longo, V., Capone, S., Forleo, A., Pappalardo, S., Montano, L., & Piscopo, M. (2022). Molecular Alterations and Severe Abnormalities in Spermatozoa of Young Men Living in the “Valley of Sacco River” (Latium, Italy): A Preliminary Study. International Journal of Environmental Research and Public Health, 19(17), 11023. https://doi.org/10.3390/ijerph191711023