
Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review


 ,
, 
Abstract
:1. Introduction
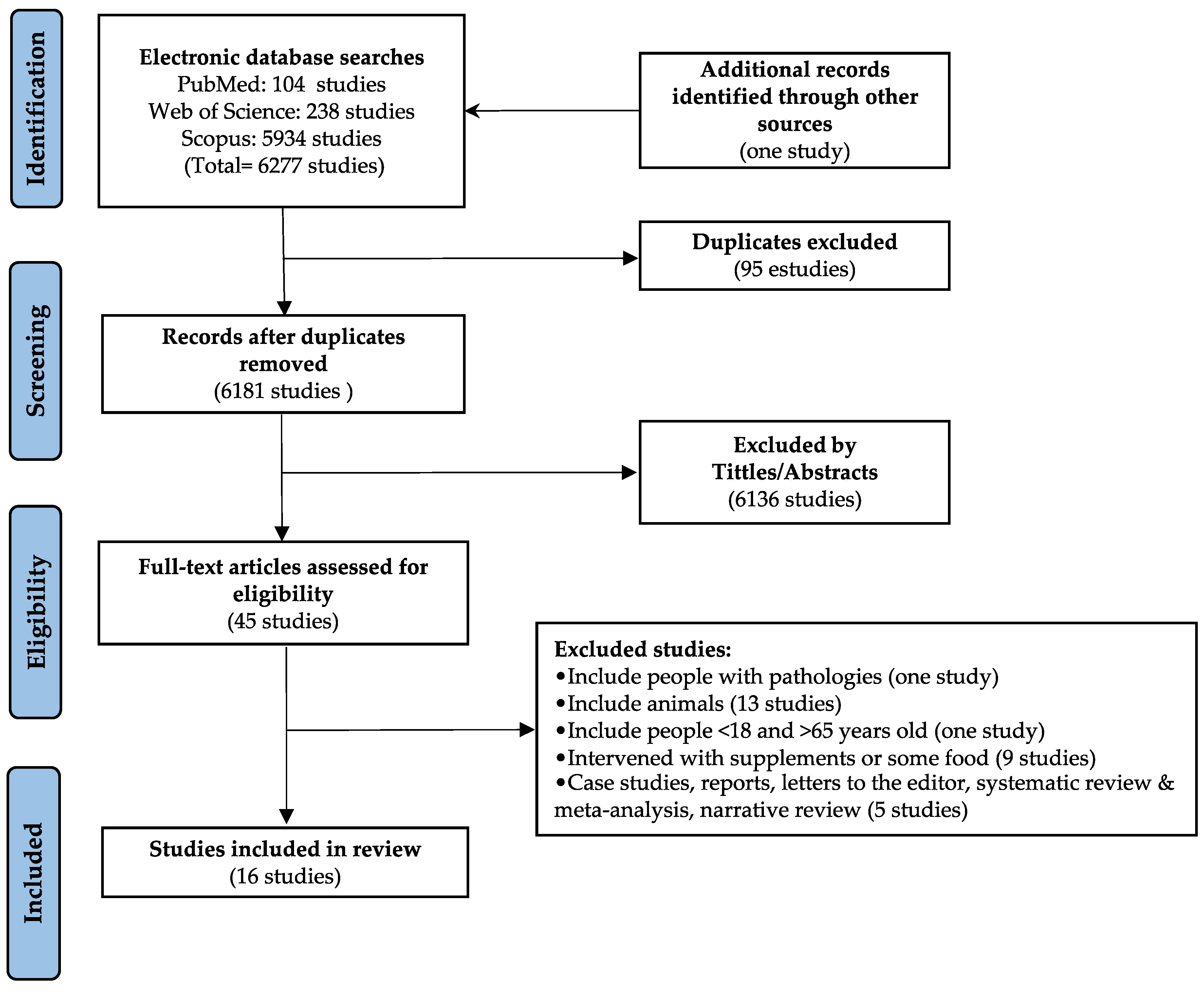
2. Materials and Methods
2.1. Search Strategy
2.2. Selection and Exclusion Criteria
2.3. Data Extraction and Reliability
2.4. Assessment of the Quality and Level of Evidence
3. Results
4. Discussion
4.1. Strategies of Analysis
4.2. Effects of Exercise Duration on Microbiota
4.3. Effects of Exercise Intensity on Microbiota
4.4. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sokol, H. Definition and roles of the gut microbiota. La Rev. Du Prat. 2019, 69, 776–782. [Google Scholar]
- Hold, G.L. The gut microbiota, dietary extremes and exercise. Gut 2014, 63, 1838–1839. [Google Scholar] [CrossRef] [Green Version]
- Mach, N.; Fuster-Botella, D. Endurance exercise and gut microbiota: A review. J. Sport Health Sci. 2016, 6, 179–197. [Google Scholar] [CrossRef]
- Rajilić-Stojanović, M.; De Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef] [PubMed]
- Pugh, J.N.; Impey, S.G.; Doran, D.A.; Fleming, S.C.; Morton, J.P.; Close, G. Acute high-intensity interval running increases markers of gastrointestinal damage and permeability but not gastrointestinal symptoms. Appl. Physiol. Nutr. Metab. 2017, 42, 941–947. [Google Scholar] [CrossRef]
- Monda, V.; Villano, I.; Messina, A.; Valenzano, A.; Esposito, T.; Moscatelli, F.; Viggiano, A.; Cibelli, G.; Chieffi, S.; Monda, M.; et al. Exercise Modifies the Gut Microbiota with Positive Health Effects. Oxid. Med. Cell. Longev. 2017, 2017, 3831972. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.M.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G.M. Community characteristics of the gut microbiomes of competitive cyclists. Microbiome 2017, 5, 98. [Google Scholar] [CrossRef]
- Schmitz, L.; Ferrari, N.; Schwiertz, A.; Rusch, K.; Woestmann, U.; Mahabir, E.; Graf, C. Impact of endurance exercise and probiotic supplementation on the intestinal microbiota: A cross-over pilot study. Pilot Feasibility Stud. 2019, 5, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, R.; Zhang, S.; Peng, X.; Yang, W.; Xu, Y.; Wu, P.; Chen, J.; Cai, Y.; Zhou, J. Characteristics of the gut microbiota in professional martial arts athletes: A comparison between different competition levels. PLoS ONE 2019, 14, e0226240. [Google Scholar] [CrossRef]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut Microbiota: The Neglected Endocrine Organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Lauretani, F.; Tana, C.; Nouvenne, A.; Ridolo, E.; Meschi, T. Exercise and immune system as modulators of intestinal microbiome: Implications for the gut-muscle axis hypothesis. Exerc. Immunol. Rev. 2019, 25, 84–95. [Google Scholar] [PubMed]
- Keohane, D.M.; Woods, T.; O’Connor, P.; Underwood, S.; Cronin, O.; Whiston, R.; O’Sullivan, O.; Cotter, P.; Shanahan, F.; Molloy, M.G. Four men in a boat: Ultra-endurance exercise alters the gut microbiome. J. Sci. Med. Sport 2019, 22, 1059–1064. [Google Scholar] [CrossRef]
- Caspersen, C.J.; Powell, K.E.; Christenson, G.M. Physical activity, exercise, and physical fitness: Definitions and distinctions for health-related research. Public Health Rep. 1985, 100, 126–131. [Google Scholar]
- Ortega, F.B.; Ruiz, J.R.; Castillo, M.J.; Sjöström, M. Physical fitness in childhood and adolescence: A powerful marker of health. Int. J. Obes. 2008, 32, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karhu, E.; Forsgård, R.A.; Alanko, L.; Alfthan, H.; Pussinen, P.; Hämäläinen, E.; Korpela, R. Exercise and gastrointestinal symptoms: Running-induced changes in intestinal permeability and markers of gastrointestinal function in asymptomatic and symptomatic runners. Eur. J. Appl. Physiol. 2017, 117, 2519–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.; Mach, N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morishima, S.; Aoi, W.; Kawamura, A.; Kawase, T.; Takagi, T.; Naito, Y.; Tsukahara, T.; Inoue, R. Intensive, prolonged exercise seemingly causes gut dysbiosis in female endurance runners. J. Clin. Biochem. Nutr. 2021, 68, 253–258. [Google Scholar] [CrossRef]
- Tota, Ł.; Piotrowska, A.; Pałka, T.; Morawska, M.; Mikuľáková, W.; Mucha, D.; Żmuda-Pałka, M.; Pilch, W. Muscle and intestinal damage in triathletes. PLoS ONE 2019, 14, e0210651. [Google Scholar] [CrossRef] [Green Version]
- Grosicki, G.J.; Durk, R.P.; Bagley, J.R. Rapid gut microbiome changes in a world-class ultramarathon runner. Physiol. Rep. 2019, 7, e14313. [Google Scholar] [CrossRef] [Green Version]
- Dalton, A.; Mermier, C.; Zuhl, M. Exercise influence on the microbiome–gut–brain axis. Gut Microbes 2019, 10, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, Z.; Hu, B.; Huang, W.; Yuan, C.; Zou, L. Response of Gut Microbiota to Metabolite Changes Induced by Endurance Exercise. Front. Microbiol. 2018, 9, 765. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef]
- Haynes, A.; Kersbergen, I.; Sutin, A.; Daly, M.; Robinson, E. A systematic review of the relationship between weight status perceptions and weight loss attempts, strategies, behaviours and outcomes. Obes. Rev. 2017, 19, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Molina-Garcia, P.; Migueles, J.H.; Cadenas-Sanchez, C.; Esteban-Cornejo, I.; Mora-Gonzalez, J.; Rodriguez-Ayllon, M.; Plaza-Florido, A.; Vanrenterghem, J.; Ortega, F.B. A systematic review on biomechanical characteristics of walking in children and adolescents with overweight/obesity: Possible implications for the development of musculoskeletal disorders. Obes. Rev. 2019, 20, 1033–1044. [Google Scholar] [CrossRef]
- Van Ekris, E.; Altenburg, T.; Singh, A.S.; Proper, K.I.; Heymans, M.; Chinapaw, M. An evidence-update on the prospective relationship between childhood sedentary behaviour and biomedical health indicators: A systematic review and meta-analysis. Obes. Rev. 2016, 17, 833–849. [Google Scholar] [CrossRef] [PubMed]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [Green Version]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 2018, 67, 625–633. [Google Scholar] [CrossRef]
- Sadowska-Krępa, E.; Rozpara, M.; Rzetecki, A.; Bańkowski, S.; Żebrowska, A.; Pilch, W. Strenuous 12-h run elevates circulating biomarkers of oxidative stress, inflammation and intestinal permeability in middle-aged amateur runners: A preliminary study. PLoS ONE 2021, 16, e0249183. [Google Scholar] [CrossRef] [PubMed]
- Tabone, M.; Bressa, C.; García-Merino, J.A.; Moreno-Pérez, D.; Van, E.C.; Castelli, F.A.; Fenaille, F.; Larrosa, M. The effect of acute moderate-intensity exercise on the serum and fecal metabolomes and the gut microbiota of cross-country endurance athletes. Sci. Rep. 2021, 11, 3558. [Google Scholar] [CrossRef]
- Bycura, D.; Santos, A.; Shiffer, A.; Kyman, S.; Winfree, K.; Sutliffe, J.; Pearson, T.; Sonderegger, D.; Cope, E.; Caporaso, J. Impact of Different Exercise Modalities on the Human Gut Microbiome. Sports 2021, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Craven, J.; Cox, A.J.; Bellinger, P.; Desbrow, B.; Irwin, C.; Buchan, J.; McCartney, D.; Sabapathy, S. The influence of exercise training volume alterations on the gut microbiome in highly-trained middle-distance runners. Eur. J. Sport Sci. 2021; in press. [Google Scholar] [CrossRef]
- Kulecka, M.; Fraczek, B.; Mikula, M.; Zeber-Lubecka, N.; Karczmarski, J.; Paziewska, A.; Ambrozkiewicz, F.; Jagusztyn-Krynicka, K.; Cieszczyk, P.; Ostrowski, J. The composition and richness of the gut microbiota differentiate the top Polish endurance athletes from sedentary controls. Gut Microbes 2020, 11, 1374–1384. [Google Scholar] [CrossRef] [PubMed]
- Moitinho-Silva, L.; Wegener, M.; May, S.; Schrinner, F.; Akhtar, A.; Boysen, T.J.; Schaeffer, E.; Hansen, C.; Schmidt, T.; Rühlemann, M.C. Short-term physical exercise impacts on the human holobiont obtained by a randomised intervention study. BMC Microbiol. 2021, 21, 162. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Gordon, J.I. The core gut microbiome, energy balance and obesity. J. Physiol. 2009, 587, 4153–4158. [Google Scholar] [CrossRef] [PubMed]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef]
- Sato, Y.; Atarashi, K.; Plichta, D.R.; Arai, Y.; Sasajima, S.; Kearney, S.M.; Suda, W.; Takeshita, K.; Sasaki, T.; Okamoto, S.; et al. Novel bile acid biosynthetic pathways are enriched in the microbiome of centenarians. Nature 2021, 599, 458–464. [Google Scholar] [CrossRef]
- Scheiman, J.; Luber, J.M.; Chavkin, T.; Macdonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef]
- Curk, T.; Brackley, C.A.; Farrell, J.D.; Xing, Z.; Joshi, D.; Direito, S.; Bren, U.; Angioletti-Uberti, S.; Dobnikar, J.; Eiser, E.; et al. Computational design of probes to detect bacterial genomes by multivalent binding. Proc. Natl. Acad. Sci. USA 2020, 117, 8719–8726. [Google Scholar] [CrossRef] [Green Version]
- Clarridge, J.E. Impact of 16S rRNA Gene Sequence Analysis for Identification of Bacteria on Clinical Microbiology and Infectious Diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef] [Green Version]
- Peterson, D.; Bonham, K.S.; Rowland, S.; Pattanayak, C.W.; Klepac-Ceraj, V.; RESONANCE Consortium; Deoni, S.C.L.; D’Sa, V.; Bruchhage, M.; Volpe, A.; et al. Comparative Analysis of 16S rRNA Gene and Metagenome Sequencing in Pediatric Gut Microbiomes. Front. Microbiol. 2021, 12, 670336. [Google Scholar] [CrossRef] [PubMed]
- German Network for Bioinformatic Infrastructure. SILVA rRNA Database Project. Available online: https://www.arb-silva.de (accessed on 22 May 2022).
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragón-Vela, J.; Solis-Urra, P.; Ruiz-Ojeda, F.J.; Álvarez-Mercado, A.I.; Olivares-Arancibia, J.; Plaza-Diaz, J. Impact of Exercise on Gut Microbiota in Obesity. Nutrients 2021, 13, 3999. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.; Gibson, G.R.; Walton, G.E.; Magistro, D.; Kinnear, W.; Hunter, K. Systematic Review of the Effects of Exercise and Physical Activity on the Gut Microbiome of Older Adults. Nutrients 2022, 14, 674. [Google Scholar] [CrossRef]
- Mc Gettigan, N.; O’Toole, A.; Boland, K. “Role of exercise in preventing and restoring gut dysbiosis in patients with inflammatory bowel disease”: A letter to the editor. World J. Gastroenterol. 2022, 28, 878–890. [Google Scholar] [CrossRef]
- Cataldi, S.; Bonavolontà, V.; Poli, L.; Clemente, F.M.; De Candia, M.; Carvutto, R.; Silva, A.F.; Badicu, G.; Greco, G.; Fischetti, F. The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review. Biology 2022, 11, 479. [Google Scholar] [CrossRef]
- Ley, R.E. Gut microbiota in 2015: Prevotella in the gut: Choose carefully. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 69–70. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, H.; Wang, X.; Yu, X.; Hu, C.; Zhang, X. The family Coriobacteriaceae is a potential contributor to the beneficial effects of Roux-en-Y gastric bypass on type 2 diabetes. Surg. Obes. Relat. Dis. 2018, 14, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.M.; Petriz, B.; Marques, G.; Kamilla, L.H.; Franco, O.L. Is There an Exercise-Intensity Threshold Capable of Avoiding the Leaky Gut? Front. Nutr. 2021, 8, 627289. [Google Scholar] [CrossRef] [PubMed]
- Yücel, G.; Zhao, Z.; El-Battrawy, I.; Lan, H.; Lang, S.; Li, X.; Buljubasic, F.; Zimmermann, W.-H.; Cyganek, L.; Utikal, J.; et al. Lipopolysaccharides induced inflammatory responses and electrophysiological dysfunctions in human-induced pluripotent stem cell derived cardiomyocytes. Sci. Rep. 2017, 7, 2935. [Google Scholar] [CrossRef] [PubMed]
- Ngkelo, A.; Meja, K.; Yeadon, M.; Adcock, I.; Kirkham, P.A. LPS induced inflammatory responses in human peripheral blood mononuclear cells is mediated through NOX4 and Giα dependent PI-3kinase signalling. J. Inflamm. 2012, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Pedrinolla, A.; Venturelli, M.; Kirmizi, E.; Moschetta, F.; Zardini, M.; Rudi, D.; Bacchi, E.; Schena, F.; Moghetti, P.; Lanza, M. Role of Exercise in Vascular Function and Inflammatory Profile in Age-Related Obesity. J. Immunol. Res. 2018, 2018, 7134235. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.D.; Suckling, C.A.; Peedle, G.Y.; Murphy, J.A.; Dawkins, T.G.; Roberts, M.G. An Exploratory Investigation of Endotoxin Levels in Novice Long Distance Triathletes, and the Effects of a Multi-Strain Probiotic/Prebiotic, Antioxidant Intervention. Nutrients 2016, 8, 733. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.S.; Seguro, C.S.; Naves, M.M.V. Gut microbiota and physical exercise in obesity and diabetes—A systematic review. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 863–877. [Google Scholar] [CrossRef] [PubMed]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vázquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The Treatment-Naive Microbiome in New-Onset Crohn’s Disease. Cell Host Microbe 2014, 15, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Gomes-Neto, J.C.; Boekschoten, M.V.; Schwab, C.; Urich, T.; et al. Lifestyle and Horizontal Gene Transfer-Mediated Evolution of Mucispirillum schaedleri, a Core Member of the Murine Gut Microbiota. mSystems 2017, 2, e00171-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getachew, B.; Aubee, J.I.; Schottenfeld, R.S.; Csoka, A.B.; Thompson, K.M.; Tizabi, Y. Ketamine interactions with gut-microbiota in rats: Relevance to its antidepressant and anti-inflammatory properties. BMC Microbiol. 2018, 18, 222. [Google Scholar] [CrossRef]
- Miquel, S.; Martín, R.; Bridonneau, C.; Robert, V.; Sokol, H.; Bermudez-Humaran, L.G.; Thomas, M.; Langella, P. Ecology and metabolism of the beneficial intestinal commensal bacteriumFaecalibacterium prausnitzii. Gut Microbes 2014, 5, 146–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meehan, C.; Beiko, R.G. A Phylogenomic View of Ecological Specialization in the Lachnospiraceae, a Family of Digestive Tract-Associated Bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Säemann, M.D.; Böhmig, G.A.; Österreicher, C.H.; Burtscher, H.; Parolini, O.; Diakos, C.; Stöckl, J.; Hörl, W.H.; Zlabinger, G.J. Anti-inflammatory effects of sodium butyrate on human monocytes: Potent inhibition of IL-12 and up-regulation of IL-10 production. FASEB J. 2000, 14, 2380–2382. [Google Scholar] [CrossRef] [PubMed]
- Kern, T.; Blond, M.B.; Hansen, T.H.; Rosenkilde, M.; Quist, J.S.; Gram, A.S.; Ekstrøm, C.T.; Hansen, T.; Stallknecht, B. Structured exercise alters the gut microbiota in humans with overweight and obesity—A randomized controlled trial. Int. J. Obes. 2019, 44, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Motiani, K.K.; Collado, M.C.; Eskelinen, J.J.; Virtanen, K.A.; Löyttyniemi, E.; Salminen, S.; Nuutila, P.; Kalliokoski, K.K.; Hannukainen, J.C. Exercise training modulates gut microbiota profile and improves endotoxemia. Med. Sci. Sports Exerc. 2020, 52, 94–104. [Google Scholar] [CrossRef] [Green Version]
- McKee, L.S.; La Rosa, S.L.; Westereng, B.; Eijsink, V.G.; Pope, P.B.; Larsbrink, J. Polysaccharide degradation by the Bacteroidetes: Mechanisms and nomenclature. Environ. Microbiol. Rep. 2021, 13, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davila, A.-M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.-H.; Sanz, Y.; Tomé, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newgard, C.B. Interplay between Lipids and Branched-Chain Amino Acids in Development of Insulin Resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.-S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shou, J.; Chen, P.-J.; Xiao, W.-H. The Effects of BCAAs on Insulin Resistance in Athletes. J. Nutr. Sci. Vitaminol. 2019, 65, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Suryani, D.; Alfaqih, M.S.; Gunadi, J.W.; Sylviana, N.; Goenawan, H.; Megantara, I.; Lesmana, R. Type, Intensity, and Duration of Exercise as Regulator of Gut Microbiome Profile. Curr. Sports Med. Rep. 2022, 21, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Dupuit, M.; Rance, M.; Morel, C.; Bouillon, P.; Boscaro, A.; Martin, V.; Vazeille, E.; Barnich, N.; Chassaing, B.; Boisseau, N. Effect of Concurrent Training on Body Composition and Gut Microbiota in Postmenopausal Women with Overweight or Obesity. Med. Sci. Sports Exerc. 2021, 54, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Munukka, E.; Ahtiainen, J.P.; Puigbó, P.; Jalkanen, S.; Pahkala, K.; Keskitalo, A.; Kujala, U.M.; Pietilä, S.; Hollmén, M.; Elo, L.; et al. Six-Week Endurance Exercise Alters Gut Metagenome That Is not Reflected in Systemic Metabolism in Over-weight Women. Front. Microbiol. 2018, 9, 2323. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Xia, J.; Sun, H. Moderate Treadmill Exercise Modulates Gut Microbiota and Improves Intestinal Barrier in High-Fat-Diet-Induced Obese Mice via the AMPK/CDX2 Signaling Pathway. Diabetes Metab. Syndr. Obes. 2022, 15, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Estaki, M.; Pither, J.; Baumeister, P.; Little, J.P.; Gill, S.K.; Ghosh, S.; Ahmadi-Vand, Z.; Marsden, K.R.; Gibson, D.L. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 2016, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.-C.; Chen, Y.-H.; Chuang, H.-L.; Chiu, C.-C. Investigation of the Effects of Microbiota on Exercise Physiological Adaption, Performance, and Energy Utilization Using a Gnotobiotic Animal Model. Front. Microbiol. 2019, 10, 1906. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Lei, O.K.; Nie, J.; Shi, Q.; Xu, Y.; Kong, Z. Effects of Low-Carbohydrate Diet and Exercise Training on Gut Microbiota. Front. Nutr. 2022, 9, 884550. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Database | Search Strategy | Limits | Filters |
|---|---|---|---|
| Web of Science | (ALL (Physical activity AND gut microbiota OR Physical activity AND intestinal barrier OR Physical activity AND intestinal permeability OR Physical exercise AND gut microbiota OR Physical exercise AND intestinal barrier OR Physical exercise AND intestinal permeability)) | Title Articles English | 238 items filtered |
| PubMed | (Physical activity OR physical exercise) AND (gut microbiota OR intestinal barrier OR intestinal permeability) | Title Articles English Humans | 104 items filtered |
| Scopus | TITLE-ABS-KEY (physical AND activity AND gut AND microbiota) OR (physical AND activity AND intestinal AND barrier) OR (physical AND activity AND intestinal AND permeability) OR (physical AND exercise AND gut AND microbiota) OR (physical AND exercise AND intestinal AND barrier) OR (physical AND exercise AND intestinal AND permeability) AND (LIMIT-TO (OA, “all”)) AND (LIMIT-TO (PUBYEAR, 2022) OR LIMIT-TO (PUBYEAR, 2021) OR LIMIT-TO (PUBYEAR, 2020) OR LIMIT-TO (PUBYEAR, 2019) OR LIMIT-TO (PUBYEAR, 2018) OR LIMIT-TO (PUBYEAR, 2017) OR LIMIT-TO (PUBYEAR, 2016) OR LIMIT-TO (PUBYEAR, 2015)) AND (LIMIT-TO (DOCTYPE, “ar”)) AND (LIMIT-TO (LANGUAGE, “English”)) | Title Articles English | 5934 items filtered |
| Author, Year | Type of Study | AIM | Sample | Study Design | Results |
|---|---|---|---|---|---|
| Pugh et al. (2017) [5] | Quasi-Experimental | Characterize the HIIT effects on small intestinal damage markers | n = 11 (men runners trained) Aged 33.1 ± 10.4; VO2max 60.0 ± 3.2 mL/kg/min | Acute HIIT episode markers of intestinal permeability and damage were evaluated and compared with resting conditions. Minimum running performance of 10 km (39 min) and a minimum of 5 workout sessions per week, using serum sampling, pre-exercise, after each set of exercises, and 2 h post-baseline | HIIT significantly increased the serum lactulose: rhamnose ratio and sucrose concentrations compared with rest. In contrast, urinary lactulose: rhamnose or sucrose concentrations did not vary between study groups. Plasma I-FABP augmented in the recuperation period from HIIT only. After 24 h of HIIT, the researchers found mild symptoms of GI distress |
| Liang et al. (2019) [9] | Cross-sectional | Whether the intestinal microbiota is distinctive between higher-level and lower-level athletes | n = 31 (professional martial arts athletes). 15 women and 16 men; aged 20–24 | Martial arts athletes; Wushu routine, vigorous, fast and dynamic sports. The researchers used 16S rRNA gene sequencing to determine the intestinal changes | Higher-level athletes have augmented metabolic capacity and diversity in the intestinal microbiota compared with lower-level athletes. |
| Petersen et al. (2017) [7] | Cross-sectional | Determine the presence of distinctive organisms in professional and amateur level competitive cyclists | n = 33 (professional and amateur level competitive cyclists); 11 women and 22 men; aged 19–49 | The study used metatranscriptomic (RNA-Seq) sequencing and mWGS | The increase in Prevotella was associated with time reported exercising during an average week. Several professional cyclists have augmented levels of Methanobrevibacter smithii transcripts compared with amateur cyclists. |
| Bressa et al. (2017) [27] | Cross-sectional | Compare intestinal composition among two groups divided by physical exercise levels | n = 40 (premenopausal women). 19 active and 21 sedentary Aged 18–40; BMI 20–25 kg/m2 | The researchers used 16S rRNA gene sequencing to determine the intestinal changes | Performance of physical activity was associated with the presence of health-promoting bacteria (R. hominis, A. muciniphila, Bifidobacterium spp., and F. prausnitzii). Decreased levels of diversity were correlated with sedentary parameters |
| Karhu et al. (2017) [16] | Quasi-experimental | Evaluate the effect of running on GI function markers | n = 17 (active runners); 8 women and 9 men; aged 18–45 | The researchers measured secondary variables, such as zonulin, levels of serum intestinal I-FABP, and bacterial LPS, among others | Both, serum I-FABP and intestinal permeability increased after running, without differences amongst groups. No changes were observed in the bacterial LPS in serum |
| Keohane et al. (2019) [13] | Long-term | Analyze the changes in the intestinal microbiota of four well-trained male athletes to prolonged, high-intensity trans-oceanic rowing | n = 4 (men athletes transatlantic rowing). Aged 25–27; BMI 23–25 kg/m2; VO2Max 46–50 mL/kg/min | Metagenomic whole-genome shotgun sequencing was used | Intense exercise clearly impacts the diversity of the intestinal microbiota, with changes in specific bacteria related to metabolic pathways |
| Bycura et al. (2021) [31] | Quasi-experimental | Impact of CRE or RTE on intestinal microbiota | n = 56 n = 28 CRE group (21 women; Aged 20.7; BMI 24.5 kg/m2 and 7 men; aged 20; BMI 24.0 kg/m2. n = 28 RTE group (17 women; aged 20.4; BMI 23.2 kg/m2 and 11 men; aged 22.6; BMI 24.59 kg/m2 | Intestinal microbiota was measured using 16S rRNA gene sequencing | The observed changes were associated only with the CRE group, resulting in disturbance of the intestinal microbiota |
| Morishima et al. (2020) [18] | Cross-sectional | Effects of highly intensive endurance exercise on the intestinal microbiota and its relationship with the onset of the exercise-induced GI disorders | n = 29 (15 women Japanese endurance runners and 14 nonathletic but healthy women). Aged 20–21; BMI 20.7–21.9 kg/m2 | Fecal microbiota was tested using 16S rRNA metagenomics, and other variables such as moisture content, organic acids, and putrefactive metabolites concentrations were examined | Female elite endurance runners have more abundance of Faecalibacterium, and these changes could be associated with the succinate concentration in this group |
| Tota et al. (2019) [19] | Long-term | Evaluate intestinal and muscle damage in triathletes | n = 15 (triathletes). Aged 6–14; VO2max 58.8 ± 4.5 mL/kg/min | Variables used for the analysis were: cortisol, c-reactive protein, zonulin, and TNF-α | Zonulin and variables of permeability were augmented after the race |
| Zhao et al. (2018) [22] | Quasi-experimental | The gut microbiota immediately responds to the enteric changes in amateur half-marathon runners | n = 20 (4 women and 16 men amateur half-marathon runners). Aged 31.3; BMI 22.6 kg/m2 | Fecal samples were analyzed before and after the marathon using 16 rDNA sequencing analyses | Coriobacteriaceae changes were related to the exercise role in avoiding disease and refining health outcomes. |
| Moitinho-Silva et al. (2021) [34] | Randomized controlled trial | Analyze the changes in the intestinal microbiota on previously physically inactive, healthy adults in comparison to controls that did not perform regular exercise | n = 36 (11 controls; 13 endurance group; 12 strength group). Aged 22–41.3; BMI 19.7–32.5 kg/m2 | Fecal microbiota was tested using 16S rRNA metagenomics | Mucosal damage and inflammation were found after short-term resistance training. No changes were observed in intestinal microbiota |
| Sadowska-Krepa et al. (2021) [29] | Quasi-experimental | Evaluate intestinal damage in middle-aged male subjects | n = 10 (amateur long-distance runners). Aged 21–35 | Variables used for the analysis were: TAS, TOS/TOC, hs-CRP, I-FABP, and zonulin | After the exercise, the levels of intestinal permeability biomarkers as, hs-CRP, I-FABP, zonulin, and inflammation were augmented |
| Kulecka et al. (2020) [33] | Quasi-experimental | Evaluate differences in intestinal microbiota amongst healthy controls and endurance athletes | n = 71 n = 14 marathon runners; n = 11 cross-country skiers; n = 46 healthy control individuals | Fecal microbiota was tested using 16S rRNA metagenomics | Excessive training is associated with changes in Bacteroides and Prevotella and bacterial diversity |
| Tabone et al. (2021) [30] | Quasi-experimental | Determine whether the changes are driven by exercise on the gut microbiota (with 16S rRNA gene) and the serum and fecal metabolome | n = 40 (men endurance cross-country runners). Aged 35.8 ± 8.0; BMI 22.8 ± 2.1 kg/m2; VO2max 58.8 ± 3.24 mL/kg/min | Fecal microbiota was tested using 16S rRNA metagenomics | The changes in gut microbiota could be related to physiological changes in ammonia, uric acid, and lactate |
| Barton et al. (2017) [28] | Cross-Sectional | Evaluate differences in intestinal microbiota amongst exercise and a more sedentary state | n = 86 (40 men professional international rugby union players and 46 men controls) | Fecal microbiota was tested using 16S rRNA metagenomics | Professional international rugby union players had more favorable effects in metabolic pathways than the control group |
| Craven et al. (2021) [32] | Quasi-experimental | Evaluate differences in intestinal microbiota according to training volume | n = 14 (highly trained middle-distance runners). n = 6 women; aged 22.0 ± 3.4; VO2max 59.0 ± 3.2 mL/kg/min n = 8 men; aged 20.7 ± 3.2; VO2max 70.1 ± 4.3 mL/kg/min | Fecal microbiota was tested using 16S rRNA metagenomics | No changes were observed in intestinal microbiota according to training volume in upper taxons. Changes in family, genus, and species were observed, these changes did not return to pre-levels |
| Criteriums According to Kind of Study | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Authors | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | Percentage Reached | Quality Level |
| Pugh et al. [5] | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 66.7 | MQ | ||||
| Liang et al. [9] | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 75.0 | HQ | |||||
| Petersen et al. [7] | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 75.0 | HQ | |||||
| Bressa et al. [27] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 100.0 | HQ | |||||
| Karhu et al. [16] | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 55.6 | MQ | ||||
| Keohane et al. [13] | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 87.5 | HQ | |||||
| Bycura et al. [31] | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 88.9 | HQ | ||||
| Morishima et al. [18] | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 50.0 | MQ | |||||
| Tota et al. [19] | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 50.0 | MQ | |||||
| Zhao et al. [22] | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 55.6 | MQ | ||||
| Moitinho-Silva et al. [34] | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 69.2 | MQ |
| Sadowska-Krepa et al. [29] | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 55.6 | MQ | ||||
| Kulecka et al. [33] | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 88.9 | HQ | ||||
| Tabone et al. [30] | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 66.7 | MQ | ||||
| Barton et al. [28] | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 75.0 | HQ | |||||
| Craven et al. [32] | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 55.6 | MQ | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonomini-Gnutzmann, R.; Plaza-Díaz, J.; Jorquera-Aguilera, C.; Rodríguez-Rodríguez, A.; Rodríguez-Rodríguez, F. Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review. Int. J. Environ. Res. Public Health 2022, 19, 9518. https://doi.org/10.3390/ijerph19159518
Bonomini-Gnutzmann R, Plaza-Díaz J, Jorquera-Aguilera C, Rodríguez-Rodríguez A, Rodríguez-Rodríguez F. Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review. International Journal of Environmental Research and Public Health. 2022; 19(15):9518. https://doi.org/10.3390/ijerph19159518
Chicago/Turabian StyleBonomini-Gnutzmann, Romina, Julio Plaza-Díaz, Carlos Jorquera-Aguilera, Andrés Rodríguez-Rodríguez, and Fernando Rodríguez-Rodríguez. 2022. "Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review" International Journal of Environmental Research and Public Health 19, no. 15: 9518. https://doi.org/10.3390/ijerph19159518
APA StyleBonomini-Gnutzmann, R., Plaza-Díaz, J., Jorquera-Aguilera, C., Rodríguez-Rodríguez, A., & Rodríguez-Rodríguez, F. (2022). Effect of Intensity and Duration of Exercise on Gut Microbiota in Humans: A Systematic Review. International Journal of Environmental Research and Public Health, 19(15), 9518. https://doi.org/10.3390/ijerph19159518