
Soil and Vegetation Development on Coal-Waste Dump in Southern Poland

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Plant Sampling
2.3. Plant Chemical Composition
2.4. Soil Sampling
2.5. Statistical Analyses
2.6. Soil Organic Matter
3. Results
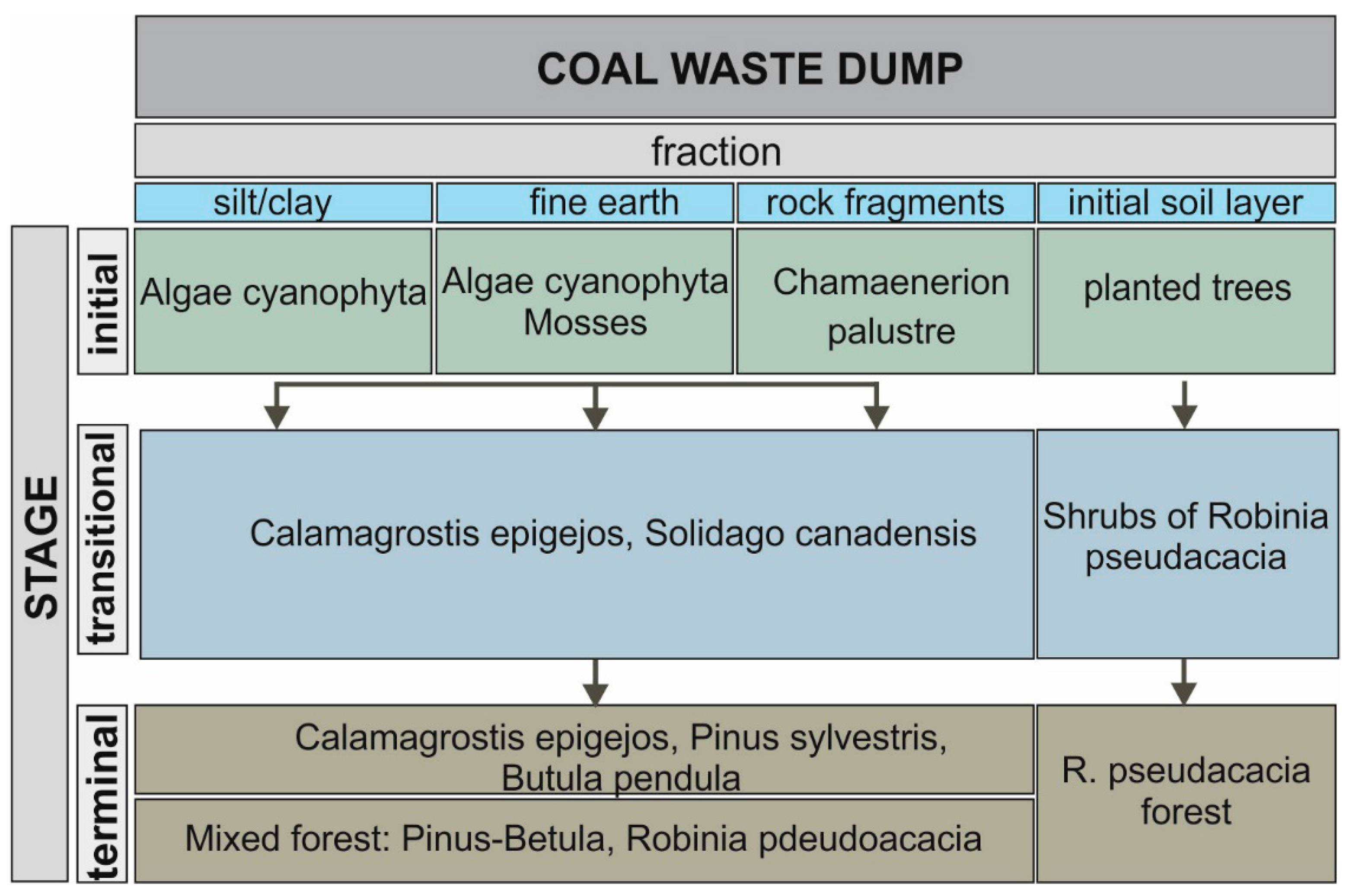
3.1. Vegetation Changes
3.2. Primary Succession
3.3. Secondary Succession
3.4. Flora Diversity
3.5. Soil Morphology
3.6. Soil Physico-Chemical Features
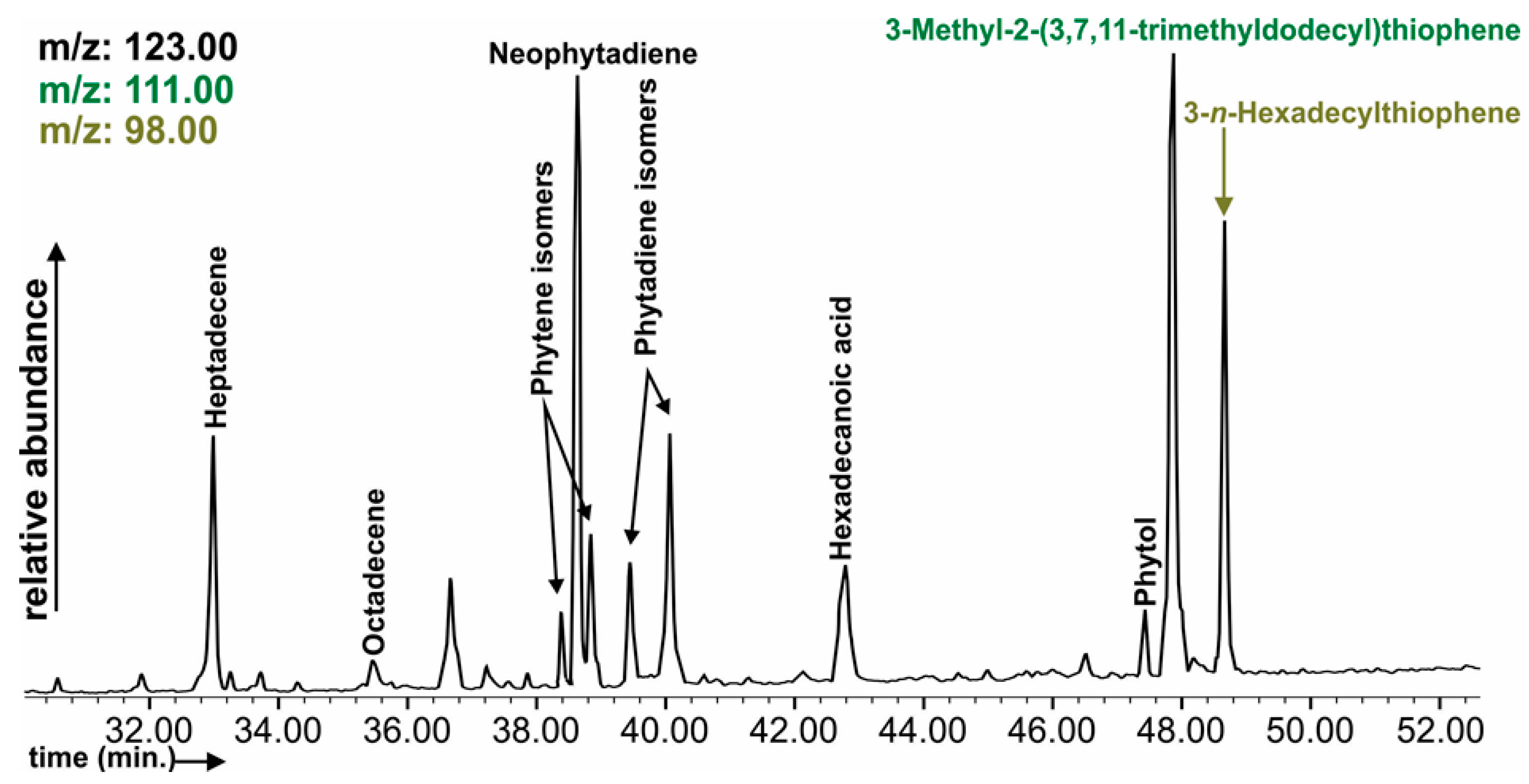
3.7. Soil Organic Matter Composition
3.8. The Major Element Composition of Ch. palustre, C. epigejos, R. pseudacacia, and Soil
3.9. The Heavy Metal Content of Ch. palustre, C. epigejos, R. pseudacacia, and Soil
4. Discussion
4.1. Habitat Conditions and the Course of Succession
4.2. Soil Properties on the Coal-Waste Dump
4.3. Soil Organic Matter Differentiation
4.4. The Contents of Heavy Metals in Soil and Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hilson, G. Pollution prevention and cleaner production in the mining industry: An analysis of current issues. J. Clean. Prod. 2000, 8, 119–126. [Google Scholar] [CrossRef]
- Alekseenko, V.A.; Bech, J.; Alekseenko, A.V.; Shvydkaya, N.V.; Roca, N. Environmental impact of disposal of coal mining wastes on soils and plants in Rostov Oblast, Russia. J. Geochem. Explor. 2018, 184, 261–270. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, J.; Liu, G.B.; Yamanaka, N. Soil properties in natural grassland, Caragana korshinskii planted shrubland, and Robinia pseudacacia planted forest in gullies on the hilly Loess Plateau, China. Catena 2014, 119, 116–124. [Google Scholar] [CrossRef]
- Rahmonov, O.; Cabała, J.; Krzysztofik, R. Vegetation and Environmental Changes on Contaminated Soil Formed on Waste from an Historic Zn-Pb Ore-Washing Plant. Biology 2021, 10, 1242. [Google Scholar] [CrossRef] [PubMed]
- Rahmonov, O.; Skreczko, S.; Rahmonov, M. Changes in Soil Features and Phytomass during Vegetation Succession in Sandy Areas. Land 2021, 10, 265. [Google Scholar] [CrossRef]
- Hendrychová, M.; Kabrna, M. An analysis of 200-year-long changes in a landscape affected by large-scale surface coal mining: History, present and future. Appl. Geogr. 2016, 74, 151–159. [Google Scholar] [CrossRef]
- Kodir, A.; Hartono, D.M.; Haeruman, H.; Mansur, I. Integrated post mining landscape for sustainable land use: A case study in South Sumatera, Indonesia. Sustain. Environ. Res. 2017, 27, 203–213. [Google Scholar] [CrossRef]
- Różkowski, J.; Rahmonov, O.; Zarychta, R.; Zarychta, A. Environmental Transformation and the Current State of Hydrogeo-logical Condition in the Wojkowice Area—Southern Poland. Resources 2021, 10, 54. [Google Scholar] [CrossRef]
- Abramowicz, A.; Rahmonov, O.; Chybiorz, R.; Ciesielczuk, J. Vegetation as an indicator of underground smoldering fire on coal-waste dumps. Fire Saf. J. 2021, 121, 103287. [Google Scholar] [CrossRef]
- Środek, D.; Rahmonov, O. The properties of Black Locust Robinia pseudacacia L. to selectively accumulate chemical ele-ments from soils of ecologically transformed areas. Forests 2021, 13, 7. [Google Scholar] [CrossRef]
- Krzysztofik, R.; Rahmonov, O.; Kantor-Pietraga, I.; Dragan, W. The Perception of Urban Forests in Post-Mining Areas: A Case Study of Sosnowiec-Poland. Int. J. Environ. Res. Public Health 2022, 19, 3852. [Google Scholar] [CrossRef] [PubMed]
- Kirmer, A.; Mahn, E.G. Spontaneous and initiated succession on unvegetated slope sites in the former lignite mining area ‘Goitsche’ (Central Germany). Appl. Veg. Sci. 2001, 4, 19–28. [Google Scholar] [CrossRef]
- Wiegleb, G.; Felinks, B. Primary succession in post mining landscapes of lower Lusatia—Chance or necessity. Ecol. Eng. 2001, 17, 199–217. [Google Scholar] [CrossRef]
- Bradshaw, A.D. The use of natural processes in reclamation—Advantages and difficulties. Lands Urban Plan. 2000, 51, 89–100. [Google Scholar] [CrossRef]
- Rostański, A. Spontaneous Plant Cover on Colliery Spoil Heaps in Upper Silesia (Southern Poland); University of Silesia: Katowice, Poland, 2006. [Google Scholar]
- Woźniak, G. Diversity of Vegetation on Coal-Mine Heaps of the Upper Silesia (Poland); W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2010. [Google Scholar]
- Prach, K.; Hobbs, R. Spontaneous succession versus technical reclamation in the restoration of disturbed sites. Restor. Ecol. 2008, 16, 363–366. [Google Scholar] [CrossRef]
- Prach, K.; Lencová, K.; Řehounková, K.; Dvořáková, H.; Jírová, A.; Konvalinková, P.; Mudrák, O.; Novák, J.; Trnková, R. Spontaneous vegetation succession at different central European mining sites: A comparison across seres. Environ. Sci. Pollut. Res. 2013, 20, 7680–7685. [Google Scholar] [CrossRef]
- Urbancová, L.; Lacková, E.; Kvíčala, M.; Čecháková, L.; Čechák, J.; Stalmachová, B. Plant communities on brownfield sites in Upper Silesia (Czech Republic). Carpath. J. Earth Environ. 2014, 9, 171–177. [Google Scholar]
- Frouz, J.; Voborilová, V.; Janoušová, I.; Kadochová, S.; Matejícek, L. Spontaneous establishment of late successional tree spe-cies English oak (Quercus robur) and European beech (Fagus sylvatica) at reclaimed alder plantation and unreclaimed post mining sites. Ecol. Eng. 2015, 77, 1–8. [Google Scholar] [CrossRef]
- Mudrák, O.; Frouz, J.; Velichová, V. Understory vegetation in reclaimed and unreclaimed post-mining forest stands. Ecol. Eng. 2010, 36, 783–790. [Google Scholar] [CrossRef]
- Rahmonov, O.; Krzysztofik, R.; Środek, D.; Smolarek-Lach, J. Vegetation and environmental changes on non-reclaimed spoil heaps in Southern Poland. Biology 2020, 9, 164. [Google Scholar] [CrossRef]
- Baasch, A.; Kirmer, S.; Tischew, S. Nine years of vegetation development in a postmining site: Effects of spontaneous and assisted site recovery. J. Appl. Ecol. 2011, 49, 251–260. [Google Scholar] [CrossRef]
- Ivanov, M.; Faimon, J.; Jarmara, P.; Pešák, L. Evolution of minesoils at a coal waste pile: A case study from Rosice-Oslavany (Czech Republic). Geologia 2009, 54, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Alday, J.G.; Marrs, R.H.; Martínez-Ruiz, C. Soil and vegetation development during early succession on restored coal wastes: A 6 year permanent plot study. Plant Soil 2012, 353, 305–320. [Google Scholar] [CrossRef]
- Kumar, S.; Maiti, S.K.; Chaudhuri, S. Soil development in 2–21 years old coalmine reclaimed spoil with trees: A case study from Sonepur-Bazari opencast project, Raniganj Coalfield, India. Ecol. Eng. 2015, 84, 311–324. [Google Scholar] [CrossRef]
- Marynowski, L.; Rahmonov, O.; Smolarek-Lach, J.; Rybicki, M.; Simoneit, B.R. Origin and significance of saccharides during initial pedogenesis in a temperate climate region. Geoderma 2020, 361, 114064. [Google Scholar] [CrossRef]
- Kruszewski, Ł.; Kisiel, M.; Cegiełka, M. Soil development in a coal-burning environment: The Upper Silesian waste heaps of Poland. Geol. Q. 2021, 65, 24. [Google Scholar] [CrossRef]
- Rahmonov, O.; Snytko, V.A.; Szczypek, T.; Parusel, T. Vegetation development on post-industrial territories of the Silesian Upland (Southern Poland). Geogr. Nat. Res. 2013, 34, 96–103. [Google Scholar] [CrossRef]
- Kompała-Bąba, A.; Bierza, W.; Błońska, A.; Sierka, E.; Magurno, F.; Chmura, D.; Besenyei, L.; Radosz, Ł.; Woźniak, G. Vegeta-tion diversity on coal mine spoil heaps—How important is the texture of the soil substrate? Biologia 2019, 74, 419–436. [Google Scholar] [CrossRef] [Green Version]
- Prach, K.; Bartha, S.; Joyce, C.B.; Pyšek, P.; van Diggelen, R.; Wiegleb, G. The role of spontaneous vegetation succession in ecosystem restoration: A perspective. Appl. Veg. Sci. 2001, 4, 111–114. [Google Scholar] [CrossRef]
- Rahmonov, O.; Kowalski, W.J.; Bednarek, R. Characterization of the Soil Organic Matter and Plant Tissues in an Initial Stage of the Plant Succession and Soil Development by Means of Curie-point Pyrolysis Coupled with GC-MS. Eurasian Soil Sci. 2010, 43, 1557–1568. [Google Scholar] [CrossRef]
- Rahmonov, O.; Cabała, J.; Bednarek, R.; Rożek, D.; Florkiewicz, A. Role of soil algae on the initial stages of soil formation in sandy polluted areas. Ecol. Chem. Eng. 2015, 22, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Howard, J. Anthropogenic Soils; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Howard, J.L. Urban anthropogenic soils—A review. Adv. Agron. 2021, 165, 1–57. [Google Scholar] [CrossRef]
- Raport Roczny. 2018. Available online: https://www.jsw.pl/raportroczny-2018/strategia/ochrona-srodowiska (accessed on 20 January 2022).
- Matuszkiewicz, W.; Faliński, J.B.; Kostrowicki, A.S.; Matuszkiewicz, J.M.; Olaczek, R.; Wojterski, T. Potencjalna Roślinność Naturalna Polski. In Mapa Przeglądowa 1:300 000; Arkusze 1–12; IGiPZ PAN: Warszwa, Poland, 1995. [Google Scholar]
- Bednarek, R.; Dziadowiec, H.; Pokojska, U.; Prusinkiewicz, Z. Badania Ekologiczno–Gleboznawcze; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2004. [Google Scholar]
- Philp, R.P. Fossil Fuel Biomarkers: Application and Spectra; Elsevier: Amsterdam, The Netherlands, 1985. [Google Scholar]
- McLafferty, F.W.; Stauffer, D.B. The Wiley Registry of Mass Spectral Data; John Wiley and Sons: New York, NY, USA, 1989. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015 International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106.; FAO: Rome, Italy, 2015. [Google Scholar]
- Nádudvari, Á.; Marynowski, L.; Fabiańska, M.J. Application of organic environmental markers in the assessment of recent and fossil organic matter input in coal wastes and river sediments: A case study from the Upper Silesia Coal Basin (Poland). Int. Coal Geol. 2018, 196, 302–316. [Google Scholar] [CrossRef]
- Nádudvari, Á.; Fabiańska, M.J. Coal-related sources of organic contamination in sediments and water samples of Bierawka River (Poland). Int. J. Coal Geol. 2015, 152, 94–109. [Google Scholar] [CrossRef]
- Ukiya, M.; Akihisa, T.; Tokuda, H.; Suzuki, H.; Mukainaka, T.; Ichiishi, E.; Yasukawa, K.; Kasahara, Y.; Nishino, H. Constitu-ents of Compositae plants III. Anti-tumor promoting effects and cytotoxic activity against human cancer cell lines of triter-pene diols and triols from edible chrysanthemum flowers. Cancer Lett. 2002, 177, 7–12. [Google Scholar] [CrossRef]
- Rüdiger, A.L.; Veiga-Junior, V.F. Chemodiversity of ursane- and oleanane-type triterpenes in amazonian burseraceae oleores-ins. Chem. Biodivers. 2013, 10, 1142–1153. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Zhao, H.L. Changes in soil particles fraction and their effects on stability of soil-vegetation system in restoration processes of degraded sandy grassland. Ecol. Environ. Sci. 2009, 18, 1395–1401. [Google Scholar]
- Abramowicz, A.; Rahmonov, O.; Chybiorz, R. Environmental Management and Landscape Transformation on Self-Heating Coal-Waste Dumps in the Upper Silesian Coal Basin. Land 2020, 10, 23. [Google Scholar] [CrossRef]
- de Kroon, H.; Hutchings, M. Morphological plasticity in clonal plants: The foraging concept reconsidered. J. Ecol. 1995, 83, 143–152. [Google Scholar] [CrossRef]
- Bróż, E.; Podgórska, M. Expansion of Chamaenerion palustre (Onagraceae) in the Wyżyna Małopolska Upland. Fragm. Florist. Et Geobot. Pol. 2008, 15, 21–42. [Google Scholar]
- Stafanowicz, A.M.; Kapusta, P.; Błonska, A.; Kompała-Bomba, A.; Woźniak, G. Effects of Calamagrostis epigejos, Chamaene-rion palustre and Tussilago farfara on nutrient availability and microbial activity in the surface layer of spoil heaps after hard coal mining. Ecol. Eng. 2015, 83, 328–337. [Google Scholar] [CrossRef]
- Foltyn, S.; Bogda, A.; Szopka, K.; Karczewska, A. Properties of anthropogenic soils on a mine spoil “Kościelniok” in Pawłowice (hard coal mine Pniówek). Soil Sci. Annual. 2011, 62, 79–85. [Google Scholar]
- Weber, B.; Büdel, B.; Belnap, J. (Eds.) Biological Soil Crusts: An Organizing Principle in Drylands, 1st ed.; Springer: Cham, Switzerland, 2016; 549p. [Google Scholar]
- Rebele, F.; Lehmann, C. Biological Flora of Central Europe: Calamagrostis epigejos (L.) Roth. Flora 2001, 196, 325–344. [Google Scholar] [CrossRef]
- Rahmonov, O. The chemical composition of plant litter of black locust (Robinia pseudacacia L.) and its ecological role in sandy ecosystems. Acta Ecol. Sin. 2009, 29, 237–243. [Google Scholar] [CrossRef]
- Piekarska-Stachowiak, A.; Szary, M.; Ziemer, B.; Besenyei, L.; Woźniak, G. An application of the plant functional group con-cept to restoration practice on coal mine spoil heaps. Ecol. Res. 2014, 29, 843–853. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, C. Clonal diversity of populations of Calamagrostis epigejos in relation to environmental stress and habitat het-erogeneity. Ecography 1997, 20, 483–490. [Google Scholar] [CrossRef]
- Lehmann, C.; Rebele, F. Successful management of Calamagrostis epigejos (L.) Roth on a sandy landfill site. J. Appl. Bot. 2002, 76, 77–81. [Google Scholar]
- Kooijman, A.M.; Dopheide, J.C.R.; Sevink, J.; Takken, I.; Verstraten, J.M. Nutrient limitations and their implications on the effects of atmospheric deposition in coastal dunes; lime-poor and lime-rich sites in the Netherlands. J. Ecol. 1998, 86, 511–526. [Google Scholar] [CrossRef]
- Prach, K. Spontaneous succession in Central-European man-made habitats: What information can be used in restoration prac-tice? Appl. Veg. Sci. 2003, 6, 125–129. [Google Scholar] [CrossRef]
- Patrzałek, A. The importance of grass species and cultivars in the development of turfing process on reclaimed land. Biul. Inst. Hod. I Aklim. Roślin 2003, 225, 359–363. [Google Scholar]
- Raport Zintegrowany. 2019. Available online: https://www.jsw.pl/raportroczny-2019/zrownowazony-rozwoj-w-jsw/ochrona-srodowiska#gospodarka_odpadami-tab (accessed on 20 January 2022).
- Klatka, S.; Malec, M.; Kruk, E.; Ryczek, M. Evaluation of possibility of natural utilisation of coal mine waste used for surface levelling. Acta Agrophysica 2017, 24, 253–262. [Google Scholar]
- Jiang, X.; Lu, W.X.; Zhao, H.Q.; Yang, C.; Yang, Z.P. Potential ecological risk assessment and prediction of soil heavy-metal pollution around coal gangue dump. Nat. Hazards Earth Syst. Sci. 2014, 14, 1599–1610. [Google Scholar] [CrossRef] [Green Version]
- Arefieva, O.; Nazarkina, A.V.; Gruschakova, N.V.; Skurichina, J.E.; Kolycheva, B. Impact of mine waters on chemical composi-tion of soil in the Partizansk Coal Basin, Russia. Int. Soil Water Conserv. Res. 2019, 7, 57–63. [Google Scholar] [CrossRef]
- Ketris, M.P.; Yudovich, Y.E. Estimations of clarkes for carbonaceous biolithes: World averages for trace element con tents in black shales and coals. Int. J. Coal Geol. 2009, 78, 135–148. [Google Scholar] [CrossRef]
- Narendrula-Kotha, R.; Theriault, G.; Mehes-Smith, M.; Kalubi, K.; Nkongolo, K. Metal toxicity and resistance in plants and microorganisms in terrestrial ecosystems. Rev. Environ. Contam. Toxicol. 2019, 249, 1–27. [Google Scholar] [CrossRef]
- Tobin-Janzen, T.; Shade, A.; Marshall, L.; Torres, K.; Beblo, C.; Janzen, C.; Lenig, J.; Martinez, A.; Ressler, D. Nitrogen changes and do main bacteria ribotype diversity in soils overlying the Centralia, Pennsylvania underground coal mine fire. Soil Sci. 2005, 170, 191–201. [Google Scholar] [CrossRef]
- Bray, E.E.; Evans, E.D. Distribution of n-paraffins as a clue to recognition of source beds. Geochim. Cosmochim. Acta 1961, 22, 2–15. [Google Scholar] [CrossRef]
- Peters, K.E.; Walters, C.C.; Moldowan, J.M. The Biomarker Guide, Biomarkers and Isotopes in Petroleum Exploration and Earth History, 2nd ed.; University Press: Cambridge, UK, 2005; 1155p. [Google Scholar]
- Maeda, H.; DellaPenna, D. Tocopherol functions in photosynthetic organisms. Curr. Opin. Plant Biol. 2007, 10, 260–265. [Google Scholar] [CrossRef]
- Green, G.; Skerratt, J.H.; Leeming, R.; Nichols, P.D. Hydrocarbon and Coprostanol Levels in Seawater, SeaIce Algae and Sed-iments near Davis-Station in Eastern Antarctica—A Regional Survey and Preliminary Results for a Field Fuel Spill Experiment. Mar. Pollut. Bull. 1992, 25, 293–302. [Google Scholar] [CrossRef]
- Readman, J.W.; Fillmann, G.; Tolosa, I.; Bartocci, J.; Mee, L.D. The use of steroid markers to assess sewage contamination of the Black Sea. Mar. Pollut. Bull. 2005, 50, 310–318. [Google Scholar] [CrossRef]
- Abdel-Aal, E.I.; Haroon, A.M.; Mofeed, J. Successive solvent extraction and GC–MS analysis for the evaluation of the phyto-chemical constituents of the filamentous green alga Spirogyra longata. Egypt. J. Aquat. Res. 2015, 41, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.L. Cholesterol autoxidation 1981–1986. Chem. Phys. Lipids 1987, 44, 87–125. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Merkel, J.P. Sterols of freshwater microalgae: Potential implications for zooplankton nutrition. Plank-Ton Res. 2016, 38, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Goad, L.J. Sterol biosynthesis and metabolism in marine invertebrates. Pure. Appl. Chem. 1981, 53, 837–852. [Google Scholar] [CrossRef]
- Patterson, G.W. Sterols of algae. In Physiology and Biochemistry of Sterols; Patterson, G.W., Nes, W.D., Eds.; American Oil Chemists’ Society: Champaign, IL, USA, 1991; pp. 118–157. [Google Scholar]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food. Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quan-titative analysis, and health-promoting uses. Prog. Lipid. Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Mahato, S.B.; Banerjee, S. Microbiological transformations of beta-sitosterol and stigmasterol by a soil pseudomonad. Expe-Rientia 1980, 36, 515–516. [Google Scholar] [CrossRef]
- Svoboda, J.A.; Thompson, M.J.; Robbins, W.E.; Elden, T.C. Unique pathways of sterol metabolism in the Mexican bean beetle, a plant-feeding insect. Lipids 1975, 10, 524–527. [Google Scholar] [CrossRef]
- Huang, W.Y.; Meinschein, W.G. Sterols as ecological indicators. Geochim. Cosmochim. Acta 1979, 43, 739–745. [Google Scholar] [CrossRef]
- Gooday, G.W. The dynamics of hyphal growth. Mycol. Res. 1995, 99, 385–394. [Google Scholar] [CrossRef]
- Steele, D.H.; Thornburg, M.J.; Stanley, J.S.; Miller, R.R.; Brooke, R.; Cushman, J.R.; Cruzan, G. Determination of styrene in selected foods. J. Agric. Food Chem. 1994, 42, 1661–1665. [Google Scholar] [CrossRef]
- Hedges, J.I.; Ertel, J.R. Characterization of lignin by gas capillary chromatography of cupric oxide oxida-tion products. Anal. Chem. 1982, 54, 174–178. [Google Scholar] [CrossRef]
- Jen, J.J.; Mackinney, G. On the photodecomposition of chlorophyll in vitro-11. Intermediate and breakdown products. Photo-Chem. Photobiol. 1970, 11, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Damsté, J.S.S.; ten Haven, H.L.; de Leeuw, J.W.; Schenck, P.A. Organic geochemical studies of a Messinian evaporitic basin, northern Apennines (Italy). II Isoprenoid and n-alkyl thiophenes and thiolanes. Org. Geochem. 1986, 10, 791–805. [Google Scholar] [CrossRef] [Green Version]
- Damsté, J.S.S.; de Leeuw, J.W. The origin and fate of C20 and C15 isoprenoid sulphur compounds in sediments and oils. Int. J. Environ. Anal. Chem. 1987, 28, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, K.; Yasukawa, M.; Muto, N.; Uemura, T.; Ishiwatari, R. Formation of C20 isoprenoid thiophenes in modern sedi-ments. Org. Geochem. 1992, 18, 83–91. [Google Scholar] [CrossRef]
- Morita, K.; Kobayashi, S. Isolation, structure, and synthesis of lenthionine and its analogs. Chem. Pharm. Bull. 1967, 15, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Nádudvari, Á.; Fabiańska, M.J.; Marynowski, L.; Kozielska, B.; Konieczyński, J.; Smołka-Danielowska, D.; Ćmiel, S. Distribu-tion of coal and coal combustion related organic pollutants in the environment of the Upper Silesian Industrial Region. Sci. Total Environ. 2018, 628–629, 1462–1488. [Google Scholar] [CrossRef] [PubMed]
- Poulin, J.; Helwig, K. Class Id resinite from Canada: A new sub-class containing succinic acid. Org. Geochem. 2012, 44, 37–44. [Google Scholar] [CrossRef]
- Nasir, S.; Augie, N.M.; Sani, A.A.; Abbott, G. Geochemical applications and limitations of Pristane to Phytane ratio in deter-mining the redox condition of North Sea oil and Kimmeridge source rocks’ paleoenvironment of deposition. Dutse J. Pure Appl. Sci. 2015, 1, 96–103. [Google Scholar]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señoráns, F.J.; Ibánez, E. Screening for bioactive com-pounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Venkata, R.B.; Samuel, L.A.; Pardha, S.M.; Narashimha, R.B.; Krishna, N.V.; Sudhakar, M.; Radhakrishnan, T.M. Antibacterial, antioxidant activity and GC–MS analysis of Eupatorium odoratum. Asian J. Pharm. Clin. Res. 2012, 5, 99–106. [Google Scholar]
- Zhang, Z.; Sachs, J.P. Hydrogen isotope fractionation in freshwater algae: I. Variations among lipids and species. Org. Geochem. 2007, 38, 582–608. [Google Scholar] [CrossRef]
- Sorigué, D.; Légeret, B.; Cuiné, S.; Morales, P.; Mirabella, B.; Guédeney, G.; Li-Beisson, Y.; Jetter, R.; Peltier, G.; Beisson, F. Microalgae Synthesize Hydrocarbons from Long-Chain Fatty Acids via a Light-Dependent Pathway. Plant Physiol. 2016, 171, 2393–2405. [Google Scholar] [CrossRef] [Green Version]
- Opgrande, J.L.; Brown, E.E.; Hesser, M.; Andrews, J. Benzaldehyde. Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons: Hoboken, NJ, USA, 2001. [Google Scholar]
- Marynowski, L.; Pięta, M.; Janeczek, J. Composition and source of polycyclic aromatic compounds in deposited dust from selected sites around the Upper Silesia, Poland. Geol. Quart. 2004, 48, 169–180. [Google Scholar]
- Safaei-Ghomi, J.; Bamoniri, A.; Hatami, A.; Batooli, H. Composition of the essential oil of Stachys acerosa growing in central. Iran. Chem. Nat. Compd. 2007, 43, 37–39. [Google Scholar] [CrossRef]
- Spychalski, W. Pierwiastki śladowe w glebach wytworzonych z gruntów pogórniczych. Ochr. Sr. Zasobów Nat. 2007, 33, 108–113. [Google Scholar]
- Klojzy-Karczmarczyk, B.; Mazurek, J.; Staszczak, J. Analiza jakości odpadów z nieczynnej hałdy górnictwa węgla ka-miennego w odniesieniu do wymagań stawianych odpadom wydobywczym obojętnym. Zesz. Nauk. Inst. Gos-Podarki Surowcami Miner. Energią Pol. Akad. Nauk 2016, 95, 227–242. [Google Scholar]
- Li, X.; Yang, H.; Zhang, C.; Zeng, G.; Liu, Y.; Xu, W.; Wu, Y.; Lan, S. Spatial distribution and transport characteristics of heavy metals around an antimony mine area in central China. Chemosphere 2017, 170, 17–24. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Mukherjee, A.B. Trace Elements from Soil to Human; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2007; 550p. [Google Scholar]
- Lis, J.; Pasieczna, A. Atlas Geochemiczny Górnego Śląska; Państwowy Instytut Geologiczny: Warszawa, Poland, 1995. [Google Scholar]
- Act of law, 2016, Dz.U. 2016 poz. 1395. Polish Minister of Environment: Warszawa, Poland. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=wdu20160001395 (accessed on 23 July 2022).
- Curyło, T.; Krauze, A.; Kuczyńska, I.; Sapek, B. Liczby graniczne zawartości Fe, Cu, Mn, Zn, Co, J, Se i Mo w roślinności łąk i pastwisk pod kątem oceny ich wartości paszowej. Pract. Kom. Nauk PTG 1985, 93, 43–60. [Google Scholar]
- Gorlach, E. Zawartość pierwiastków śladowych w roślinach pastewnych jako miernik ich wartości. Zesz. Nauk. AR W Kra-Kowie. 1991, 34, 13–22. [Google Scholar]
- Gloser, V. Seasonal changes of nitrogen storage compounds in a rhizomatous grass Calamagrostis epigeios. Biol. Plant. 2002, 45, 563–568. [Google Scholar] [CrossRef]
- Gajić, G.; Pavlović, P.; Kostić, O.; Jarić, S.; Đurđević, L.; Pavlović, D.; Mitrović, M. Ecophysiological and biochemical traits of three herbaceous plants growing on the disposed coal combustion fly ash of different weathering stage. Arch. Biol. Sci. 2014, 65, 1651–1667. [Google Scholar] [CrossRef] [Green Version]
- Ranđelović, D.; Jakovljević, K.; Mihailović, N.; Jovanović, S. Metal accumulation in populations of Calamagrostis epigejos (L.) Roth from diverse anthropogenically degraded sites (SE Europe, Serbia). Environ. Monitor. Assess. 2018, 190, 183. [Google Scholar] [CrossRef]
- Fiala, K.; Tůma, I.; Holub, P. Effect of nitrogen addition and drought on above-ground biomass of expanding tall grasses Calamagrostis epigejos and Arrhenatherum elatius. Biologia 2011, 66, 275–281. [Google Scholar] [CrossRef]
- Schlapfer, B.; Ryser, P. Leaf and root turnover of three ecologically contrasting grass species in relation to their performance along a productivity gradient. Oikos 1996, 75, 398–406. [Google Scholar] [CrossRef]
- Ranđelović, D.; Jakovljević, K.; Jovanović, S. The application of Calamagrostis epigejos (L.) Roth. in phytoremediation tech-nologies. In Phytoremediation Potential of Perennial Grasses; Pande, V.C., Singh, D.P., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2020; pp. 259–282. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhao, Z.; Niu, S.; Li, X.; Wang, Y.; Bai, Z. Reclamation promotes the succession of the soil and vegetation in open-cast coal mine: A case study from Robinia pseudacacia reclaimed forests, Pingshuo mine, China. Catena 2018, 165, 72–79. [Google Scholar] [CrossRef]
- Samecka-Cymerman, A.; Stankiewicz, A.; Kolon, K.; Kempers, A.J. Self-organizing feature map (neural networks) as a tool to select the best indicator of road traffic pollution (soil, leaves or bark of Robinia pseudacacia L.). Environ. Pollut. 2009, 157, 2061–2065. [Google Scholar] [CrossRef] [PubMed]
- Ussiri, D.A.N.; Lal, R.; Jacinthe, P.A. Soil Properties and Carbon Sequestration of Afforested Pastures in Reclaimed Minesoils of Ohio. Soil Sci. Soc. Am. J. 2006, 70, 1797–1806. [Google Scholar] [CrossRef]
- Tilman, D.; Lehman, C. Human-caused environmental change: Impacts on plant diversity and evolution. Proc. Natl. Acad. Sci. USA 2001, 98, 5433–5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmonov, O.; Różkowski, J.; Klys, G. The Managing and Restoring of Degraded Land in Post-Mining Areas. Land 2022, 11, 269. [Google Scholar] [CrossRef]
- Palowski, B.; Małkowska, E.; Kurtyka, R.; Szymanowska-Pułka, J.; Gucwa-Przepióra, E.; Małkowski, Ł.; Woźnica, A.; Małkowski, E. Bioaccumulation of heavy metals in selected organs of black locust (Robinia pseudacacia) and their poten-tial use as air contamination bioindicators. Pol. J. Environ. Study 2016, 25, 2085–2096. [Google Scholar] [CrossRef]
- Tissot, B.; Welte, D.H. Petroleum Formation and Occurrence: A New Approach to Oil and Gas Exploration, 2nd ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1984; p. 699. [Google Scholar]
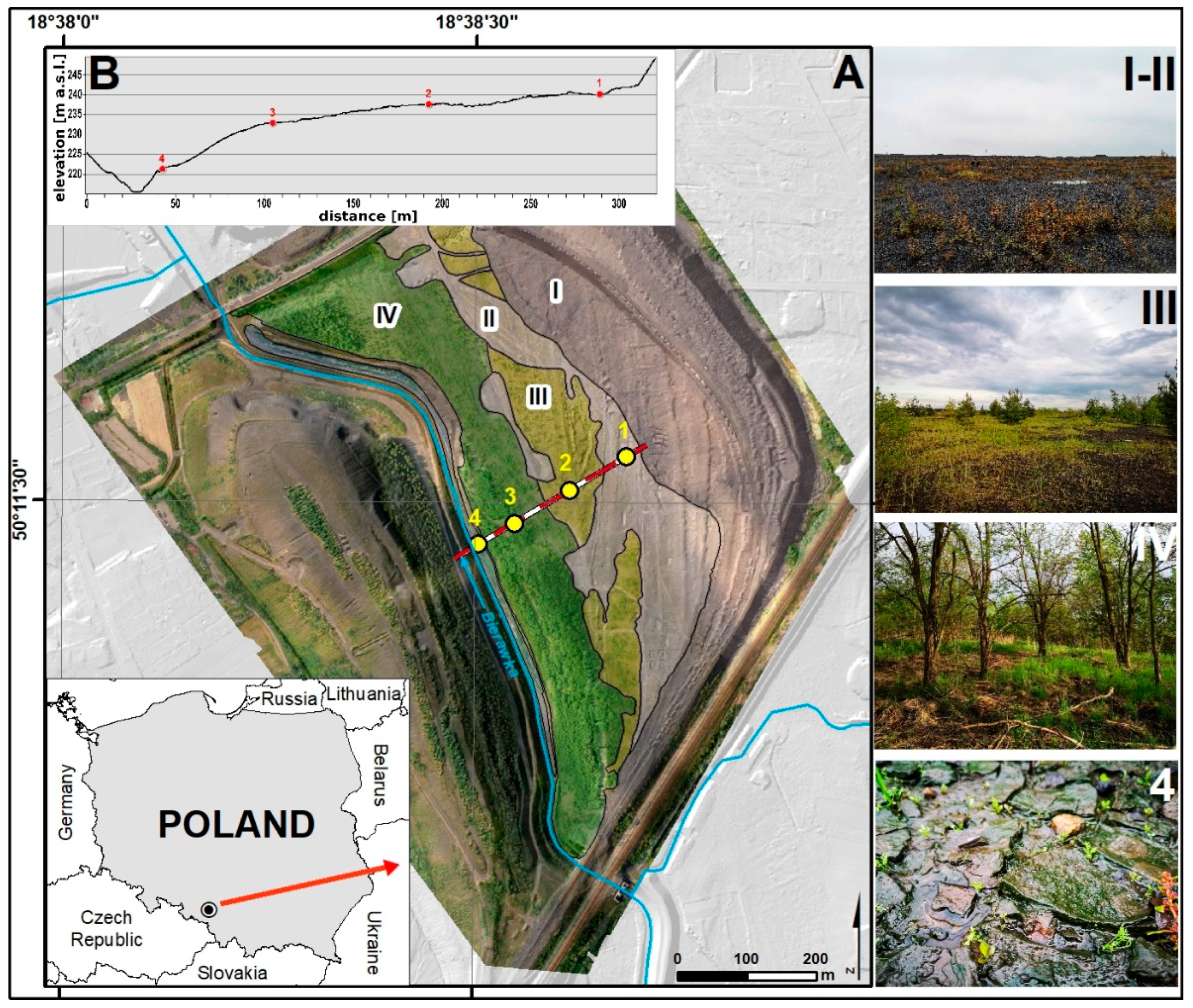
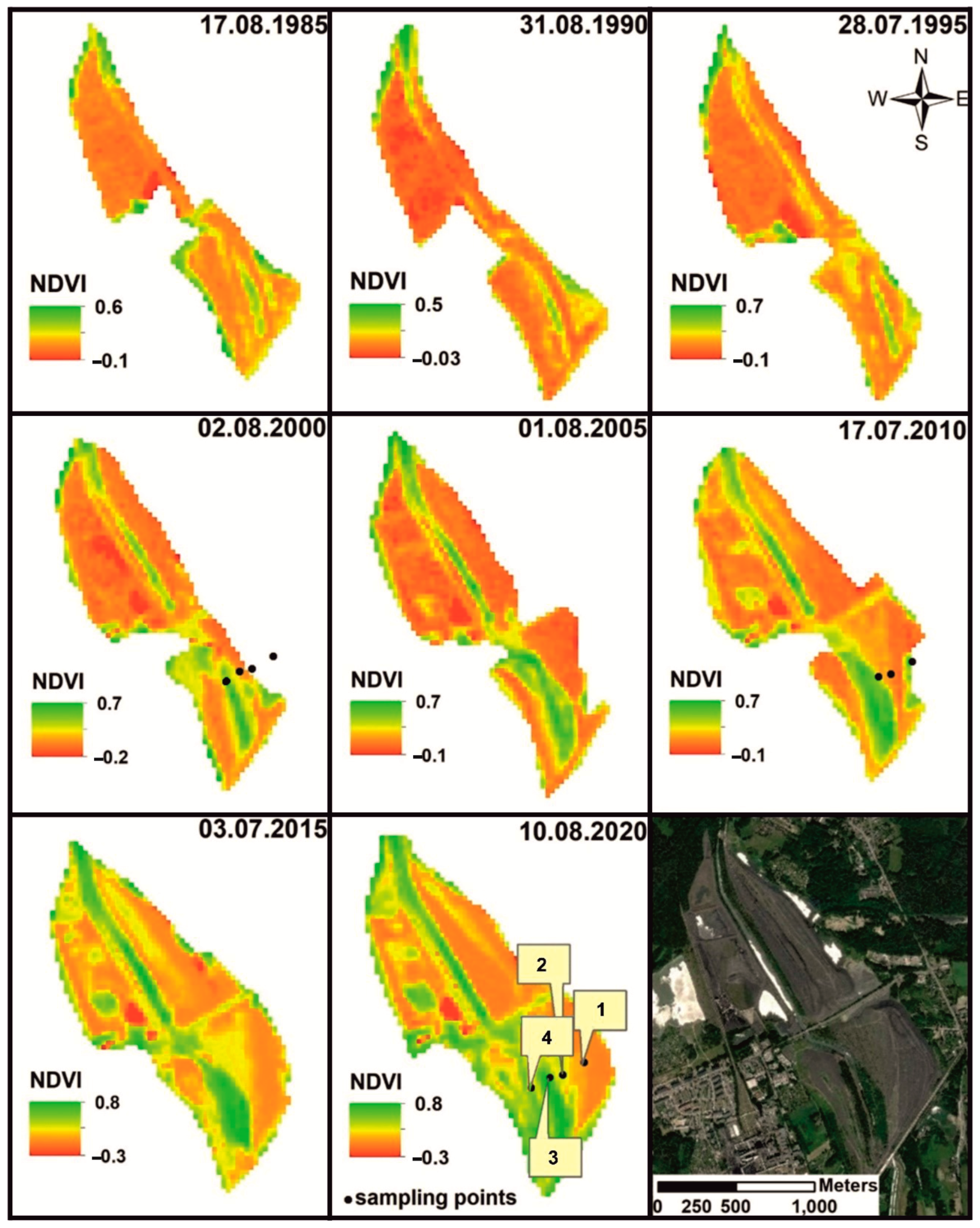

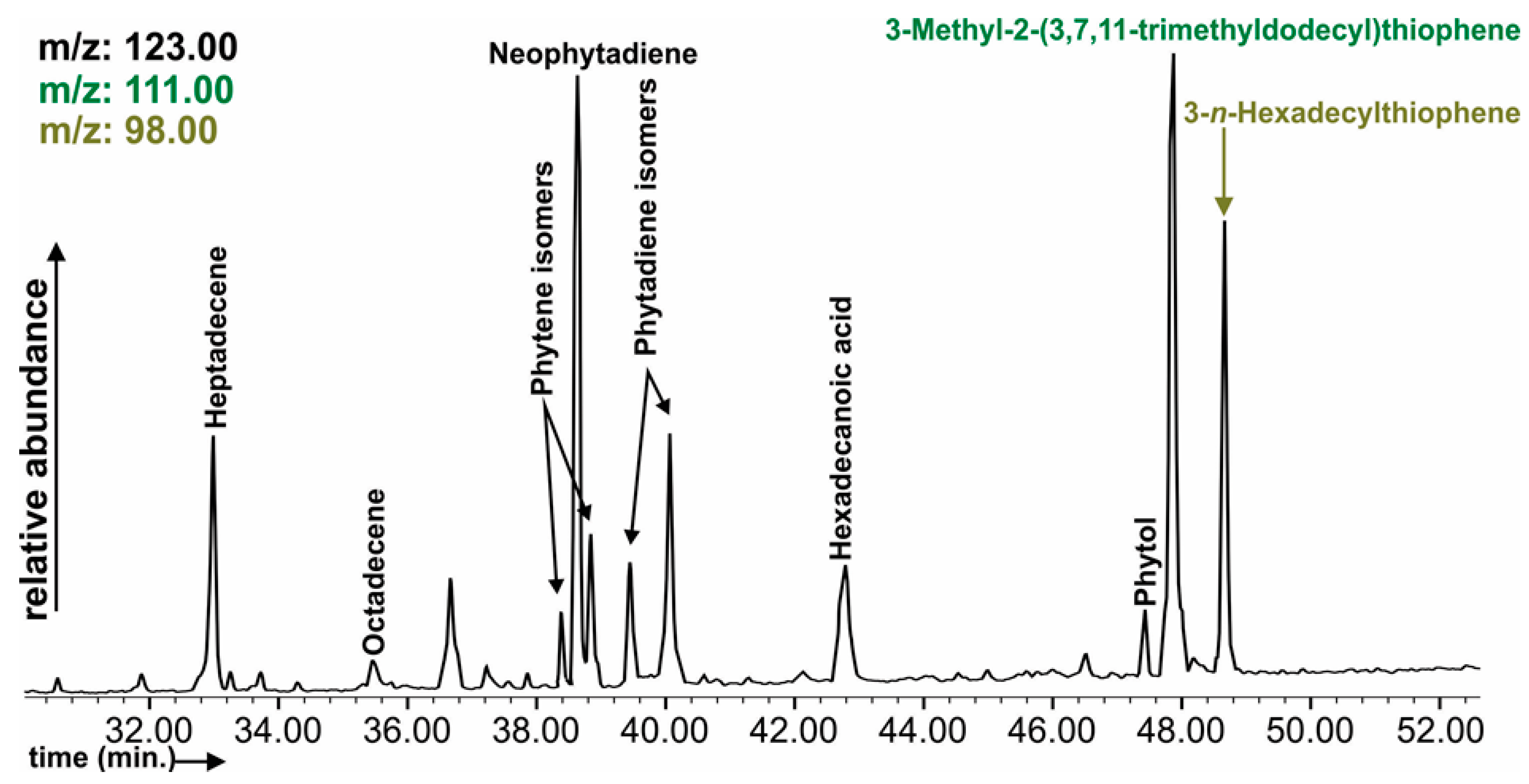

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [mm] | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Horizon | Depth [cm] | >20.0 | 20.0–10.0 | 10.0–5.0 | 5.0–2.0 | 2.0–1.0 | 1.0–0.5 | 0.5–0.25 | 0.25–0.1 | 0.1–0.05 | 0.05–0.02 | 0.02–0.006 | 0.006–0.002 | <0.002 |
| [%] | ||||||||||||||
| Profile 1 Communities with Chamaenerion palustre | ||||||||||||||
| C1 | 0–14 | 9.7 | 20.0 | 17.3 | 21.1 | 12.9 | 4.0 | 5.5 | 2.7 | 2.3 | 0.9 | 1.7 | 1.1 | 0.8 |
| C1 | 14–90 | 4.1 | 22.4 | 20.6 | 21.9 | 13.8 | 4.6 | 5.1 | 2.0 | 1.4 | 0.7 | 1.0 | 0.9 | 1.5 |
| Profile 2 Communities with Calamagrostis epigejos | ||||||||||||||
| A | 0–13 | 4.2 | 12.9 | 17.1 | 20.5 | 12.0 | 3.6 | 9.9 | 4.7 | 3.0 | 2.3 | 4.3 | 1.8 | 3.7 |
| ACq | 13–45 | - | 11.4 | 17.9 | 24.4 | 10.5 | 6.4 | 6.9 | 3.9 | 2.5 | 1.8 | 4.6 | 2.9 | 6.8 |
| Cq | 45–65 | 9.8 | 25.7 | 24.6 | 17.2 | 7.9 | 3.7 | 4.3 | 1.7 | 1.2 | 0.7 | 1.8 | 1.0 | 0.4 |
| Profile 3 Communities with Robinia pseudacacia | ||||||||||||||
| Olfh | 4–0 | 3.7 | 12.1 | 4.2 | 4.5 | 8.6 | 10.2 | 8 | 16.8 | 13.6 | 7.2 | 11.1 | ||
| A | 0–23 | - | 16.3 | 11.7 | 14.1 | 16.0 | 2.6 | 5.7 | 4.8 | 3.0 | 5.5 | 5.5 | 4.3 | 10.5 |
| ACq | 23–60 | - | 13.7 | 20.9 | 27.5 | 14.4 | 5.6 | 5.5 | 2.5 | 1.6 | 1.9 | 2.3 | 1.5 | 2.6 |
| Cq | 60–80 | 7.5 | 19.1 | 20.9 | 22.4 | 5.7 | 2.2 | 3.9 | 2.7 | 1.7 | 3.2 | 3.4 | 2.2 | 5.1 |
| Profile 4 Organic mineral crusts with green algae | ||||||||||||||
| (1) OC | 0.4 | - | - | 1.7 | 4.1 | 11.8 | 1.6 | 1.7 | 1.8 | 3.4 | 4.6 | 23.9 | 19.8 | 25.6 |
| (2) OC | 0.5 | - | - | 0.4 | 3.3 | 6.5 | 2.7 | 4.6 | 5.3 | 9.9 | 34.0 | 18.0 | 4.5 | 10.8 |
| (3) OC | 0.6 | - | - | - | - | 1.1 | 0.9 | 1.1 | 1.1 | 3.9 | 28.7 | 35.6 | 14.8 | 12.8 |
| (4) OC | 0.5 | - | - | - | - | 1.9 | 0.9 | 1.3 | 0.9 | 2.9 | 23.5 | 40.2 | 15.7 | 12.7 |
| (5) OC | 0.9 | - | - | 0.6 | 2.0 | 9.3 | 1.2 | 3.1 | 2.8 | 5.3 | 27.3 | 29.9 | 11.5 | 7.0 |
| Soil Horizons | Depth [cm] | pH | Loss on Ignition | Corg. | Nt | C/N | Mg Avail. | P Avail. | Pt | K Avail. | Hh [cmol(+)/kg] | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H2O | KCl | [%] | [mg·kg−1] | |||||||||
| Profile 1 Chamaenerion palustre | ||||||||||||
| C1 | 0–14 | 6.9 | 6.7 | 34.75 | 25.8 | 0.538 | 48 | 343.5 | 0.90 | 558.0 | 197.0 | 1.92 |
| C1 | 14–90 | 6.6 | 6.5 | 34.14 | 31.2 | 0.316 | 99 | 498.0 | 1.00 | 572.0 | 249.0 | 1.84 |
| Profile 2 Communities with Calamagrostis epigejos | ||||||||||||
| A | 0–13 | 6.9 | 6.5 | 30.86 | 26.7 | 0.688 | 39 | 319.5 | 0.00 | 387.0 | 139.0 | 2.00 |
| ACq | 13–45 | 7.1 | 6.8 | 43.48 | 41.6 | 0.531 | 78 | 284.5 | 1.05 | 705.0 | 159.0 | 1.60 |
| Cq | 45–65 | 4.9 | 4.8 | 40.85 | 32.9 | 0.754 | 44 | 475.0 | 1.40 | 578.0 | 136.5 | 5.68 |
| Profile 3 Communities with Robinia pseudacacia | ||||||||||||
| Olfh | 4–0 | 5.8 | 5.3 | 46.5 | 26.4 | 0.751 | 35 | 299.0 | 0.70 | 440.0 | 191.5 | 4.36 |
| A | 0–23 | 4.8 | 4.1 | 18.73 | 10.7 | 0.884 | 12 | 326.0 | 1.40 | 196.0 | 143.0 | 5.36 |
| ACq | 23–60 | 5.5 | 5.2 | 32.84 | 27.5 | 0.402 | 68 | 357.5 | 1.92 | 281.6 | 132.0 | 3.76 |
| Cq | 60–80 | 5.4 | 5.1 | 24.37 | 14.5 | 0.706 | 21 | 393.5 | 0.60 | 318.0 | 143.0 | 3.92 |
| Profile 4 Organic mineral crusts with green algae | ||||||||||||
| (1) OC | 0.4 | 7.6 | 7.1 | 24.82 | 20.7 | 0.297 | 70 | 481.0 | 0.00 | 455.0 | 251.5 | 2.00 |
| (2) OC | 0.5 | 6.8 | 6.5 | 14.83 | 9.6 | 1.132 | 8 | 263.5 | 16.20 | 954.0 | 207.0 | 3.12 |
| (3) OC | 0.6 | 7.1 | 6.9 | 21.46 | 14.0 | 0.450 | 31 | 345.5 | 7.32 | 1588.0 | 199.0 | 2.32 |
| (4) OC | 0.5 | 7.1 | 6.8 | 22.47 | 16.1 | 0.446 | 36 | 507.0 | 7.40 | 1580.0 | 202.0 | 2.80 |
| (5) OC | 0.9 | 7.4 | 7.3 | 22.12 | 16.0 | 0.610 | 26 | 670.0 | 9.77 | 1310.0 | 277.0 | 2.04 |
| Ca | K | Na | Mg | P | Fe | S | Al | |
|---|---|---|---|---|---|---|---|---|
| [mg·kg−1] | ||||||||
| plants parts | plant materials | |||||||
| Chamaenerion palustre | ||||||||
| aboveground | 7000 | 6200 | 30 | 3480 | 710 | 310 | 2200 | <100 |
| roots | 8400 | 5700 | 30 | 4040 | 530 | 710 | 1600 | 200 |
| Calamagrostis epigejos | ||||||||
| aboveground | 1400 | 9700 | 20 | 850 | 1118 | 250 | 1300 | <100 |
| roots | 1100 | 3200 | 110 | 760 | 8600 | 12,610 | 2200 | 4000 |
| Robinia pseudacacia | ||||||||
| leaves | 12,400 | 13,700 | 3 | 3140 | 1350 | 170 | 2100 | <100 |
| bark | 16,800 | 3300 | 2 | 510 | 320 | 210 | 1700 | <100 |
| roots | 3800 | 11,600 | 5 | 1320 | 1030 | 2510 | 3000 | 1000 |
| pods | 2900 | 9300 | <1 | 1470 | 2510 | 6 | 1900 | <100 |
| horizon and depth [cm] | soil materials | |||||||
| Profile 1 under Chamaenerion palustre | ||||||||
| C1 (0–14) | 4200 | 1400 | 80 | 3100 | 580 | 15,900 | 3500 | 5400 |
| C2 (14–90) | 5700 | 1600 | 90 | 4500 | 530 | 18,100 | 5800 | 5500 |
| Profile 2 under Calamagrostis epigejos | ||||||||
| A (0–13) | 6600 | 1500 | 90 | 4700 | 310 | 13,800 | 2600 | 6100 |
| ACq (13–45) | 3200 | 1600 | 90 | 2600 | 340 | 18,700 | 7700 | 5300 |
| Cq (45–65) | 1600 | 1700 | 90 | 1600 | 300 | 18,800 | 9600 | 5200 |
| Profile 3 under Robinia pseudacacia | ||||||||
| A (0–23) | 1500 | 1600 | 70 | 2700 | 290 | 26,200 | 1300 | 6600 |
| ACq (23–60) | 1000 | 1600 | 70 | 1900 | 140 | 11,700 | 2100 | 5600 |
| Cq (60–80) | 1600 | 1700 | 70 | 2100 | 380 | 16,300 | 2300 | 6700 |
| Zn | Pb | Cd | Co | Ni | Hg | Cu | As | Cr | |
|---|---|---|---|---|---|---|---|---|---|
| [mg·kg−1] | |||||||||
| plant parts | plant materials | ||||||||
| Chamaenerion palustre | |||||||||
| aboveground | 65.4 | 2.89 | 0.19 | 0.44 | 1.8 | 0.023 | 19.83 | <0.1 | 2.3 |
| roots | 35.4 | 2.06 | 0.11 | 1.16 | 4.1 | 0.01 | 8.71 | <0.1 | 2.1 |
| Calamagrostis epigejos | |||||||||
| aboveground | 22.8 | 0.98 | 0.16 | 0.35 | 1.6 | 0.01 | 2.98 | 0.3 | 1.4 |
| roots | 70.0 | 21.16 | 1.12 | 17.50 | 25.9 | 0.193 | 37.53 | 10.3 | 9.8 |
| Robinia pseudacacia | |||||||||
| leaves | 29.4 | 0.99 | 0.14 | 0.52 | 5.0 | 0.028 | 4.88 | <0.1 | 1.7 |
| bark | 14.8 | 3.32 | 0.18 | 0.51 | 2.2 | 0.007 | 5.39 | <0.1 | 1.7 |
| roots | 35.3 | 5.81 | 0.26 | 3.26 | 12.2 | 0.031 | 15.54 | 0.9 | 4.2 |
| pods | 27.7 | 0.21 | 0.08 | 0.77 | 10.1 | >0.001 | 9.76 | <0.1 | 1.8 |
| horizon and depth [cm] | soil materials | ||||||||
| Profile 1—with Chamaenerion palustre | |||||||||
| C1 (0–14) | 49 | 23.6 | <0.1 | 7.7 | 18.1 | 0.08 | 39.7 | 3.9 | 8 |
| C2 (14–90) | 42 | 21.2 | <0.1 | 8.8 | 21.8 | 0.09 | 39.5 | 4.0 | 9 |
| Profile 2—with Calamagrostis epigejos | |||||||||
| A (0–13) | 50 | 21.4 | 0.2 | 7.6 | 22.9 | 0.13 | 32.7 | 5.1 | 10 |
| ACq (13–45) | 44 | 21.5 | <0.1 | 8.7 | 25.4 | 0.20 | 35.7 | 10.0 | 9 |
| Cq (45–65) | 46 | 25.2 | <0.1 | 10.7 | 31.5 | 0.28 | 40.2 | 14.4 | 8 |
| Profile 3—with Robinia pseudacacia | |||||||||
| A (0–23) | 74 | 17.2 | 0.2 | 8.5 | 25.5 | 0.09 | 26.7 | 4.8 | 14 |
| ACq (23–60) | 46 | 16.5 | <0.1 | 8.7 | 24.3 | 0.07 | 31.7 | 2.4 | 10 |
| Cq (60–80) | 62 | 20.5 | 0.2 | 10.2 | 27.4 | 0.13 | 30.7 | 5.6 | 12 |
| Sample | CRUST WITH ALGAE (Profile 4) | |||||||
|---|---|---|---|---|---|---|---|---|
| [mg·kg−1] | ||||||||
| Ca | K | Na | Mg | P | Fe | S | Al | |
| 1 (OC) | 5400 | 2100 | 570 | 4000 | 1100 | 19,100 | 2600 | 8700 |
| 2 (OC) | 4800 | 1500 | 200 | 2800 | 1550 | 21,400 | 2700 | 8400 |
| 3 (OC) | 9000 | 1800 | 220 | 4600 | 2460 | 28,800 | 3200 | 13,000 |
| 4 (OC) | 12,700 | 1900 | 330 | 5500 | 2820 | 33,600 | 3500 | 14,400 |
| 5 (OC) | 5900 | 2000 | 540 | 3800 | 1240 | 18,800 | 2400 | 9400 |
| Sample | CRUST WITH ALGAE (Profile 4) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| [mg·kg−1] | |||||||||
| Zn | Pb | Cd | Co | Ni | Hg | Cu | As | Cr | |
| 1 (OC) | 277 | 41.8 | 1.4 | 15.9 | 31.9 | 0.12 | 52.9 | 6.9 | 23 |
| 2 (OC) | 308 | 41.8 | 1.8 | 12.4 | 27.6 | 0.14 | 41.4 | 6.7 | 26 |
| 3 (OC) | 649 | 66.4 | 3.4 | 16.9 | 36.3 | 0.18 | 63.7 | 11.0 | 44 |
| 4 (OC) | 736 | 76.0 | 4.2 | 19.7 | 40.6 | 0.24 | 72.9 | 12.6 | 50 |
| 5 (OC) | 319 | 38.2 | 1.7 | 13.0 | 29.6 | 0.11 | 42.6 | 6.8 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmonov, O.; Czajka, A.; Nádudvari, Á.; Fajer, M.; Spórna, T.; Szypuła, B. Soil and Vegetation Development on Coal-Waste Dump in Southern Poland. Int. J. Environ. Res. Public Health 2022, 19, 9167. https://doi.org/10.3390/ijerph19159167
Rahmonov O, Czajka A, Nádudvari Á, Fajer M, Spórna T, Szypuła B. Soil and Vegetation Development on Coal-Waste Dump in Southern Poland. International Journal of Environmental Research and Public Health. 2022; 19(15):9167. https://doi.org/10.3390/ijerph19159167
Chicago/Turabian StyleRahmonov, Oimahmad, Agnieszka Czajka, Ádám Nádudvari, Maria Fajer, Tomasz Spórna, and Bartłomiej Szypuła. 2022. "Soil and Vegetation Development on Coal-Waste Dump in Southern Poland" International Journal of Environmental Research and Public Health 19, no. 15: 9167. https://doi.org/10.3390/ijerph19159167
APA StyleRahmonov, O., Czajka, A., Nádudvari, Á., Fajer, M., Spórna, T., & Szypuła, B. (2022). Soil and Vegetation Development on Coal-Waste Dump in Southern Poland. International Journal of Environmental Research and Public Health, 19(15), 9167. https://doi.org/10.3390/ijerph19159167