
The Effect of Endurance Training on Pulmonary

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Anthropometric Characteristics
2.2. Study Design
2.3. Double Leg Incremental Step Test
2.4. Constant Load Exercises
2.5. Endurance Training
2.6. Data Treatment
2.7. Statistics
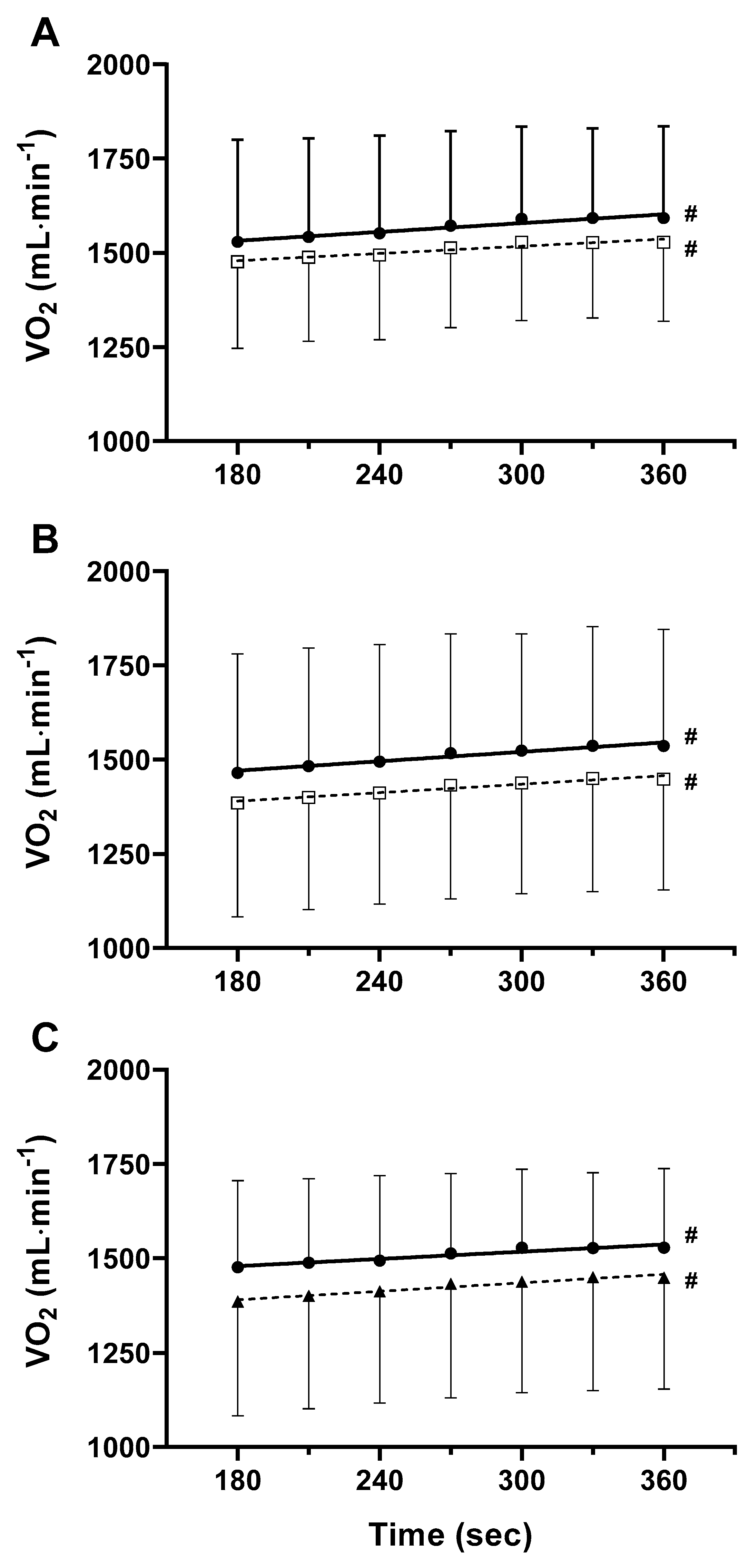
3. Results
3.1. Patient Characteristics and the Exercise Training Regimen
3.2. Double Leg Moderate Constant Load Test
3.3. Double Leg Heavy Constant Load Test
4. Discussion
4.1. O2p Kinetics Parameters and Moderate-Intensity Exercise
4.2. Exercise Responses to Heavy Intensity Exercise
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, T.J.; Mckenna, M.J. Exercise Limitation Following Transplantation. Compr. Physiol. 2012, 2, 1937–1979. [Google Scholar] [CrossRef] [PubMed]
- Duscha, B.D.; Slentz, C.A.; Johnson, J.L.; Houmard, J.A.; Bensimhon, D.R.; Knetzger, K.J.; Kraus, W.E. Effects of exercise training amount and intensity on peak oxygen consumption in middle-age men and women at risk for cardiovascular disease. Chest 2005, 128, 2788–2793. [Google Scholar] [CrossRef] [PubMed]
- Grassi, B.; Porcelli, S.; Salvadego, D.; Zoladz, J.A. Slow O2 kinetics during moderate-intensity exercise as markers of lower metabolic stability and lower exercise tolerance. Eur. J. Appl. Physiol. 2011, 111, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Jones, A.M. Oxygen uptake kinetics. Compr. Physiol. 2012, 2, 933–996. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Schaffartzik, W.; Knight, D.R.; Derion, T.; Kennedy, B.; Guy, H.J.; Prediletto, R.; Wagner, P.D. Contribution of exercising legs to the slow component of oxygen uptake kinetics in humans. J. Appl. Physiol. 1991, 71, 1245–1253. [Google Scholar] [CrossRef]
- Colosio, A.L.; Caen, K.; Bourgois, J.G.; Boone, J.; Pogliaghi, S. Bioenergetics of the VO2 slow component between exercise intensity domains. Pflug. Arch. Eur. J. Physiol. 2020, 472, 1447–1456. [Google Scholar] [CrossRef]
- Tam, E.; Bruseghini, P.; Capelli, C.; Oliboni, E.; Pezzato, A.; Pogliaghi, S.; Mucelli, R.P.; Schena, F.; Calabria, E. Effect of endurance and strength training on the slow component of O2 kinetics in elderly humans. Front. Physiol. 2018, 9, 1353. [Google Scholar] [CrossRef]
- Grassi, B.; Porcelli, S.; Marzorati, M.; Lanfranconi, F.; Vago, P.; Marconi, C.; Morandi, L. Metabolic myopathies: Functional evaluation by analysis of oxygen uptake kinetics. Med. Sci. Sports Exerc. 2009, 41, 2120–2127. [Google Scholar] [CrossRef]
- Grassi, B. Regulation of oxygen consumption at exercise onset: Is it really controversial? Exerc. Sport Sci. Rev. 2001, 29, 134–138. [Google Scholar] [CrossRef]
- Willis, W.T.; Jackman, M.R. Mitochondrial function during heavy exercise. Med. Sci. Sport Exer. 1994, 26, 1347–1353. [Google Scholar] [CrossRef]
- Barstow, T.J.; Jones, A.M.; Nguyen, P.H.; Casaburi, R. Influence of muscle fiber type and pedal frequency on oxygen uptake kinetics of heavy exercise. J. Appl. Physiol. 1996, 81, 1642–1650. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.A.; Gladden, L.B.; Hogan, M.C.; Nieckarz, Z.; Grassi, B. Progressive recruitment of muscle fibers is not necessary for the slow component of O2 kinetics. J. Appl. Physiol. 2008, 105, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Mercier, J.G.; Hokanson, J.F.; Brooks, G.A. Effects of cyclosporine A on skeletal muscle mitochondrial respiration and endurance time in rats. Am. J. Respir. Crit. Care Med. 1995, 151, 1532–1536. [Google Scholar] [CrossRef] [PubMed]
- Hokanson, J.F.; Mercier, J.G.; Brooks, G.A. Cyclosporine A decreases rat skeletal muscle mitochondrial respiration in vitro. Am. J. Respir. Crit. Care Med. 1995, 151, 1848–1851. [Google Scholar] [CrossRef] [PubMed]
- Patti, A.; Neunhaeuserer, D.; Ortolan, S.; Roman, F.; Gasperetti, A.; Battista, F.; Di Bella, C.; Gobbo, S.; Bergamin, M.; Furian, L.; et al. A clinical evaluation of VO2 kinetics in kidney transplant recipients. Eur. J. Appl. Physiol. 2021, 121, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Tomczak, C.R.; Warburton, D.E.R.; Riess, K.J.; Jendzjowsky, N.G.; Esch, B.T.; Liang, Y.; Haennel, R.G.; Haykowsky, M.J. Pulmonary oxygen uptake and heart rate kinetics during the six-minute walk test in transplant recipients. Transplantation 2008, 85, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Jendzjowsky, N.G.; Tomczak, C.R.; Lawrance, R.; Taylor, D.A.; Tymchak, W.J.; Riess, K.J.; Warburton, D.E.R.; Haykowsky, M.J. Impaired pulmonary oxygen uptake kinetics and reduced peak aerobic power during small muscle mass exercise in heart transplant recipients. J. Appl. Physiol. 2007, 103, 1722–1727. [Google Scholar] [CrossRef][Green Version]
- Grassi, B.; Marconi, C.; Meyer, M.; Rieu, M.; Cerretelli, P. Gas exchange and cardiovascular kinetics with different exercise protocols in heart transplant recipients. J. Appl. Physiol. 1997, 82, 1952–1962. [Google Scholar] [CrossRef]
- Tomczak, C.R.; Tymchak, W.J.; Haykowsky, M.J. Effect of exercise training on pulmonary oxygen uptake kinetics in heart transplant recipients. Am. J. Cardiol. 2013, 112, 1489–1492. [Google Scholar] [CrossRef]
- Inglis, E.C.; Iannetta, D.; Murias, J.M. Association between O2 kinetics and O2max in groups differing in fitness status. Eur. J. Appl. Physiol. 2021, 121, 1921–1931. [Google Scholar] [CrossRef]
- Skattebo, Ø.; Bjerring, A.W.; Auensen, M.; Sarvari, S.I.; Cumming, K.T.; Capelli, C.; Hallén, J. Blood volume expansion does not explain the increase in peak oxygen uptake induced by 10 weeks of endurance training. Eur. J. Appl. Physiol. 2020, 120, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Skattebo, Ø.; Capelli, C.; Rud, B.; Auensen, M.; Calbet, J.A.L.; Hallén, J. Increased oxygen extraction and mitochondrial protein expression after small muscle mass endurance training. Scand. J. Med. Sci. Sports 2020, 30, 1615–1631. [Google Scholar] [CrossRef]
- Corvino, R.B.; Oliveira, M.F.M.; Denadai, B.S.; Rossiter, H.B.; Caputo, F. Speeding of oxygen uptake kinetics is not different following low-intensity blood-flow-restricted and high-intensity interval training. Exp. Physiol. 2019, 104, 1858–1867. [Google Scholar] [CrossRef] [PubMed]
- Abbiss, C.R.; Karagounis, L.G.; Laursen, P.B.; Peiffer, J.J.; Martin, D.T.; Hawley, J.A.; Fatehee, N.N.; Martin, J.C. Single-leg cycle training is superior to double-leg cycling in improving the oxidative potential and metabolic profile of trained skeletal muscle. J. Appl. Physiol. 2011, 110, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.; Paterson, D.H.; Kowalchuk, J.M.; Moy, A.P.; Thorp, D.B.; Noble, E.G.; Taylor, A.W.; Cunningham, D.A. Determinants of oxygen uptake kinetics in older humans following single-limb endurance exercise training. Exp. Physiol. 2001, 86, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Borg, G.A.V. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]
- Lepretre, P.M.; Koralsztein, J.P.; Billat, V.L. Effect of exercise intensity on relationship between O2max and cardiac output. Med. Sci. Sports Exerc. 2004, 36, 1357–1363. [Google Scholar] [CrossRef]
- Whipp, B.J.; Davis, J.A.; Torres, F.; Wasserman, K. A test to determine parameters of aerobic function during exercise. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 50, 217–221. [Google Scholar] [CrossRef]
- Herman, T.; Dall, C.; Christensen, S.; Goetze, J.; Prescott, E.; Gustafsson, F. Effect of High Intensity Exercise on Peak Oxygen Uptake and Endothelial Function in Long-Term Heart Transplant Recipients. Am. J. Transplant. 2011, 11, 536–541. [Google Scholar] [CrossRef]
- Buchheit, M.; Laursen, P.B. High-intensity interval training, solutions to the programming puzzle: Part I: Cardiopulmonary emphasis. Sport. Med. 2013, 43, 313–338. [Google Scholar] [CrossRef]
- Ciolac, E.G.; Mantuani, S.S.; Neiva, C.M.; Verardi, C.E.L.; Pessôa-Filho, D.M.; Pimenta, L. Rating of perceived exertion as a tool for prescribing and self regulating interval training: A pilot study. Biol. Sport 2015, 32, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Capelli, C.; Adami, A.; Antonutto, G.; Cautero, M.; Tam, E. Oxygen deficits and oxygen delivery kinetics during submaximal intensity exercise in humans after 14 days of head-down tilt-bed rest. Eur. J. Appl. Physiol. 2009, 107, 51–59. [Google Scholar] [CrossRef]
- Whipp, B.J.; Rossiter, H.B. The kinetics of oxygen uptake. In Oxygen Uptake Kinetics in Sports, Exercise and Medicine; Jones, A.M., Poole, D.C., Eds.; Routledge: London, UK, 2005; pp. 62–94. [Google Scholar]
- Lamarra, N.; Whipp, B.J.; Ward, S.A.; Wasserman, K. Effect of interbreath fluctuations on characterizing exercise gas exchange kinetics. J. Appl. Physiol. 1987, 62, 2003–2012. [Google Scholar] [CrossRef] [PubMed]
- Peronnet, F.; Massicotte, D. Table of nonprotein respiratory quotient: An update. Can. J. Sport Sci. 1991, 16, 23–29. [Google Scholar] [PubMed]
- Salvadego, D.; Sartorio, A.; Agosti, F.; Tringali, G.; Patrizi, A.; Isola, M.; LoMauro, A.; Aliverti, A.; Grassi, B. Respiratory muscle endurance training reduces the O2 cost of cycling and perceived exertion in obese adolescents. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R487–R495. [Google Scholar] [CrossRef] [PubMed]
- Billat, V.L.; Richard, R.; Binsse, V.M.; Koralsztein, J.P.; Haouzi, P. The O2 slow component for severe exercise depends on type of exercise and is not correlated with time to fatigue. J. Appl. Physiol. 1998, 85, 2118–2124. [Google Scholar] [CrossRef]
- Motulsky, H.; Christopoulos, A. Fitting Models to Biological Data Using Linear and Non-Linear Regression: A Practical Guide to Curve Fitting; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Zar, Z.H. Biostatistical Analysis; Prentice-Hall: Englewood Cliffs, NJ, USA, 1974. [Google Scholar]
- Del Vecchio, A.; Casolo, A.; Negro, F.; Scorcelletti, M.; Bazzucchi, I.; Enoka, R.; Felici, F.; Farina, D. The increase in muscle force after 4 weeks of strength training is mediated by adaptations in motor unit recruitment and rate coding. J. Physiol. 2019, 597, 1873–1887. [Google Scholar] [CrossRef]
- Marquardt, D.M. An algorithm for least-squares estimation of nonlinear parameters. Soc. Ind. J. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
- MacPhee, S.L.; Shoemaker, J.K.; Paterson, D.H.; Kowalchuk, J.M. Kinetics of O2 uptake, leg blood flow, and muscle deoxygenation are slowed in the upper compared with lower region of the moderate-intensity exercise domain. J. Appl. Physiol. 2005, 99, 1822–1834. [Google Scholar] [CrossRef]
- Kempeneers, G.; Noakes, T.D.; van Zyl-Smit, R.; Myburgh, K.H.; Lambert, M.; Adams, B.; Wiggins, T. Skeletal Muscle Limits the Exercise Tolerance of Renal Transplant Recipients: Effects of a Graded Exercise Training Program. Am. J. Kidney Dis. 1990, 16, 57–65. [Google Scholar] [CrossRef]
- Richard, R.; Verdier, J.; Doutreleau, S.; Piquard, F. Exercise Limitation in Trained Heart and Kidney Transplant Recipients: Central and Peripheral Limitations. J. Heart Lung Transplant. 2005, 24, 1774–1780. [Google Scholar] [CrossRef]
- Jones, A.M.; Carter, H. The effect of endurance training on parameters of aerobic fitness./Effet de l’entrainement d’endurance sur les parametres de la capacite aerobie. Sport. Med. 2000, 29, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.; Koziel, A.; Woyda-PLoSzczyca, A.; Celichowski, J.; Jarmuszkiewicz, W. Endurance training increases the efficiency of rat skeletal muscle mitochondria. Pflug. Arch. 2016, 468, 1709–1724. [Google Scholar] [CrossRef]
- Holloszy, J.O. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J. Biol. Chem. 1967, 242, 2278–2282. [Google Scholar] [CrossRef]
- Carra, J.; Candau, R.; Keslacy, S.; Giolbas, F.; Borrani, F.; Millet, G.P.; Varray, A.; Ramonatxo, M. Addition of inspiratory resistance increases the amplitude of the slow component of O2 uptake kinetics. J. Appl. Physiol. 2003, 94, 2448–2455. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Grassi, B.; Christensen, P.M.; Krustrup, P.; Bangsbo, J.; Poole, D.C. Slow component of O2 kinetics: Mechanistic bases and practical applications. Med. Sci. Sports Exerc. 2011, 43, 2046–2062. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Anthropometrics | SL-ETGRP (n = 17) | DL-ETGRP (n = 16) | p |
|---|---|---|---|
| Age (years) | 56 (10) | 55 (10) | 0.82 |
| BM (Kg) | 83 (15) | 78(18) | 0.35 |
| BMI | 26.7 (3.3) | 26.3 (5.3) | 0.76 |
| Years post-transplant | 6.2 (6.9) | 8.9 (7.7) | 0.30 |
| Cardio-respiratory parameters | |||
| O2peak (mL·min−1) | 1747 (420) | 1719 (483) | 0.86 |
| O2peak (mL·kg−1·min−1) | 21.8 (7.7) | 22.2 (4.7) | 0.85 |
| Epeak (L·min−1) | 78.6 (16.9) | 77.5 (21.3) | 0.88 |
| O2 at VT1 (mL·min−1) | 1311 (216) | 1275 (290) | 0.55 |
| HRpeak (bpm) | 124 (24) | 143 (22) | 0.03 |
| SpO2 (%) | 96 (2) | 96 (3) | 0.93 |
| Peak power output () | 133 (32) | 132 (33) | 0.95 |
| Power output () at VT1 | 76 (15) | 76 (20) | 0.87 |
| Medications | SL-ETGRP (n: 17) | DL-ETGRP (n: 16) |
|---|---|---|
| Immunosuppressant | 17 (100%) | 16 (100%) |
| Corticosteroids | 7 (41%) | 4 (25%) |
| NSAID | 10 (59%) | 9 (56%) |
| ACE-inhibitors | 2 (12%) | 1 (6%) |
| Angiotensin 2 receptor blockers | 4 (23%) | 1 (6%) |
| α-blockers | 6 (35%) | 4 (25%) |
| β-blockers | 11 (65%) | 4 (25%) |
| Diuretics | 3 (18%) | 2 (13%) |
| Calcium channel blockers | 4 (23%) | 3 (19%) |
| Statins | 6 (35%) | 3 (19%) |
| Lipid lowering agents | 2 (12%) | 0 (0%) |
| Metformin | 1 (6%) | 1 (6%) |
| Insulin | 1 (6%) | 0 (0%) |
| Thyroid hormones | 1 (6%) | 4 (25%) |
| Proton pump inhibitors | 10 (59%) | 6 (38%) |
| Xanthine oxidase inhibitors | 3 (18%) | 4 (25%) |
| Hypouricemic agents | 7 (41%) | 5 (31%) |
| Kinase inhibitor agents | 1 (6%) | 2 (13%) |
| Bisphosphonates | 1 (6%) | 1 (6%) |
| Dopamine agonists | 1 (6%) | 0 (0%) |
| Bronchodilators | 1 (6%) | 0 (0%) |
| Antigout agents | 1 (6%) | 1 (6%) |
| Antiarrhythmic agents | 1 (6%) | 0 (0%) |
| SL-ETGRP (n = 14) | DL-ETGRP (n = 13) | Effect Size | *** p Values | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| O2p Kinetics | PRE | POST | Mean Difference (95% CI) | PRE | POST | Mean Difference (95% CI) | ηp2 | G | T | G × T |
| O2 Def (mL O2) | 728 (168) | 596 (131) † | 132 (31; 233) | 734 (278) | 537 (204) † | 197 (101; 293) | 0.54 | 0.707 | <0.0001 | 0.276 |
| MRT (s) | 52.1 (15.9) | 43.5 (15.2) † | 8.5 (3 to 14) | 53.3 (14.4) | 38.6 (9.5) † | 14.6 (9; 20) | 0.63 | 0.707 | <0.0001 | 0.083 |
| SCamp (mL O2) | 207 (57) | 131 (68) † | 75.5 (27; 124) | 207 (84) | 118 (90) † | 90 (40; 138) | 0.51 | 0.767 | <0.0001 | 0.645 |
| HR kinetics | ||||||||||
| Baseline | 71.6 (11.6) ▲ | 70.3 (10.3) | 1.3 (−4.2; 6.8) | 81.2 (10.7) | 78.3 (12.3) | 3 (−2.3; 8.3) | 0.07 | 0.040 | 0.193 | 0.615 |
| Amplitude | 21.6 (6.9) ▲ | 19.6 (6.6) | 2 (−1; 5) | 26.4 (6) | 22.9 (5.5) † | 3.5 (0.7; 6.4) | 0.29 | 0.082 | 0.038 | 0.375 |
| Time delay (s) | 13.4 (10.7) | 17.8 (19.5) | −4.4 (−12.3; 3.5) | 11.9 (13.7) | 12.3 (13.8) | −0.4 (−8.2; 7.3) | 0.04 | 0.510 | 0.312 | 0.403 |
| Time constant (s) | 78.4 (79.9) | 50.6 (59) † | 27.7 (11.8; 43.7) | 62.9 (36.4) | 41.9 (28.7) † | 21 (5.7; 36.2) | 0.53 | 0.557 | <0.0001 | 0.468 |
| MRT (s) | 91.7 (87) | 68.4 (65.3) † | 23.3 (8.2; 38.4) | 74.8 (48.1) | 54.2 (39.2) † | 20.5 (6.2; 34.9) | 0.51 | 0.513 | <0.0001 | 0.655 |
| 95% CI for time constant | 58–81 | 39–55 | - | 52–65 | 37–48 | - | - | - | - | |
| DL-MOD | SL-ETGRP (n = 14) | DL-ETGRP (n = 15) | Effect Size | *** p Values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Steady-State Parameters | PRE | POST | Mean Difference (95% CI) | PRE | POST | Mean Difference (95% CI) | ηp2 | G | T | G × T |
| O2p-ss (mL·min−1) | 1206 (173) | 1180 (169) | 25 (−11; 61) | 1135 (226) | 1131 (229) | 4 (−30; 40) | 0.07 | 0.424 | 0.169 | 0.328 |
| CO2p-ss (mL·min−1) | 1153 (154) | 1090 (141) † | 62 (20 to 104) | 1060 (197) | 1019 (201) † | 41 (1 to 81) | 0.40 | 0.215 | 0.0002 | 0.398 |
| E-ss (L·min−1) | 41.8 (4.4) | 39.8 (4.9) # | 2 (−0.1 to 4.1) | 38.5 (8.7) | 36.1 (6.9) † | 2.4 (0.4; 4.4) | 0.32 | 0.154 | 0.001 | 0.734 |
| RER | 0.96 (0.04) | 0.93 (0.04) † | 0.03 (0.01; 0.06) | 0.94 (0.04) | 0.90 (0.03) † | 0.03 (0.01; 0.06) | 0.40 | 0.080 | 0.0002 | 0.873 |
| Gross Efficiency (%) | 14.7 (1.2) | 15 (1.4) | −0.3 (−0.8 to 0.1) | 14.4 (2.3) | 14.5 (2.1) | 0.1 (−0.5 to 0.3) | 0.11 | 0.595 | 0.100 | 0.404 |
| O2 cost (mL·Watt−1) | 14.0 (1.1) | 13.9 (1) | 0.1 (−0. to 0.7) | 14.4 (3.2) | 14.4 (2.4) | 0 (−0.6 to 0.6) | 0.00 | 0.524 | 0.727 | 0.807 |
| HRss (bpm) | 97 (17) ⁑ | 91 (15) † | 5 (1 to 9) | 111 (12) | 102 (12) † | 9 (5; 13) | 0.58 | 0.022 | <0.0001 | 0.094 |
| [La]b (mmol·L−1) | 3.15 (1.11) | 2.45 (1.15) † | 0.7 (0.3 to 1.1) | 3.18 (0.84) | 1.98 (0.75) † | 1.15 (0.8; 1.5) | 0.76 | 0.689 | <0.0001 | 0.057 |
| Power () | 62 (13) | 62 (13) | - | 58 (14) | 58 (14) | - | - | - | - | - |
| DL-HVY | SL-ETGRP (n = 17) | DL-ETGRP (n = 16) | Effect Size | p Values *** | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| End-Exercise Parameters | PRE | POST | Mean Difference (95% CI) | PRE | POST | Mean Difference (95% CI) | ηp2 | G | T | G × T |
| O2p (mL·min−1) | 1712 (293) | 1636 (249) † | 76 (15; 137) | 1665 (344) | 1563 (351) † | 103 (40; 166) | 0.43 | 0.579 | <0.0001 | 0.469 |
| CO2p (mL·min−1) | 1651 (263) | 1554 (238) † | 97 (37; 158) | 1645 (334) | 1478 (329) † | 167(104; 229) | 0.62 | 0.689 | <0.0001 | 0.069 |
| RER | 0.97 (0.04) | 0.95 (0.03) # | 0.02 (0; 0.04) | 0.99 (0.06) | 0.95 (0.04) † | 0.04 (0.02; 0.07) | 0.40 | 0.540 | <0.0001 | 0.067 |
| O2 cost (mL·Watt−1) | 14.2 (1.2) | 13.6 (1.4) | 0.6 (−0.2; 1.4) | 14.8 (2.9) | 13.5 (1.7) † | 1.3 (0.5; 2.1) | 0.33 | 0.635 | <0.0005 | 0.158 |
| E (L·min−1) | 70.3 (13.3) | 62.8 (10.2) † | 7.4 (2.9; 12) | 68.0 (13.8) | 56.5 (13.3) † | 11.5 (6.8; 16.2) | 0.60 | 0.313 | <0.0001 | 0.158 |
| O2-RM | 116 (27) | 104 (24) † | 12 (1.5; 22.7) | 113 (29) | 93 (24) † | 20 (9; 31) | 0.44 | 0.394 | <0.0001 | 0.230 |
| HR (bpm) | 126 (24) | 118 (22) † | 8 (3; 13) | 141 (18) | 128 (19) † | 13 (8; 18) | 0.60 | 0.098 | <0.0001 | 0.137 |
| [La]b (mmol·L−1) | 6.2 | 5.2 † | 1 (0.1; 2) | 6.6 | 4.6 † | 2 (1.1; 2.9) | 0.52 | 0.881 | <0.0001 | 0.088 |
| RPEdyspena | 15.1 (1.5) | 13.6 (1.6) † | 1.5 (0.1; 2.9) | 16.2 (1.6) | 14.1 (1.8) † | 2.1 (0.7; 3.4) | 0.40 | 0.072 | 0.0001 | 0.523 |
| RPEleg pain | 6.2 (1.6) | 5.1 (1.7) † | 1.1 (0.1; 2.2) | 6.3 (1.4) | 5.1 (1.9) † | 1.2 (0.1; 2.2) | 0.32 | 0.964 | 0.001 | 0.932 |
| Power () | 95 (24) | 95 (24) | - | 94 (25) | 94 (25) | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
del Torto, A.; Capelli, C.; Peressutti, R.; Di Silvestre, A.; Livi, U.; Nalli, C.; Sponga, S.; Amici, G.; Baccarani, U.; Lazzer, S.
The Effect of Endurance Training on Pulmonary
del Torto A, Capelli C, Peressutti R, Di Silvestre A, Livi U, Nalli C, Sponga S, Amici G, Baccarani U, Lazzer S.
The Effect of Endurance Training on Pulmonary
del Torto, Alessio, Carlo Capelli, Roberto Peressutti, Adriana Di Silvestre, Ugolino Livi, Chiara Nalli, Sandro Sponga, Giampaolo Amici, Umberto Baccarani, and Stefano Lazzer.
2022. "The Effect of Endurance Training on Pulmonary
del Torto, A., Capelli, C., Peressutti, R., Di Silvestre, A., Livi, U., Nalli, C., Sponga, S., Amici, G., Baccarani, U., & Lazzer, S.
(2022). The Effect of Endurance Training on Pulmonary