1. Introduction
Phytoplankton are the main marine primary producer. As the basic link in the marine food web structure [
1], phytoplankton play an important role in the material cycle and energy transformation of marine ecosystems [
2]. A change in the phytoplankton community directly affects the succession of food webs in ecosystems, particularly the characteristics of the top consumer community [
3], and the species composition, and community structure of phytoplankton are often used to characterize the water environment [
4].
The community characteristics, distribution, and quantitative dynamics of dominant phytoplankton species are closely related to a variety of environmental factors [
5]. Temperature is an important factor that affects the photosynthesis and respiration of phytoplankton, and it is closely related to the biological efficiency of phytoplankton [
6,
7,
8]. An increase in temperature is related to changes in the standing stock of biomass of primary producers [
7]. Salinity affects phytoplankton community composition and cell abundance [
9]. For example, diatoms are usually dominant in low-salinity seawater, whereas dinoflagellates and cyanobacteria are more tolerant to high-salinity seawater. When seawater salinity exceeds 30 psu, the biomass of phytoplankton becomes negatively correlated with salinity [
10]. In addition, light, water mass, current, monsoon conditions, and climate also affect the biomass, community structure, and distribution of phytoplankton [
11,
12,
13].
At present, seawater warming and eutrophication are the two major environmental problems encountered by offshore ecosystems [
14]. Diatoms and dinoflagellates are the two dominant phytoplankton groups [
15] in offshore ecosystems, which cause the highest frequency of offshore algal blooms. Therefore, an analysis of the response of phytoplankton communities, especially diatoms and dinoflagellates, to seawater warming and eutrophication is very important to understand the impact of future environmental changes on the structure and function of offshore ecosystems [
16]. Over the last 20 years, related studies have focused mainly on the quantitative dynamic relationship between nutrient input and phytoplankton, the evolutionary trends of dominant species and community structure of offshore phytoplankton under eutrophication [
17,
18,
19], and the interaction of various stress factors to explore the effects of multiple factors on phytoplankton productivity and community structure [
20]. The aims of this study were: (1) to investigate the response characteristics of phytoplankton species to environmental stressors; (2) to determine how these response characteristics are affected by these stressors, and (3) to assess the potential tradeoffs that may be involved, which could help to improve our conceptual understanding of the effects of multiple stressors on different phytoplankton populations [
1]. Therefore, long-term monitoring of phytoplankton is crucial to understanding the coastal ecological environment and identifying the key environmental factors which affect it.
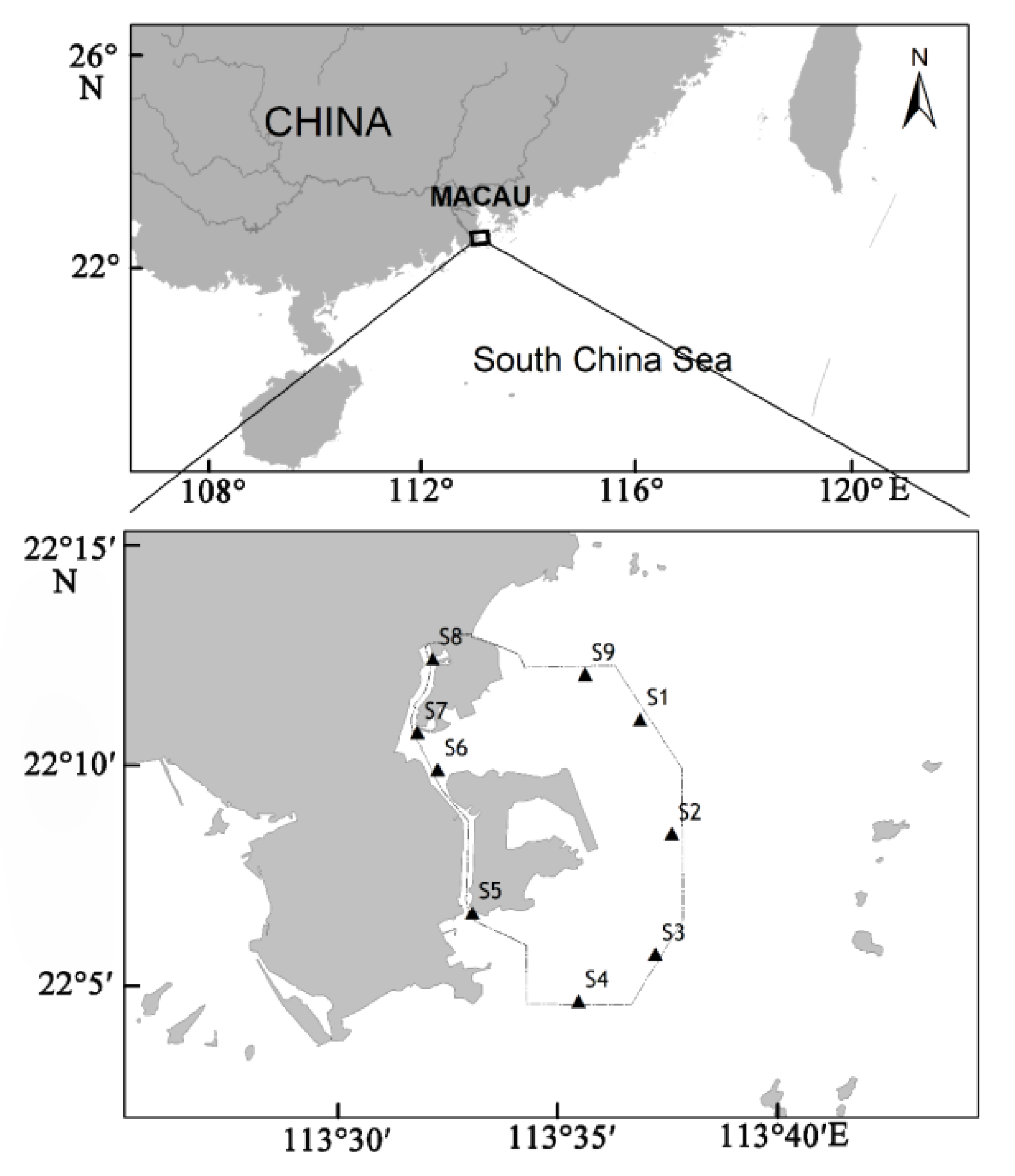
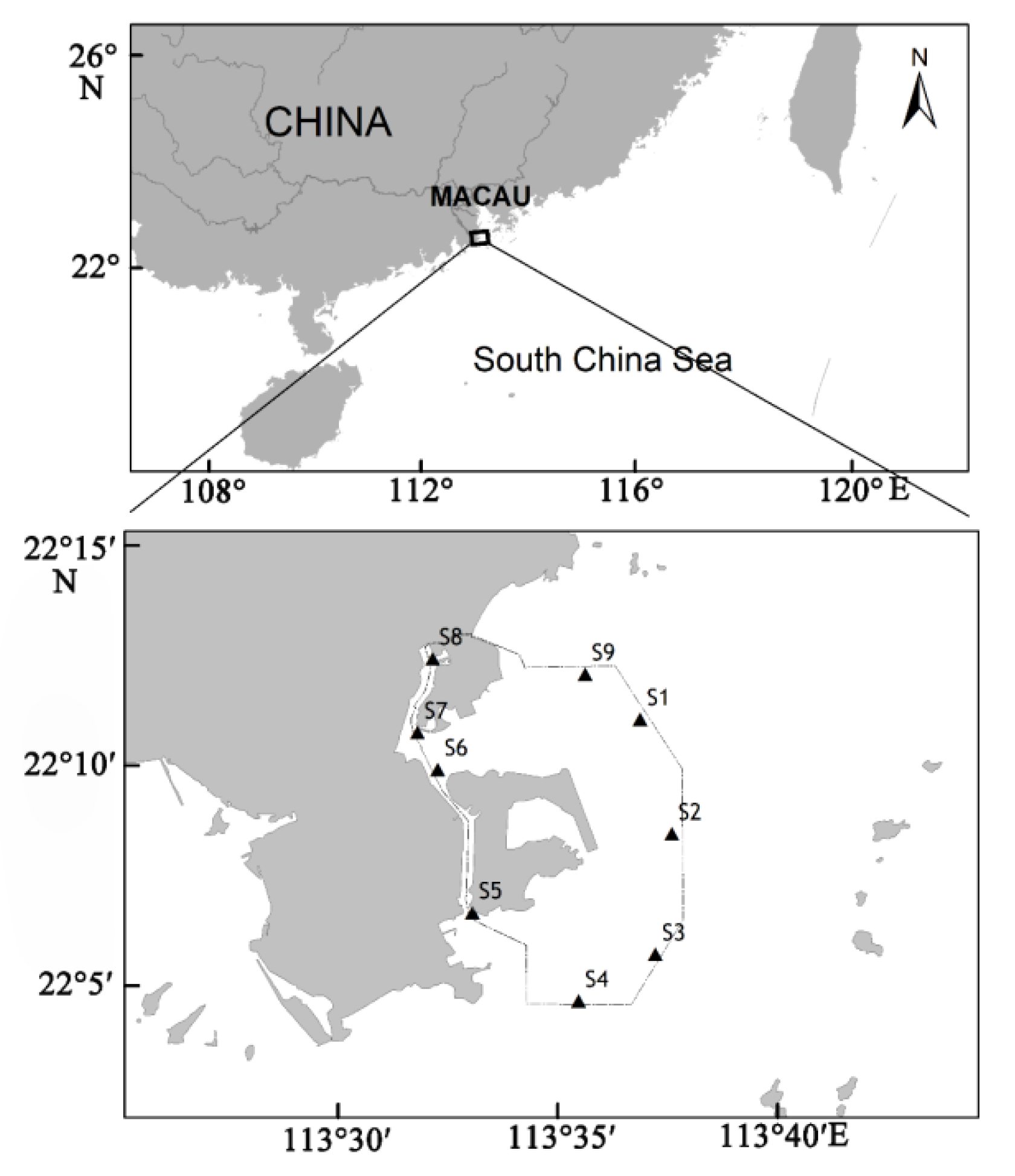
The waters around Macau are located in the channel between Lingdingyang Bay and Modaomen watercourse in the Pearl River Estuary and represent an important component of the Pearl River Estuary. Macau is one of the four central cities of the Guangdong–Hong Kong–Macau greater bay area. It has a population density of more than 20,000 inhabitants/km2, which makes it one of the most densely populated cities in the world, with a per capita GDP of about US$80,000, dominating the top of the ranking among the most economically developed regions in the world. With economic development and the discharge of pollutants such as domestic sewage and industrial wastewater, the ecological environment of Macau’s surrounding waters has been seriously damaged. According to a report on the environmental condition of Macau (2018), in the past 10 years, eutrophication along the coast of Macau has become serious, and masses of dead fish, black and odorous waters, and red tides frequently occur.
In the last decade, numerous studies have been performed on the ecology of phytoplankton in the Pearl River Estuary and its adjacent waters in China, and these have discussed the ecological characteristics of phytoplankton, mechanism of red tides, and grain size structure and spatial distribution of primary productivity [
21,
22,
23,
24,
25,
26,
27,
28,
29]. However, there is still a lack of research on the community structure of phytoplankton and the relationship between phytoplankton and environmental factors in the waters around Macau.
In the present study, based on the survey data of phytoplankton in the waters around Macau, the phytoplankton community was analyzed by a multivariate statistical method, and the relationship between the phytoplankton community and environmental factors was evaluated. The primary objective of this study was to provide a reference to protect the biodiversity and resources in the waters around Macau and to provide basic data for the economic construction and offshore ecosystem management in the Guangdong–Hong Kong–Macau greater bay area.
4. Discussion
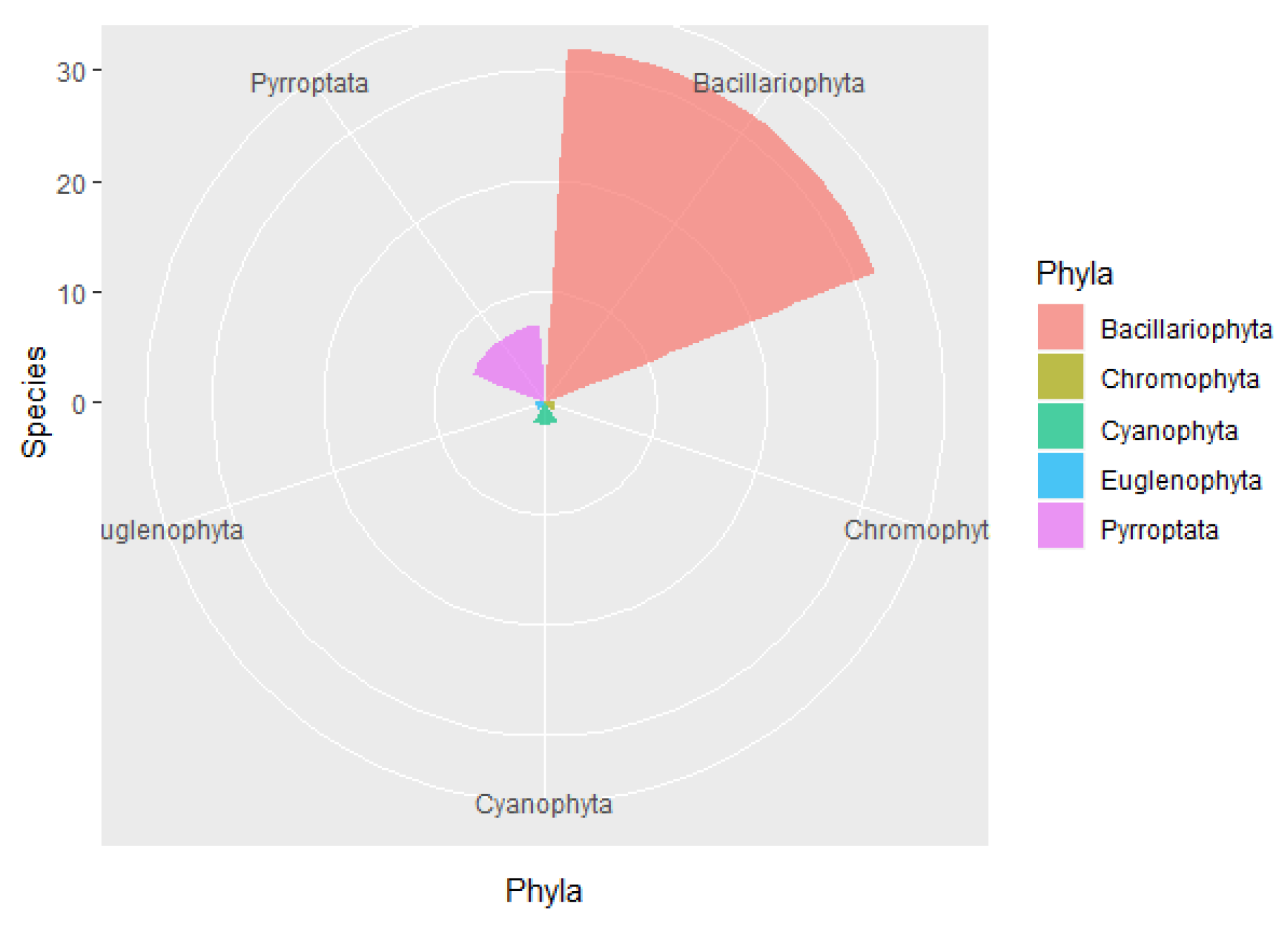
The results of the present study indicated that diatoms are the major phytoplankton species in the waters around Macau, which is consistent with data from the inshore waters of the northern South China Sea [
22,
23,
24,
35,
36]. The phytoplankton species that can thrive under normal and warm temperature conditions were dominant and showed obvious tropical and subtropical characteristics [
22,
23]. The surveyed waters are affected by fresh water and tidal currents. The phytoplankton species could be divided into different ecological types, such as low-salt nearshore, widespread offshore, and high-salt offshore, and this classification is consistent with that reported by Jia et al. (2019) [
21]. For example,
S. costatum is a nearshore low-salt phytoplankton species, whereas
Rhizosolenia setigera Brightwell,
T. nitzschioides, Ditylum brightwellii, T. frauenfeidii, etc., are widespread offshore phytoplankton species with a wide range of adaptations to temperature and salinity.
Rhizosolenia styliformis Brightwell and
C. siamense Ostenfeld are species that can adapt to high salinities and varying temperatures and could have been transported to the study area from the open sea by tidal currents. The phytoplankton taxa predominantly detected in the study area belonged to the widely distributed offshore group, and these results are consistent with those reported in previous studies [
28,
37].
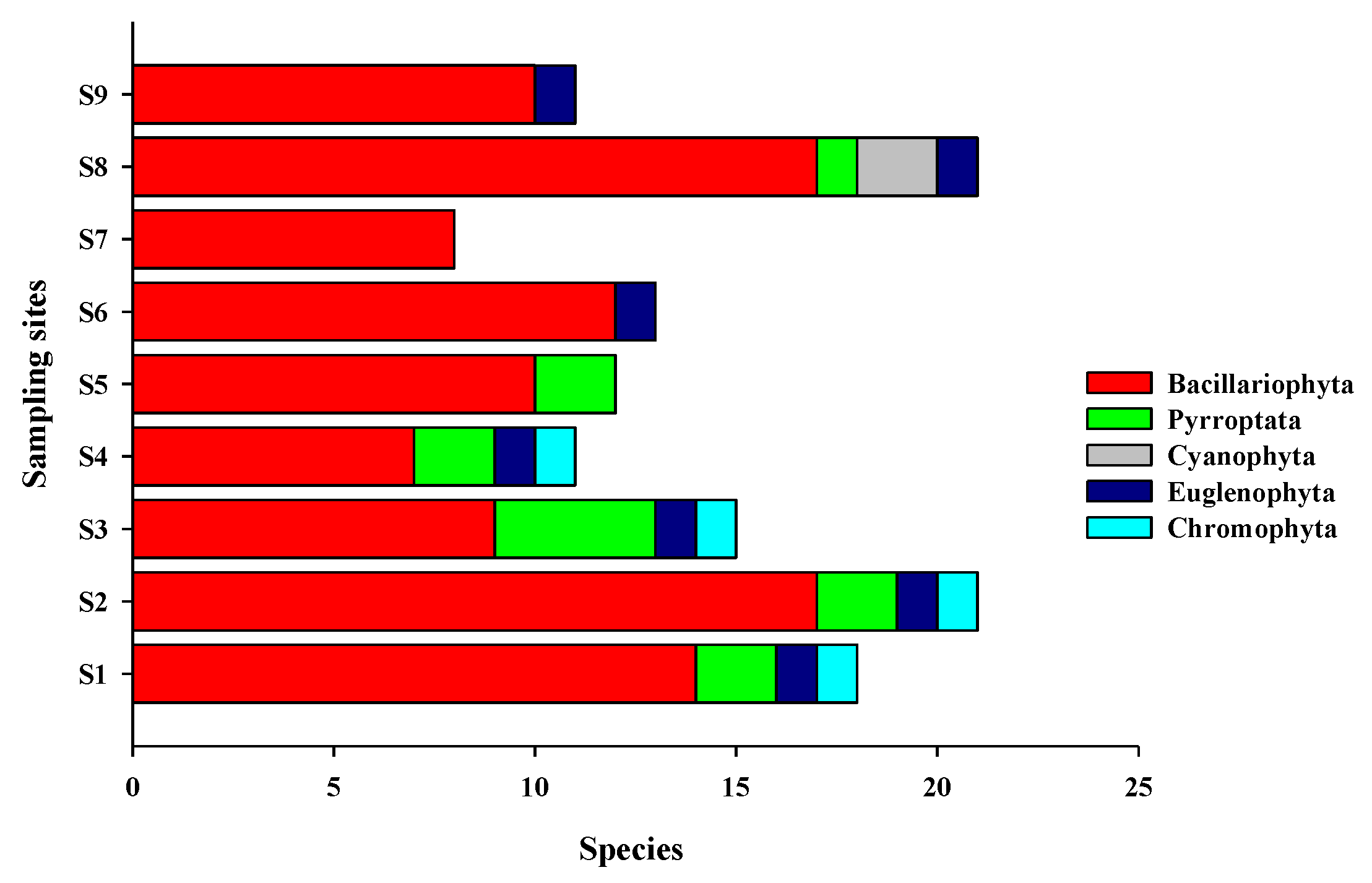
The dominant phytoplankton species in the waters around Macau were
S. costatum,
A. granulata, T. frauenfeidii, and
T. nitzschioides, similar to those identified in previous studies [
28,
38,
39].
S. costatum is a red tide alga that is abundant in the coastal waters of Guangdong [
40]. Between the years 2000 and 2009, 36 red tides were recorded in the waters near the Pearl River Estuary. Diatoms, mainly
S. costatum, are the predominant red tide algae [
41,
42], especially in the high water season [
23], and numerous studies have reported similar findings in other waters, such as the East China Sea [
43]. While
S. costatum does not produce phycotoxins, its major effect on the marine ecosystem is oxygen depletion caused by the appearance of red tides [
44].
A. granulata is a universal phytoplankton species, which is particularly dominant in summer and in medium-eutrophic water. It is often used as an indicator of eutrophication and polluted water [
45,
46].
T. frauenfeidii is a widespread phytoplankton species, which can also cause red tides and is one of the dominant phytoplankton species in the coastal waters, bays, and estuaries of China [
47]. As a common diatom found worldwide,
T. nitzschioides is distributed in all major seas and shallow seas from the equator to high latitudes, except in the southern and arctic regions [
48]. Furthermore,
T. nitzschioides has been reported to be a good indicator of change in seawater temperature [
49].
In marine phytoplankton studies, the diversity index is a commonly used biodiversity measurement tool that can better reflect the attributes of the phytoplankton community and its relationship with the environment [
50]. The results of the present study showed that the spatial distribution of phytoplankton biodiversity in the waters around Macau is much lower than that in the open sea, suggesting that the phytoplankton community in the waterway area is significantly disturbed. Frequent interference by human activities (such as shipping and flood discharge) and eutrophication are the main factors that affect the diversity of phytoplankton communities in waterways. The results of the present study are consistent with the phytoplankton biodiversity observed in the adjacent waters of the Pearl River Estuary [
22,
23,
24,
28,
39]. Correlation analysis between the diversity index and environmental factors revealed a significant negative correlation between phytoplankton abundance and turbidity, and a significant positive correlation between phytoplankton biodiversity and nitrogen nutrients in the water. The instability of the water body in the waterway area is influenced by multiple stresses, such as waterway transportation and drainage, which affects the growth of phytoplankton on the surface of the water. Furthermore, land source discharge and shipping disturbance increase the turbidity of the water body, which affects the absorption of light by plants. The light restriction also decreases phytoplankton abundance. The differences in nutrient levels may potentially limit the absorption and utilization of nutrients by phytoplankton and regulate the phytoplankton population structure. While the growth of some phytoplankton species could be restricted by the lack of certain nutrients, other phytoplankton species with relatively low demand for these nutrients can rapidly grow and become dominant, resulting in simplification of the species composition and a decrease in biodiversity. In the present study, the low Shannon–Wiener index and moderate species richness reflected the uneven distribution of the phytoplankton species, with diatoms dominating the phytoplankton community in the study sites.
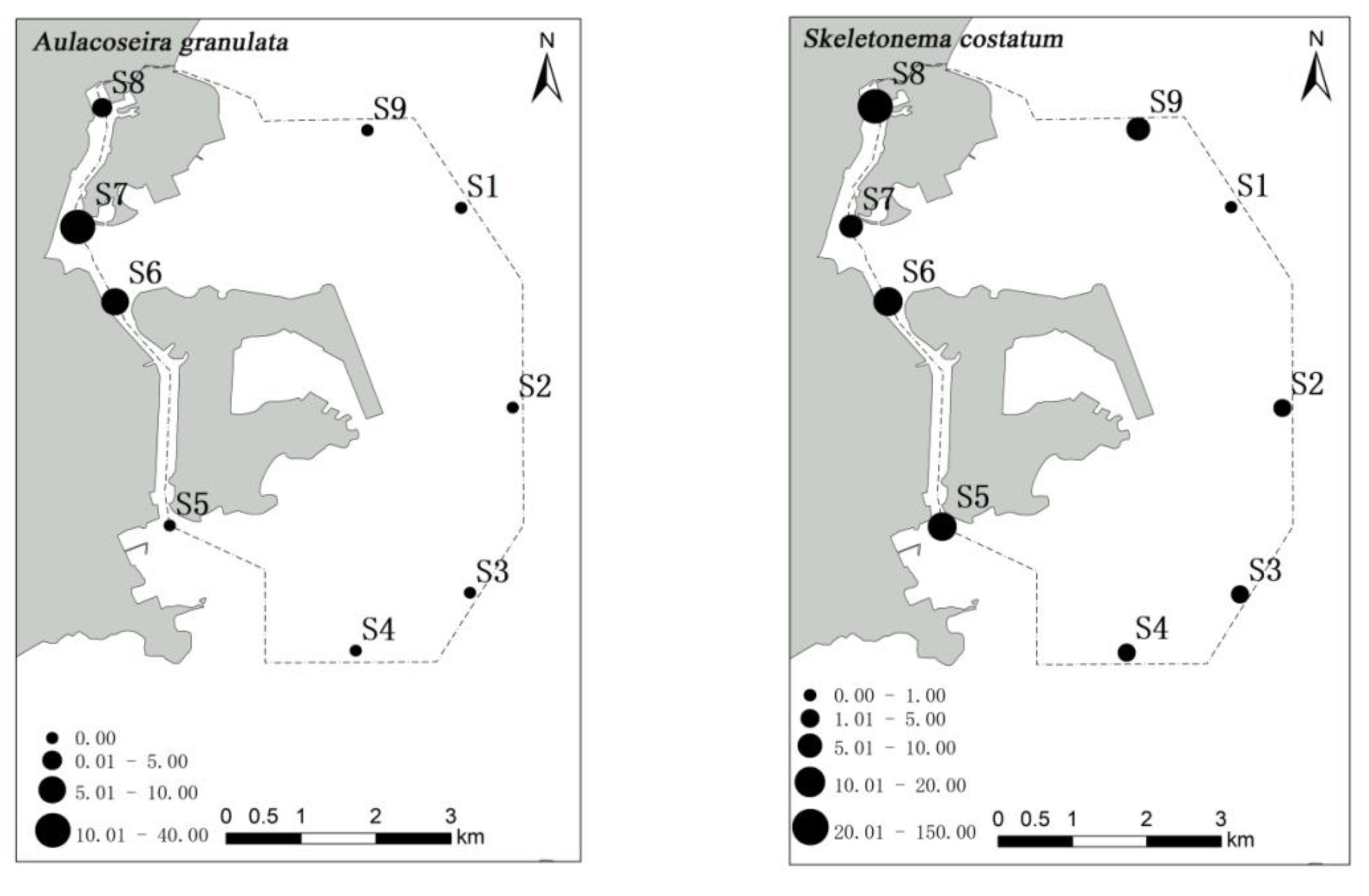
Spatial changes in phytoplankton communities are an important component of many aspects of ecology, including the maintenance of species diversity and community stability [
51,
52]. The spatial distribution of phytoplankton communities is mainly caused by the heterogeneity of the environment on the spatial scale [
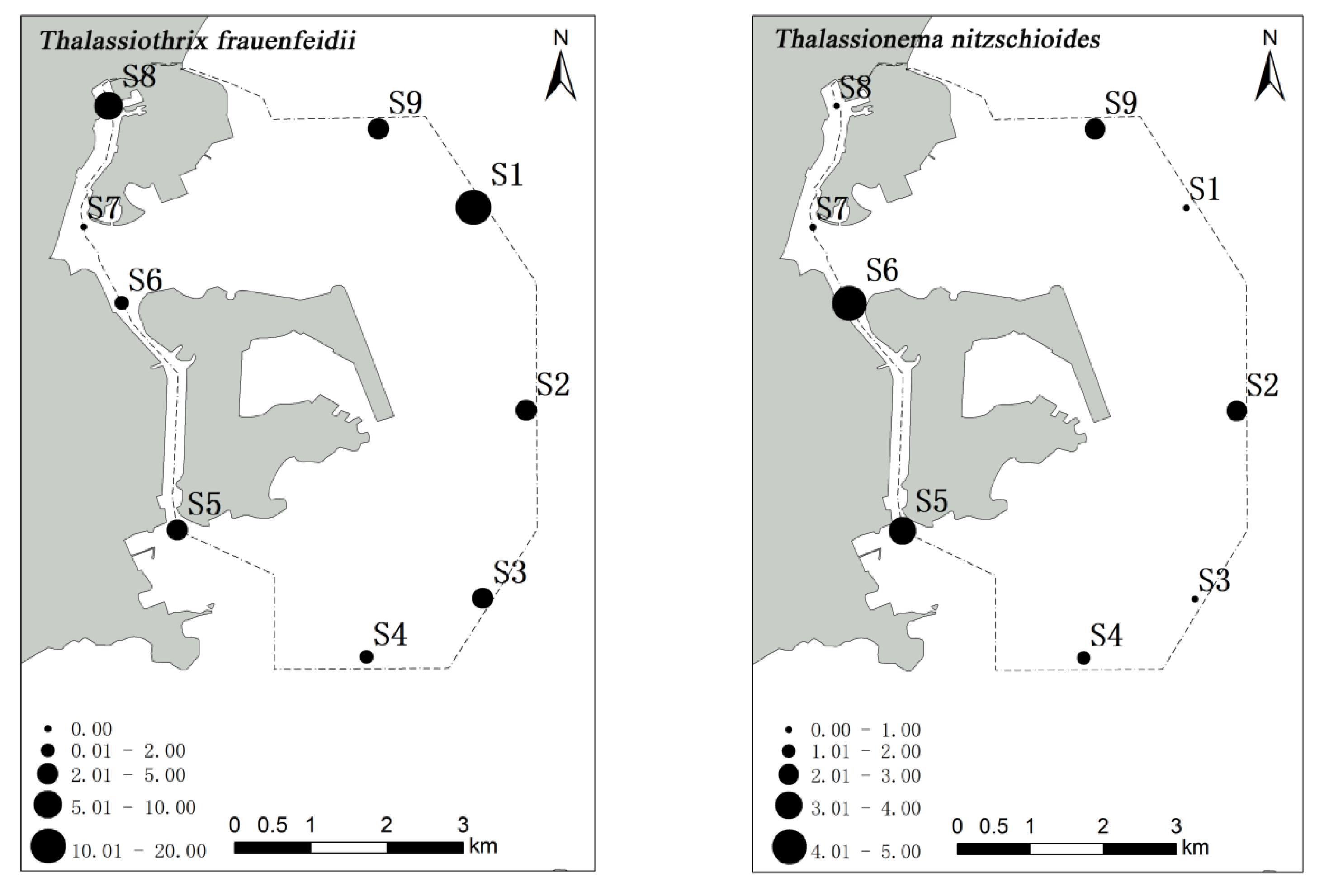
53]. In the present study, a high abundance of phytoplankton was observed in stations S7 and S8 of the Qianshan waterway and station S6 of the Shizimen waterway. The abundance of
S. costatum in stations S8 and S7 reached 1380 and 289 cells/m
3, respectively. The distribution of
S. costatum is negatively correlated with salinity and transparency [
54]. In the waters around Macau, the inflow of fresh water from the Pearl River decreases the salinity and transparency, making it suitable for the growth of
S. costatum. Similar results have also been noted in the Yangtze Estuary [
55]. Some studies have shown that
S. costatum can grow better in water with a high N/P ratio, such as the Pearl River Estuary. Some studies have shown that eutrophic water bodies are more suitable for
A. granulata growth, and that the mixed stirring environment is more conducive to maintaining
A. granulata in the true light layer [
56]. In the present study,
A. granulata was distributed in stations S6 and S8, but its abundance was much lower than that observed in S7. Furthermore, a high number of
T. frauenfeidii (221 cells/m
3) was observed in station S1, which may be related to the temperature, water depth, salinity, or nutrient concentration [
57,
58].
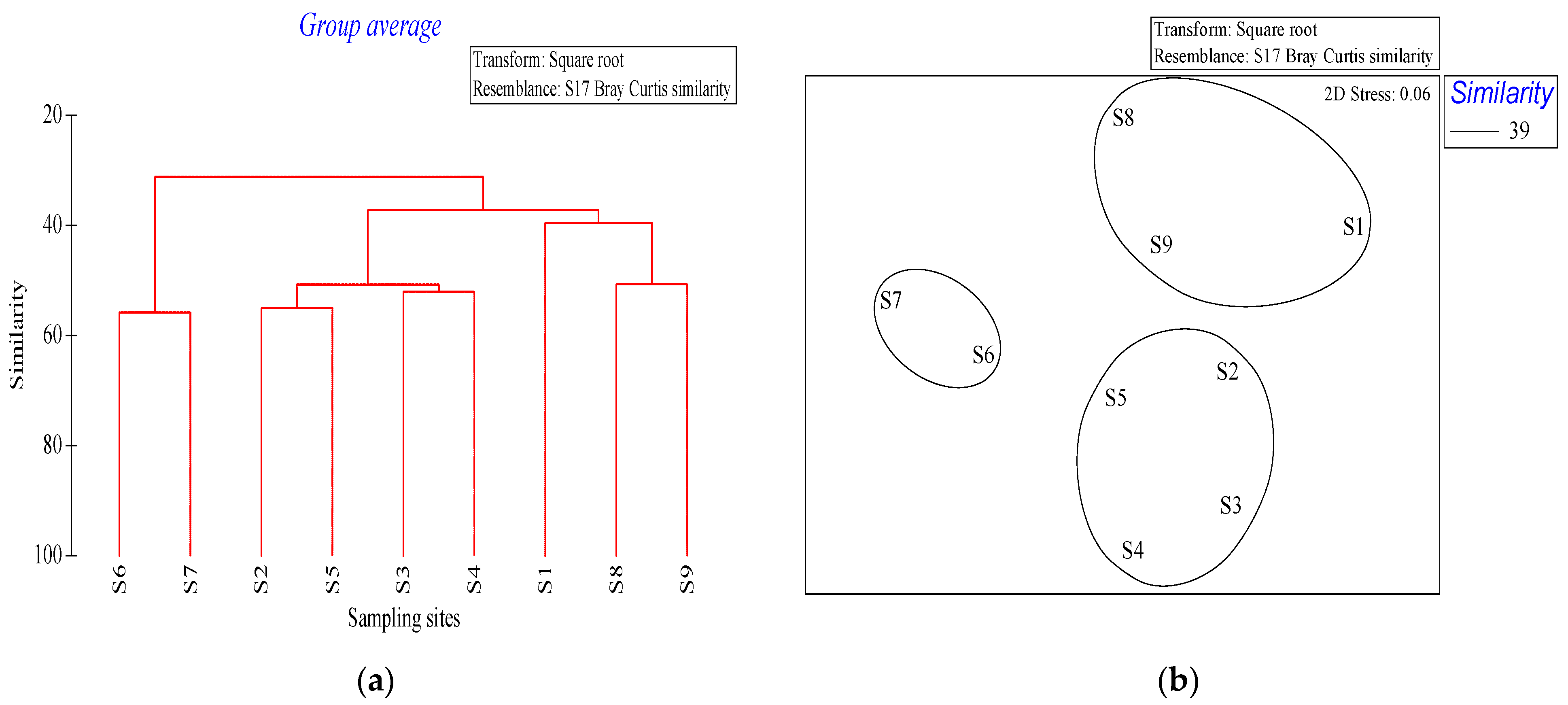
The NMDS results revealed that the phytoplankton communities in the waters around Macau could be mainly divided into the
A. granulata—
S. costatum,
S. costatum—T. frauenfeidii—T. nitzschioides, and
T. frauenfeidii—C. curvisetus—C. diadema—
S. costatum communities. The
A. granulata—S. costatum community was detected in the Qianshan waterway, which has a high nutrient content. The Qianshan waterway is affected by the freshwater inflow from the Pearl River, resulting in relatively low salinity, which promotes
S. costatum growth [
59]. The salinity of stations S6 and S7 was 5.14–7.6 psu, Huo et al. (2001) reported 10–20 psu was the optimal salinity range for
S. costatum growth [
60]. The
S. costatum—T. frauenfeidii—T. nitzschioides community was detected in the waters southeast of Macau, mainly owing to the ecological characteristics of these species.
Thalassiothrix frauenfeidii and
T. nitzschioides can tolerate a wide range of temperatures and salinities, and
T. nitzschioides populations arranged in chain. The
T. frauenfeidii—C. curvisetus—C. diadema—S. costatum community was detected in the northeast of the survey waters.
C. diadema is a coastal species distributed from the northern temperate zone to the Arctic region. It has adapted to a wide range of temperatures and salinities and is widely distributed around the world. In this study,
C.
diadema was detected in the offshore stations S1 and S9.
Chaetoceros curvisetus Cleve is a eurythermic coastal species, which presented the second highest cell abundance; however, its distribution range was small, indicating its narrow ecological adaptation range.
Phytoplankton must absorb inorganic nutrients (mainly nitrate, nitrite, silicate, and phosphate) from the water body for their own growth and reproduction. Therefore, the nutrient concentration in a water body is the primary factor affecting phytoplankton communities [
61]. In the northern part of the South China Sea, nutrient concentration is the primary limiting factor for phytoplankton growth. Owing to terrestrial nutrient supplementation, the Pearl River Estuary and coastal areas do not usually face nutrient limitations. As a result, phytoplankton cell abundance is higher in these areas, when compared with those in the shelf and open sea areas [
62]. Long et al. analyzed the surface water of the northern South China Sea and concluded that the growth of phytoplankton and nitrite content in the northern South China Sea are more closely related than other nutrient factors [
63]. Song et al. (2013) suggested that Hong Kong’s coastal waters are more likely to be restricted by N content in various seasons [
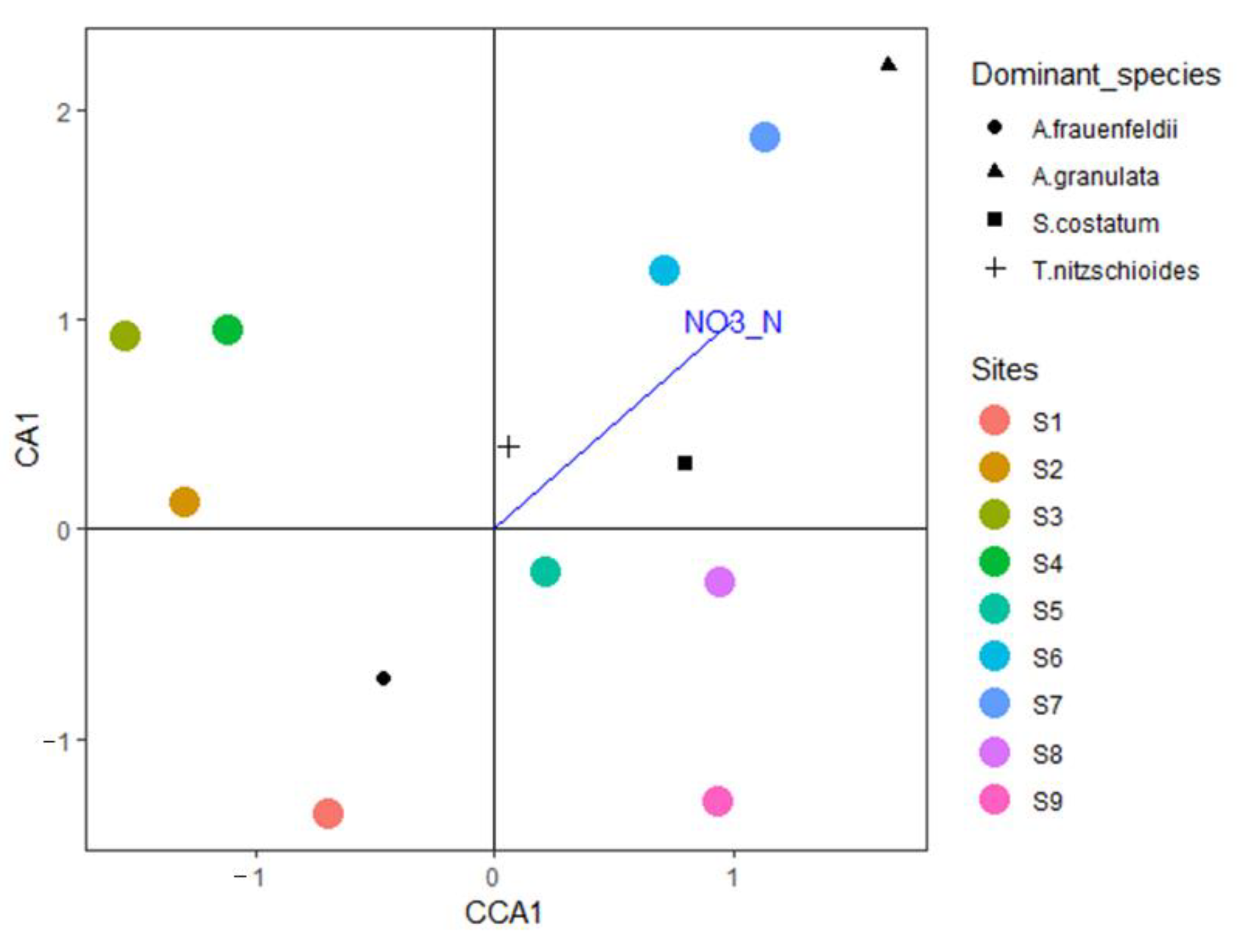
64]. Considering the minimum nutrient threshold, winter nutrients do not restrict the growth of phytoplankton. In the present study, the CCA of the phytoplankton community and environmental factors revealed a significant correlation between nitrite nitrogen and phytoplankton abundance. The CCA triplot indicated that
S. costatum,
A. granulata, and
T. nitzschioides growth, especially
S. costatum growth, is closely related to nitrite nitrogen content. Previous studies have shown that different N/P ratios in the environment affect the specific growth rate and cell state of
S. costatum, and that the N content restricts growth more than P content.
S. costatum growth is significantly correlated with total nitrogen and dissolved inorganic nitrogen contents in Baltic coastal waters [
65]; however, Lagus et al. indicated that
S. costatum growth was mainly restricted by P content.
A. granulata is known to adapt to low-salinity conditions [
66], and its distribution is mainly determined by the nutrient distribution pattern, as well as salinity [
67]. Furthermore, previous studies have demonstrated that the growth of
T. nitzschioides is mainly controlled by temperature and nutrient concentration.
T. nitzschioides is considered to be a good indicator of changes in the nutrient concentration in the South-North Sea [
68].
In coastal areas where river runoff injects a high level of nutrients, a continuous influx of ammonium-utilizing, nitrite-utilizing, and nitrifying microorganisms can cause competition and restriction in the phytoplankton population and maintain a temporary coexisting population of nitrifying bacteria [
69]. The coastal phytoplankton communities with a high degree of eutrophication tend to be of smaller size [
70,
71], and small diatoms (such as
Skeletonema sp. and
Chaetoceros sp.) may be well represented in areas with a high degree of eutrophication. Nevertheless, further studies are needed to perform long-term monitoring of phytoplankton communities to determine the response of phytoplankton to multiple stresses in the coastal waters of Macau.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}