
The Role of Grass Compost and Zea Mays in Alleviating Toxic Effects of Tetracycline on the Soil Bacteria Community




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Objective
2.2. Experiment Design
2.3. Methods of Laboratory Analyses
2.3.1. Microbiological Analyses
- Count of microorganisms
- DNA isolation
- Amplicon sequencing
- Bioinformatic analysis
2.3.2. Biochemical Analyses
2.3.3. Chemical and Physicochemical Analyses
2.4. Data and Statistical Analysis
3. Results
3.1. Response of Zea Mays to Soil Contamination with Tetracycline
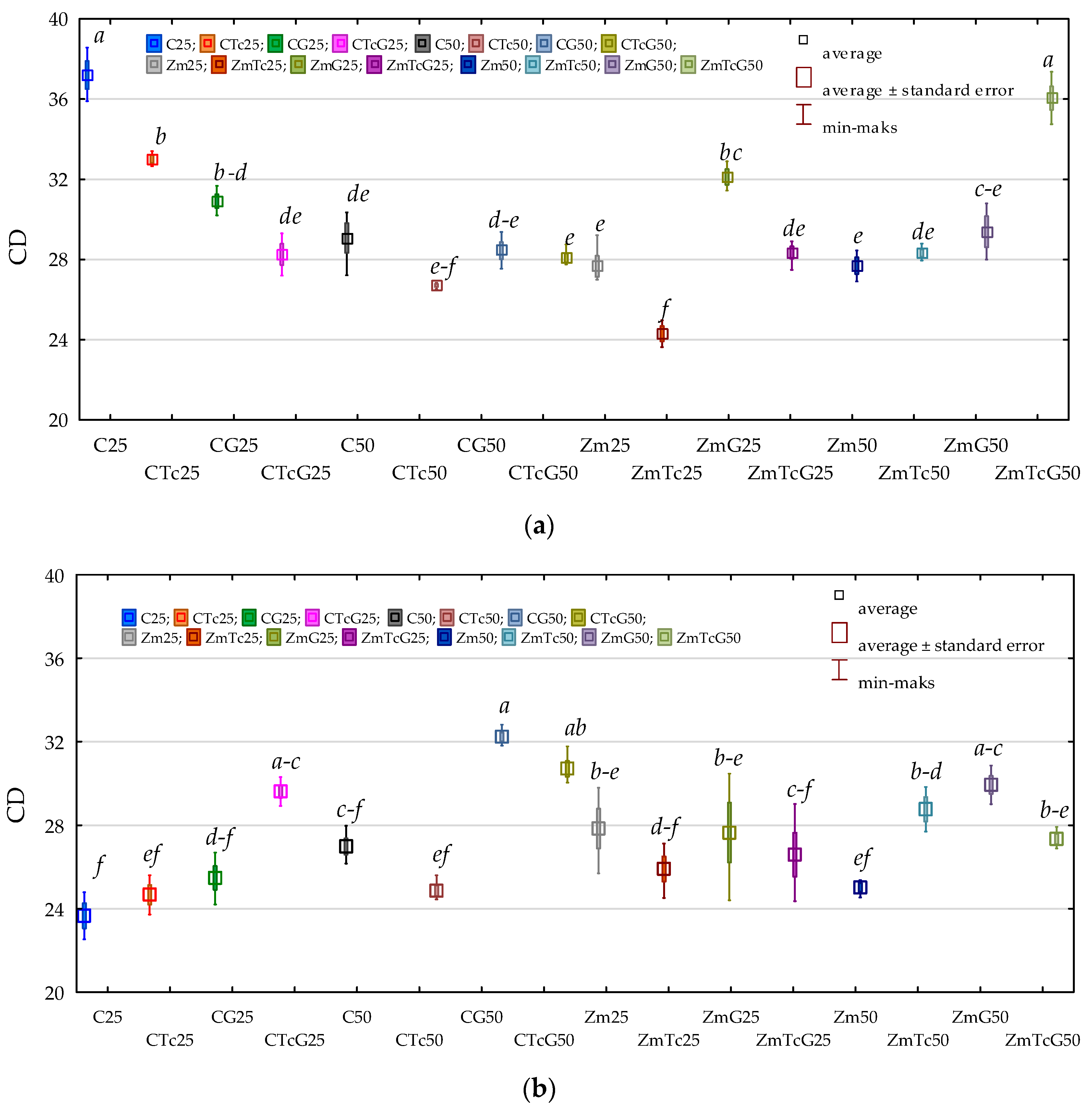
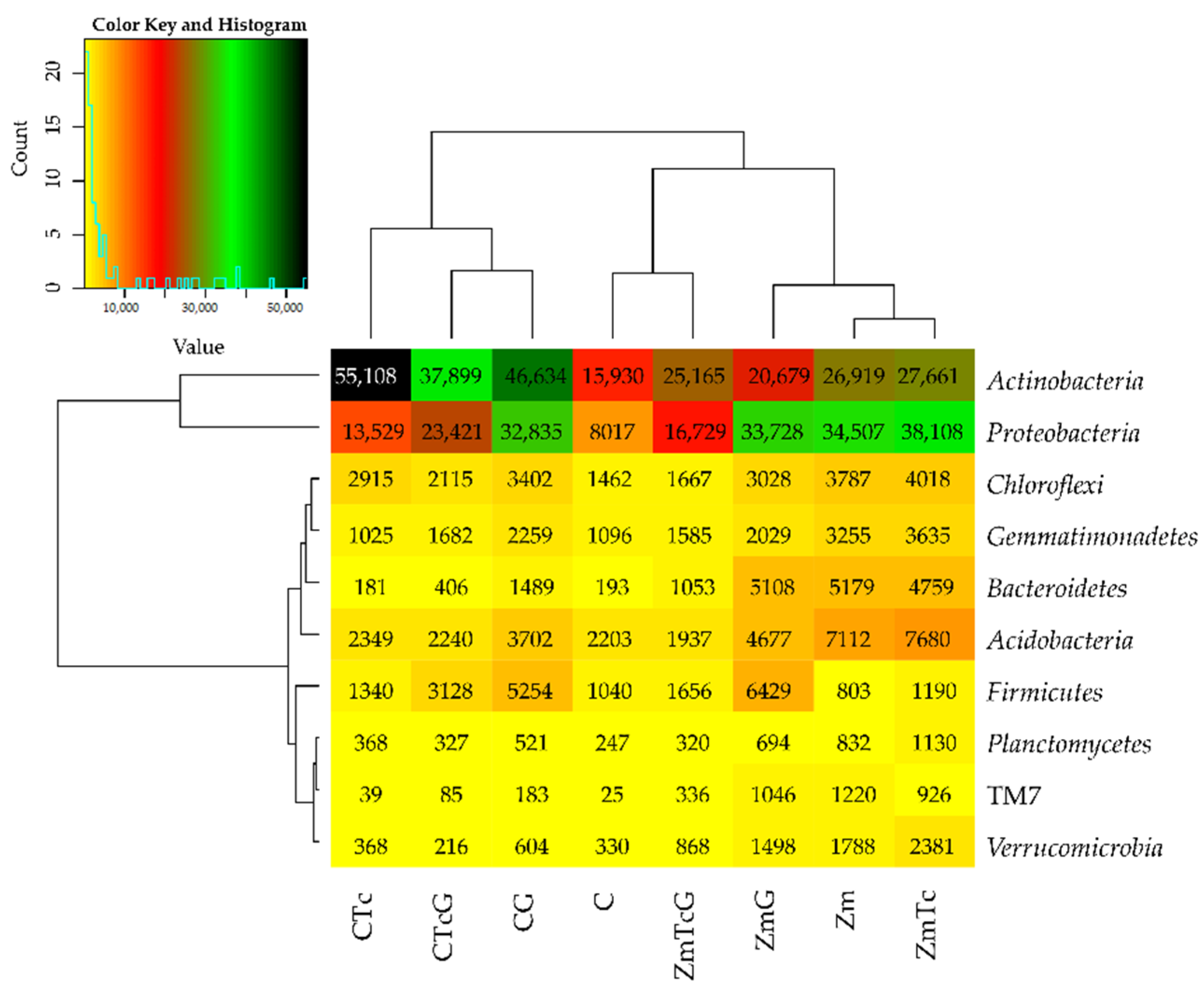
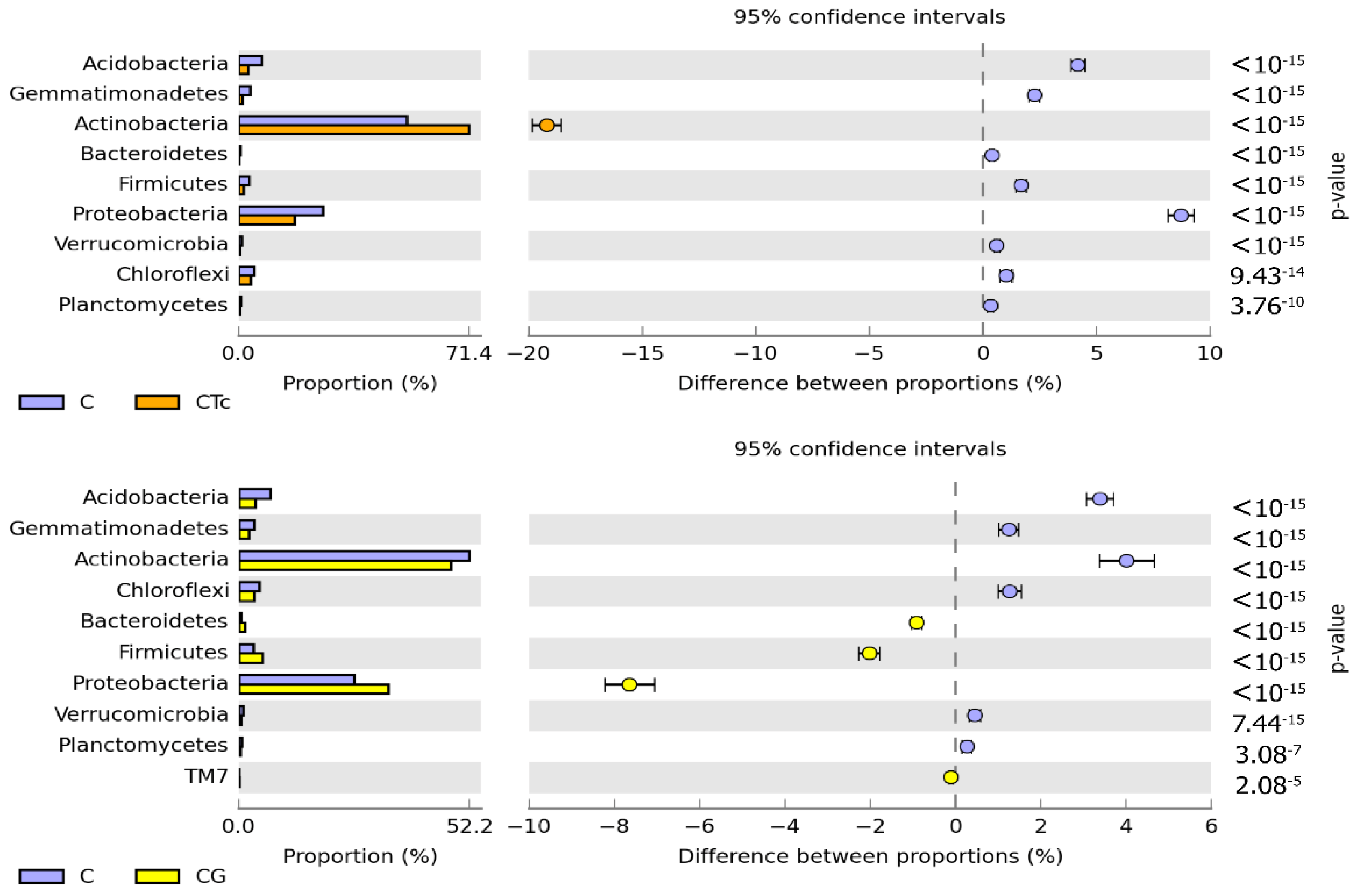
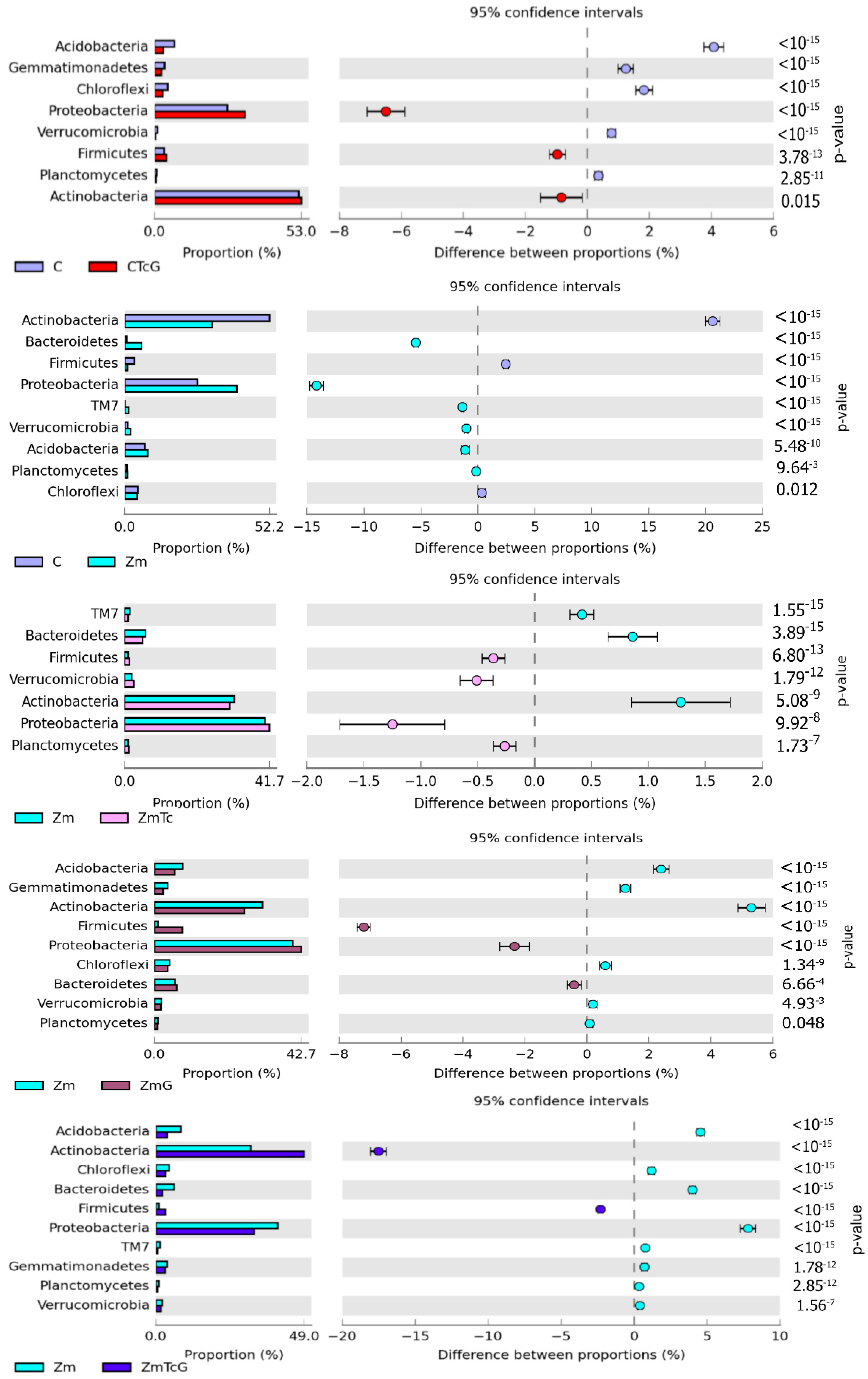
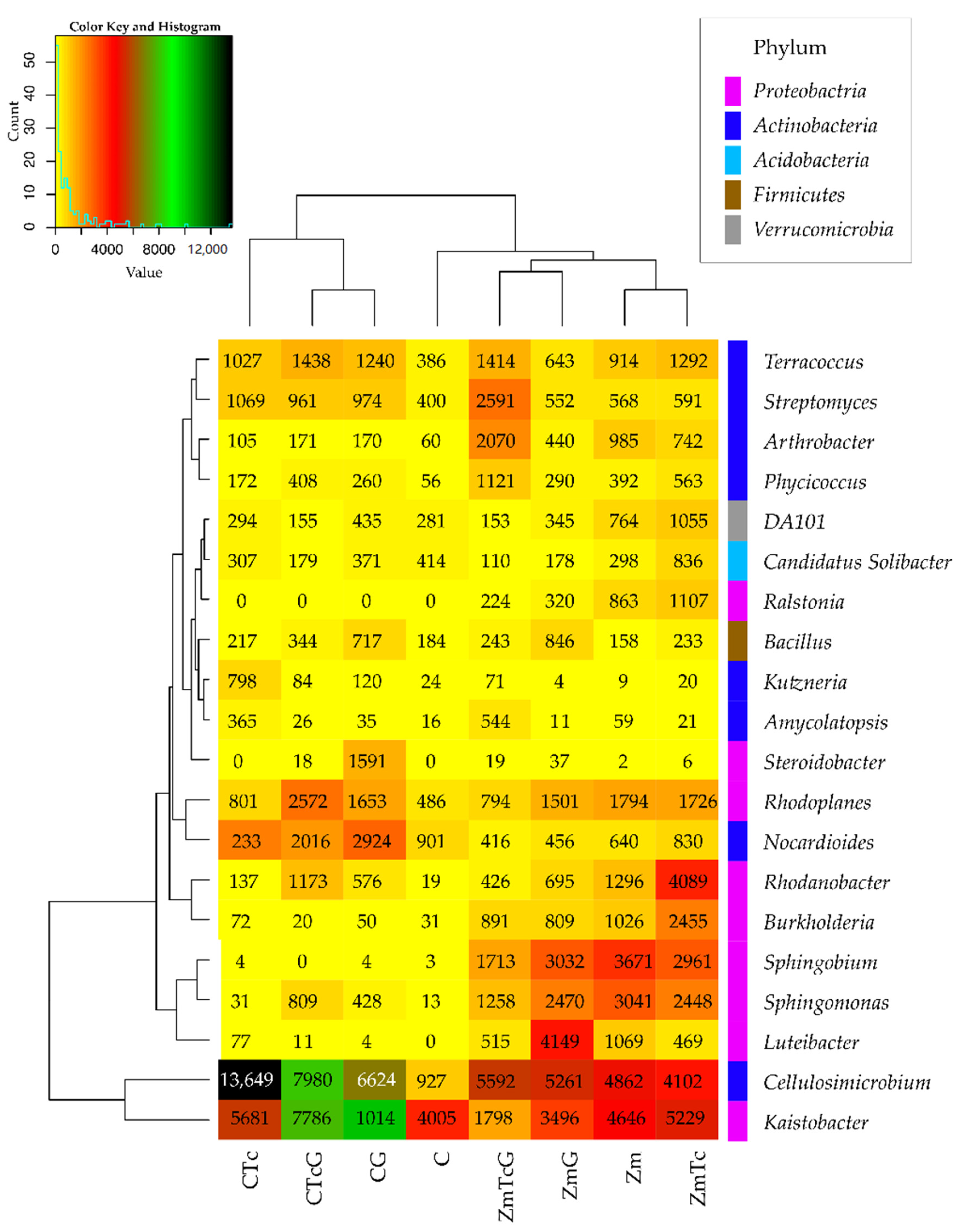
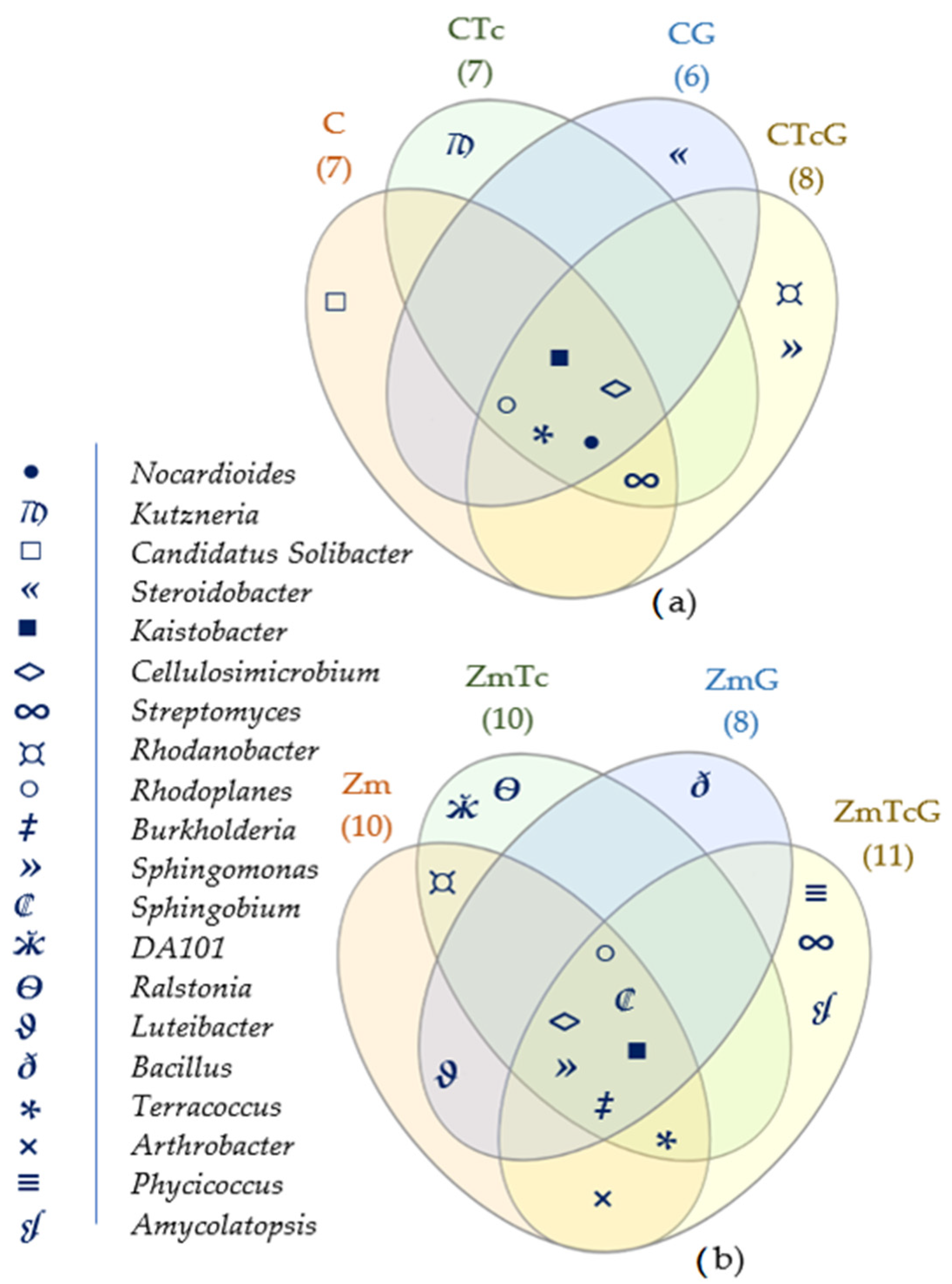
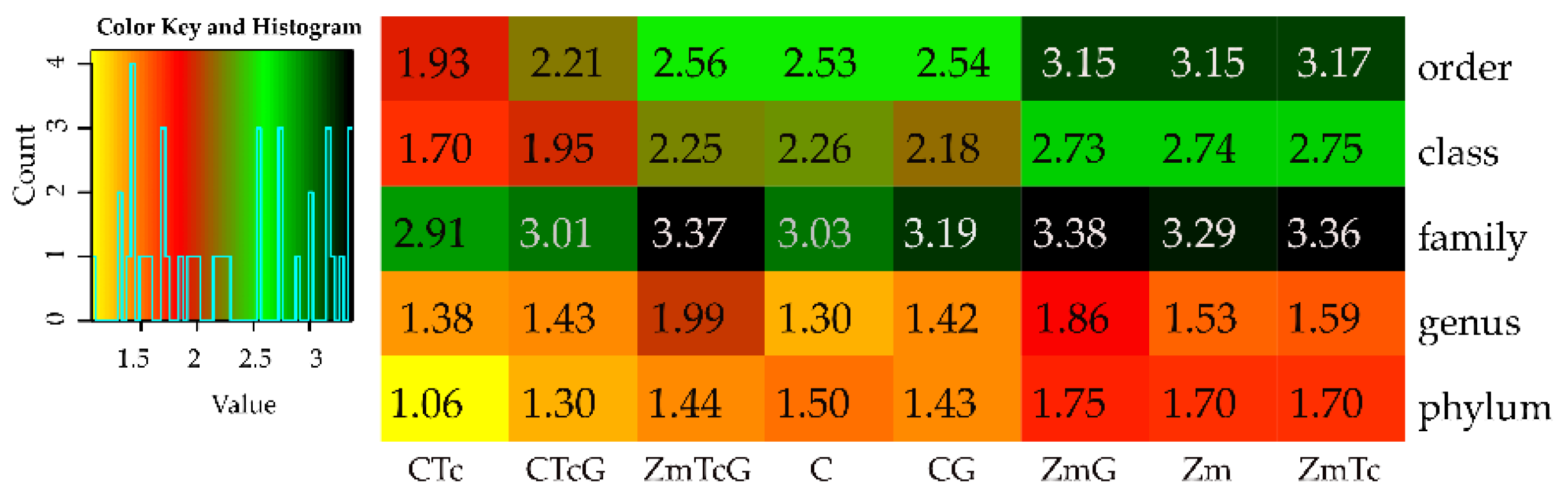
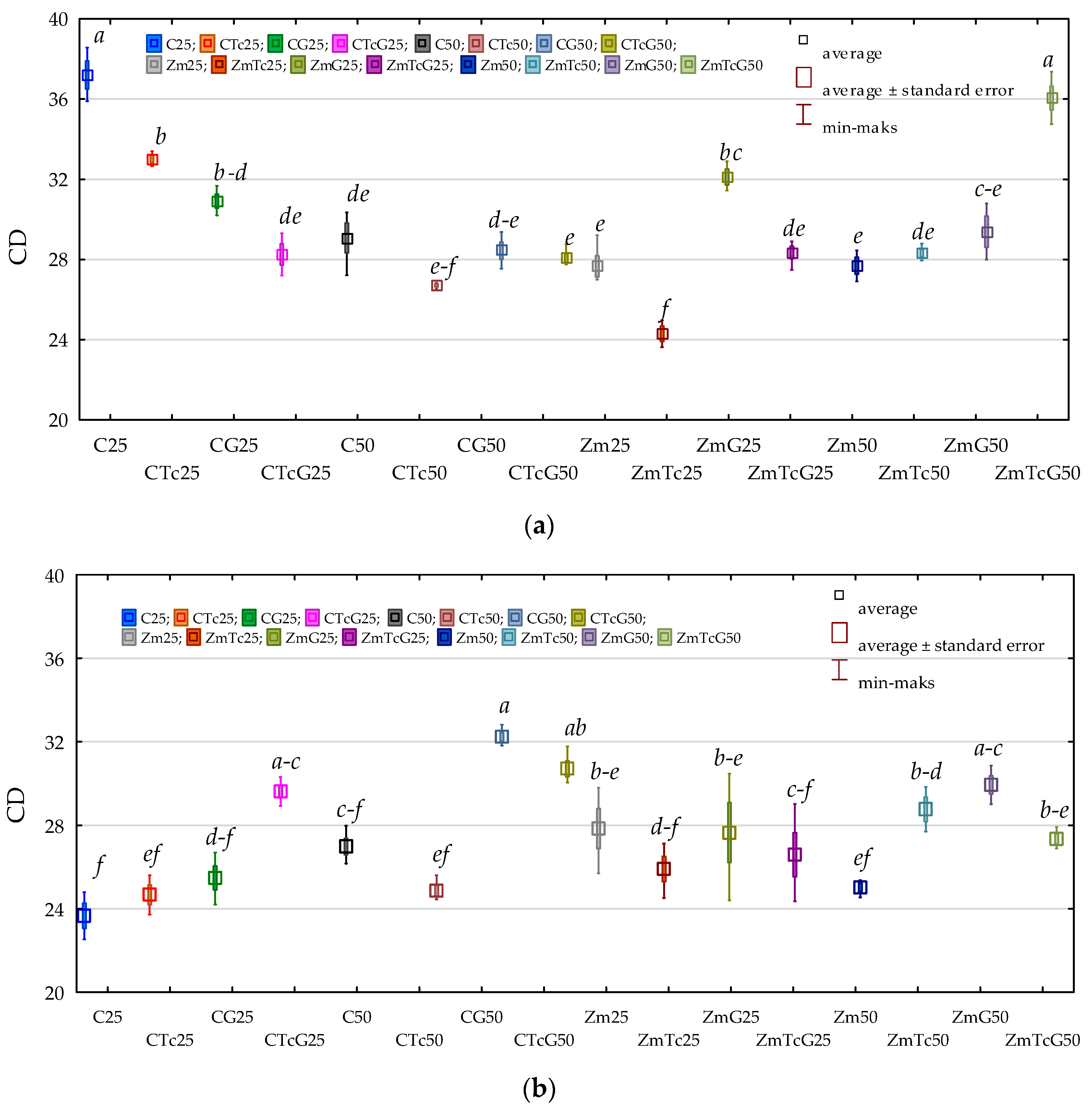
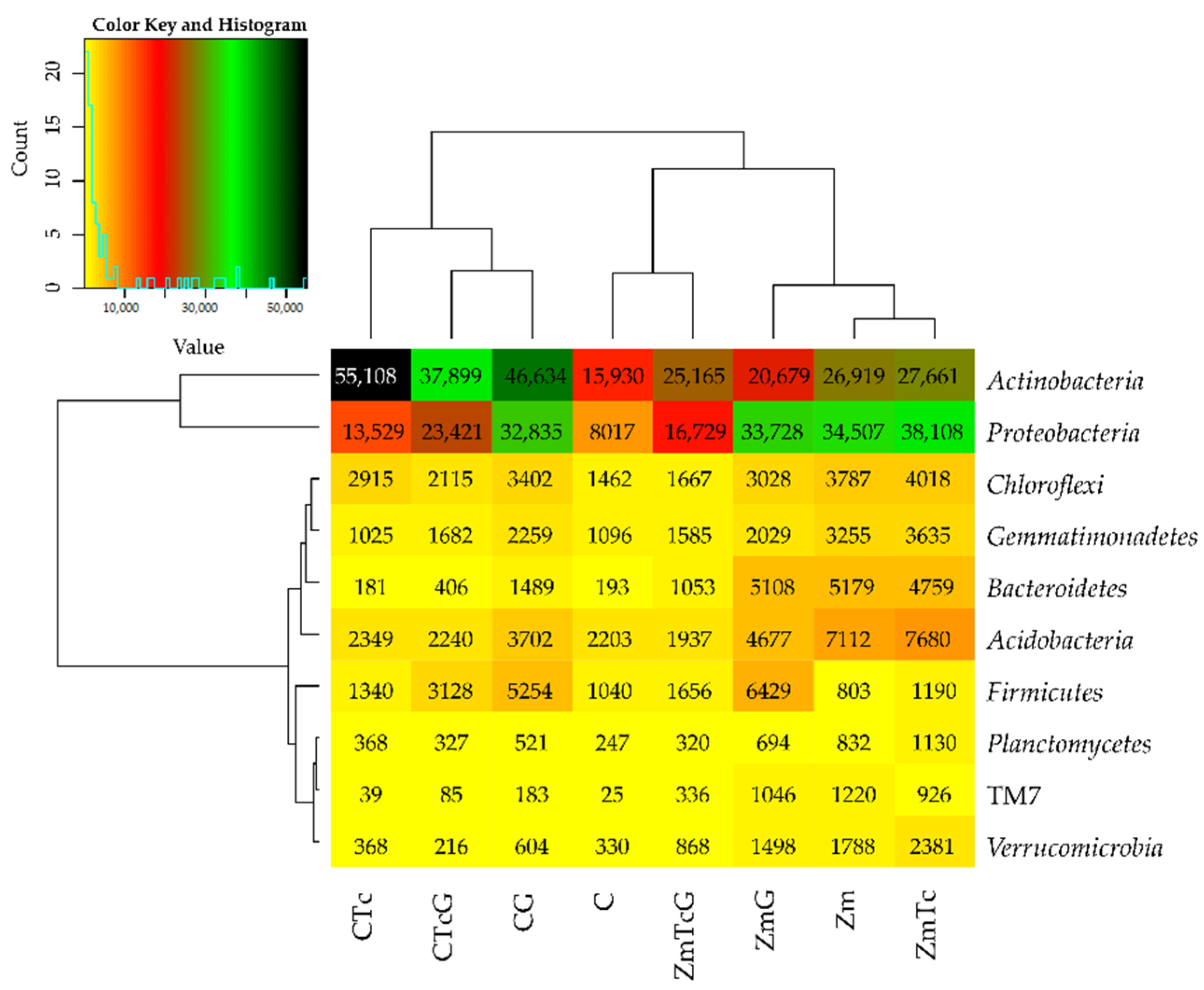
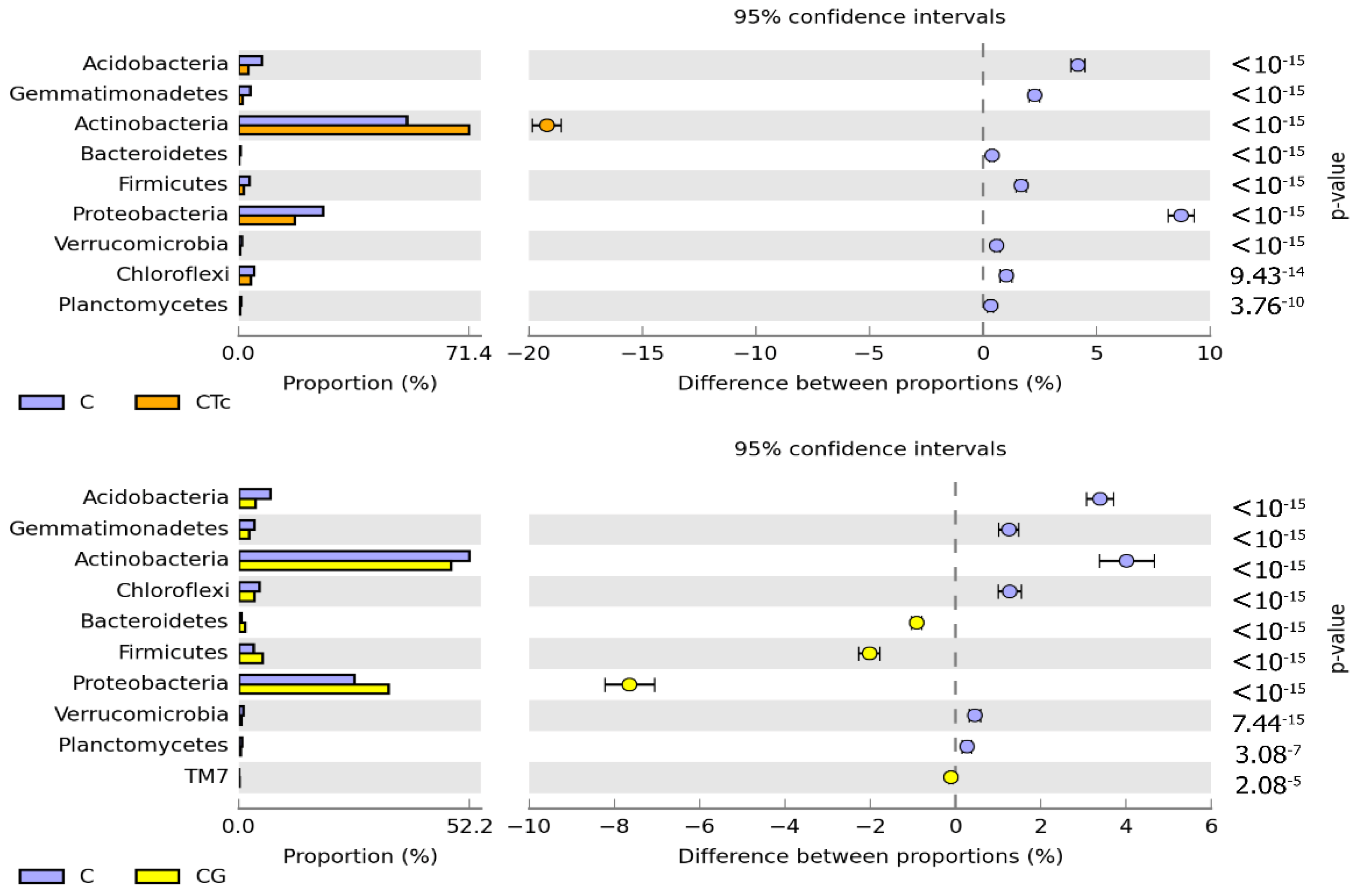
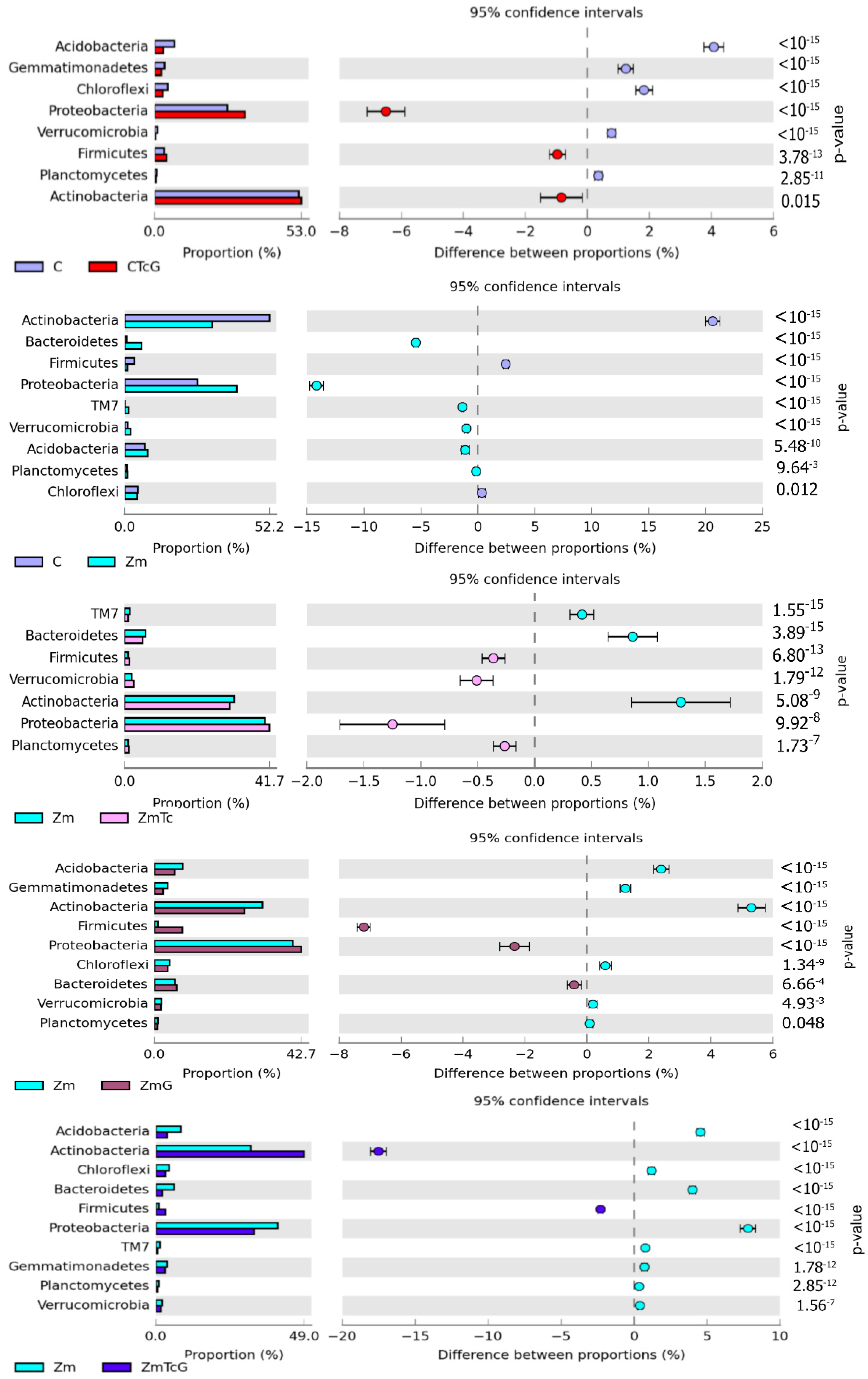
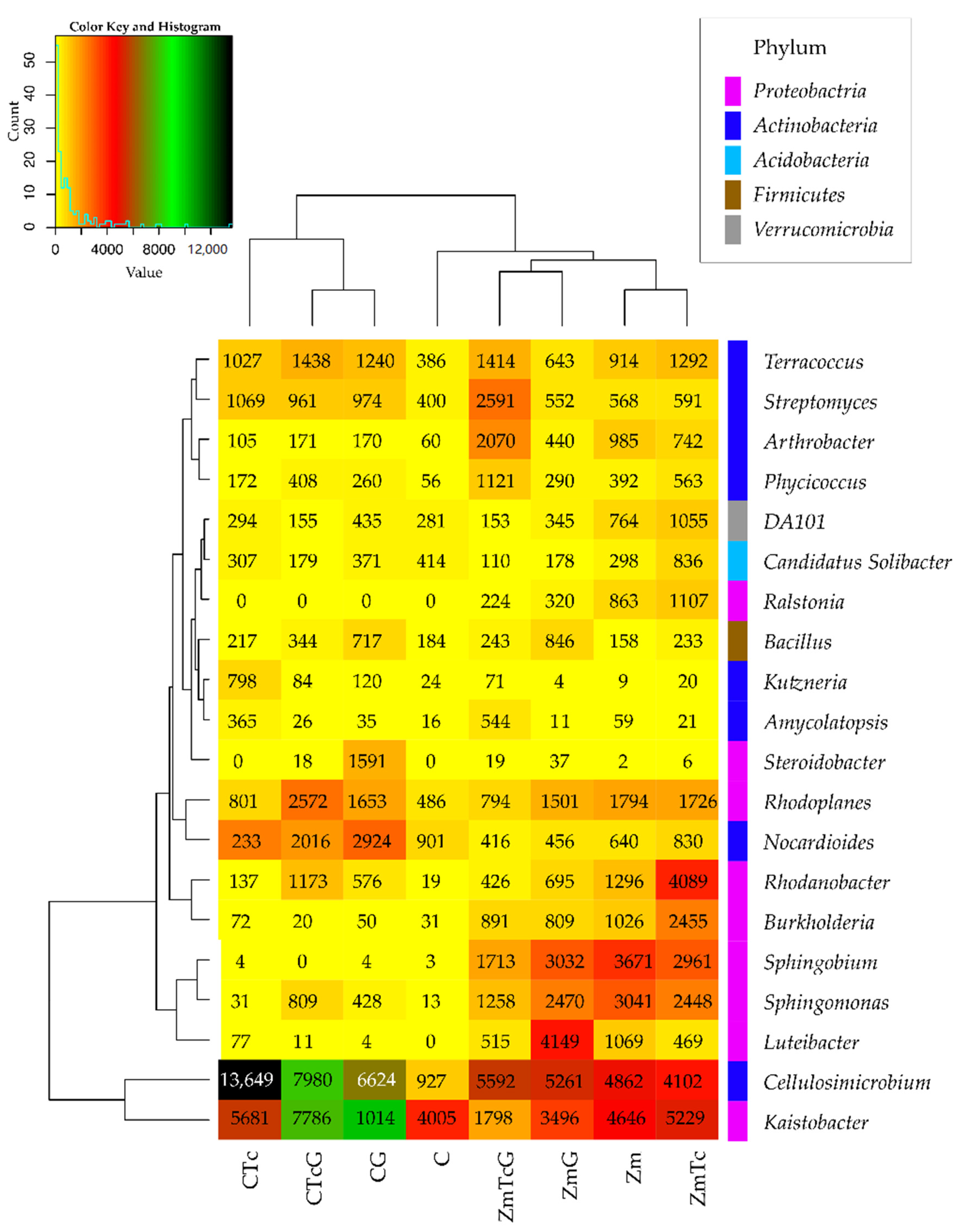
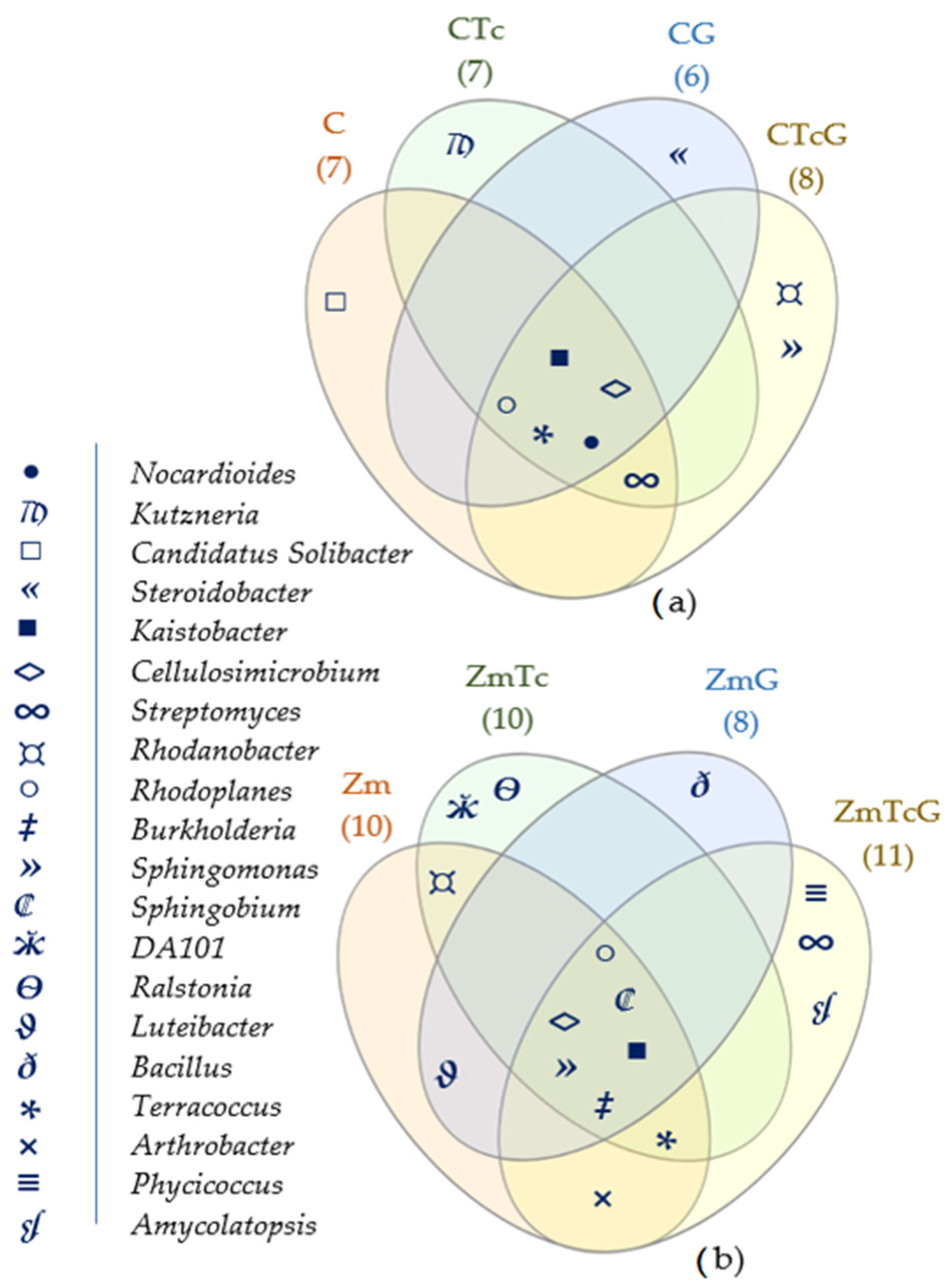
3.2. Response of Bacteria to Soil Contamination with Tetracycline
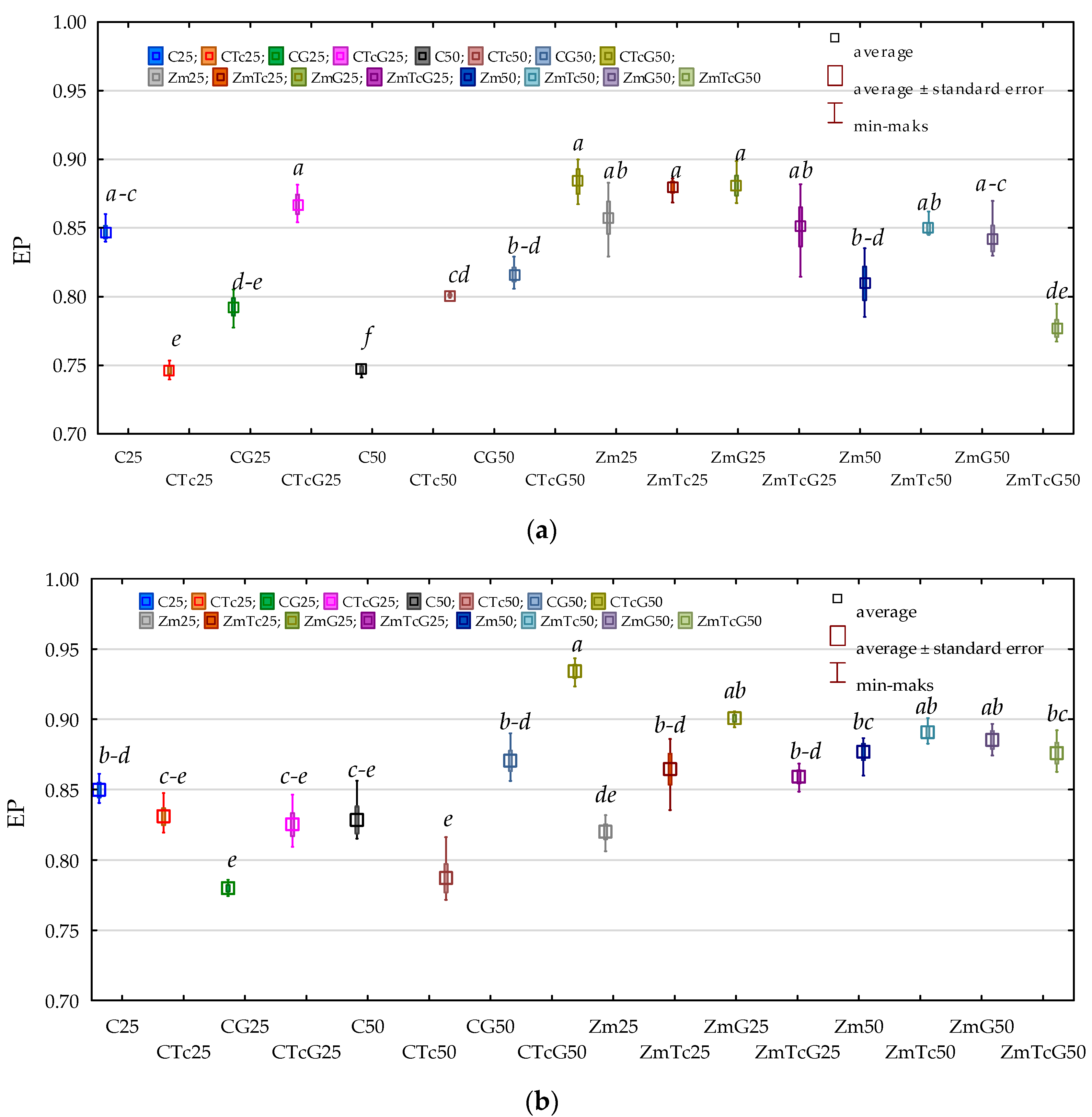
3.3. Response of Oxidoreductases to Soil Contamination with Tetracycline
4. Discussion
4.1. Effect of Tetracycline and Soil Fertilization on Plants
4.2. Effect of Tetracycline, Fertilization with Compost, and Cultivation of Zea Mays on Soil Bacteria Community
4.3. Effect of Tetracycline, Fertilization with Compost, and Cultivation of Zea Mays on Activities of Soil Oxidoreductases
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusu, A.; Buta, E.L. The Development of third-generation tetracycline antibiotics and new perspectives. Pharmaceutics 2021, 13, 2085. [Google Scholar] [CrossRef] [PubMed]
- Granados-Chinchilla, F.; Rodríguez, C. Tetracyclines in food and feedingstuffs: From regulation to analytical methods, bacterial resistance, and environmental and health implications. J. Anal. Methods Chem. 2017, 2017, e1315497. [Google Scholar] [CrossRef] [PubMed]
- Fuoco, D. Classification framework and chemical biology of tetracycline-structure-based drugs. Antibiotics 2012, 1, 1–13. [Google Scholar] [CrossRef]
- OIE. OIE Annual Report on Antimicrobial Agents Intended for Use in Animals: Better Understanding of the Global Situation; Fifth Report; OIE: Paris, France, 2021; p. 136. [Google Scholar]
- EMA. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2019 and 2020. EMA; Publications Office of the European Union: Luxembourg, Germany, 2021; p. 130. [Google Scholar]
- WHO. Critically Important Antimicrobials for Human Medicine, 6th Revision; 6th Revision; World Health Organization: Geneva, Switzerland, 2019; p. 52. [Google Scholar]
- Ahn, J.G.; Cho, H.-K.; Li, D.; Choi, M.; Lee, J.; Eun, B.-W.; Jo, D.S.; Park, S.E.; Choi, E.H.; Yang, H.-J.; et al. Efficacy of tetracyclines and fluoroquinolones for the treatment of macrolide-refractory mycoplasma pneumoniae pneumonia in children: A systematic review and meta-analysis. BMC Infect. Dis. 2021, 21, 1003. [Google Scholar] [CrossRef]
- Furlong-Silva, J.; Cross, S.D.; Marriott, A.E.; Pionnier, N.; Archer, J.; Steven, A.; Merker, S.S.; Mack, M.; Hong, Y.-K.; Taylor, M.J.; et al. Tetracyclines improve experimental lymphatic filariasis pathology by disrupting interleukin-4 receptor–mediated lymphangiogenesis. J. Clin. Investig. 2021, 131, e140853. [Google Scholar] [CrossRef]
- Mdegela, R.H.; Mwakapeje, E.R.; Rubegwa, B.; Gebeyehu, D.T.; Niyigena, S.; Msambichaka, V.; Nonga, H.E.; Antoine-Moussiaux, N.; Fasina, F.O. Antimicrobial use, residues, resistance and governance in the food and agriculture sectors, Tanzania. Antibiotics 2021, 10, 454. [Google Scholar] [CrossRef]
- Van, T.T.H.; Yidana, Z.; Smooker, P.M.; Coloe, P.J. Antibiotic use in food animals worldwide, with a focus on africa: Pluses and minuses. J. Glob. Antimicrob. Resist. 2020, 20, 170–177. [Google Scholar] [CrossRef]
- Albernaz-Gonçalves, R.; Olmos Antillón, G.; Hötzel, M.J. Linking animal welfare and antibiotic use in pig farming—A review. Animals 2022, 12, 216. [Google Scholar] [CrossRef]
- Doidge, C.; West, H.; Kaler, J. Antimicrobial resistance patterns of escherichia coli isolated from sheep and beef farms in england and wales: A Comparison of disk diffusion interpretation methods. Antibiotics 2021, 10, 453. [Google Scholar] [CrossRef]
- Hao, H.; Cheng, G.; Iqbal, Z.; Ai, X.; Hussain, H.I.; Huang, L.; Dai, M.; Wang, Y.; Liu, Z.; Yuan, Z. Benefits and risks of antimicrobial use in food-producing animals. Front. Microbiol. 2014, 5, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasho, R.P.; Cho, J.Y. Veterinary antibiotics in animal waste, its distribution in soil and uptake by plants: A review. Sci. Total Environ. 2016, 563–564, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Daghrir, R.; Drogui, P. Tetracycline antibiotics in the environment: A review. Environ. Chem. Lett. 2013, 11, 209–227. [Google Scholar] [CrossRef]
- Santás-Miguel, V.; Fernández-Sanjurjo, M.J.; Núñez-Delgado, A.; Álvarez-Rodríguez, E.; Díaz-Raviña, M.; Arias-Estévez, M.; Fernández-Calviño, D. Use of waste materials to prevent tetracycline antibiotics toxicity on the growth of soil bacterial communities. Environ. Res. 2021, 193, 110404. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Guo, Y.; Shi, M.; Qiu, T.; Gao, M.; Tian, S.; Wang, X. Effect of antibiotic type and vegetable species on antibiotic accumulation in soil-vegetable system, soil microbiota, and resistance genes. Chemosphere 2021, 263, 128099. [Google Scholar] [CrossRef]
- Awad, Y.M.; Kim, S.-C.; Abd El-Azeem, S.A.M.; Kim, K.-H.; Kim, K.-R.; Kim, K.; Jeon, C.; Lee, S.S.; Ok, Y.S. Veterinary antibiotics contamination in water, sediment, and soil near a swine manure composting facility. Environ. Earth Sci. 2014, 71, 1433–1440. [Google Scholar] [CrossRef]
- An, J.; Chen, H.; Wei, S.; Gu, J. Antibiotic contamination in animal manure, soil, and sewage sludge in Shenyang, Northeast China. Environ. Earth Sci. 2015, 74, 5077–5086. [Google Scholar] [CrossRef]
- Li, C.; Chen, J.; Wang, J.; Ma, Z.; Han, P.; Luan, Y.; Lu, A. Occurrence of antibiotics in soils and manures from greenhouse vegetable production bases of Beijing, China and an associated risk assessment. Sci. Total Environ. 2015, 521–522, 101–107. [Google Scholar] [CrossRef]
- Andreu, V.; Vazquez-Roig, P.; Blasco, C.; Picó, Y. Determination of tetracycline residues in soil by pressurized liquid extraction and liquid chromatography tandem mass spectrometry. Anal. Bioanal. Chem. 2009, 394, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Conde-Cid, M.; Álvarez-Esmorís, C.; Paradelo-Núñez, R.; Nóvoa-Muñoz, J.C.; Arias-Estévez, M.; Álvarez-Rodríguez, E.; Fernández-Sanjurjo, M.J.; Núñez-Delgado, A. Occurrence of tetracyclines and sulfonamides in manures, agricultural soils and crops from different areas in Galicia (NW Spain). J. Clean. Prod. 2018, 197, 491–500. [Google Scholar] [CrossRef]
- Hamscher, G.; Sczesny, S.; Höper, H.; Nau, H. Determination of persistent tetracycline residues in soil fertilized with liquid manure by high-performance liquid chromatography with electrospray ionization tandem mass spectrometry. Anal. Chem. 2002, 74, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Łukaszewicz, P.; Białk-Bielińska, A.; Dołżonek, J.; Kumirska, J.; Caban, M.; Stepnowski, P. A New approach for the extraction of tetracyclines from soil matrices: Application of the microwave-extraction technique. Anal. Bioanal. Chem. 2018, 410, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Nie, G.; Feng, G.; Han, J.; Huang, L.; Zhang, X. Genome-wide identification, characterization, and expression analysis of the NAC transcription factor family in orchardgrass (Dactylis Glomerata L.). BMC Genom. 2021, 22, 178. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Yang, L.; Zhang, L.; Ye, B.; Wang, L. Antibiotics in soil and water in china–a systematic review and source analysis. Environ. Pollut. 2020, 266, 115147. [Google Scholar] [CrossRef]
- Wei, R.; Ge, F.; Zhang, L.; Hou, X.; Cao, Y.; Gong, L.; Chen, M.; Wang, R.; Bao, E. Occurrence of 13 veterinary drugs in animal manure-amended soils in Eastern China. Chemosphere 2016, 144, 2377–2383. [Google Scholar] [CrossRef]
- Xu, L.; Wang, W.; Xu, W. Effects of tetracycline antibiotics in chicken manure on soil microbes and antibiotic resistance genes (ARGs). Environ. Geochem. Health 2022, 44, 273–284. [Google Scholar] [CrossRef]
- Chen, C.; Li, J.; Chen, P.; Ding, R.; Zhang, P.; Li, X. Occurrence of antibiotics and antibiotic resistances in soils from wastewater irrigation areas in Beijing and Tianjin, China. Environ. Pollut. 2014, 193, 94–101. [Google Scholar] [CrossRef]
- He, L.-Y.; He, L.-K.; Gao, F.-Z.; Wu, D.-L.; Zou, H.-Y.; Bai, H.; Zhang, M.; Ying, G.-G. Dissipation of antibiotic resistance genes in manure-amended agricultural soil. Sci. Total Environ. 2021, 787, 147582. [Google Scholar] [CrossRef]
- Huang, R.; Guo, Z.; Gao, S.; Ma, L.; Xu, J.; Yu, Z.; Bu, D. Assessment of veterinary antibiotics from animal manure-amended soil to growing alfalfa, alfalfa silage, and milk. Ecotoxicol. Environ. Saf. 2021, 224, 112699. [Google Scholar] [CrossRef]
- Haenni, M.; Dagot, C.; Chesneau, O.; Bibbal, D.; Labanowski, J.; Vialette, M.; Bouchard, D.; Martin-Laurent, F.; Calsat, L.; Nazaret, S.; et al. Environmental contamination in a high-income country (france) by antibiotics, antibiotic-resistant bacteria, and antibiotic resistance genes: Status and possible causes. Environ. Int. 2022, 159, 107047. [Google Scholar] [CrossRef]
- Stachurová, T.; Sýkorová, N.; Semerád, J.; Malachová, K. Resistant genes and multidrug-resistant bacteria in wastewater: A study of their transfer to the water reservoir in the Czech Republic. Life 2022, 12, 147. [Google Scholar] [CrossRef] [PubMed]
- Zhi, S.; Zhou, J.; Yang, F.; Tian, L.; Zhang, K. Systematic analysis of occurrence and variation tendency about 58 typical veterinary antibiotics during animal wastewater disposal processes in Tianjin, China. Ecotoxicol. Environ. Saf. 2018, 165, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Scaria, J.; Anupama, K.V.; Nidheesh, P.V. Tetracyclines in the environment: An overview on the occurrence, fate, toxicity, detection, removal methods, and sludge management. Sci. Total Environ. 2021, 771, 145291. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Wang, M.; Wu, J.; Jiang, Q.; Jin, J.; Jin, Q.; Yang, W.; Chen, J.; Wang, Y.; Xiao, M. Spread of chloramphenicol and tetracycline resistance genes by plasmid mobilization in agricultural soil. Environ. Pollut. 2020, 260, 113998. [Google Scholar] [CrossRef] [PubMed]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- ISO 11261; Soil Quality—Determination of Total Nitrogen—Modified Kjeldahl Method. ISO: Geneva, Switzerland, 1995.
- Nelson, D.; Sommers, L. Total carbon, organic carbon, and organic matter. In Method of Soil Analysis: Chemical Methods; Sparks, D.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1996; pp. 1201–1229. [Google Scholar]
- Egner, H.; Riehm, H.; Domingo, W. Untersuchungen Über Die Chemische Bodenanalyse Als Grundlage Für Die Beurteilung Des Nährstoffzustandes Der böden. II. Chemische extractionsmethoden zur phospor- und kaliumbestimmung. Ann R. Agric. Coll. Swed. 1960, 26, 199–215. [Google Scholar]
- Schlichting, E.; Blume, H.; Stahr, K. Bodenkundliches Praktikum. Pareys Studientexte 81; Blackwell Wissenschafts-Verlag: Berlin, Germany, 1995. [Google Scholar]
- ISO 10390; Soil Quality—Determination of PH. International Organization for Standardization: Geneva, Switzerland, 2005.
- Klute, A. Methods of Soil Analysis; American Society of Agronomy, Agronomy Monograph 9: Madison, WI, USA, 1996. [Google Scholar]
- Thomas, G.W. Exchangeable Cations. In Methods of Soil Analysis; Soil Science Society of America, American Society of Agronomy, Inc.: Madison, WI, USA, 1982; pp. 159–165. [Google Scholar]
- Bunt, J.S.; Rovira, A.D. Microbiological studies of some subantarctic soils. J. Soil Sci. 1955, 6, 119–128. [Google Scholar] [CrossRef]
- Ohta, H.; Hattori, T. Oligotrophic bacteria on organic debris and plant roots in a paddy field soil. Soil Biol. Bioch. 1983, 15, 1–8. [Google Scholar]
- Parkinson, D.; Gray, T.R.G.; Williams, S.T. Methods for Studying the Ecology of Soil Microorganisms; IBP Handbook 19; Blackwell Scientific Publication: Oxford, UK, 1971. [Google Scholar]
- Öhlinger, R. Dehydrogenase activity with the substrate TTC. In Methods in Soil Biology; Schinner, F., Ohlinger, R., Kandler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 241–243. [Google Scholar]
- Johnson, J.L.; Temple, K.L. Some variables affecting the measurement of “catalase activity” in soil. Soil Sci. Soc. Am. J. 1964, 28, 207–209. [Google Scholar] [CrossRef]
- Babić, S.; Horvat, A.J.M.; Mutavdžić Pavlović, D.; Kaštelan-Macan, M. Determination of PKa Values of active pharmaceutical ingredients. Trends Anal. Chem. 2007, 26, 1043–1061. [Google Scholar] [CrossRef]
- Chen, M.; Yi, Q.; Hong, J.; Zhang, L.; Lin, K.; Yuan, D. Simultaneous Determination of 32 antibiotics and 12 pesticides in sediment using ultrasonic-assisted extraction and high performance liquid chromatography-tandem mass spectrometry. Anal. Methods 2015, 7, 1896–1905. [Google Scholar] [CrossRef]
- Conde-Cid, M.; Núñez-Delgado, A.; Fernández-Sanjurjo, M.J.; Álvarez-Rodríguez, E.; Fernández-Calviño, D.; Arias-Estévez, M. Tetracycline and sulfonamide antibiotics in soils: Presence, fate and environmental risks. Processes 2020, 8, 1479. [Google Scholar] [CrossRef]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the soil environment—degradation and their impact on microbial activity and diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- Erenstein, O.; Chamberlin, J.; Sonder, K. Estimating the global number and distribution of maize and wheat farms. Glob. Food Sec. 2021, 30, 100558. [Google Scholar] [CrossRef]
- OECD-FAO. OECD-FAO Agricultural Outlook 2021–2030; OECD Publishing: Paris, France, 2021; p. 337. Available online: https://www.oecd.org/development/oecd-fao-agricultural-outlook-19991142.htm (accessed on 12 February 2022).
- Chao, H.; Zheng, X.; Xia, R.; Sun, M.; Hu, F. Incubation Trial indicated the earthworm intestinal bacteria as promising biodigestor for mitigating tetracycline resistance risk in anthropogenic disturbed forest soil. Sci. Total Environ. 2021, 798, 149337. [Google Scholar] [CrossRef] [PubMed]
- Chessa, L.; Pusino, A.; Garau, G.; Mangia, N.P.; Pinna, M.V. Soil microbial response to tetracycline in two different soils amended with cow manure. Environ. Sci. Pollut. Res. 2016, 23, 5807–5817. [Google Scholar] [CrossRef] [PubMed]
- Borowik, A.; Wyszkowska, J.; Kucharski, M.; Kucharski, J. Implications of soil pollution with diesel oil and bp petroleum with ACTIVE Technology for soil health. Int. J. Environ. Res. Public Health 2019, 16, 2474. [Google Scholar] [CrossRef] [Green Version]
- Ferris, M.J.; Muyzer, G.; Ward, D.M. Denaturing gradient gel electrophoresis profiles of 16s rrna-defined populations inhabiting a hot spring microbial mat community. Appl. Environ. Microbiol. 1996, 62, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Zaborowska, M.; Wyszkowska, J.; Borowik, A.; Kucharski, J. Bisphenol A—A dangerous pollutant distorting the biological properties of soil. Int. J. Mol. Sci. 2021, 22, 12753. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Wyszkowski, M. Resistance of aerobic microorganisms and soil enzyme response to soil contamination with Ekodiesel Ultra Fuel. Environ. Sci. Pollut. Res. 2017, 24, 24346–24363. [Google Scholar] [CrossRef] [Green Version]
- De Leij, F.A.A.M.; Whipps, J.M.; Lynch, J.M. The Use of colony development for the characterization of bacterial communities in soil and on roots. Microb. Ecol. 1993, 27, 81–97. [Google Scholar] [CrossRef]
- TIBCO Software Inc. Statistica (Data Analysis Software System), Version 13. Available online: https://www.tibco.com/products/data-science (accessed on 12 February 2022).
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RStudio Team. R Studio: Integrated Development; RStudio, Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com/ (accessed on 5 March 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austri, 2019; Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 5 March 2022).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, M.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data. R Package Version 2.17.0. 2020. Available online: https://CRAN.R-Project.org/package=gplots (accessed on 5 March 2022).
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Carballo, M.; Rodríguez, A.; de la Torre, A. Phytotoxic effects of antibiotics on terrestrial crop plants and wild plants: A systematic review. Arch. Environ. Contam. Toxicol. 2022, 82, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ying, G.-G.; Tao, R.; Zhao, J.-L.; Yang, J.-F.; Zhao, L.-F. Effects of six selected antibiotics on plant growth and soil microbial and enzymatic activities. Environ. Pollut. 2009, 157, 1636–1642. [Google Scholar] [CrossRef]
- Wei, X.; Wu, S.C.; Nie, X.P.; Yediler, A.; Wong, M.H. The effects of residual tetracycline on soil enzymatic activities and plant growth. J. Environ. Sci. Health B 2009, 44, 461–471. [Google Scholar] [CrossRef]
- Pan, M.; Chu, L.M. Phytotoxicity of veterinary antibiotics to seed germination and root elongation of crops. Ecotoxicol. Environ. Saf. 2016, 126, 228–237. [Google Scholar] [CrossRef]
- Wang, R.; Wang, J.; Wang, J.; Zhu, L.; Zhang, W.; Zhao, X.; Ahmad, Z. Growth inhibiting effects of four antibiotics on cucumber, rape and chinese cabbage. Bull. Environ. Contam. Toxicol. 2019, 103, 187–192. [Google Scholar] [CrossRef]
- Timmerer, U.; Lehmann, L.; Schnug, E.; Bloem, E. Toxic effects of single antibiotics and antibiotics in combination on germination and growth of Sinapis alba L. Plants 2020, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Liu, W.; Pan, N.; Jiao, W.; Wang, M. Oxytetracycline on functions and structure of soil microbial community. J. Soil Sci. Plant Nutr. 2013, 13, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Lyu, H.; Cheng, Z.; Wang, Y.; Tang, J. Insight into the mechanisms of ball-milled biochar addition on soil tetracycline degradation enhancement: Physicochemical properties and microbial community structure. Chemosphere 2022, 291, 132691. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, J.; Gao, L.; Kong, F.; Shen, G.; Wang, R.; Gao, J.; Zhang, J. The Effects of tetracycline residues on the microbial community structure of tobacco soil in pot experiment. Sci. Rep. 2020, 10, 8804. [Google Scholar] [CrossRef] [PubMed]
- Santás-Miguel, V.; Rodríguez-González, L.; Núñez-Delgado, A.; Álvarez-Rodríguez, E.; Díaz-Raviña, M.; Arias-Estévez, M.; Fernández-Calviño, D. Time-course evolution of bacterial community tolerance to tetracycline antibiotics in agricultural soils: A laboratory experiment. Chemosphere 2022, 291, 132758. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.S.; Wang, W.Z.; Deng, J.B.; Xu, W.H. The residue of tetracycline antibiotics in soil and brassica juncea var. gemmifera, and the diversity of soil bacterial community under different livestock manure treatments. Environ. Geochem. Health, 2022; Online ahead of print. [Google Scholar] [CrossRef]
- Santás-Miguel, V.; Arias-Estévez, M.; Díaz-Raviña, M.; Fernández-Sanjurjo, M.J.; Álvarez-Rodríguez, E.; Núñez-Delgado, A.; Fernández-Calviño, D. Interactions between soil properties and tetracycline toxicity affecting to bacterial community growth in agricultural soil. Appl. Soil Ecol. 2020, 147, 103437. [Google Scholar] [CrossRef]
- Tong, X.; Wang, X.; He, X.; Wang, Z.; Li, W. Effects of Antibiotics on microbial community structure and microbial functions in constructed wetlands treated with artificial root exudates. Environ. Sci. Processes Impacts 2020, 22, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lin, H.; Ma, J.; Zhu, R.; Sun, W.; Lin, X.; Zhang, J.; Zheng, H.; Zhang, X. Degradation of tetracycline antibiotics by Arthrobacter nicotianae OTC-16. J. Hazard. Mater. 2021, 403, 123996. [Google Scholar] [CrossRef]
- Ma, J.; Lin, H.; Sun, W.; Wang, Q.; Yu, Q.; Zhao, Y.; Fu, J. Soil microbial systems respond differentially to tetracycline, sulfamonomethoxine, and ciprofloxacin entering soil under pot experimental conditions alone and in combination. Environ. Sci. Pollut. Res. 2014, 21, 7436–7448. [Google Scholar] [CrossRef]
- Santás-Miguel, V.; Díaz-Raviña, M.; Martín, A.; García-Campos, E.; Barreiro, A.; Núñez-Delgado, A.; Álvarez-Rodríguez, E.; Arias-Estévez, M.; Fernández-Calviño, D. Soil enzymatic activities and microbial community structure in soils polluted with tetracycline antibiotics. Agronomy 2021, 11, 906. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; MMS., C.-P.; Chaturvedi, A.K.; Shabnam, A.A.; Subrahmanyam, G.; Mondal, R.; Gupta, D.K.; Malyan, S.K.; Kumar, S.S.; et al. Lead toxicity: Health hazards, influence on food chain, and sustainable remediation approaches. Int. J. Environ. Res. Public Health 2020, 17, 2179. [Google Scholar] [CrossRef] [Green Version]
- Torres-Cortés, G.; Millán, V.; Ramírez-Saad, H.C.; Nisa-Martínez, R.; Toro, N.; Martínez-Abarca, F. Characterization of novel antibiotic resistance genes identified by functional metagenomics on soil samples. Environ. Microbiol. 2011, 13, 1101–1114. [Google Scholar] [CrossRef]
- Chen, Q.; An, X.; Li, H.; Su, J.; Ma, Y.; Zhu, Y.-G. Long-term field application of sewage sludge increases the abundance of antibiotic resistance genes in soil. Environ. Int. 2016, 92–93, 1–10. [Google Scholar] [CrossRef]
- Kyselková, M.; Jirout, J.; Vrchotová, N.; Schmitt, H.; Elhottová, D. Spread of tetracycline resistance genes at a conventional dairy farm. Front. Microbiol. 2015, 6, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, Y.; Liu, Y.-J.; Wang, J.; Vukanti, R.; Ge, Y. Enrichment of potential degrading bacteria accelerates removal of tetracyclines and their epimers from cow manure biochar amended soil. Chemosphere 2021, 278, 130358. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, A.; Kukal, S.S.; Saha, D.; Sharma, H.; Kalia, A.; Sharma, S. Potential Indicators of soil health degradation in different land use-based ecosystems in the Shiwaliks of Northwestern India. Sustainability 2019, 11, 3908. [Google Scholar] [CrossRef] [Green Version]
- Kucharski, J.; Tomkiel, M.; Baćmaga, M.; Borowik, A.; Wyszkowska, J. Enzyme activity and microorganisms diversity in soil contaminated with the Boreal 58 WG Herbicide. J. Environ. Sci. Health B 2016, 51, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Steffan, J.J.; Brevik, E.C.; Burgess, L.C.; Cerdà, A. The effect of soil on human health: An overview. Eur. J. Soil Sci. 2018, 69, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Wyszkowska, J.; Borowik, A.; Olszewski, J.; Kucharski, J. Soil Bacterial community and soil enzyme activity depending on the cultivation of Triticum aestivum, Brassica napus, and Pisum sativum ssp. arvense. Diversity 2019, 11, 246. [Google Scholar] [CrossRef] [Green Version]
- Kessler, N.C.H.; Sampaio, S.C.; do Prado, N.V.; Remor, M.B.; dos Reis, R.R.; Cordovil, C.M. Effects of tetracyclines on enzymatic activity and soil nutrient availability. J. Soil Sci. Plant Nutr. 2020, 20, 2657–2670. [Google Scholar] [CrossRef]
- Liu, B.; Li, Y.; Zhang, X.; Wang, J.; Gao, M. Effects of chlortetracycline on soil microbial communities: Comparisons of enzyme activities to the functional diversity via Biolog EcoPlatesTM. Eur. J. Soil Biol. 2015, 68, 69–76. [Google Scholar] [CrossRef]
- Zhao, X.; Li, X.; Li, Y.; Zhang, X.; Zhai, F.; Ren, T.; Li, Y. Metagenomic analysis reveals functional genes in soil microbial electrochemical removal of tetracycline. J. Hazard. Mater. 2021, 408, 124880. [Google Scholar] [CrossRef]
- Hammerschmiedt, T.; Holatko, J.; Kucerik, J.; Mustafa, A.; Radziemska, M.; Kintl, A.; Malicek, O.; Baltazar, T.; Latal, O.; Brtnicky, M. Manure maturation with biochar: Effects on plant biomass, manure quality and soil microbiological characteristics. Agriculture 2022, 12, 314. [Google Scholar] [CrossRef]
- Khalifa, T.H.; Mariey, S.A.; Ghareeb, Z.E.; Khatab, I.A.; Alyamani, A. Effect of organic amendments and nano-zinc foliar application on alleviation of water stress in some soil properties and water productivity of barley yield. Agronomy 2022, 12, 585. [Google Scholar] [CrossRef]
- Wojewódzki, P.; Lemanowicz, J.; Debska, B.; Haddad, S.A. Soil enzyme activity response under the amendment of different types of biochar. Agronomy 2022, 12, 569. [Google Scholar] [CrossRef]
- Kaczyńska, G.; Borowik, A.; Wyszkowska, J. Soil dehydrogenases as an indicator of contamination of the environment with petroleum products. Water Air Soil Pollut. 2015, 226, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucharski, J.; Wieczorek, K.; Wyszkowska, J. Changes in the enzymatic activity in sandy loam soil exposed to zinc pressure. J. Elem. 2011, 16, 577–589. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Boros-Lajszner, E.; Lajszner, W.; Kucharski, J. Reaction of soil enzymes and spring barley to copper chloride and copper sulphate. Environ. Earth Sci. 2017, 76, 403. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Sarkar, B.; Bolan, N.S.; Ok, Y.S.; Churchman, G.J. Decomposition of soil organic matter as affected by clay types, pedogenic oxides and plant residue addition rates. J. Hazard. Mater. 2019, 374, 11–19. [Google Scholar] [CrossRef]
- Zhang, N.; He, X.-D.; Gao, Y.-B.; Li, Y.-H.; Wang, H.-T.; Ma, D.; Zhang, R.; Yang, S. Pedogenic Carbonate and soil dehydrogenase activity in response to soil organic matter in Artemisia ordosica community. Pedosphere 2010, 20, 229–235. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Soil Sample Description | Maximum Antibiotics, Concentration, µg kg−1 | References |
|---|---|---|---|
| city of Shenyan, northeast China | depth of 0–15 cm | 976.17 | [20] |
| China, Beijing, | soils from vegetable greenhouses | 74.4 | [21] |
| Spain, South surrounding of Valencia | agricultural soils with 20% of clay | 64.4 | [22] |
| Korea, Hongcheon, Gangwon province, | paddy and upland soils | 177.6 | [19] |
| Spain, NW, Galicia | sand 40%, silt—38%, clay—22% | 600 | [23] |
| Germany | 0–10, 10–20 and 20–30 cm soils sand 91.6%, silt—6.0%, clay—2.4% | 86.2; 198.7; 171.7 | [24] |
| Poland, areas of Pomeranian Voivodeship | soils from agricultural areas of Pomeranian Voivodeship | 14.5 | [25] |
| Abbreviation | Properties | Unit | Soil | Compost | References |
|---|---|---|---|---|---|
| Chemical and physicochemical properties | |||||
| Ntot | total nitrogen | g kg−1 DM | 0.83 ± 0.05 | 20.18 ± 0.86 | [39] |
| Corg | organic carbon | 10.00 ± 0.60 | 146.61 ± 6.19 | [40] | |
| SOM | soil organic matter | 17.24 ± 1.03 | 252.76 ± 10.67 | [40] | |
| P | phosphorus | mg kg−1 DM | 81.10 ± 4.13 | 3.41 ± 0.18 | [41] |
| K | potassium | 145.25 ± 7.18 | 9.25 ± 0.24 | [41] | |
| Mg | magnesium | 71.00 ± 3.00 | 5.69 ± 0.17 | [42] | |
| pH | pHKCl—soil reaction | 1 mol KCl dm−3 | 4.40 ± 0.15 | 6.1 ± 0.15 | [43] |
| EBC | sum of exchangeable base cations | mmol (+) kg−1 DM | 63.60 ± 4.50 | 659.76 ± 32.01 | [44] |
| HAC | hydrolytic acidity | 26.10 ± 1.92 | 82.04 ± 3.98 | [44] | |
| CEC | cation exchange capacity | 89.70 | 741.80 | [44] | |
| ACS | alkaline cation saturation | % | 70.90 | 88.94 | [44] |
| Exchangeable cations | |||||
| K+ | potassium | mg kg−1 DM | 168.00 ± 8.30 | NA | [45] |
| Ca++ | calcium | 1190.5 ± 52.70 | NA | [45] | |
| Na+ | sodium | 10.00 ± 0.43 | NA | [45] | |
| Mg++ | magnesium | 82.10 ± 4.70 | NA | [45] | |
| Microorganisms number per 1 kg DM | |||||
| Org | organotrophic bacteria | cfu | 36.728 ± 1.986 × 109 | NA | [46] |
| Olig | oligotrophic bacteria | 7.740 ± 0.581 × 109 | NA | [47] | |
| Cop | copiotrophic bacteria | 11.240 ± 0.127 × 109 | NA | [47] | |
| Act | actinomyces | 17.008 ± 0.537 × 109 | NA | [48] | |
| Enzymatic activity per 1 kg DM h−1 | |||||
| Deh | dehydrogenases | µmol TFF | 4.042 ± 0.136 | NA | [49] |
| Cat | catalase | mol O2 | 0.212 ± 0.001 | NA | [50] |
| Tc Content (mg kg−1 DM of soil) | Yield, (DM g pot−1) | Zea Mays Development Phase SPAD | |||
|---|---|---|---|---|---|
| Shoots | Roots | Together | 4 Leaves | 8 Leaves | |
| −G | |||||
| 0 | 55.33 b ± 0.92 | 13.51 bc ± 0.44 | 68.84 c ± 0.44 | 38.10 a ± 2.74 | 22.62 b ± 4.89 |
| 100 | 56.49 b ± 1.55 | 14.69 ab ± 0.42 | 71.18 bc ± 0.64 | 38.69 a ± 3.62 | 28.00 a ± 2.36 |
| +G | |||||
| 0 | 60.46 a ± 0.93 | 12.07 c ± 1.44 | 72.53 b ± 1.12 | 34.36 c ± 2.36 | 29.01 a ± 1.54 |
| 100 | 62.48 a ± 1.03 | 15.83 a ± 0.72 | 78.31 a ± 1.37 | 36.33 b ± 2.61 | 27.48 a ± 1.58 |
| The Dose of Grass Compost in g C kg−1 DM Soil | −Zm | +Zm | ||
|---|---|---|---|---|
| Analysis Day | ||||
| 25 | 50 | 25 | 50 | |
| Organotrophic bacteria (Org) | ||||
| 0 | −0.352 d ± 0.011 | −0.514 e ± 0.046 | −0.122 ab ± 0.007 | −0.123 ab ± 0.014 |
| 4 | −0.093 ab ± 0.005 | −0.161 bc ± 0.012 | −0.236 c ± 0.025 | −0.061 a ± 0.002 |
| Oligotrophic bacteria (Olig) | ||||
| 0 | −0.262 f ± 0.022 | −0.009 d ± 0.001 | −0.469 g ± 0.027 | −0.052 de ± 0.005 |
| 4 | −0.102 e ± 0.008 | 0.223 b ± 0.017 | 0.687 a ± 0.029 | 0.134 c ± 0.008 |
| Copiotrophic bacteria (Cop) | ||||
| 0 | 0.214 c–e ± 0.015 | 0.375 bc ± 0.053 | 0.300 cd ± 0.064 | 0.591 a ± 0.007 |
| 4 | 0.070 e ± 0.006 | 0.388 bc ± 0.051 | 0.139 de ± 0.005 | 0.529 ab ± 0.051 |
| Actinomycetes (Act) | ||||
| 0 | −0.633 c ± 0.060 | −0.265 b ± 0.067 | −0.063 b ± 0.006 | −0.259 b ± 0.016 |
| 4 | −0.222 ab ± 0.021 | −0.217 ab ± 0.037 | −0.054 a ± 0.005 | −0.118 ab ± 0.53 |
| Tc Content (mg kg−1 DM of soil) | −Zm | +Zm | ||
|---|---|---|---|---|
| Analysis Day | ||||
| 25 | 50 | 25 | 50 | |
| Organotrophic bacteria (Org) | ||||
| 0 | 0.289 c ± 0.060 | 0.150 c–e ± 0.015 | 0.203 cd ± 0.021 | −0.060 f ± 0.014 |
| 100 | 0.804 b ± 0.083 | 0.988 a ± 0.047 | 0.046 d–f ± 0.004 | 0.005 ef ± 0.002 |
| Oligotrophic bacteria (Olig) | ||||
| 0 | −0.333 e ± 0.029 | −0.118 cd ± 0.019 | −0.353 e ± 0.045 | −0.189 d ± 0.017 |
| 100 | −0.189 d ± 0.007 | 0.089 b ± 0.023 | 1.057 a ± 0.036 | −0.031 c ± 0.010 |
| Copiotrophic bacteria (Cop) | ||||
| 0 | 0.190 ab ± 0.019 | 0.058 c ± 0.020 | 0.278 a ± 0.033 | 0.074 c ± 0.036 |
| 100 | 0.049 c ± 0.005 | 0.068 c ± 0.015 | 0.120 ab ± 0.053 | 0.032 c ± 0.021 |
| Actinomycetes (Act) | ||||
| 0 | 1.020 b ± 0.075 | 0.878 bc ± 0.034 | 0.311 d ± 0.029 | 0.528 cd ± 0.078 |
| 100 | 3.278 a ± 0.374 | 1.000 b ± 0.146 | 0.323 d ± 0.043 | 0.819 bc ± 0.089 |
| Tc Content (mg kg−1 DM of Soil) | Microorganisms | |||||||
|---|---|---|---|---|---|---|---|---|
| Org | Olig | Cop | Act | |||||
| Analysis Day | ||||||||
| 25 | 50 | 25 | 50 | 25 | 50 | 25 | 50 | |
| −G | ||||||||
| 0 | 0.563 e ± 0.049 | 1.405 b ± 0.049 | 0.417 b ± 0.008 | 0.402 b ± 0.019 | 0.071 bc ± 0.007 | 0.027 c ± 0.009 | 1.429 b ± 0.114 | 1.204 ab ± 0.066 |
| 100 | 1.120 c ± 0.016 | 3.345 a ± 0.069 | 0.018 d ± 0.005 | 0.342 ab ± 0.027 | 0.147 a–c ± 0.057 | 0.188 ab ± 0.022 | 5.194 a ± 0.547 | 1.222 ab ± 0.049 |
| +G | ||||||||
| 0 | 0.459 f ± 0.025 | 0.965 d ± 0.029 | 0.375 ab ± 0.038 | 0.289 ab ± 0.039 | 0.150 a–c ± 0.048 | 0.042 c ± 0.012 | 0.576 c ± 0.095 | 0.793 ab ± 0.093 |
| 100 | 0.229 g ± 0.039 | 1.198 c ± 0.015 | 1.583 a ± 0.045 | 0.194 cd ± 0.018 | 0.224 a ± 0.028 | 0.148 a–c ± 0.038 | 0.916 ab ± 0.098 | 1.021 ab ± 0.101 |
| The Dose of Grass Compost in g C kg−1 DM Soil | −Zm | +Zm | ||
|---|---|---|---|---|
| Analysis Day | ||||
| 25 | 50 | 25 | 50 | |
| Dehydrogenases | ||||
| 0 | −0.087 b ± 0.016 | −0.040 ab ± 0.008 | −0.074 b ± 0.018 | −0.245 d ± 0.021 |
| 4 | −0.004 a ± 0.001 | −0.061 ab ± 0.009 | −0.078 b ± 0.016 | −0.170 c ± 0.013 |
| Catalase | ||||
| 0 | −0.042 b ± 0.010 | 0.047 a ± 0.010 | −0.089 c ± 0.014 | 0.051 a ± 0.002 |
| 4 | 0.016 a ± 0.004 | −0.054 bc ± 0.012 | −0.037 b ± 0.010 | −0.026 b ± 0.004 |
| Tc Content (mg kg−1 DM of Soil) | −Zm | +Zm | ||
|---|---|---|---|---|
| Analysis Day | ||||
| 25 | 50 | 25 | 50 | |
| Dehydrogenases | ||||
| 0 | 0.061 d ± 0.053 | 0.046 d ± 0.005 | 0.043 d ± 0.011 | 0.583 b ± 0.014 |
| 100 | 0.158 c ± 0.008 | 0.022 d ± 0.033 | 0.038 d ± 0.018 | 0.742 a ± 0.028 |
| Catalase | ||||
| 0 | 0.076 bc ± 0.005 | 0.156 a ± 0.005 | 0.081 bc ± 0.028 | 0.094 b ± 0.005 |
| 100 | 0.141 a ± 0.015 | 0.045 cd ± 0.013 | 0.141 a ± 0.015 | 0.014 d ± 0.012 |
| Tc Content (mg kg−1 DM of Soil) | Dehydrogenases | Catalase | ||
|---|---|---|---|---|
| Analysis Day | ||||
| 25 | 50 | 25 | 50 | |
| −G | ||||
| 0 | 0.237 ef ± 0.007 | 0.982 c ± 0.055 | 0.051 ab ± 0.003 | 0.078 a ± 0.014 |
| 100 | 0.254 e ± 0.025 | 0.558 d ± 0.040 | 0.000 c ± 0.003 | 0.082 a ± 0.003 |
| +G | ||||
| 0 | 0.215 ef ± 0.012 | 2.000 a ± 0.026 | 0.055 ab ± 0.007 | 0.020 bc ± 0.003 |
| 100 | 0.124 f ± 0.019 | 1.654 b ± 0.043 | 0.000 c ± 0.003 | 0.050 ab ± 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyszkowska, J.; Borowik, A.; Kucharski, J. The Role of Grass Compost and Zea Mays in Alleviating Toxic Effects of Tetracycline on the Soil Bacteria Community. Int. J. Environ. Res. Public Health 2022, 19, 7357. https://doi.org/10.3390/ijerph19127357
Wyszkowska J, Borowik A, Kucharski J. The Role of Grass Compost and Zea Mays in Alleviating Toxic Effects of Tetracycline on the Soil Bacteria Community. International Journal of Environmental Research and Public Health. 2022; 19(12):7357. https://doi.org/10.3390/ijerph19127357
Chicago/Turabian StyleWyszkowska, Jadwiga, Agata Borowik, and Jan Kucharski. 2022. "The Role of Grass Compost and Zea Mays in Alleviating Toxic Effects of Tetracycline on the Soil Bacteria Community" International Journal of Environmental Research and Public Health 19, no. 12: 7357. https://doi.org/10.3390/ijerph19127357
APA StyleWyszkowska, J., Borowik, A., & Kucharski, J. (2022). The Role of Grass Compost and Zea Mays in Alleviating Toxic Effects of Tetracycline on the Soil Bacteria Community. International Journal of Environmental Research and Public Health, 19(12), 7357. https://doi.org/10.3390/ijerph19127357