
Recurrent Plasmodium vivax Cases of Both Short and Long Latency Increased with Transmission Intensity and Were Distributed Year-Round in the Most Affected Municipalities of the RACCN, Nicaragua, 2013–2018


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Study Area
2.2. Data Cleaning and Analysis
3. Results
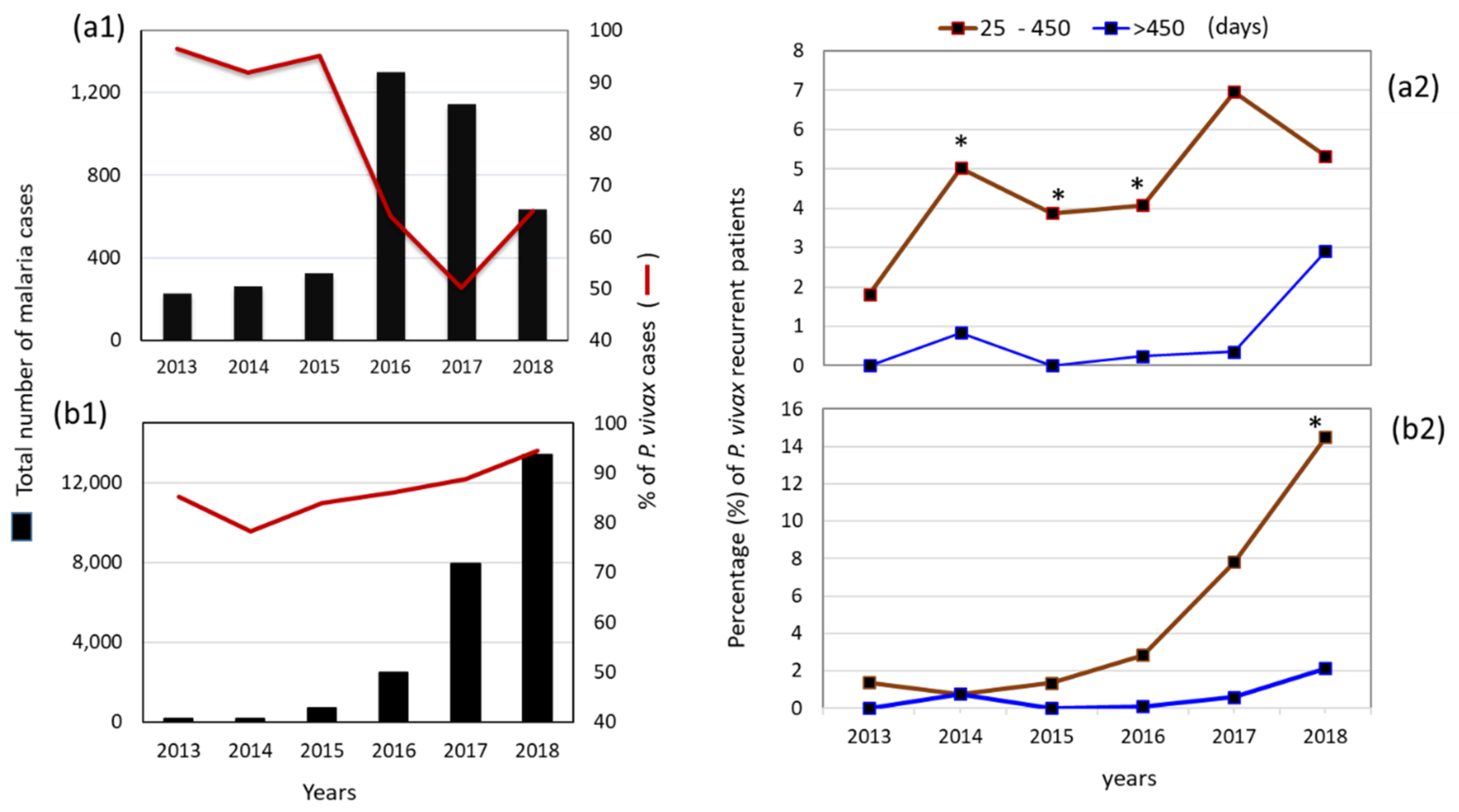
3.1. Malaria Cases per Species per Year at a National Level
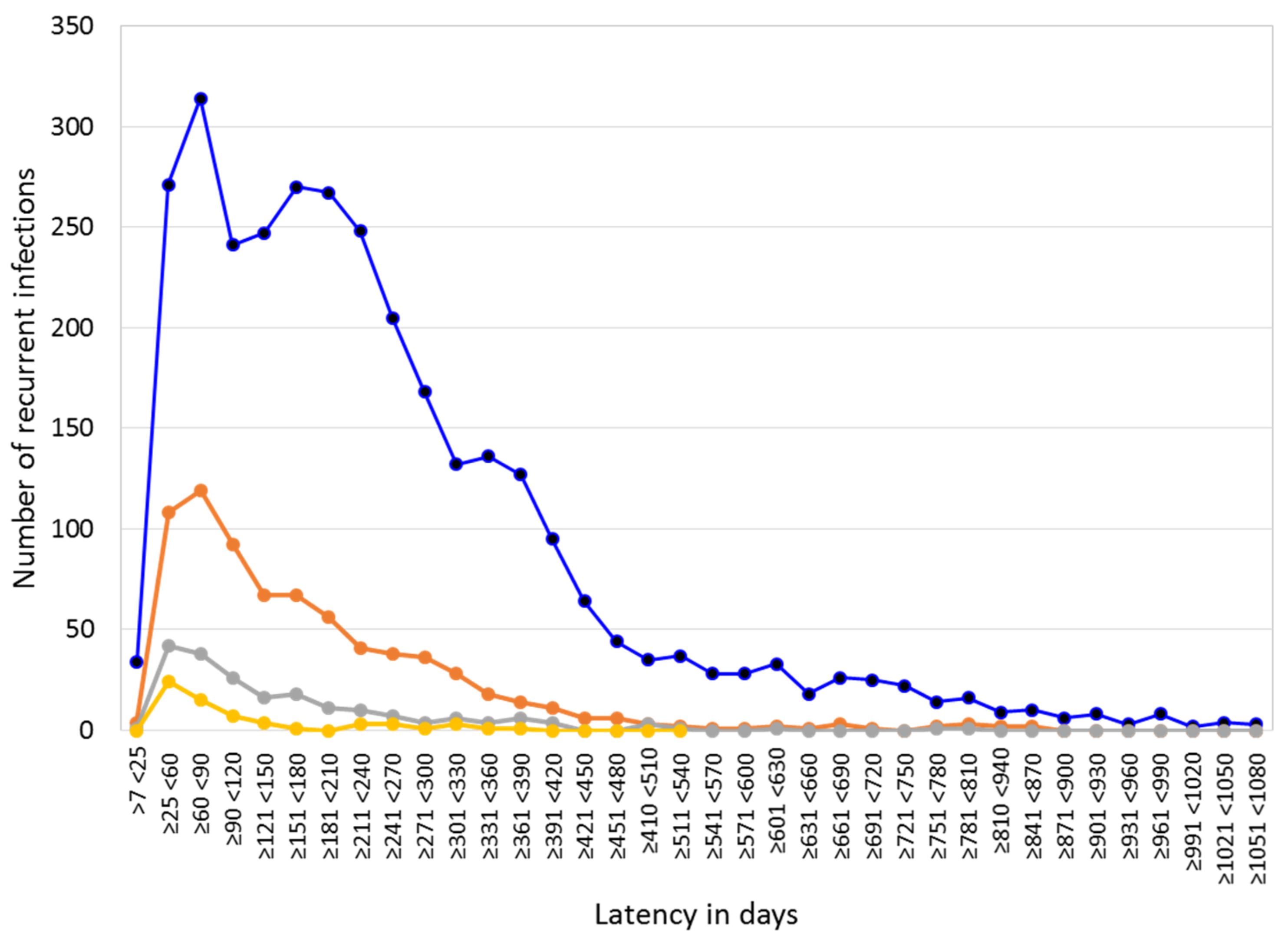
3.2. Patients with Recurrent P. vivax Infections Nationwide
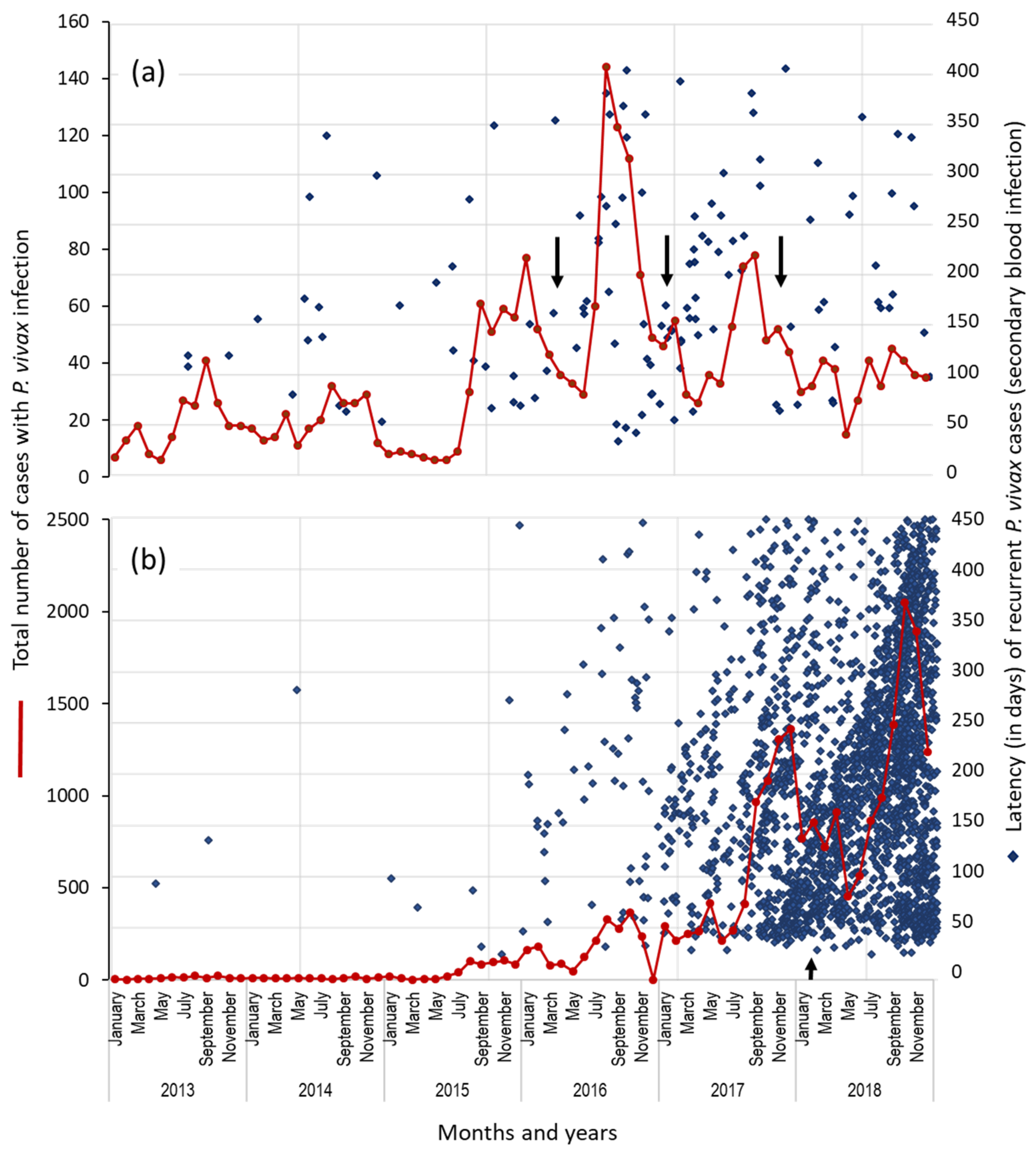
3.3. Proportion and Latency of P. vivax Recurrent Cases in Municipalities: Puerto Cabezas and Rosita, RACCN (2013–2018)
3.4. Temporal Distribution of First Recurrent Episodes (by the Latency) and Total P. vivax Infections in Rosita and Puerto Cabezas, RACCN
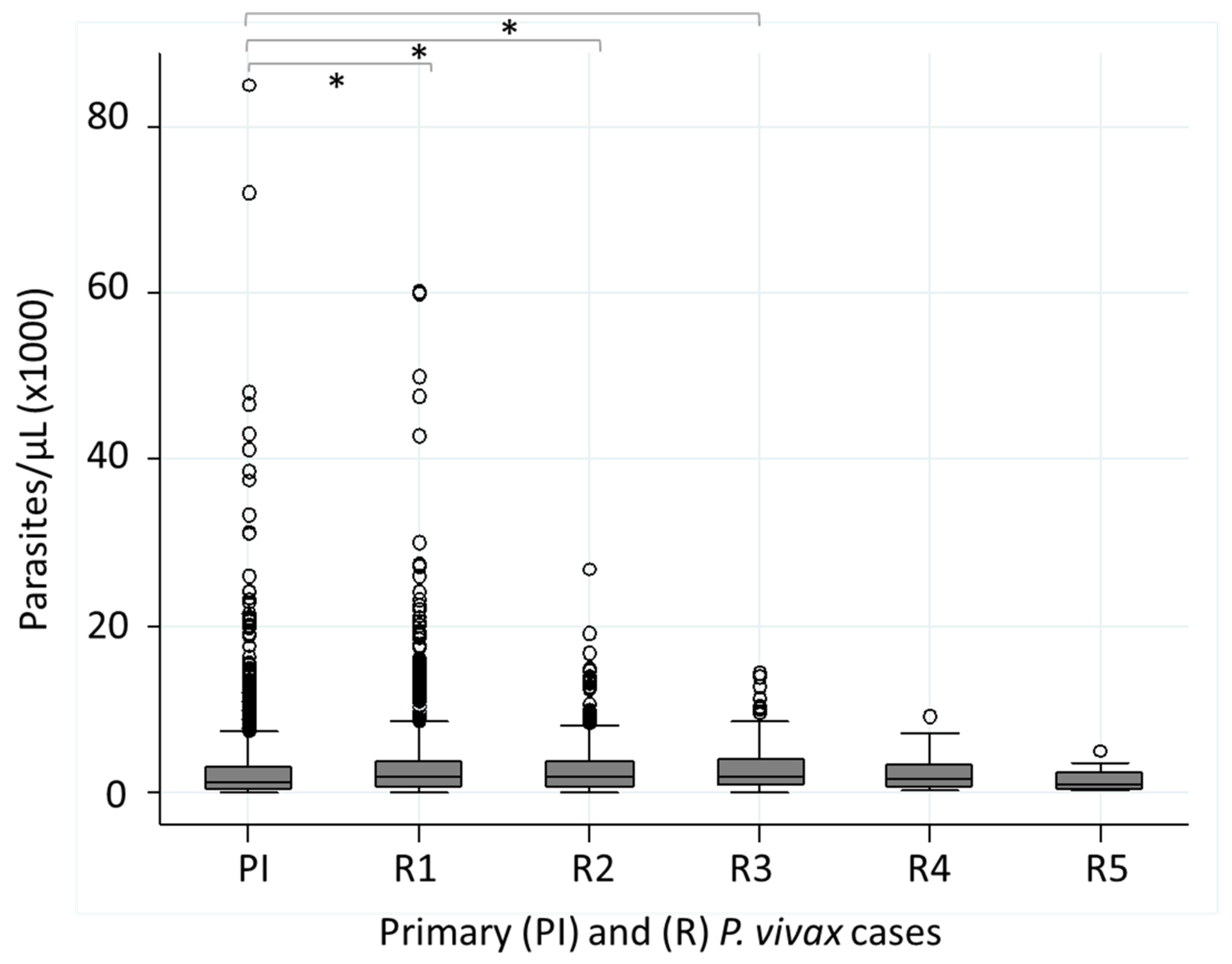
3.5. Parasitemia (Parasites/µL) in Patients with Recurrent P. vivax Infections
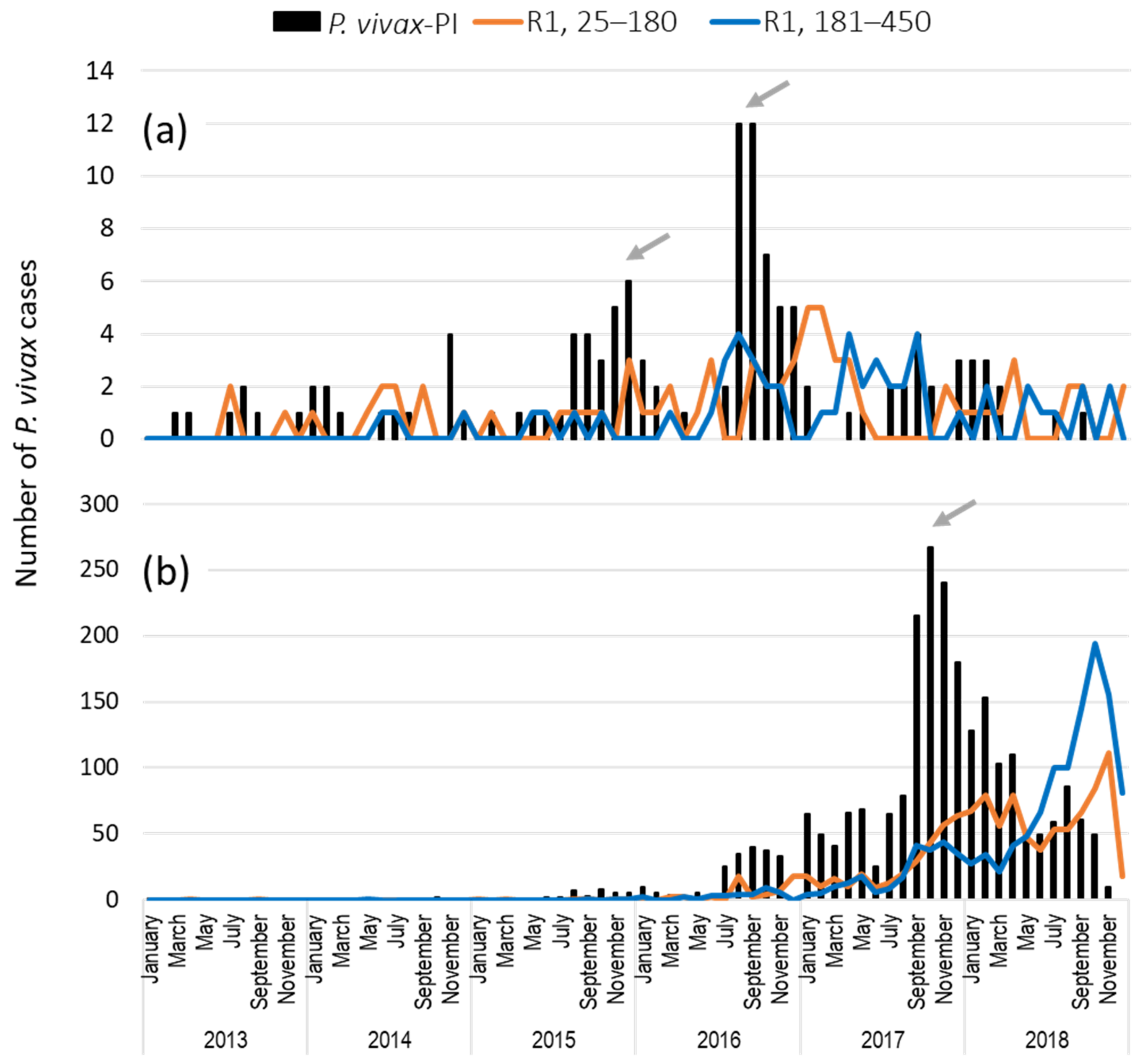
3.6. Comparison of Recurrent P. vivax and P. falciparum Infections (25–450 Days) in RACCN Municipalities: Puerto Cabezas and Rosita
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lover, A.A.; Baird, J.K.; Gosling, R.; Price, R.N. Malaria Elimination: Time to Target All Species. Am. J. Trop. Med. Hyg. 2018, 99, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Battle, K.E.; Karhunen, M.S.; Bhatt, S.; Gething, P.W.; Howes, R.E.; Golding, N.; Van Boeckel, T.P.; Messina, J.P.; Shanks, G.D.; Smith, D.L.; et al. Geographical variation in Plasmodium vivax relapse. Malar. J. 2014, 13, 144. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Treatment of Malaria. 2015. Available online: https://www.ncbi.nlm.nih.gov/books/NBK294440/ (accessed on 16 May 2022).
- Howes, R.E.; Battle, K.E.; Mendis, K.N.; Smith, D.L.; Cibulskis, R.E.; Baird, J.K.; Hay, S.I. Global Epidemiology of Plasmodium vivax. Am. J. Trop. Med. Hyg. 2016, 95, 15–34. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for Malaria. Available online: https://apps.who.int/iris/handle/10665/343751 (accessed on 16 May 2022).
- Cedillos, R.A.; Warren, M.; Jeffery, G.M. Field evaluation of primaquine in the control of Plasmodium vivax. Am. J. Trop. Med. Hyg. 1978, 27, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Mason, J. Patterns of Plasmodium vivax recurrence in a high-incidence coastal area of El Salvador, C.A. Am. J. Trop. Med. Hyg. 1975, 24, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ceron, L.; Mu, J.; Santillan, F.; Joy, D.; Sandoval, M.A.; Camas, G.; Su, X.; Choy, E.V.; Torreblanca, R. Molecular and epidemiological characterization of Plasmodium vivax recurrent infections in southern Mexico. Parasites Vectors 2013, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ceron, L.; Rodriguez, M.H.; Sandoval, M.A.; Santillan, F.; Galindo-Virgen, S.; Betanzos, A.F.; Rosales, A.F.; Palomeque, O.L. Effectiveness of combined chloroquine and primaquine treatment in 14 days versus intermittent single dose regimen, in an open, non-randomized, clinical trial, to eliminate Plasmodium vivax in southern Mexico. Malar. J. 2015, 14, 426. [Google Scholar] [CrossRef][Green Version]
- Contacos, P.G.; Collins, W.E.; Jeffery, G.M.; Krotoski, W.A.; Howard, W.A. Studies on the characterization of Plasmodium vivax strains from Central America. Am. J. Trop. Med. Hyg. 1972, 21, 707–712. [Google Scholar] [CrossRef]
- WHO. World Malaria Report. 2021. Available online: https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2021 (accessed on 16 May 2022).
- WHO. World Malaria Report. 2019. Available online: https://www.who.int/publications/i/item/9789241565721 (accessed on 16 May 2022).
- PAHO. Report on the Situation of Malaria in the Americas. 2017. Available online: https://www3.paho.org/hq/index.php?option=com_docman&view=download&category_slug=datos-estadisticos-mapas-8110&alias=48336-situation-of-malaria-in-the-region-of-the-americas-2017-1&Itemid=270&lang=es (accessed on 16 May 2022).
- BANCO MUNDIAL. Población Total Nicaragua. Available online: https://datos.bancomundial.org/indicador/SP.POP.TOTL?locations=NI (accessed on 16 May 2022).
- Gobierno de Nicaragua. Perfil Demográfico Nicaragua. 2020. Available online: https://pronicaragua.gob.ni/media/publications/Perfil_Demografico_2020_PWsyOuB.pdf (accessed on 16 May 2022).
- Plan Nacional de Nicaragua en el Marco del Plan Mesoamericano Para Mejorar el Control de la Malaria Hacia su Eliminación. 2015. Available online: http://www.proyectomesoamerica.org:8088/smsp/phocadownload/Institucional/PlanesNacionales/PNMalaria/NIC%20PN%20Malaria.pdf (accessed on 16 May 2022).
- Biología y Ecología de Anopheles albimanus Wiedemann en Centroamérica. Bol. Oficina Sanit. Panam. 1996, 121, 32.
- Sinka, M.E.; Rubio-Palis, Y.; Manguin, S.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Van Boeckel, T.; Kabaria, C.W.; Harbach, R.E.; Hay, S.I. The dominant Anopheles vectors of human malaria in the Americas: Occurrence data, distribution maps and bionomic precis. Parasites Vectors 2010, 3, 72. [Google Scholar] [CrossRef]
- Sequeira, M.E.H.; Amador, J.J.; Domingo, G.; Quintanilla, M.; de los Santos, T. Malaria in Nicaragua: A Review of Control Status, Trends, and Needs. Available online: https://path.azureedge.net/media/documents/TS_nicaragua_malaria_rpt.pdf (accessed on 16 May 2022).
- Covell, G. Relationship between malarial parasitaemia and symptoms of the disease: A review of the literature. Bull. World Health Organ. 1960, 22, 605–619. [Google Scholar] [PubMed]
- Stepniewska, K.; Taylor, W.R.; Mayxay, M.; Price, R.; Smithuis, F.; Guthmann, J.P.; Barnes, K.; Myint, H.Y.; Adjuik, M.; Olliaro, P.; et al. In vivo assessment of drug efficacy against Plasmodium falciparum malaria: Duration of follow-up. Antimicrob. Agents. Chemother. 2004, 48, 4271–4280. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Galappaththy, G.N.; Tharyan, P.; Kirubakaran, R. Primaquine for preventing relapse in people with Plasmodium vivax malaria treated with chloroquine. Cochrane Database Syst. Rev. 2013, 10, CD004389. [Google Scholar] [CrossRef]
- Kitchener, S.; Nasveld, P.; Bennett, S.; Torresi, J. Adequate primaquine for vivax malaria. J. Travel. Med. 2005, 12, 133–135. [Google Scholar] [CrossRef][Green Version]
- Carmona Fonseca, J. Primaquine and relapses of Plasmodium vivax. Meta analysis of controlled clinical trials. Rev. Bras. Epidemiol. 2015, 18, 174–193. [Google Scholar] [CrossRef]
- Ministerio de Salud. Esquema Nacional de Tratamiento de Malaria. 2014. Available online: https://www.paho.org/es/documentos/esquema-nacional-tratamiento-malaria-miniosterio-salud-nicaragua-2014 (accessed on 16 May 2022).
- Rodriguez, M.H.; Betanzos-Reyes, A.F.; Hernandez-Avila, J.E.; Mendez-Galvan, J.F.; Danis-Lozano, R.; Altamirano-Jimenez, A. The participation of secondary clinical episodes in the epidemiology of vivax malaria during pre- and post-implementation of focal control in the state of Oaxaca, Mexico. Am. J. Trop. Med. Hyg. 2009, 80, 889–895. [Google Scholar] [CrossRef]
- Hurtado, L.A.; Rigg, C.A.; Calzada, J.E.; Dutary, S.; Bernal, D.; Koo, S.I.; Chaves, L.F. Population Dynamics of Anopheles albimanus (Diptera: Culicidae) at Ipeti-Guna, a Village in a Region Targeted for Malaria Elimination in Panama. Insects 2018, 9, 164. [Google Scholar] [CrossRef]
- Haghdoost, A.A.; Mazhari, S.; Bahaadini, K. Estimating the relapse risk of Plasmodium vivax in Iran under national chemotherapy scheme using a novel method. J. Vector. Borne Dis. 2006, 43, 168–172. [Google Scholar]
- Taylor, W.R.; Hoglund, R.M.; Peerawaranun, P.; Nguyen, T.N.; Hien, T.T.; Tarantola, A.; von Seidlein, L.; Tripura, R.; Peto, T.J.; Dondorp, A.M.; et al. Development of weight and age-based dosing of daily primaquine for radical cure of vivax malaria. Malar. J. 2021, 20, 366. [Google Scholar] [CrossRef]
- Mac Donald-Ottevanger, M.S.; Adhin, M.R.; Jitan, J.K.; Bretas, G.; Vreden, S.G. Primaquine double dose for 7 days is inferior to single-dose treatment for 14 days in preventing Plasmodium vivax recurrent episodes in Suriname. Infect. Drug Resist. 2018, 11, 3–8. [Google Scholar] [CrossRef]
- Orjuela-Sanchez, P.; da Silva, N.S.; da Silva-Nunes, M.; Ferreira, M.U. Recurrent parasitemias and population dynamics of Plasmodium vivax polymorphisms in rural Amazonia. Am. J. Trop. Med. Hyg. 2009, 81, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.; Rodrigues, E.; Ribatski-Silva, D.; Teixeira, L.; Ferreira, A.; Fernandes, C. Factors associated with recurrent Plasmodium vivax malaria in Porto Velho, Rondônia State, Brazil, 2009. Cad. Saude Publica 2014, 30, 1403–1417. [Google Scholar]
- Daher, A.; Silva, J.; Stevens, A.; Marchesini, P.; Fontes, C.J.; Ter Kuile, F.O.; Lalloo, D.G. Evaluation of Plasmodium vivax malaria recurrence in Brazil. Malar. J. 2019, 18, 18. [Google Scholar] [CrossRef] [PubMed]
- PAHO Malaria Technical Advisory Group. Strategies to Decrease Relapses in P. vivax Cases. Available online: https://www3.paho.org/hq/index.php?option=com_docman&view=download&slug=malaria-technical-group-session-6-strategy-for-reducing-the-relapses-of-p-vivax-for-the-control-and-elimination-of-malaria-in-the-americas&Itemid=270&lang=es (accessed on 16 May 2022).
- Mueller, I.; Galinski, M.R.; Tsuboi, T.; Arevalo-Herrera, M.; Collins, W.E.; King, C.L. Natural acquisition of immunity to Plasmodium vivax: Epidemiological observations and potential targets. Adv. Parasitol. 2013, 81, 77–131. [Google Scholar] [CrossRef]
- Michon, P.; Cole-Tobian, J.L.; Dabod, E.; Schoepflin, S.; Igu, J.; Susapu, M.; Tarongka, N.; Zimmerman, P.A.; Reeder, J.C.; Beeson, J.G.; et al. The risk of malarial infections and disease in Papua New Guinean children. Am. J. Trop. Med. Hyg. 2007, 76, 997–1008. [Google Scholar] [CrossRef]
- Collins, W.E.; Jeffery, G.M.; Roberts, J.M. A retrospective examination of reinfection of humans with Plasmodium vivax. Am. J. Trop. Med. Hyg. 2004, 70, 642–644. [Google Scholar] [CrossRef]
- Cucunuba, Z.M.; Guerra, A.P.; Rahirant, S.J.; Rivera, J.A.; Cortes, L.J.; Nicholls, R.S. Asymptomatic Plasmodium spp. infection in Tierralta, Colombia. Mem. Inst. Oswaldo Cruz. 2008, 103, 668–673. [Google Scholar] [CrossRef]
- Camargo, E.P.; Alves, F.; Pereira da Silva, L.H. Symptomless Plasmodium vivax infections in native Amazonians. Lancet 1999, 353, 1415–1416. [Google Scholar] [CrossRef]
- Lin, J.T.; Saunders, D.L.; Meshnick, S.R. The role of submicroscopic parasitemia in malaria transmission: What is the evidence? Trends. Parasitol. 2014, 30, 183–190. [Google Scholar] [CrossRef]
- Silva, R.G.; Nunes, J.E.; Canduri, F.; Borges, J.C.; Gava, L.M.; Moreno, F.B.; Basso, L.A.; Santos, D.S. Purine nucleoside phosphorylase: A potential target for the development of drugs to treat T-cell- and apicomplexan parasite-mediated diseases. Curr. Drug Targets 2007, 8, 413–422. [Google Scholar] [CrossRef]
- Gutierrez, S.; Gonzalez-Ceron, L.; Montoya, A.; Sandoval, M.A.; Torres, M.E.; Cerritos, R. Genetic structure of Plasmodium vivax in Nicaragua, a country in the control phase, based on the carboxyl terminal region of the merozoite surface protein-1. Infect. Genet. Evol. 2016, 40, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ceron, L.; Montoya, A.; Corzo-Gomez, J.C.; Cerritos, R.; Santillan, F.; Sandoval, M.A. Genetic diversity and natural selection of Plasmodium vivax multi-drug resistant gene (pvmdr1) in Mesoamerica. Malar. J. 2017, 16, 261. [Google Scholar] [CrossRef] [PubMed]
- Douglas, N.M.; Nosten, F.; Ashley, E.A.; Phaiphun, L.; van Vugt, M.; Singhasivanon, P.; White, N.J.; Price, R.N. Plasmodium vivax recurrence following falciparum and mixed species malaria: Risk factors and effect of antimalarial kinetics. Clin. Infect. Dis. 2011, 52, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Ashley, E.A.; Phyo, A.P.; Carrara, V.I.; Tun, K.M.; Nosten, F.; Smithuis, F.; White, N.J. Plasmodium vivax Relapse Rates Following Plasmodium falciparum Malaria Reflect Previous Transmission Intensity. J. Infect. Dis. 2019, 220, 100–104. [Google Scholar] [CrossRef]
- Commons, R.J.; Simpson, J.A.; Watson, J.; White, N.J.; Price, R.N. Estimating the Proportion of Plasmodium vivax Recurrences Caused by Relapse: A Systematic Review and Meta-Analysis. Am. J. Trop. Med. Hyg. 2020, 103, 1094–1099. [Google Scholar] [CrossRef]
- Sagara, I.; Sangare, D.; Dolo, G.; Guindo, A.; Sissoko, M.; Sogoba, M.; Niambele, M.B.; Yalcoue, D.; Kaslow, D.C.; Dicko, A.; et al. A high malaria reinfection rate in children and young adults living under a low entomological inoculation rate in a periurban area of Bamako, Mali. Am. J. Trop. Med. Hyg. 2002, 66, 310–313. [Google Scholar] [CrossRef]
- Larranaga, N.; Mejia, R.E.; Hormaza, J.I.; Montoya, A.; Soto, A.; Fontecha, G.A. Genetic structure of Plasmodium falciparum populations across the Honduras-Nicaragua border. Malar. J. 2013, 12, 354. [Google Scholar] [CrossRef]
- Rodriguez, M.H.; Betanzos-Reyes, A.F. Plan to improve malaria control towards its elimination in Mesoamerica. Salud. Publica Mex. 2011, 53 (Suppl. S3), S333–S348. [Google Scholar]
- Balieiro, A.A.S.; Siqueira, A.M.; Melo, G.C.; Monteiro, W.M.; Sampaio, V.S.; Mueller, I.; Lacerda, M.V.G.; Villela, D.A.M. Short-Time Recurrences of Plasmodium vivax Malaria as a Public Health Proxy for Chloroquine-Resistance Surveillance: A Spatio-Temporal Study in the Brazilian Amazon. Int. J. Environ. Res. Public Health 2021, 18, 5061. [Google Scholar] [CrossRef]
- Llanos-Cuentas, A.; Lacerda, M.V.G.; Hien, T.T.; Velez, I.D.; Namaik-Larp, C.; Chu, C.S.; Villegas, M.F.; Val, F.; Monteiro, W.M.; Brito, M.A.M.; et al. Tafenoquine versus Primaquine to Prevent Relapse of Plasmodium vivax Malaria. N. Engl. J. Med. 2019, 380, 229–241. [Google Scholar] [CrossRef]
- WHO. New Opportunities to Prevent Plasmodium vivax Malaria Relapse. Available online: https://www.who.int/news/item/25-02-2019-new-opportunities-to-prevent-p-vivax-malaria-relapse#:~:text=For%20the%20treatment%20of%20P,a%20full%202%2Dweek%20period (accessed on 16 May 2022).
- Kondrashin, A.; Baranova, A.M.; Ashley, E.A.; Recht, J.; White, N.J.; Sergiev, V.P. Mass primaquine treatment to eliminate vivax malaria: Lessons from the past. Malar. J. 2014, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Mendis, K. Mass drug administration should be implemented as a tool to accelerate elimination: Against. Malar. J. 2019, 18, 279. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmodium Species | Years (Number of Cases) | Total | |||||
|---|---|---|---|---|---|---|---|
| 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | ||
| P. vivax | 950 | 924 | 1880 | 4343 | 8959 | 14,501 | 31,557 |
| P. falciparum | 211 | 139 | 320 | 1104 | 2033 | 1319 | 5126 |
| Mixed Pv–Pf | 0 | 0 | 3 | 17 | 35 | 49 | 104 |
| Total | 1161 | 1063 | 2203 | 5464 | 11,027 | 15,869 | 36,787 |
| % P. vivax | 81.8 | 86.9 | 85.3 | 79.5 | 81.2 | 91.4 | 85.8 |
| % P. falciparum | 18.2 | 13.1 | 14.5 | 20.3 | 18.4 | 8.3 | 13.9 |
| % Pv–Pf | 0 | 0 | 0.1 | 0.3 | 0.3 | 0.3 | 0.3 |
| NI | 4 | 2 | 0 | 70 | 0 | 1 | 77 |
| Number of P. vivax Infections | Nationwide (%) n = 3163 Patients | RACCN, Puerto Cabezas (%) n = 2789 Patients | RACCN, Rosita (%) n = 139 Patients |
|---|---|---|---|
| 2 | 77.2 | 75.3 | 90.7 |
| 3 | 16.7 | 17.9 | 7.2 |
| 4 | 4 | 4.4 | 2.1 |
| 5 | 1.2 | 1.3 | 0.7 |
| 6 | 0.6 | 0.7 | - |
| 7 | 0.09 | 0.1 | - |
| (a) | ||||||
| Parameters | Season/Period | Statistics (Comparing All Groups) | ||||
| Rain | Dry | Rain | Dry | Rain | ||
| June–November 2015 | December 2015–May 2016 | June–November 2016 | December 2016–May 2017 | June–November 2017 | ||
| Number of P. vivax cases | 216 | 302 | 533 | 241 | 338 | χ2 = 9.6, p = 0.046 |
| Linked to recurrent patients (%) | 8.3 | 3.9 | 7 | 3 | 3.7 | |
| Number and latency of R1: | χ2 = 11.04, p = 0.026 | |||||
| n | 6 | 9 | 21 | 28 | 13 | |
| Short: 25–180 days (%) | 66.6 | 88.8 | 33.3 | 71.4 | 15.3 | |
| Long: 181–450 days (%) | 33.3 | 11.2 | 66.7 | 28.6 | 84.7 | |
| Statistics: | χ2 = 7.7, p = 0.005 | χ2 = 16.6, p < 0.0001 | ||||
| χ2 = 1.1, p = 0.69 | χ2 = 9.03, p = 0.007 | |||||
| (b) | ||||||
| Parameters | Season/Period | Statistics (Comparing All Groups) | ||||
| Dry | Rain | Dry | Rain | |||
| December 2016–May 2017 | June–November 2017 | December 2017–May 2018 | June–November 2018 | |||
| Number of P. vivax cases | 1439 | 4251 | 5062 | n.d. | χ2 = 76.7, p < 0.0001 | |
| Linked to recurrent patients (%) | 20 | 20 | 14 * | n.d. | ||
| Number and latency of R1: | χ2 = 211.8, p < 0.0001 | |||||
| n | 142 | 325 | 598 | 1111 | ||
| Short: 25–180 days (%) | 64.7 | 52.6 | 65 | 35 | ||
| Long: 181–450 days (%) | 35.3 | 47.4 | 35 | 65 | ||
| Statistics: | χ2 = 5.9, p = 0.014 | χ2 = 132.2, p < 0.0001 | ||||
| χ2 = 10.1, p = 0.001 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto, A.M.; González-Cerón, L.; Santillán-Valenzuela, F.; Parrales, M.E.; Montoya, A. Recurrent Plasmodium vivax Cases of Both Short and Long Latency Increased with Transmission Intensity and Were Distributed Year-Round in the Most Affected Municipalities of the RACCN, Nicaragua, 2013–2018. Int. J. Environ. Res. Public Health 2022, 19, 6195. https://doi.org/10.3390/ijerph19106195
Soto AM, González-Cerón L, Santillán-Valenzuela F, Parrales ME, Montoya A. Recurrent Plasmodium vivax Cases of Both Short and Long Latency Increased with Transmission Intensity and Were Distributed Year-Round in the Most Affected Municipalities of the RACCN, Nicaragua, 2013–2018. International Journal of Environmental Research and Public Health. 2022; 19(10):6195. https://doi.org/10.3390/ijerph19106195
Chicago/Turabian StyleSoto, Aida M., Lilia González-Cerón, Frida Santillán-Valenzuela, María E. Parrales, and Alberto Montoya. 2022. "Recurrent Plasmodium vivax Cases of Both Short and Long Latency Increased with Transmission Intensity and Were Distributed Year-Round in the Most Affected Municipalities of the RACCN, Nicaragua, 2013–2018" International Journal of Environmental Research and Public Health 19, no. 10: 6195. https://doi.org/10.3390/ijerph19106195
APA StyleSoto, A. M., González-Cerón, L., Santillán-Valenzuela, F., Parrales, M. E., & Montoya, A. (2022). Recurrent Plasmodium vivax Cases of Both Short and Long Latency Increased with Transmission Intensity and Were Distributed Year-Round in the Most Affected Municipalities of the RACCN, Nicaragua, 2013–2018. International Journal of Environmental Research and Public Health, 19(10), 6195. https://doi.org/10.3390/ijerph19106195