Abstract
Over the past several decades, much attention has been focused on the dispersal of aquatic nonindigenous species via ballast tanks of shipping vessels worldwide. The recently reclassified dinoflagellate Pseudocochlodinium profundisulcus (previously identified as Cochlodinium sp., Cochlodinium geminatum, or Polykrikos geminatus) was not reported in China until 2006. However, algal blooming events caused by this organism have been reported almost every year since then in the Pearl River Estuary and its adjacent areas in China. Whether P. profundisulcus is an indigenous or an invasive species has thus become an ecological question of great scientific and practical significance. In this study, we collected the sediments from ballast tanks of ships arriving in the ports of China and North America and characterized dinoflagellate resting cysts via a combined approach. We germinated two dark brownish cysts from the tank of an international ship (Vessel A) arriving at the Jiangyin Port (China) into vegetative cells and identified them as P. profundisulcus by light and scanning electron microscopy and phylogenetic analyses for partial LSU rDNA sequences. We also identified P. profundisulcus cyst from the ballast tank sediment of a ship (Vessel B) arriving in the port of North America via single-cyst PCR and cloning sequencing, which indicated that this species could be transported as resting cyst via ship. Since phylogenetic analyses based on partial LSU rDNA sequences could not differentiate all sequences among our cysts from those deposited in the NCBI database into sub-groups, all populations from China, Australia, Japan, and the original sources from which the cysts in the two vessels arrived in China and North America were carried over appeared to share a very recent common ancestor, and the species may have experienced a worldwide expansion recently. These results indicate that P. profundisulcus cysts may have been extensively transferred to many regions of the world via ships’ ballast tank sediments. While our work provides an exemplary case for both the feasibility and complexity (in tracking the source) of the bio-invasion risk via the transport of live resting cysts by ship’s ballast tanks, it also points out an orientation for future investigation.
1. Introduction
A ship’s ballast tanks carrying ballast water and sediments were proved to be responsible for the dispersal of toxic phytoplankton worldwide [1,2,3,4,5,6,7]. Recently, a large number of studies have focused on the introduction potency of ballast sediments, which can be accumulated in the bottom of the tanks and are difficult to be washed off [8,9,10]. For instance, tens to hundreds of tons of fine mud with a mean particle size of <20 μm were found in tanks, with the thickness ranging from several centimeters to >30 cm [11]. The ballast sediments provide a suitable environment for dinoflagellates resting cysts, which could remain and maintain viable in sediments for years [12,13,14,15,16]. Hence, there were usually more dinoflagellate cells (cysts) within ballast sediments compared to that in ballast water, e.g., >300 million Alexandrium cysts existing in sediments of one single ballast tank in a ship entering an Australian port [1], which implied the great threat of ballast tank sediments to marine ecosystem, human health, and economic development [1,10,17,18].
Gymnodiniales are the major order of dinoflagellates, many species of which have ecological significance due to the ability to form harmful algal blooms (HABs). Among them, Pseudocochlodinium profundisulcus (reported as Cochlodinium sp., Cochlodinium geminatum or Polykrikos geminatus [19]) was firstly reported to form bloom in April 2006 in the coastal water of Zhuhai, Guangdong province, China [20]. Since then, blooms caused by this species have occurred frequently in the South China Sea, whereas blooming areas from several to 300 km2 have the maximum cell density up to 4.13 × 107 cells·L−1 [21,22,23,24,25]. These blooms not only caused severe water discoloration but also led to economic loss, for instance, a small-scale bloom (89 km2) in August 2011 caused a loss of about 3.16 million RMB [24]. Moreover, P. profundisulcus bloom water exhibited a toxic effect on zooplankton Artemia salina [26]. Recently, Dong et al. (2020) demonstrated that retinal (a potential teratogenic agent) existed in cultured P. profundisulcus and also accumulated rapidly during the P. profundisulcus bloom, implying its potential risk to aquatic organisms [23]. Furthermore, P. profundisulcus culture significantly inhibited the growth of co-occurring phytoplankton, including Prorocentrum micans, Heterosigma akashiwo, Karlodinum veneficum, and Rhodomonas salina, which suggests that the allelopathy of P. profundisulcus may play an important role in competing with other phytoplankton and staying dominant throughout blooms [27].
Pseudocochlodinium profundisulcus has been reported in coastal waters of China, Japan, Australia, and other regions [19,28,29,30]. Lan and Gu (2014) reported that P. profundisulcus could produce resting cyst in the Pearl River estuary, implying that cyst germination may provide a source for blooms [28]. Recently, our group observed that the resting cysts of P. profundisulcus in some ship’s ballast sediment samples [10], which suggests this important HABs-causing species in China might be associated with bio-invasion risk. However, whether P. profundisulcus is a local population or an invasive species has become a question of great scientific and practical significance, which was urgent to be studied to understand the mechanism of bloom explosive and transmission better.
In this study, we used a combinatorial approach including morphological observation, resting cyst germination, molecular identification, and phylogenetic analyses to (1) investigate the occurrence of P. profundisulcus resting cysts in ship’s ballast tank arriving in the ports of China and North America, and (2) assess the potential bio-invasion risk of P. profundisulcus via ships’ ballast tank sediments.
2. Materials and Methods
2.1. Sampling and Sediment Collection
Ballast tank sediment sample CN was collected from an empty ballast tank with a capacity of 747.0 m3 from Vessel A arriving at Jiangyin Port, China. Vessel A is a general cargo ship built in 1997. The ballast water log of the studied tank before sampling was present in Table 1. The sediment sample was collected on 10 November 2017 with a sterile trowel and sub-packaged into sterilized tubes, which were stored at 4 °C in darkness until further analysis.

Table 1.
The ballast water log for the studied ballast tank of Vessel A.
A ballast tank sediment sample NA was collected from a double-bottom tank of Vessel B entering Windsor Port (Ontario, Canada) on 26 June 2002. This sample was related to a research program for assessing potential invasion vectors via overseas vessels arriving in the Great Lakes, with the information of ship, tank, and ballasting history provided by Johengen et al. (2005) [31]. In detail, this investigated ship was built in 1986. During the research program, the voyage of this ship was from Tampico (Mexico) to Toledo (USA), Windsor (Canada), and Detroit (USA). During the voyage, it was ballasted at Everglades (USA) with a total ballast capacity of 9000 m3 in May 2002. The sampled tank was not cleaned at the dry dock, and the number of total residuals was about 50 metric tons with sediments <25% when sampling.
2.2. Culture of Pseudocochlodinium Profundisulcus Establishment from Ballast Sediment of Vessel A Collected from Jiangyin Port, China, Light and Scanning Microscopy
Ballast tank sediment sample CN from Vessel A (2 g) was used for the resting cyst germination experiment. Cyst assemblage was concentrated using sodium polytungstate solution [32]. Individual cysts were washed at least three times with sterile f/2 medium [33] and micropipetted to a 24-well culture plate with each well containing 2.5 mL fresh f/2 medium and one single cyst. An antibiotic solution (final concentration 2%; a mixture of 10,000 I.U. penicillin and 10,000 µg·mL−1 streptomycin, Solarbio, Beijing, China) and germanium dioxide (final concentration of 20 mg·L−1) was added into the medium immediately to discourage bacterial and diatom growth. The plate was incubated at 21 ± 1°C with 12:12 h light:dark cycle under a light intensity of 100 µmol photons m−2·s−1. A clonal culture of P. profundisulcus was established through germlings. The living cyst, empty cyst, and vegetative cells were observed with light microscopes (IX73, BX53, Olympus, Tokyo, Japan) and photographed by a DP80 digital camera (Olympus, Tokyo, Japan). Cells at exponential growth stage were fixed with osmium tetroxide (OsO4, 2% final concentration) for 40–50 min, gently filtered onto an 11 μm Millipore nylon membrane, dehydrated in an acetone series (10%, 30%, 50%, 70%, 90%, and 3 times in 100%, each step 15 min), critical point-dried (automated critical point dryer, EM CPD 300, Leica, Vienna, Austria), sputter-coated with gold (Sputter/Carbon Thread, EM ACE200, Leica, Austria), and observed with an S-3400N SEM (Hitachi, Hitachinaka, Japan).
2.3. Identification of Pseudocochlodinium Profundisulcus Cyst from North America
Ballast tank sediment sample NA collected from Vessel B entering North America was used to investigate the presence of P. profundisulcus cyst. Individual cysts were micropipetted from cyst assemblage concentrated using SPT from ballast sediment [32] and washed at least three times with sterile seawater enriched with f/2 medium [33]. A pair of primer (5′-ACCCGCTGAATTTAAGCATA-3′ and 5′-GCTACTACCACCAAGATCTGC-3′) was used for single-cell sequencing. The processes including PCR, cloning, and sequencing were according to Shang et al. (2019) [10].
2.4. DNA Extraction, PCR Amplification, Sequencing, and Phylogenetic Analyses
Total DNA of P. profundisulcus established from the Chinese port was extracted using a plant DNA extraction kit (Tiangen, Beijing, China) following the manufacturer’s protocol and identified by large subunit ribosomal DNA gene sequencing. About 1400 bp of LSU rDNA sequence was amplified using primers of D1R (forward, 5′-ACCCGCTGAATTTAAGCATA-3′) and 28-1483R (reverse, 5′-GCTACTACCACCAAGATCTGC-3′) [34,35]. The PCR reactions and sequencing were conducted according to Hu et al. (2021) [19].
Phylogenetic analyses for collected LSU rDNA sequences were aligned using the default settings of MAFFT v.7.110 [36,37] (Available online: http://mafft.cbrc.jp/alignment/server/; Accessed on 27 September 2021) and modified manually using BioEdit (v7.313) [38]. The program jModelTest 2.1.4 was executed to select the most appropriate model of molecular evolution with Akaike information criterion and GTR + G + I model with gamma-distributed rate variation across sites, and a proportion of invariable sites was chosen as the most appropriate model [39]. Maximum likelihood analysis was conducted with raxmlGUI v1.3.1 using the model GTR + I + G [40,41]. Node support was assessed with 1000 bootstrap replicates. The Bayesian inference analysis was performed by MrBayes 3.2.6 using the best-fitting substitution model (GTR + I + G) [42]. Four independent Markov chain Monte Carlo processes of 10,000,000 generations were conducted. Trees were sampled every 1000 generations. The first 10% of trees were discarded as burn-in. The convergence was assessed by the average standard deviation of split frequencies (less than 0.01). The remaining trees were used to generate a consensus tree, with posterior probabilities of all branches computed via a majority-rule consensus approach. FigTree (v1.4.4) was used to view and edit trees for publication.
3. Results
3.1. Morphology of Resting Cyst and Vegetative Cell of Pseudocochlodinium Profundisulcus
A resting cyst of P. profundisulcus in the Sample NA collected from a double-bottom tank of the Vessel B entering Windsor Port was isolated and observed, which was yellowish-brown, irregularly round with lobed ornaments around the cyst wall (identified as Polykrikos geminatum, see picture in Shang et al. (2019) [10]). In addition, two dark brownish resting cysts were isolated from the tank of Vessel A, arriving at Jiangyin port, China, and successfully germinated into vegetative cells in 7 days. Light and scanning electron microscopy were used to observe the characteristics of one of the resting cysts and the associated vegetative cells and provided the morphological structure for this species (Figure 1). The cysts of P. profundisulcus were about 45 μm in length and subspherical to spherical. The cyst wall was dark brownish in appearance with lobed ornaments (Figure 1a). A yellow to dark brownish accumulation body was present (Figure 1a). The archeopyle was chasmic (Figure 1b). Vegetative cells were roughly subspherical with a slightly conical convex epicone and round hypocone (Figure 1c–f). The nucleus was located near the center of the cell, and each cell contained many yellow-brownish to brown chloroplasts scattering throughout the cell periphery (Figure 1c–f). The cingulum encircled more than one, but less than one and a half turns of the cell body (Figure 1g–i). The apical structure complex was comma-shaped (Figure 1g–i).
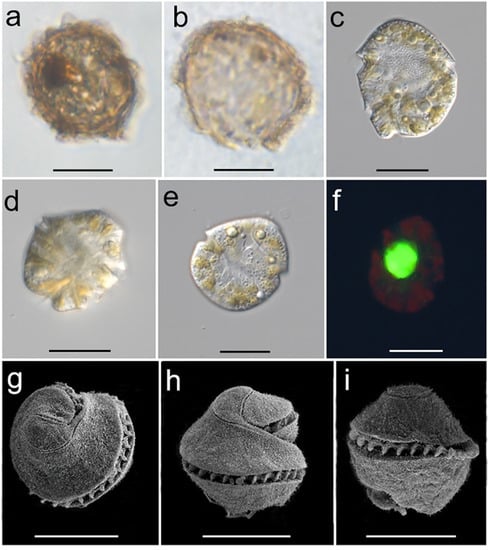
Figure 1.
Light microscopic micrographs of the Pseudocochlodinium profundisulcus resting cyst (a), empty cyst (b), and vegetative cells (c–f); scanning electron microscopic micrographs (g–i) of the vegetative cells; Scale bars = 20 μm.
3.2. Molecular Phylogeny
The accession numbers of LSU rDNA sequences for P. profundisulcus were obtained and deposited in NCBI (accession numbers: OK667806 (from China port) and OK667934 (from North America port)). The sequences were analyzed using the Basic Local Search Tool (BLAST, Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi; Accessed on 25 October 2021) in GenBank. The LSU rDNA sequences (OK667806 and OK667934) were found to be 98.5% and 97.54%, 99.8–99.9% and 99.2–99.5%, and 99.6–99.9% and 98.0%–99.5% identical to three entities in GenBank that were annotated as Cochlodinium cf. geminatum (EF616462), Po. geminatus (KF878934 and JX967270), and P. profundisulcus (MF445291, MG874046, MG874047, MG874048, MW811439, and MW811440)
Phylogenetic analyses using maximum likelihood and Bayesian inference generated similar trees but differed at a few internal nodes (Figure 2). The two entities (OK667806 and OK667934) from ballast sediments formed a coherent clade together with P. profundisulcus (MF445291, MW811439, MW811440, MG874046, MG874047, MG874048, and MG874049, from the Pearl River Estuary, China), a previously deposited entity of Po. geminatus (JX967270, KF878934, from the Pearl River Estuary, China; species name provided in GenBank), and Cochlodinium cf. geminatum (EF616462, from Australia; species name provided in GenBank) with strong support (100/1.0).
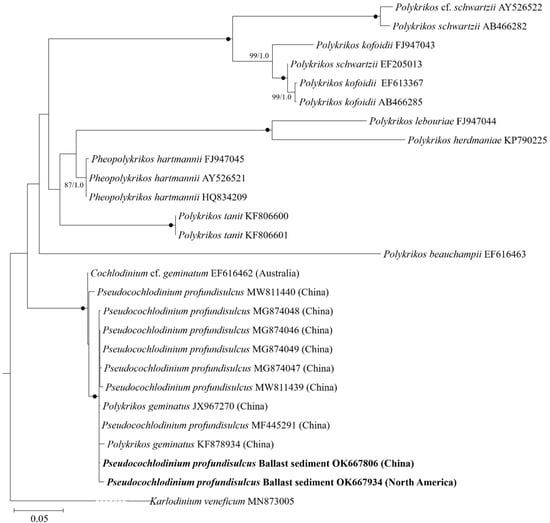
Figure 2.
Phylogenetic tree of the Pseudocochlodinium profundisulcus based on the large subunit rDNA sequences. Karlodinum veneficum was used as an outgroup. Bootstrap values >50% and posterior probabilities (PP) above 0.5 are shown. Black dots (•) indicate maximal support (PP = 1.00 in BI and bootstrap support = 100% in ML, respectively).
4. Discussion
Harmful algal blooms (HABs) are one of the most important marine ecological disasters. In the past few decades, algal blooms have occurred frequently in the world with an increasing trend [43,44,45,46]. Moreover, some HAB-forming species show a trend of spreading to other countries and regions [47,48,49]. In addition to global warming and water eutrophication associated with human activities, phytoplankton spread by ship’s ballast water and sediments is widely regarded as a very important external cause for the increase in HABs [1,49,50,51,52,53,54,55]. As one of the most ecologically important phytoplankton groups, dinoflagellates cause about 75% of the marine HABs events, and many species could produce various toxins [56]. In addition, dinoflagellates are important primary producers in the oceans, especially offshore, and molecular sequencing (e.g., rDNA) results showed that they accounted for about 50% of surface marine protists [57]. A very important biological feature of dinoflagellate is that many dinoflagellates, including these algal bloom species, can form dormant cysts during their life history [13]. The cysts could withstand harsh environment due to the thick cyst walls, allowing them to survive in marine sediments for a long period of time (from a few months to over 100 years) and germinate under suitable environmental conditions that provide seeds for reproduction and even forming blooms [12,13,14,15,16]. This characteristic of cysts has a very important ecological significance that makes them easier to be geographically dispersed through man-made or natural processes, such as the alien introduction by ballast water/sediments and farmed shellfish [1,50,54,55]. Smayda (2007) documented that those species clearly believed to be spread through ship’s ballast tanks to form harmful algal blooms in different places were all cyst- or dormant cell-producing species [54]. In turn, we inferred that the dinoflagellates that produce dormant cysts could, in theory, undergo allochthonous dispersal through the ballast tanks [58]. Since International Maritime Organization (IMO) established and adopted “The International Convention for The Control and Management of Ships Ballast Water Ballast Water and Sediments, 2004” (BWM Convention) at a Diplomatic Conference in London in 2004 and entered into force on 8 September 2017 [59], it is urgent to study the survivability of dinoflagellate cysts in an extreme environment for determining appropriate treatments on ballast sediments.
While the frequency, scale, and degree of harmful algal blooms have increased in China, it is particularly worth noting that new toxic and HABs-forming species have also been continuously uncovered [49]. As an organism that prefers high salinities and has a wide tolerance to temperature [23], P. profundisulcus was initially described as Cochlodinium geminatum Schütt [60] and transferred to the genus Polykrikos as P. geminatus by Qiu et al. (2013) [61]. Since the first record of P. profundisulcus in China in 2006, it has formed HABs frequently in the South China Sea since then [20,21,22,23,24,25]. Until now, Pseudocochlodinium species have had a global distribution [19]. In a previous study, we found 11 OTUs with 935 reads annotated as Cochlodinium sp., Cochlodinium geminatum, or Polykrikos geminatus (reclassified as P. profundisulcus by Hu et al. (2021) [19]) via metabarcoding approach, which implied many P. profundisulcus cysts present in the sediments of the ship’s tanks [10]. In this study, we further confirmed the presence of live resting cysts of P. profundisulcus in sediments of ballast tanks in an international vessel that had visited Brazil, USA, Japan, Australia, and China. As this ship loaded and discharged water in many places as well as in Huangdao Port, China, before our sampling, it was difficult to distinguish whether the cysts were from China or abroad. Moreover, we also identified a P. profundisulcus cyst by single-cyst polymerase chain reaction (PCR), and subsequent sequencing in the ballast tank sediment of an overseas vessel visited Mexico, USA, and Canada, which further indicated that this species could be transported as resting cyst via ship. Since P. profundisulcus has been reported from North America [62], the origin of the cyst in this ship is also a perplexing question.
The characteristic of P. profundisulcus to produce resting cysts could not only help to survive under unfavorable environmental conditions but also capable of enduring long-distance transport (currents, ballast tanks, or transplanted shellfish), which possibly facilitate its ability to colonize and settle in new habitats [2,10,17,54,58]. The initial outbreak and reoccurrence of P. profundisulcus bloom in China were observed near the harbor area where was subject to ship ballast waters [20,63]. In this study, we detected the P. profundisulcus resting cysts in the ship’s ballast tank sediments arriving in the ports of both China and North America. Moreover, we have germinated two P. profundisulcus cysts from one sediment sample (2 g) in the tank of Vessel A. This is important evidence that this toxic and harmful algal bloom-forming species not only exists as resting cysts in ships but also has the ability to reproduce, posing a serious threat to the accepted waters. Since dinoflagellates have the ability to reproduce sexually and form resting cysts, genetic exchange and recombination could happen in a short timescale [64,65,66]. Once the cysts in ballast tanks are de-ballasted into the accepted waters, they may hatch continuously in suitable growth conditions and supply novel genotypes to the indigenous ecosystems [67,68]. It is worthwhile mentioning that intraspecific genetical diversity is usually accompanied by substantial intraspecific trait variation, which can help the species to enhance its competitive success and adapt more quickly to changing environmental conditions [69]. As molecular tools offer the opportunity to identify genetic diversity, new insights into the physiology and ecology of phytoplankton are provided. For instance, distinct subpopulations succession of diatom (e.g., Ditylum brightwellii) and dinoflagellates (e.g., Alexandrium catenella and Margalefidinium polykrikoides) occurred to maintain long-term algal blooms [70,71,72]. This intraspecific variation ultimately results in the diversification and evolutionary adaptation of species, which can even happen over ecological time scales [73,74]. The high sequence homology found among the LSU rDNA sequences of P. profundisulcus in ballast tank sediments and the populations from China, Australia, and Japan deposited in the NCBI database, likely suggesting a recent common ancestor or origin for these P. profundisulcus. This stimulated us to propose a further investigation on the origination and genetic diversity of P. profundisulcus once there are sufficient molecular data from deep sediments and different regions. The presence of P. profundisulcus cysts in ship’s ballast tanks increased the potential of bio-invasion risk to China and North America since the dinoflagellate in tanks survived as resistant resting cysts during the voyage and hatched in the accepted ecosystems. This study suggests that corresponding supervision and treatment measures should be taken to deal with the invasion of alien species via ship’s ballast water and sediments.
5. Conclusions
Although the dinoflagellate P. profundisulcus forms blooms almost every year since 2006 in China, whether it is an indigenous or an invasive species remains unclear. In this study, we confirmed the presence of P. profundisulcus resting cysts in the ballast sediments from vessels arriving in the ports of China and North America. Moreover, two P. profundisulcus cysts were germinated into vegetative cells in the ballast sediments from Vessel A, which is a general cargo ship built in 1997 and docked numerous ports in Brazil, USA, Japan, and Australia, before sailing to Huangdao Port, Shandong Province, and Jiangyin Port, Jiangsu Province in China. As P. profundisulcus has been reported in many regions of the world, our results suggested that this organism might be extensively transferred via ships’ ballast tank sediments. The origin and genetic diversity of P. profundisulcus should be further explored to gather sufficient molecular data from different sources, including that from deep sediments and different countries.
Author Contributions
Conceptualization, Y.Z.T. and Z.H.; methodology, X.Z. and Y.L.; software, W.T. and Y.D.; validation, L.S., Z.H. and Y.H.; formal analysis, X.Z. and Y.L.; investigation, X.Z. and L.S.; resources, W.T. and Y.H.; data curation, Z.H. and Y.D.; writing—original draft preparation, L.S.; writing—review and editing, Z.H. and Y.Z.T.; visualization, X.Z., L.S. and Z.H.; supervision, Y.Z.T. and Z.H.; project administration, Y.Z.T. and W.T.; funding acquisition, Y.Z.T. and W.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (grant number 41976134), the Science and Technology Basic Resources Investigation Program of China (grant number 2018FY100200), and Research Projects of General Administration of Customs (grant number 2021HK157).
Acknowledgments
We would like to thank Yuanyuan Sun from the Institute of Oceanology, Chinese Academy of Sciences for her assistance with the electron microscopy, and Bingli Liu and Hao Shen from Integrated Technical Service Center of Jiangyin Customs for the sample collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hallegraeff, G.M.; Bolch, C.J. Transport of diatom and dinoflagellate resting spores in ships’ ballast water: Implications for plankton biogeography and aquaculture. J. Plankton. Res. 1992, 14, 1067–1084. [Google Scholar] [CrossRef]
- Hamer, J.P.; Lucas, I.A.N.; McCollin, T.A. Harmful dinoflagellate resting cysts in ships’ ballast tank sediments: Potential for introduction into English and Welsh waters. Phycologia 2001, 40, 246–255. [Google Scholar] [CrossRef]
- Casas-Monroy, O.; Roy, S.; Rochon, A. Dinoflagellate cysts in ballast sediments: Differences between Canada’s east coast, west coast and the Great Lakes. Aquat. Conserv. 2013, 23, 254–276. [Google Scholar] [CrossRef]
- Garrett, M.J.; Puchulutegui, C.; Selwood, A.I.; Wolny, J.L. Identification of the harmful dinoflagellate Vulcanodinium rugosum recovered from a ballast tank of a globally traveled ship in Port Tampa Bay, Florida, USA. Harmful Algae 2014, 39, 202–209. [Google Scholar] [CrossRef]
- Carlton, J.T. Transoceanic and interoceanic dispersal of coastal marine organisms: The biology of ballast water. Oceanogr. Mar. Biol. 1985, 23, 313–371. [Google Scholar]
- Rey, A.; Basurko, O.C.; Rodriguez-Ezpeleta, N. The challenges and promises of genetic approaches for ballast water management. J. Sea Res. 2018, 133, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xie, D.; Bowler, P.A.; Zeng, Z.; Xiong, W.; Liu, C. Non-indigenous species in marine and coastal habitats of the South China Sea. Sci. Total Environ. 2021, 759, 143465. [Google Scholar] [CrossRef]
- Lv, B.; Cui, Y.; Tian, W.; Feng, D. Composition and influencing factors of bacterial communities in ballast tank sediments: Implications for ballast water and sediment management. Mar. Environ. Res. 2017, 132, 14–22. [Google Scholar] [CrossRef]
- Maglić, L.; Frančić, V.; Zec, D.; David, M. Ballast water sediment management in ports. Mar. Pollut. Bull. 2017, 147, 237–244. [Google Scholar] [CrossRef]
- Shang, L.; Hu, Z.; Deng, Y.; Liu, Y.; Zhai, X.; Chai, Z.; Liu, X.; Zhan, Z.; Dobbs, F.C.; Tang, Y. Metagenomic sequencing identifies highly diverse assemblages of dinoflagellate cysts in sediments from ships’ ballast tanks. Microorganisms 2019, 7, 250. [Google Scholar] [CrossRef] [Green Version]
- Hamer, J.P. Ballast Tank Sediments. In Invasive Aquatic Species of Europe. Distribution, Impacts and Management; Leppäkoski, E., Gollasch, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar] [CrossRef]
- Isabel Figueroa, R.; Estrada, M.; Garces, E. Life histories of microalgal species causing harmful blooms: Haploids, diploids and the relevance of benthic stages. Harmful Algae 2018, 73, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Z.; Hu, Z.; Deng, Y. Characteristical life history (resting cyst) provides a mechanism for recurrence and geographic expansion of harmful algal blooms of dinoflagellat: A review. Stud. Mar. Sin. 2016, 51, 132–154. (In Chinese) [Google Scholar] [CrossRef]
- Ribeiro, S.; Berge, T.; Lundholm, N.; Andersen, T.J.; Abrantes, F.; Ellegaard, M. Phytoplankton growth after a century of dormancy illuminates past resilience to catastrophic darkness. Nat. Commun. 2011, 1, 311. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.; Harris, A.S.D.; Jones, K.J.; Edmonds, R.L. Long-term survival of marine planktonic diatoms and dinoflagellates in stored sediment samples. J. Plankton. Res. 1999, 21, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Fukuyo, Y.; Matsuoka, K. Cyst Methodologies. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Enevoldsen, H.O., Eds.; IOC Manuals and Guides No. 33; UNESCO: Paris, France, 1995; pp. 229–2499. [Google Scholar]
- Doblin, M.A.; Coyne, K.J.; Rinta-Kanto, J.M.; Wilhelm, S.W.; Dobbs, F.C. Dynamics and short-term survival of toxic cyanobacteria species in ballast water from NOBOB vessels transiting the Great Lakes—Implications for HAB invasions. Harmful Algae 2007, 6, 519–530. [Google Scholar] [CrossRef]
- Casas-Monroy, O.; Roy, S.; Rochon, A. Ballast sediment-mediated transport of non-indigenous species of dinoflagellates on the East Coast of Canada. Aquat. Invasions 2011, 6, 231–248. [Google Scholar] [CrossRef]
- Hu, Z.; Xu, N.; Gu, H.; Chai, Z.; Takahashi, K.; Li, Z.; Deng, Y.; Iwataki, M.; Matsuoka, K.; Tang, Y.Z. Morpho-molecular description of a new HAB species, Pseudocochlodinium profundisulcus gen. et sp. nov., and its LSU rRNA gene based genetic diversity and geographical distribution. Harmful Algae 2021, 108, 102098. [Google Scholar] [CrossRef] [PubMed]
- Ou, L.; Zhang, Y.; Yang, L.; Wang, H.; Xie, X.; Rong, Z.; Lv, S.; Qi, Y. The outbreak of Cochlodinium geminatum bloom in Zhuhai, Guangdong. J. Trop. Oceanogr. 2010, 29, 57–61. (In Chinese) [Google Scholar] [CrossRef]
- Ke, Z.; Huang, L.; Tan, Y.; Song, X. A dinoflagellate Cochlodinium geminatum bloom in the Zhujiang (Pearl) River estuary in autumn 2009. Chin. J. Oceanol. Limn. 2012, 30, 371–378. [Google Scholar] [CrossRef]
- Pang, Y.; Nie, R.; Lu, S. Preliminary environmental analysis of the evolution of Cochlodinium geminatum bloom in the Pearl River Estuary. Ecol. Env. Sci. 2015, 24, 286–293. (In Chinese) [Google Scholar] [CrossRef]
- Dong, Y.; Cui, L.; Cao, R.; Cen, J.; Zou, J.; Zhou, X.; Lu, S. Ecological characteristics and teratogenic retinal determination of Cochlodinium geminatum blooms in Pearl River Estuary, South China. Ecotoxicol. Environ. Safe 2020, 191, 110226. [Google Scholar] [CrossRef]
- Bulletin of Marine Environmental Status of Guangdong province. 2011. Available online: http://gdee.gd.gov.cn/hjzkgb/content/post_2469372.html (accessed on 8 November 2021). (In Chinese)
- Bulletin of Marine Environmental Status of Guangdong province. 2012. Available online: http://gdee.gd.gov.cn/hjzkgb/content/post_2466238.html (accessed on 8 November 2021). (In Chinese)
- Wu, N.; Jiang, T.; Jiang, T.; Lv, S.; Huan, Q. Acute toxicity of Cochlodinium geminatum bloom waters from Pearl River Estuary on larvae of brine shrimp, fish and shrimp. J. Fish. China 2013, 37, 1328–1333. (In Chinese) [Google Scholar] [CrossRef]
- Yan, J.; Liu, J.; Cai, Y.; Duan, S.; Tang, Y.; Xu, N. Allelopathic effects and mechanisms of Cochlodinium geminatum isolated from the Pearl River Estuary. J. Appl. Phycol. 2019, 31, 2957–2967. [Google Scholar] [CrossRef] [Green Version]
- Lan, D.; Gu, H. Dinoflagellate Cysts along the Coast of China; Science Press: Beijing, China, 2014. (In Chinese) [Google Scholar]
- Iwataki, M.; Kawami, H.; Takano, Y.; Law, S.P.; Lu, S.G.; Fukuyo, Y.; Matsuoka, K. Morphology and phylogenetic position of an unarmored dinoflagellate forming brownish resting cyst. In Proceedings of the 13th International Conference on Harmful Algae, Hong Kong, China, 3–7 November 2008; p. 56. [Google Scholar]
- Hallegraeff, G.M.; Bolch, C.J.S.; Huisman, J.M.; de Salas, M.F. Planktonic dinoflagellates. In Algae of Australia Phytoplankton of Temperate Coastal Waters; Hallegraeff, G.M., Bolch, C.J.S., Hill, D.R., Jameson, I., LeRoi, J.M., McMinn, A., Murray, S., de Salas, M.F., Saunders, K., Eds.; CSIRO Publishing/ABRS: Melbourne, Australia, 2010; pp. 145–212. [Google Scholar]
- Johengen, T.H.; Reid, D.; Fahnenstiel, G.; MacIsaac, H.; Dobbs, F.; Doblin, M.; Ruiz, G.; Jenkins, P. Assessment of Transoceanic NOBOB Vessels and Low-Salinity Ballast Water as Vectors for Nonindigenous Species Introductions to the Great Lakes; University of Michigan and NOAA-Great Lakes Environmental Research Laboratory: Ann Arbor, MI, USA, 2005. [Google Scholar]
- Bolch, C.J.S. The use of sodium polytungstate for the separation and concentration of living dinoflagellate cysts from marine sediments. Phycologia 1997, 36, 472–478. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of phytoplankton for feedingmarine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 29–60. [Google Scholar] [CrossRef]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- Daugbjerg, N.; Hansen, G.; Larsen, J.; Moestrup, Ø. Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmoured dinoflagellates. Phycologia 2000, 39, 302–317. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.Z.; Gu, H.; Wang, Z.; Liu, D.; Wang, Y.; Lu, D.; Hu, Z.; Deng, Y.; Shang, L.; Qi, Y. Exploration of resting cysts (stages) and their relevance for possibly HABs-causing species in China. Harmful Algae 2021, 107, 102050. [Google Scholar] [CrossRef] [PubMed]
- Smayda, T.J. Novel and nuisance phytoplankton blooms in the sea: Evidence for a global epidemic. In Toxic Marine Phytoplankton; Graneli, E., Sundstr¨om, B., Edler, L., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 29–40. [Google Scholar]
- Anderson, D. HABs in a changing world: A perspective on harmful algal blooms, their impacts, and research and management in a dynamic era of climatic and environmental change. In Proceedings of the 15th International Conference On Harmful Algae, Changwon Gyeongnam, Korea, 29 October–2 November 2012; pp. 3–17. [Google Scholar]
- Gobler, C.J. Climate change and harmful algal blooms. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in understanding harmful algal blooms: Paradigm shifts and new technologies for research, monitoring, and management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [Green Version]
- Glibert, P.M.; Berdalet, E.; Burford, M.A.; Pitcher, G.C.; Zhou, M. Global Ecology and Oceanography of Harmful Algal Blooms, 1st ed.; Springer Nature: Cham, Switzerland, 2018. [Google Scholar]
- Hallegraeff, G.M.; Bolch, C.J. Transport of toxic dinoflagellate cysts via ships’ ballast water. Mar. Pollut. Bull. 1991, 22, 27–30. [Google Scholar] [CrossRef]
- Ruiz, G.M.; Carlton, J.T.; Grosholz, E.D.; Hines, A.H. Global invasions of marine and estuarine habitats by non-indigenous species: Mechanisms, extent, and consequences. Am. Zool. 1997, 37, 621–632. [Google Scholar] [CrossRef]
- McCarthy, H.P.; Crowder, L.B. An overlooked scale of global transport: Phytoplankton species richness in ships’ ballast water. Biol. Invasions 2000, 2, 321–322. [Google Scholar] [CrossRef]
- Bolch, C.J.S.; de Salas, M.F. A review of the molecular evidence for ballast water introduction of the toxic dinoflagellates Gymnodinium catenatum and the Alexandrium “tamarensis complex” to Australasia. Harmful Algae 2007, 6, 465–485. [Google Scholar] [CrossRef]
- Smayda, T.J. Reflections on the ballast water dispersal—Harmful algal bloom paradigm. Harmful Algae 2007, 6, 601–622. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Bescot, N.L.; Mahé, F.; Audic, S.; Dimier, C.; Garet, M.J.; Poulain, J.; Wincker, P.; Vargas, C.D.; Siano, R.J.E.M. Global patterns of pelagic dinoflagellate diversity across protist size classes unveiled by metabarcoding. Environ. Microbiol. 2016, 18, 609–626. [Google Scholar] [CrossRef]
- Hu, Z.; Shang, L.; Deng, Y.; Tang, Y.Z. Retrospect and prospect: Studies on geographical expansion of resting cysts of non-indigenous harmful algal bloom (HAB)-forming dinoflagellates via ships’ ballast tanks. Mar. Sci. 2020, 44, 103–115. (In Chinese) [Google Scholar]
- IMO. International Convention for the Control and Management of Ships’ Ballast Water and Sediments; IMO: London, UK, 2004. [Google Scholar]
- Qi, Y.; Ou, L.; Li, Y.; Lu, S. Taxonomy differentiation of harmful algal bloom causative species in genus Cochlodinium in Chinese coastal waters. In Proceedings of the Fifteen Academic Conference and Celebration of 30-year Anniversary for Chinese Society of Phycology, Jinan, China, 15–18 November 2009. (In Chinese). [Google Scholar]
- Qiu, D.; Huang, L.; Liu, S.; Zhang, H.; Lin, S. Apical groove type and molecular phylogeny suggests reclassification of Cochlodinium geminatum as Polykrikos geminatum. PLoS ONE 2013, 8, e71346. [Google Scholar] [CrossRef]
- Kofoid, C.A.; Swezy, O. The free living unarmored Dinoflagellata. In Memoirs of the University of California (Berkeley); The University of California (Berkeley): Berkeley, CA, USA, 1921; pp. 1–564. [Google Scholar]
- Shen, P.-P.; Li, Y.-N.; Qi, Y.-Z.; Zhang, L.-P.; Tan, Y.-H.; Huang, L.-M. Morphology and bloom dynamics of Cochlodinium geminatum (Schutt) Schutt in the Pearl River Estuary, South China Sea. Harmful Algae 2012, 13, 10–19. [Google Scholar] [CrossRef]
- Alpermann, T.J.; Beszteri, B.; John, U.; Tillmann, U.; Cembella, A.D. Implications of life-history transitions on the population genetic structure of the toxigenic marine dinoflagellate Alexandrium tamarense. Mol. Ecol. 2009, 18, 2122–2133. [Google Scholar] [CrossRef]
- Bolch, C.J.S.; Blackburn, S.I.; Hallegraeff, G.M.; Vaillancourt, R.E. Genetic variation among strains of the toxic dinoflagellate Gymnodinium catenatum (dinophyceae). J. Phycol. 1999, 35, 356–367. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Doucette, G.J.; Gallagher, J.C.; Balech, E. Biogeography of toxic dinoflagellates in the genus Alexandrium from the northeastern United States and Canada. Mar. Biol. 1994, 120, 467–478. [Google Scholar] [CrossRef]
- Genovesi-Giunti, B.; Laabir, M.; Vaquer, A. The benthic resting cyst: A key actor in harmful dinoflagellate blooms—A review. Vie et Milieu 2006, 56, 327–337. [Google Scholar]
- Anglès, S.; Garcés, E.; Reñé, A.; Sampedro, N. Life-cycle alternations in Alexandrium minutum natural populations from the NW Mediterranean Sea. Harmful Algae 2012, 16, 1–11. [Google Scholar] [CrossRef]
- Barrett, R.D.H.; Schluter, D. Adaptation from standing genetic variation. Trends. Ecol. Evol. 2008, 23, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Rynearson, T.A.; Newton, J.A.; Armbrust, E.V. Spring bloom development, genetic variation, and population succession in the planktonic diatom Ditylum bright. Limnol. Oceanogr. 2006, 51, 1249–1261. [Google Scholar] [CrossRef]
- Park, B.S.; Kim, J.H.; Kim, J.-H.; Baek, S.H.; Han, M.-S. Intraspecific bloom succession in the harmful dinoflagellate Cochlodinium polykrikoides (Dinophyceae) extended the blooming period in Korean coastal waters in 2009. Harmful Algae 2018, 71, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Erdner, D.L.; Richlen, M.; McCauley, L.A.R.; Anderson, D.M. Diversity and dynamics of a widespread bloom of the toxic dinoflagellate Alexandrium fundyense. PLoS ONE 2011, 6, e22965. [Google Scholar] [CrossRef] [Green Version]
- Carroll, S.P.; Hendry, A.P.; Reznick, D.N.; Fox, C.W. Evolution on ecological time-scales. Funct. Ecol. 2007, 21, 387–393. [Google Scholar] [CrossRef]
- Pelletier, F.; Garant, D.; Hendry, A.P. Eco-evolutionary dynamics. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 1483–1489. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).