
Investigating Molecular Mechanisms of Immunotoxicity and the Utility of ToxCast for Immunotoxicity Screening of Chemicals Added to Food

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Data-Mining Strategy for the Identification of Immune-Relevant High-Throughput Assays
2.2. Identification of Case Study Compounds
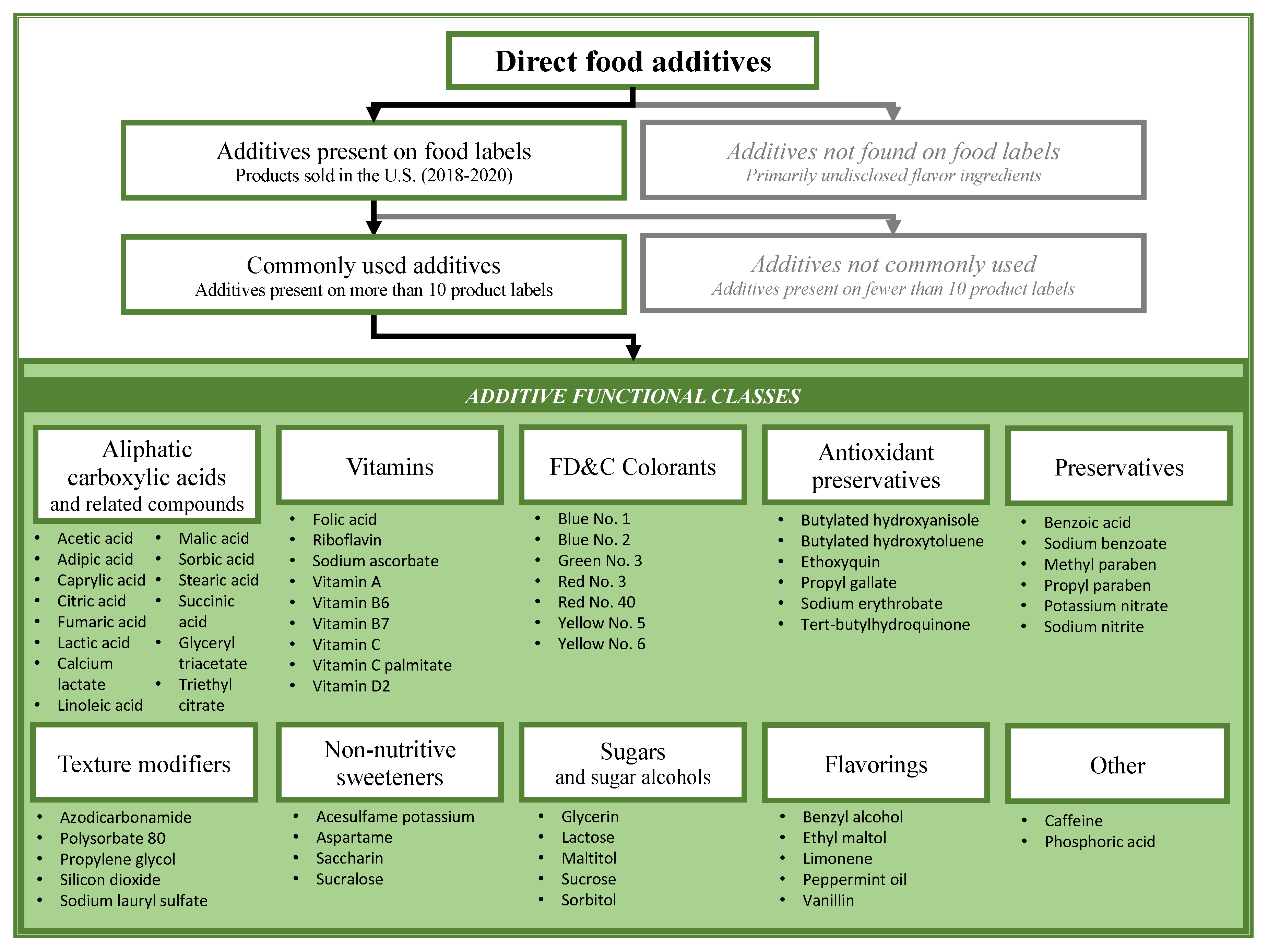
2.2.1. Direct Food Additives
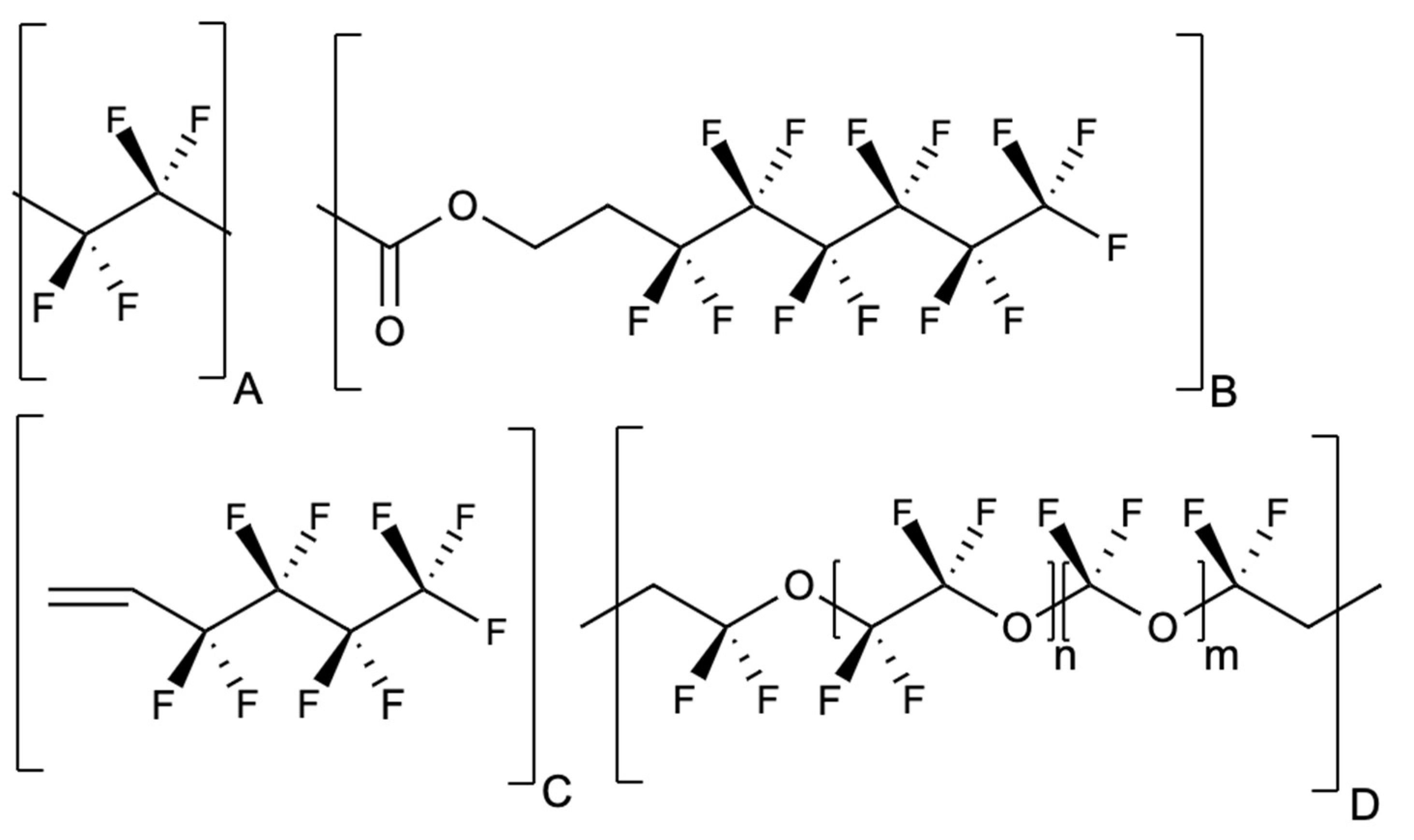
2.2.2. Indirect Additives: Per- and Polyfluoroalkyl Substances
3. Results
3.1. Identification of Case Study Compounds
3.2. Identification of ToxCast Active Assays
3.3. Identification of Immune-Specific Interactions in the Comparative Toxicogenomics Database
3.4. Analysis of Immune-Related Assays within ToxCast
3.5. Correlation Analysis of ToxCast and Immunological Data for TBHQ
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Material Tested | Migration Conditions | Detections of PFAS 1 | Findings |
|---|---|---|---|
| Cookware and food packaging United States, 2005 [91] | PFOA leaching into Miglyol and water was measured from different products, including popcorn bag, hamburger wrapper, sandwich wrapper, French fry box, and paper plates. | PFOA | Paper coatings with fluorotelomers released significantly higher amounts of PFOA than other tested products. The highest concentration of PFOA was released from microwave popcorn bags. |
| Fast food wrappers United States, 2008 [92] | PFAS migration measured from three retail fast food wrappers into food and food simulants (Miglyol, butter, water, vinegar, chocolate spread, and water/ethanol solutions (10, 20, 25, and 30% ethanol). Migration tests were run with 100 °C food/food simulant added to paper for 15 min; butter was tested at 4 °C for 40 days. | 3 PFAS species tested 2 | Reported significantly higher migration of PFAS into butter and other oil emulsion mixtures compared with migration into water, vinegar, oils, or alcohol. |
| Frying pans United States, 2007 [93] | Pans were heated for 30 min at 250 °C and the headspace gas was tested for a characteristic perfluorinated substance fragment, “-CF2-CF3”. | None reported | Did not detect PFAS compounds in the headspace gas. |
| Frying pans, cooking utensils, grill pans, pots, rice cookers, and non-stick baking papers. Korea, 2018 [94] | Analysis of PFAS migration from 312 food contact materials into food simulants, water, and corn oil at varying conditions: 4% acetic acid at 100 °C for 30 min; 50% ethanol and 50% n-heptane at 70 °C for 30 min followed by incubation at 25 °C for 1 h. | PFOA, PFNA, PFDoDA, PFTrDA, PFTeDA, PFHxDA, PFODA | Seven PFAS migrated into food simulants from 10 frying pans and 2 baking utensils. No PFAS migration was observed during subsequent testing and PFAS did not migrate from frying pans into corn oil or water. |
| Pet food paper bag 3 Spain, 2019 [95] | Analysis of PFAS migration from paper bag packaging into food simulant (Tenax) and milk at various conditions: 10 days at 40 °C; 2 h at 80 °C, 120 °C and 160 °C | PFBA, PFPeA, PFHxA, PFHpA, PFOA, PFNA, PFDA, PFUnDA, PFDoDA, PFTrDA, PFTeDA | Greater migration was observed into milk than the food simulant, and high migration percentages were observed for long-chain PFAS. Migration rates increased with temperature. |
| Muffin paper | Migration of fluorotelomers from muffin containers into butter, muffin dough, and Tenax food simulant measured after exposure to oven temperatures of 120–200 °C for 5 to 60 min. | 6:2 FTOH | The fluorotelomer concentrations were higher in the dough, butter, and Tanex food simulant after heating compared to the original levels in the baking cup paper, indicating the release of fluorotelomers from precursor compounds. |
| 8:2 FTOH | |||
| Germany, 2011 [96] | 10:2 FTOH | ||
| Microwave popcorn and foods in paper packaging Sweden, 2013 [97] | PFAS were tested in food samples before and after preparation as directed on the packaging. | 6:2 diPAP, 8:2 diPAP, 10:2 diPAP, PFHxA, PFOA, PFNA, PFDA, PFUnDA, PFTrDA | The PFAS concentrations, notably for polyfluoroalkyl phosphoric acid diesters (diPAP), increased in some packaged foods tested after heating in accordance with the package directions. |
| Instant food cups, microwave-popcorn bags, beverage cups, ice cream cups, fast food containers, dessert containers, and baking papers Thailand, 2012 [98] | The leaching of PFOA and PFOS from 34 food packaging products into methanol and saliva simulant at 80 °C during a 30 min period. | PFOA, PFOS | PFOA and PFOS migrated into saliva simulant from the majority of samples, all of which had detectable PFOA or PFOS. The highest migration of PFOA and PFOS was reported for a French-fry box and hot beverage cup. |
| Frying pans United States, 2005 [99] | The migration of PFOA from 11 frying pans into water, 10% ethanol, and 95% ethanol was measured after heating to 125 °C. | None reported | No PFOA was detected in any samples. |
| Frying pans, sandwich maker, waffle irons Germany, 2015 [100] | Volatilization of 9 PFAS into air was measured under normal use and under overheating scenarios. | PFBA, PFPeA, PFHxA, PFHpA, PFOA, PFNA, PFDA, PFUnDA, PFDoDA | PFAS release increased significantly at overheating temperatures, and all 9 PFAS were detected. PFOA emissions were lower than in prior reports. |
| Frying pans and microwave popcorn bags United States, 2007 [101] | The migration of 10 PFAS into air and water was measured from pans heated on a hotplate set to 250 °C and from microwave popcorn after microwave heating for 3 min. | PFPeA, PFHpA, PFOA, PFNA, PFDA, PFUnDA, PFDoDA, 6:2 FTOH, 8:2 FTOH | PFOA was released into the air from frying pans at normal cooking temperatures during consequent uses. One brand of microwave popcorn released much higher levels of PFAS compared to the other two. |
| Butter wrappers and dairy processing equipment Germany, 2013 [102] | Concentrations of 9 PFAS were measured in dairy products during processing. Migration of PFAS from butter wrappers was measured after 45 days at 5 °C. | PFBA, PFPeA. PFHxA, PFHpA, PFOA, PFNA, PFDA, PFDoA, 8:2 FTOH, 10:2 FTOH | Greater migration of PFOA and PFHxA relative to the longer-chain PFAS was observed from the butter wrapper. PFAS concentrations increased with greater fat concentrations in dairy products. |
| Two paper food contact materials United States, 2013 [103] | Migration of PFAS from two paper food packaging into five simulants (Miglyol oil, Miglyol oil with soy lecithin, Miglyol oil with Tween 60, 10% ethanol, 3% acetic acid) under two temperatures: 100 °C (15 min) and 40 °C (2, 24, 96 and 240 h). | PFHxA, PFHpA, PFOA, PFNA, PFDA, PFUnDA, PFDoDA | Seven PFAS migrated into food stimulants, and the shorter chain compounds migrated at a faster rate. The addition of emulsifiers increased migration efficiencies. Paper coating based on di-perfluoro-alkyloxy-amino-acid leached PFAS at a higher rate compared to the coating based on polyfluoroalkyl phosphate surfactants. |
| Paper bowl China, 2016 [104] | Migration studies of 16 PFAS from paper bowls into several food simulants: oil, water, and ethanol/water mixtures (10/90, 30/70, 50/50). Simulants preheated to 100 °C were added to a bowl, followed by a 15-min hold at room temperature. | 6:2 FTOH, 8:2 FTOH, 10:2 FTOH, 12:2 FTOH, 14:2 FTOH; 16:2 FTOH, PFBA, PFPeA, PFHxA, PFHpA, PFOA, PFNA, PFDA, PFUnDA, PFDoDA, PFTrDA, PFTeDA, PFPeDA, PFHxDA, PFHpDA, PFODA | Perfluorinated carboxylic acids and fluorotelomer alcohols readily migrated out of the paper bowls, with a greater transfer into 50% ethanol relative to 30% ethanol or water. PFBA (compound with 4 fluorinated carbons) had the greatest migration efficiency. |
| Pet food paper bag 3 Spain, 2020 [105] | Migration of 12 PFAS from unprinted pet food paper bags was measured to food simulants (Tenax, 50% ethanol, and 95% ethanol) and foods (ground cereal, parboiled rice, infant milk powder). | PFPeA, PFHxA, PFHpA, PFOA, PFNA, PFDA, 8:2 FTCA, 8:2 FTUCA | Shorter chain PFAS exhibited greater migration efficiencies than long-chain PFAS. More migration was observed into milk powder compared to food simulants. |
| Study Model | Dose | Main Findings |
|---|---|---|
| Mouse [44] | 0.0014% TBHQ diet fed to mice prior to infection with influenza virus | NK cell expression of granzyme B was decreased in the TBHQ-exposed mice, indicating impaired NK cell cytotoxicity. |
| Mouse [106] | 0.001% TBHQ diet fed to mice prior to ovalbumin exposure. | Following ovalbumin sensitization, a higher concentration of IgE and higher mast cell protease response were measured in the TBHQ group compared to controls. |
| Mouse [53] | Single intraperitoneal injection of 50 mM TBHQ | TBHQ upregulated cytokine IL-17D in an Nrf2-dependent manner and chemokine CCL2 in an Nrf2-independent manner. |
| Murine wild-type and Nrf2-null splenocytes [45] | Cells treated with 0.25–2.5 μM TBHQ | TBHQ enhanced Nrf2-dependent IgM production in B cells and decreased induction of CD22, CD25, CD69, and CD138 receptors both in wild-type and Nrf2-null B cells. |
| Murine splenocytes [57] | Cells treated with 1 μM or 5 μM TBHQ | Activation of NK cells in the presence of TBHQ decreased production of IFN-ɣ, granzyme B, and perforin, and lowered the induction of CD25 and CD69. |
| Splenocytes from Nrf2-null and wild-type mice [46] | Cells treated with 0.1–1 μM TBHQ | TBHQ inhibited the production of IL-2 and Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) in both wild-type and Nrf2-null T cells. |
| Human T cell line [58] | Jurkat cells treated with 0.1–1 μM TBHQ | TBHQ suppression of CD25 expression partly depended on Nrf2, while TBHQ inhibition of NFkB activation and IL-2 secretion was Nrf2-independent. |
| Rat thymocytes [107] | Cells treated with 10–300 μM TBHQ | TBHQ exposure activated Ca2+-dependent K+ channels and elevated intracellular Ca2+ levels. |
| Primary human CD4+ T cells [108] | Cells treated with 0.1–5 μM TBHQ | Inactivated human T cells, TBHQ inhibits the production of IL-2 and IFN-γ, inhibits the induction of CD25 and CD69, and suppresses NFκB DNA binding. |
| Human dendritic cells [109] | Cells treated with 10 μM TBHQ | TBHQ inhibited IL-12 expression in an Nrf2-dependent manner. |
| Primary human CD4+ T cells [110] | Cells treated with 50 μM TBHQ | TBHQ increased total Nrf2 levels. |
| Human T cell line [111] | Jurkat cells treated with 0.1–5 μM TBHQ | TBHQ inhibited interleukin-2 and CD25 expression and decreased NFκB transcriptional activity. |
| Murine T cells [70] | Cells treated with 0.1–1 μM TBHQ | TBHQ suppressed IFN-γ production and induced IL-4, IL-5, and IL-13 production. |
References
- Ferrario, D.; Gribaldo, L.; Hartung, T. Arsenic exposure and immunotoxicity: A review including the possible influence of age and sex. Curr. Environ. Health Rep. 2016, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, K.M.; Bai, S.; Barnette, D.; Blossom, S.J. Exposure cessation during adulthood did not prevent immunotoxicity caused by developmental exposure to low-level trichloroethylene in drinking water. Toxicol. Sci. 2017, 157, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.H.; Choi, K.C. Adverse effects of pesticides on the functions of immune system. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 235, 108789. [Google Scholar] [CrossRef]
- Grandjean, P. Delayed discovery, dissemination, and decisions on intervention in environmental health: A case study on immunotoxicity of perfluorinated alkylate substances. Environ Health. 2018, 17, 62. [Google Scholar] [CrossRef] [PubMed]
- Blossom, S.J.; Gilbert, K.M. Epigenetic underpinnings of developmental immunotoxicity and autoimmune disease. Curr. Opin. Toxicol. 2018, 10, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Dietert, R.R.; Zelikoff, J.R. Early-life environment, developmental immunotoxicology, and the risk of pediatric allergic disease including asthma. Birth Defects Res. B Dev. Reprod. Toxicol. 2008, 83, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Hessel, E.V.; Tonk, E.C.; Bos, P.M.; van Loveren, H.; Piersma, A.H. Developmental immunotoxicity of chemicals in rodents and its possible regulatory impact. Crit. Rev. Toxicol. 2015, 45, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Von der Embse, A.N.; De Witt, J.C. Developmental immunotoxicity (DIT) testing: Current recommendations and the future of DIT testing. Methods Mol. Biol. 2018, 1803, 47–56. [Google Scholar] [CrossRef]
- Vos, J.G. Immune suppression as related to toxicology. CRC Crit. Rev. Toxicol. 1977, 5, 67–101. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization International Programme on Chemical Safety. Guidance for Immunotoxicity Risk Assessment for Chemicals. 2012. Available online: http://www.inchem.org/documents/harmproj/harmproj/harmproj10.pdf (accessed on 15 February 2021).
- U.S. Congress Office of Technology Assessment. Identifying & Controlling Immunotoxic Substances Background Paper. 1991. OTA-BP-BA-75. Available online: https://ota.fas.org/reports/9124.pdf (accessed on 15 February 2021).
- Anderson, S.E.; Shane, H.L. Investigative Immunotoxicology. Methods Mol. Biol. 2018, 1803, 27–46. [Google Scholar] [CrossRef]
- Hartung, T.; Corsini, E. Immunotoxicology: Challenges in the 21st century and in vitro opportunities. ALTEX 2013, 30, 411–426. [Google Scholar] [CrossRef]
- Hastings, K.L. Immunotoxicology: A brief history. Methods Mol. Biol. 2018, 1803, 3–13. [Google Scholar] [CrossRef]
- Burleson, S.C.; Freebern, W.J.; Burleson, F.G.; Burleson, G.R.; Johnson, V.J.; Luebke, R. Host resistance assays. Methods Mol. Biol. 2018, 1803, 117–145. [Google Scholar] [CrossRef]
- European Chemical Agency. Guidance on Information Requirements and Chemical Safety Assessment Chapter, R.7a: Endpoint Specific Guidance. 2017. Available online: https://echa.europa.eu/guidance-documents/guidance-on-information-requirements-and-chemical-safety-assessment (accessed on 15 February 2021).
- U.S. Environmental Protection Agency. Health Effects Test Guidelines. OPPTS 870.7800. Immunotoxicity. 1998; EPA 712–C–96–351. Available online: https://www.epa.gov/test-guidelines-pesticides-and-toxic-substances/series-870-health-effects-test-guidelines (accessed on 15 February 2021).
- U.S. Environmental Protection Agency. Part 158 Toxicology Data Requirements: Guidance for Neurotoxicity Battery, Subchronic Inhalation, Subchronic Dermal and Immunotoxicity Studies. Office of Pesticides Programs. 2013. Available online: https://19january2017snapshot.epa.gov/sites/production/files/2014-02/documents/part158-tox-data-requirement.pdf (accessed on 15 February 2021).
- U.S. Environmental Protection Agency. Design for the Environment Program Alternatives Assessment Criteria for Hazard Evaluation Version 2.0. 2011. Available online: https://www.epa.gov/saferchoice/alternatives-assessment-criteria-hazard-evaluation (accessed on 15 February 2021).
- Rovida, C.; Barton-Maclaren, T.; Benfenati, E.; Caloni, F.; Charukeshi Chandrasekera, P.; Chesné, C.; Cronin, M.T.D.; De Knecht, J.; Dietrich, D.R.; Escher, S.E.; et al. Internationalization of read-across as a validated new approach method (NAM) for regulatory toxicology. ALTEX 2020. [Google Scholar] [CrossRef]
- Turley, A.E.; Isaacs, K.K.; Wetmore, B.A.; Karmaus, A.L.; Embry, M.R.; Krishan, M. Incorporating new approach methodologies in toxicity testing and exposure assessment for tiered risk assessment using the RISK21 approach: Case studies on food contact chemicals. Food Chem. Toxicol. 2019, 134, 110819. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.; Abedini, J.; Ceger, P.; Chang, X.; Cook, B.; Karmaus, A.L.; Lea, I.; Mansouri, K.; Phillips, J.; McAfee, E.; et al. An integrated chemical environment with tools for chemical safety testing. Toxicol. In Vitro 2020, 67, 104916. [Google Scholar] [CrossRef] [PubMed]
- Borrel, A.; Auerbach, S.S.; Houck, K.A.; Kleinstreuer, N.C. Tox21BodyMap: A webtool to map chemical effects on the human body. Nucleic Acids Res. 2020, 48, W472–W476. [Google Scholar] [CrossRef]
- Kleinstreuer, N.C.; Yang, J.; Berg, E.L.; Knudsen, T.B.; Richard, A.M.; Martin, M.T.; Reif, D.M.; Judson, R.S.; Polokoff, M.; Dix, D.J.; et al. Phenotypic screening of the ToxCast chemical library to classify toxic and therapeutic mechanisms. Nat. Biotechnol. 2014, 32, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Richard, A.M.; Judson, R.S.; Houck, K.A.; Grulke, C.M.; Volarath, P.; Thillainadarajah, I.; Yang, C.; Rathman, J.; Martin, M.T.; Wambaugh, J.F.; et al. ToxCast chemical landscape: Paving the road to 21st century toxicology. Chem. Res. Toxicol. 2016, 29, 1225–1251. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.A.; Guyton, K.Z.; Martin, M.T.; Reif, D.M.; Rusyn, I. Use of high-throughput in vitro toxicity screening data in cancer hazard evaluations by IARC monograph working groups. ALTEX 2018, 35, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.; Pham, N.; Marty, M.; Sandy, M.; Solomon, G.; Zeise, L. An integrated approach using publicly available resources for identifying and characterizing chemicals of potential toxicity concern: Proof-of-concept with chemicals that affect cancer pathways. Toxicol. Sci. 2019, 169, 14–24. [Google Scholar] [CrossRef]
- Karmaus, P.W.F.; Karmaus, A.L. Challenges for integrating immunotoxicology into the twenty-first-century toxicology testing paradigm. Methods Mol. Biol. 2018, 1803, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; McMorran, R.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. The comparative toxicogenomics database: Update 2019. Nucleic Acids Res. 2019, 47, D948–D954. [Google Scholar] [CrossRef] [PubMed]
- Kosnik, M.B.; Planchart, A.; Marvel, S.W.; Reif, D.M.; Mattingly, C.J. Integration of curated and high-throughput screening data to elucidate environmental influences on disease pathways. Comput. Toxicol. 2019, 12, 100094. [Google Scholar] [CrossRef] [PubMed]
- Judson, R.; Houck, K.; Martin, M.; Richard, A.M.; Knudsen, T.B.; Shah, I.; Little, S.; Wambaugh, J.; Setzer, R.W.; Kothya, P.; et al. Analysis of the effects of cell stress and cytotoxicity on in vitro assay activity across a diverse chemical and assay space. Toxicol. Sci. 2016, 152, 323–339. [Google Scholar] [CrossRef]
- Karmaus, A.L.; Filer, D.L.; Martin, M.T.; Houck, K.A. Evaluation of food-relevant chemicals in the ToxCast high-throughput screening program. Food Chem. Toxicol. 2016, 92, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Karmaus, A.L.; Trautman, T.D.; Krishan, M.; Filer, D.L.; Fix, L.A. Curation of food-relevant chemicals in ToxCast. Food Chem. Toxicol. 2017, 103, 174–182. [Google Scholar] [CrossRef]
- National Toxicology Program. Report on Carcinogens, Fourteenth Edition. Butylated Hydroxyanisole. Available online: https://ntp.niehs.nih.gov/ntp/roc/content/profiles/butylatedhydroxyanisole.pdf (accessed on 15 February 2021).
- European Food Safety Authority. Risks to human health related to the presence of perfluoroalkyl substances in food. EFSA J. 2020, 18, e06223. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Packaging & Food Contact Substances (FCS). Available online: https://www.fda.gov/food/food-ingredients-packaging/packaging-food-contact-substances-fcs (accessed on 15 February 2021).
- U.S. Food and Drug Administration. Indirect Food Additives: Paper and Paperboard Components. 81 Fed. Reg. 5. Available online: https://www.federalregister.gov/documents/2016/01/04/2015-33026/indirect-food-additives-paper-and-paperboard-components (accessed on 15 February 2021).
- Rice, P.A. C6-Perfluorinated compounds: The new greaseproofing agents in food packaging. Curr. Environ. Health Rep. 2015, 2, 33–40. [Google Scholar] [CrossRef]
- Schaider, L.A.; Balan, S.A.; Blum, A.; Andrews, D.Q.; Strynar, M.J.; Dickinson, M.E.; Lunderberg, D.M.; Lang, J.R.; Peaslee, G.F. Fluorinated compounds in U.S. fast food packaging. Environ. Sci. Technol. Lett. 2017, 4, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Susmann, H.P.; Schaider, L.A.; Rodgers, K.M.; Rudel, R.A. Dietary habits related to food packaging and population exposure to PFASs. Environ. Health Perspect. 2019, 127, 107003. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. FDA Announces the Voluntary Phase-Out by Industry of Certain PFAS Used in Food Packaging. Constituent Update 31 July 2020. Available online: https://www.fda.gov/food/cfsan-constituent-updates/fda-announces-voluntary-phase-out-industry-certain-pfas-used-food-packaging (accessed on 15 February 2021).
- Trier, X.; Taxvig, X.; Rosenmai, A.K.; Pedersen, G.A. PFAS in Paper and Board for Food Contact: Options for Risk Management of Poly- and Perfluorinated Substances. Nordisk Ministerråd. 2017. Available online: https://orbit.dtu.dk/files/149769110/Rapport_PFAS_in_paper_and_board_for_food_contact_Options_for_risk_management_of_poly_and_perfluorina.pdf (accessed on 15 February 2021).
- Rice, P.A.; Aungst, J.; Cooper, J.; Bandele, O.; Kabadi, S.V. Comparative analysis of the toxicological databases for 6:2 fluorotelomer alcohol (6:2 FTOH) and perfluorohexanoic acid (PFHxA). Food Chem. Toxicol. 2020, 138, 111210. [Google Scholar] [CrossRef]
- Boss, A.; Freeborn, R.; Liu, S.; Zagorski, J.; Turley, A.; Gardner, E.; Rockwell, C. The food additive tBHQ impairs NK cell cytotoxicity against influenza infection (FS12-02-19). Curr. Dev. Nutr. 2019, 3, nzz049-FS12. [Google Scholar] [CrossRef]
- Bursley, J.K.; Rockwell, C.E. Nrf2-dependent and -independent effects of tBHQ in activated murine B cells. Food Chem. Toxicol. 2020, 145, 111595. [Google Scholar] [CrossRef]
- Zagorski, J.W.; Turley, A.E.; Freeborn, R.A.; Van Den Berg, K.R.; Dover, H.E.; Kardell, B.R.; Liby, K.T.; Rockwell, C.E. Differential effects of the Nrf2 activators tBHQ and CDDO-Im on the early events of T cell activation. Biochem. Pharmacol. 2018, 147, 67–76. [Google Scholar] [CrossRef]
- DeWitt, J.C.; Blossom, S.J.; Schaider, L.A. Exposure to per-fluoroalkyl and polyfluoroalkyl substances leads to immunotoxicity: Epidemiological and toxicological evidence. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 148–156. [Google Scholar] [CrossRef]
- Houck, K.A.; Dix, D.J.; Judson, R.S.; Kavlock, R.J.; Yang, J.; Berg, E.L. Profiling bioactivity of the ToxCast chemical library using BioMAP primary human cell systems. J. Biomol. Screen. 2009, 14, 1054–1066. [Google Scholar] [CrossRef]
- Rosen, M.B.; Das, K.P.; Rooney, J.; Abbott, B.; Lau, C.; Corton, J.C. PPARα-independent transcriptional targets of perfluoroalkyl acids revealed by transcript profiling. Toxicology 2017, 387, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Ait Bamai, Y.; Goudarzi, H.; Araki, A.; Okada, E.; Kashino, I.; Miyashita, C.; Kishi, R. Effect of prenatal exposure to per- and polyfluoroalkyl substances on childhood allergies and common infectious diseases in children up to age 7 years: The Hokkaido study on environment and children’s health. Environ. Int. 2020, 143, 105979. [Google Scholar] [CrossRef]
- Salihovic, S.; Lind, L.; Larsson, A.; Lind, P.M. Plasma perfluoroalkyls are associated with decreased levels of proteomic inflammatory markers in a cross-sectional study of an elderly population. Environ. Int. 2020, 145, 106099. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kong, A.N. Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. 2009, 48, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Seelige, R.; Saddawi-Konefka, R.; Adams, N.M.; Picarda, G.; Sun, J.C.; Benedict, C.A.; Bui, J.D. Interleukin-17D and Nrf2 mediate initial innate immune cell recruitment and restrict MCMV infection. Sci. Rep. 2018, 8, 13670. [Google Scholar] [CrossRef] [PubMed]
- Gharavi, N.; El-Kadi, A.O. tert-Butylhydroquinone is a novel aryl hydrocarbon receptor ligand. Drug Metab. Dispos. 2005, 33, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, T.D.; Köhle, C.; Buckler, F.; Schmohl, S.; Braeuning, A.; Schmiechen, A.; Schwarz, M.; Münzel, P.A. Regulation of CYP1A1 gene expression by the antioxidant tert-butylhydroquinone. Drug Metab. Dispos. 2006, 34, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Vázquez, C.; Quintana, F.J. Regulation of the immune response by the aryl hydrocarbon receptor. Immunity 2018, 48, 19–33. [Google Scholar] [CrossRef]
- Boss, A.P.; Freeborn, R.A.; Duriancik, D.M.; Kennedy, R.C.; Gardner, E.M.; Rockwell, C.E. The Nrf2 activator tBHQ inhibits the activation of primary murine natural killer cells. Food Chem. Toxicol. 2018, 121, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Zagorski, J.W.; Maser, T.P.; Liby, K.T.; Rockwell, C.E. Nrf2-dependent and-independent effects of tert-butylhydroquinone, CDDO-Im, and H2O2 in human Jurkat T cells as determined by CRISPR/Cas9 gene editing. J. Pharmacol. Exp. Ther. 2017, 361, 259–267. [Google Scholar] [CrossRef]
- Koh, K.; Cha, Y.; Kim, S.; Kim, J. tBHQ inhibits LPS-induced microglial activation via Nrf2-mediated suppression of p38 phosphorylation. Biochem. Biophys. Res. Commun. 2009, 380, 449–453. [Google Scholar] [CrossRef]
- Tsatsakis, A.; Petrakis, D.; Nikolouzakis, T.K.; Docea, A.O.; Calina, D.; Vinceti, M.; Goumenou, M.; Kostoff, R.N.; Mamoulakis, C.; Aschner, M.; et al. COVID-19, an opportunity to reevaluate the correlation between long-term effects of anthropogenic pollutants on viral epidemic/pandemic events and prevalence. Food Chem. Toxicol. 2020, 141, 111418. [Google Scholar] [CrossRef]
- Punt, A.; Firman, J.; Boobis, A.; Cronin, M.; Gosling, J.P.; Wilks, M.F.; Hepburn, P.A.; Thiel, A.; Fussell, K.C. Potential of ToxCast data in the safety assessment of food chemicals. Toxicol. Sci. 2020, 174, 326–340. [Google Scholar] [CrossRef]
- Maffini, M.V.; Neltner, T.G.; Vogel, S. We are what we eat: Regulatory gaps in the United States that put our health at risk. PLoS Biol. 2017, 15, e2003578. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Guidance for Industry and Other Stakeholders: Redbook 2000. Toxicological Principles for the Safety Assessment of Food Ingredients. July 2007. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-and-other-stakeholders-redbook-2000 (accessed on 15 February 2021).
- U.S. Food and Drug Administration. Guidance for Industry: Preparation of Food Contact Notifications for Food Contact Substances (Toxicology Recommendations). April 2002. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-preparation-food-contact-notifications-food-contact-substances-toxicology (accessed on 15 February 2021).
- Groh, K.J.; Geueke, B.; Martin, O.; Maffini, M.; Muncke, J. Overview of intentionally used food contact chemicals and their hazards. Environ. Int. 2021, 150, 106225. [Google Scholar] [CrossRef] [PubMed]
- Ogungbesan, A.; Neal-Kluever, A.; Rice, P. Exploring the use of current immunological assays for the developmental immunotoxicity assessment of food contact materials. Food Chem. Toxicol. 2019, 133, 110801. [Google Scholar] [CrossRef]
- Chappell, G.A.; Britt, J.K.; Borghoff, S.J. Systematic assessment of mechanistic data for FDA-certified food colors and neurodevelopmental processes. Food Chem. Toxicol. 2020, 140, 111310. [Google Scholar] [CrossRef] [PubMed]
- Mestas, J.; Hughes, C.C.W. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef]
- Shay, T.; Jojic, V.; Rothamel, K.; Puyraimon-Zemmour, D.; Feng, T.; Wakamatsu, E.; Benoist, C.; Koller, D.; Regev, A.; ImmGen Consortium. Conservation and divergence in the transcriptional programs of the human and mouse immune systems. Proc. Natl. Acad. Sci. USA 2013, 110, 2946–2951. [Google Scholar] [CrossRef]
- Rockwell, C.E.; Zhang, M.; Fields, P.E.; Klaassen, C.D. Th2 skewing by activation of Nrf2 in CD4+ T cells. J. Immunol. 2012, 188, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Food Additives and Contaminants. WHO Food Additives Series 40. World Health Organization, Geneva. 1998. tert-Butylhydroquinone (TBHQ). Available online: http://www.inchem.org/documents/jecfa/jecmono/v040je02.htm (accessed on 15 February 2021). tert-Butylhydroquinone (TBHQ).
- National Toxicology Program. NTP Toxicology and Carcinogenesis Studies of t-Butylhydroquinone (CAS No. 1948-33-0) in F344/N Rats and B6C3F(1) Mice (Feed Studies). Natl Toxicol Program Tech Rep Ser. 459:1-326. Available online: https://ntp.niehs.nih.gov/go/tr459 (accessed on 15 February 2021).
- European Food Safety Authority. Opinion of the Scientific Panel on food additives, flavourings, processing aids and materials in contact with food (AFC) on a request from the Commission related to tertiary-Butylhydroquinone (TBHQ). EFSA J. 2004, 2, 84. [Google Scholar] [CrossRef]
- Dalsager, L.; Christensen, N.; Husby, S.; Kyhl, H.; Nielsen, F.; Høst, A.; Grandjean, P.; Jensen, T.K. Association between prenatal exposure to perfluorinated compounds and symptoms of infections at age 1–4 years among 359 children in the Odense Child Cohort. Environ. Int. 2016, 96, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Granum, B.; Haug, L.S.; Namork, E.; Stolevik, S.B.; Thomsen, C.; Aaberge, I.S.; van Loveren, H.; Løvik, M.; Nygaard, U.C. Pre-natal exposure to perfluoroalkyl substances may be associated with altered vaccine antibody levels and immune-related health outcomes in early childhood. J. Immunotoxicol. 2013, 10, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Averina, M.; Brox, J.; Huber, S.; Furberg, A.S.; Sørensen, M. Serum perfluoroalkyl substance (PFAS) and risk of asthma and various allergies in adolescents. The Tromsø study Fit Futures in Northern Norway. Environ. Res. 2019, 169, 114–121. [Google Scholar] [CrossRef]
- Buser, M.C.; Scinicariello, F. Perfluoroalkyl substances and food allergies in adolescents. Environ. Int. 2016, 88, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Corsini, E.; Luebke, R.W.; Germolec, D.R.; DeWitt, J.C. Perfluorinated compounds: Emerging POPs with potential immunotoxicity. Toxicol. Lett. 2014, 230, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Le, Y.; Bu, P.; Cheng, X. Regulation of Hox and ParaHox genes by perfluorochemicals in mouse liver. Toxicology 2020, 441, 152521. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Jia, X.; Gao, N.; Wu, Y.; Liu, Z.; Lu, X.; Du, Q.; He, J.; Li, N.; Chen, B.; et al. Role of the Nrf2-ARE pathway in perfluorooctanoic acid (PFOA)-induced hepatotoxicity in Rana nigromaculata. Environ. Pollut. 2018, 238, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yang, B.; Wu, L.; Zou, W.; Pan, X.; Zou, T.; Liu, F.; Xia, L.; Wang, X.; Zhang, D. Involvement of NRF2 in perfluorooctanoic acid-induced testicular damage in male mice. Biol. Reprod. 2015, 93, 41. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.B.; Lee, J.S.; Ren, H.; Vallanat, B.; Liu, J.; Waalkes, M.P.; Abbott, B.D.; Lau, C.; Corton, J.C. Toxicogenomic dissection of the perfluorooctanoic acid transcript profile in mouse liver: Evidence for the involvement of nuclear receptors PPAR alpha and CAR. Toxicol. Sci. 2008, 103, 46–56. [Google Scholar] [CrossRef]
- Rani, A.; Murphy, J.J. STAT5 in cancer and immunity. J. Interferon Cytokine Res. 2016, 36, 226–237. [Google Scholar] [CrossRef]
- Bansal, A.; Henao-Mejia, J.; Simmons, R.A. Immune system: An emerging player in mediating effects of endocrine disruptors on metabolic health. Endocrinology 2018, 159, 32–45. [Google Scholar] [CrossRef]
- Ng, C.A.; Hungerbuehler, K. Exploring the use of molecular docking to identify bioaccumulative perfluorinated alkyl acids (PFAAs). Environ. Sci. Technol. 2015, 49, 12306–12314. [Google Scholar] [CrossRef]
- Li, C.H.; Ren, X.M.; Guo, L.H. Adipogenic activity of oligomeric hexafluoropropylene oxide (perfluorooctanoic acid alternative) through peroxisome proliferator-activated receptor γ pathway. Environ. Sci. Technol. 2019, 53, 3287–3295. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Ren, X.M.; Wan, B.; Guo, L.H. Comparative in vitro and in vivo evaluation of the estrogenic effect of hexafluoropropylene oxide homologues. Environ. Sci. Technol. 2019, 53, 8371–8380. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, N.J.M.; Wargenau, A.; Sorenson, C.; Pedersen, J.; Tufenkji, N.; Novak, P.J.; Simcik, M.F. Partitioning and accumulation of perfluoroalkyl substances in model lipid bilayers and bacteria. Environ. Sci. Technol. 2018, 52, 10433–10440. [Google Scholar] [CrossRef]
- Nouhi, S.; Ahrens, L.; Campos Pereira, H.; Hughes, A.V.; Campana, M.; Gutfreund, P.; Palsson, G.K.; Vorobiev, A.; Hellsing, M.S. Interactions of perfluoroalkyl substances with a phospholipid bilayer studied by neutron reflectometry. J. Colloid. Interface Sci. 2018, 511, 474–481. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, J.C.; Germolec, D.R.; Luebke, R.W.; Johnson, V.J. Associating changes in the immune system with clinical diseases for interpretation in risk assessment. Curr. Protoc. Toxicol. 2016, 67, 18.1.1–18.1.22. [Google Scholar] [CrossRef] [PubMed]
- Begley, T.H.; White, K.; Honigfort, P.; Twaroski, M.L.; Neches, R.; Walker, R.A. Perfluorochemicals: Potential sources of and migration from food packaging. Food Addit. Contam. 2005, 22, 1023–1031. [Google Scholar] [CrossRef]
- Begley, T.H.; Hsu, W.; Noonan, G.; Diachenko, G. Migration of fluorochemical paper additives from food-contact paper into foods and food simulants. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 384–390. [Google Scholar] [CrossRef]
- Bradley, E.L.; Read, W.A.; Castle, L. Investigation into the migration potential of coating materials from cookware products. Food Addit. Contam. 2007, 24, 326–335. [Google Scholar] [CrossRef]
- Choi, H.; Bae, I.A.; Choi, J.C.; Park, S.J.; Kim, M. Perfluorinated compounds in food simulants after migration from fluorocarbon resin-coated frying pans, baking utensils, and non-stick baking papers on the Korean market. Food Addit. Contam. Part B Surveill. 2018, 11, 264–272. [Google Scholar] [CrossRef]
- Elizalde, M.P.; Gomez-Lavin, S.; Urtiaga, A.M. Migration of perfluorinated compounds from paper bag to Tenax® and lyophilised milk at different temperatures. Intern. J. Environm. Anal. Chem. 2019, 98, 1423–1433. [Google Scholar] [CrossRef]
- Fengler, R.; Schlummer, M.; Gruber, L.; Fiedler, D.; Weise, N. Migration of Fluorinated Telomer Alcohols (FTOH) from Food Contact Materials into Food at Elevated Temperatures. In Proceedings of the Conference paper presented at 1th International Symposium on Halogenated Persistent Organic Pollutants (POPs)-Dioxin, Brussels, Belgium, 21–25 August 2011; Available online: http://dev.dioxin20xx.org/wp-content/uploads/pdfs/2011/2107.pdf (accessed on 15 February 2021).
- Gebbink, W.A.; Ullah, S.; Sandblom, O.; Berger, U. Polyfluoroalkyl phosphate esters and perfluoroalkyl carboxylic acids in target food samples and packaging--method development and screening. Environ. Sci. Pollut. Res. Int. 2013, 20, 7949–7958. [Google Scholar] [CrossRef] [PubMed]
- Poothong, A.; Boontanon, S.K.; Boontanon, N. Determination of perfluorooctane sulfonate and perfluorooctanoic acid in food packaging using liquid chromatography coupled with tandem mass spectrometry. J. Hazard. Mater. 2012, 205-206, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Poweley, C.R.; Michalczyk, M.J.; Kaiser, M.A.; Buxton, W. Determination of perfluorooctanoic acid (PFOA) extractable from the surface of commercial cookware under simulated cooking conditions by LC/MS/MS. Analyst 2005, 130, 1299–1302. [Google Scholar] [CrossRef]
- Schlummer, M.; Sölch, C.; Meisel, T.; Still, M.; Gruber, L.; Wolz, G. Emission of perfluoroalkyl carboxylic acids (PFCA) from heated surfaces made of polytetrafluoroethylene (PTFE) applied in food contact materials and consumer products. Chemosphere 2015, 129, 46–53. [Google Scholar] [CrossRef]
- Sinclair, E.; Kim, S.K.; Akinleye, H.B.; Kannan, K. Quantitation of gas-phase perfluoroalkyl surfactants and fluorotelomer alcohols released from nonstick cookware and microwave popcorn bags. Environ. Sci. Technol. 2007, 41, 1180–1185. [Google Scholar] [CrossRef]
- Still, M.; Schlummer, M.; Gruber, L.; Fiedler, D.; Wolz, G. Impact of industrial production and packaging processes on the concentration of per- and polyfluorinated compounds in milk and dairy products. J. Agric. Food Chem. 2013, 61, 9052–9062. [Google Scholar] [CrossRef]
- Xu, Y.; Noonan, G.O.; Begley, T.H. Migration of perfluoroalkyl acids from food packaging to food simulants. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2013, 30, 899–908. [Google Scholar] [CrossRef]
- Yuan, G.; Peng, H.; Huang, C.; Hu, J. Ubiquitous occurrence of fluorotelomer alcohols in eco-friendly paper-made food-contact materials and their implication for human exposure. Environ. Sci. Technol. 2016, 50, 942–950. [Google Scholar] [CrossRef]
- Zabaleta, I.; Blanco-Zubiaguirre, L.; Baharli, E.N.; Olivares, M.; Prieto, A.; Zuloaga, O.; Elizalde, M.P. Occurrence of per- and polyfluorinated compounds in paper and board packaging materials and migration to food simulants and foodstuffs. Food Chem. 2020, 321, 126746. [Google Scholar] [CrossRef]
- Jin, Y.N.; Bursley, J.; Dover, H.; Gangur, V.; Rockwell, C.E. The effect of the common food additive tBHQ in OVA-elicited food allergy. J. Immunol. 2018, 200, 104.4. [Google Scholar]
- Takeda, M.; Oyama, K.; Kamemura, N.; Kanemaru, K.; Yuasa, K.; Yokoigawa, K.; Oyama, Y. Change in plasma membrane potential of rat thymocytes by tert-butylhydroquinone, a food additive: Possible risk on lymphocytes. Food Chem. Toxicol. 2017, 109, 296–301. [Google Scholar] [CrossRef]
- Turley, A.E.; Zagorski, J.W.; Rockwell, C.E. The Nrf2 activator tBHQ inhibits T cell activation of primary human CD4 T cells. Cytokine 2015, 71, 289–295. [Google Scholar] [CrossRef]
- Macoch, M.; Morzadec, C.; Génard, R.; Pallardy, M.; Kerdine-Römer, S.; Fardel, O.; Vernhet, L. Nrf2-dependent repression of interleukin-12 expression in human dendritic cells exposed to inorganic arsenic. Free Radic. Biol. Med. 2015, 88, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Morzadec, C.; Macoch, M.; Sparfel, L.; Kerdine-Römer, S.; Fardel, O.; Vernhet, L. Nrf2 expression and activity in human T lymphocytes: Stimulation by T cell receptor activation and priming by inorganic arsenic and tert-butylhydroquinone. Free Radic. Biol. Med. 2014, 71, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Zagorski, J.W.; Turley, A.E.; Dover, H.E.; Van Den Berg, K.R.; Compton, J.R.; Rockwell, C.E. The Nrf2 activator, tBHQ, differentially affects early events following stimulation of Jurkat cells. Toxicol. Sci. 2013, 136, 63–71. [Google Scholar] [CrossRef] [PubMed]


| PFAS Reported to Migrate to Food, with CAS Numbers | Number of Assays with Half-Maximal Activity Concentration (AC50) < Cytotoxicity Limit |
|---|---|
| Perfluorooctanesulfonic acid (1763-23-1) | 48 |
| Perfluoroundecanoic acid (2058-94-8) | 45 |
| Perfluorooctanoic acid (335-67-1) | 41 |
| Perfluorohexanoic acid (307-24-4) | 22 |
| Perfluorodecanoic acid (335-76-2) | 18 |
| Potassium perfluorooctanesulfonate (2795-39-3) | 13 |
| Ammonium perfluorooctanoate (3825-26-1) | 11 |
| Perfluorononanoic acid (375-95-1) | 9 |
| Perfluoroheptanoic acid (375-85-9) | 5 |
| 6:2 fluorotelomer alcohol (647-42-7) | 4 |
| 8:2 fluorotelomer alcohol (678-39-7) | 1 |
| 6:2 fluorotelomer methacrylate (2144-53-8) | 1 |
| Lithium perfluorooctanesulfonate (29457-72-5) | 0 |
| Direct Food Additives | Number of Assays with Half-Maximal Activity Concentration (AC50) < Cytotoxicity Limit |
|---|---|
| Tert-butylhydroquinone (TBHQ) | 58 |
| FD&C Red No. 3 (erythrosine) | 46 |
| Propyl paraben | 23 |
| Propyl gallate | 21 |
| Ethoxyquin, FD&C Blue No. 1, folic acid, sodium lauryl sulfate, sorbic acid, vitamin D2 | 11–16 |
| Acetic acid, caprylic acid, FD&C Green No. 3, maltol, methyl paraben, sodium ascorbate, stearic acid, triethyl citrate, vitamin A | 6–10 |
| Acesulfame potassium, adipic acid, ascorbyl palmitate, aspartame, azodicarbonamide, benzoic acid, benzyl alcohol, butylated hydroxytoluene, caffeine, calcium lactate, citric acid, ethyl maltol, FD&C Red No. 40, FD&C Yellow No. 5, FD&C Yellow No. 6, fumaric acid, glycerin, glyceryl triacetate, limonene, linoleic acid, malic acid, peppermint oil, phosphoric acid, potassium nitrate, propylene glycol, riboflavin, saccharin, silicon dioxide, sodium erythorbate, sodium nitrite, sorbitol, sucralose, sugar, vanillin, vitamin B6, vitamin C | 1–5 |
| Butylated hydroxyanisole, FD&C Blue No. 2, lactic acid, lactose, polysorbate 80, sodium benzoate, succinic acid, vitamin B7 | 0 |
| Gene Name and Function | ToxCast Assay Name | TBHQ | FD&C Red 3 | PFOS | PFOA | PFNA | PFDA | PFUnDA |
|---|---|---|---|---|---|---|---|---|
| CCL2 (chemokine (C-C motif) ligand 2) Chemokine with chemotactic activity for monocytes and basophils | BSK_3C_MCP1_down | ✓ | ||||||
| BSK_CASM3C_MCP1_down | ✓ | |||||||
| BSK_KF3CT_MCP1_down | ✓ | ✓ | ✓ | |||||
| BSK_LPS_MCP1_down | ✓ | ✓ | ||||||
| BSK_SAg_MCP1_down | ✓ | |||||||
| CCL26 (chemokine (C-C motif) ligand 26) Chemokine with chemotactic activity for eosinophils and basophils | BSK_4H_Eotaxin3_down | ✓ | ||||||
| CD38 molecule Transmembrane receptor expressed on macrophages, dendritic cells, and NK cells | BSK_SAg_CD38_down | ✓ | ||||||
| CD40 molecule Transmembrane receptor expressed on antigen-presenting cells | BSK_LPS_CD40_down | ✓ | ||||||
| BSK_SAg_CD40_down | ✓ | |||||||
| CD69 molecule Transmembrane receptor expressed on activated T cells and NK cells | BSK_SAg_CD69_down | ✓ | ||||||
| CSF1 (macrophage colony-stimulating factor) Cytokine that controls the differentiation and function of macrophages | BSK_hDFCGF_MCSF_down | ✓ | ✓ | ✓ | ||||
| BSK_LPS_MCSF_down | ✓ | ✓ | ||||||
| CXCL10 (chemokine (C-X-C motif) ligand 10) Chemokine involved in the stimulation of monocytes, NK cells, and T cells | BSK_BE3C_IP10_down | ✓ | ✓ | ✓ | ✓ | |||
| BSK_hDFCGF_IP10_down | ✓ | ✓ | ||||||
| BSK_KF3CT_IP10_down | ✓ | ✓ | ✓ | |||||
| CXCL8 (chemokine (C-X-C motif) ligand 8) Chemokine secreted by macrophages, neutrophils, eosinophils, and T cells | BSK_hDFCGF_IL8_down | ✓ | ✓ | |||||
| BSK_LPS_IL8_down | ✓ | |||||||
| BSK_SAg_IL8_down | ✓ | |||||||
| CXCL9 (chemokine (C-X-C motif) ligand 9) Chemokine involved in immune regulation and inflammatory processes | BSK_BE3C_MIG_down | ✓ | ✓ | |||||
| BSK_hDFCGF_MIG_down | ✓ | ✓ | ||||||
| BSK_SAg_MIG_down | ✓ | |||||||
| HLA-DRA (major histocompatibility complex class II) Antigen-presenting molecule | BSK_3C_HLADR_down | ✓ | ||||||
| BSK_BE3C_HLADR_down | ✓ | ✓ | ✓ | ✓ | ||||
| ICAM1 (intercellular adhesion molecule 1) Cell adhesion molecule | BSK_KF3CT_ICAM1_down | ✓ | ||||||
| IL-1α (Interleukin-1, alpha) Cytokine produced by macrophages | BSK_BE3C_IL1a_down | ✓ | ✓ | ✓ | ||||
| BSK_KF3CT_IL1a_down | ✓ | ✓ | ||||||
| BSK_LPS_IL1a_down | ✓ | |||||||
| LTB4R (leukotriene B4 receptor) Transmembrane receptor on immune cells | NVS_GPCR_gLTB4 | ✓ | ✓ | ✓ | ||||
| E-selectin Cell adhesion molecule | BSK_LPS_Eselectin_down | ✓ | ✓ * | |||||
| BSK_SAg_Eselectin_down | ✓ | |||||||
| P-selectin Cell adhesion molecule | BSK_4H_Pselectin_down | ✓ | ||||||
| Prostaglandin E receptor 2 Transmembrane receptor on immune cells | BSK_LPS_PGE2_down | ✓ | ✓ | |||||
| TGF-β1 (transforming growth factor, beta 1) Growth factor that regulates immune responses | BSK_BE3C_TGFb1_down | ✓ | ✓ | ✓ | ||||
| BSK_KF3CT_TGFb1_down | ✓ | ✓ | ✓ | |||||
| TNF (tumor necrosis factor) Pro-inflammatory cytokine primarily secreted by macrophages | BSK_LPS_TNFa_down | ✓ | ||||||
| VCAM1 (vascular cell adhesion molecule 1) Cell adhesion molecule | BSK_3C_VCAM1_down | ✓ | ||||||
| BSK_4H_VCAM1_down | ✓ | |||||||
| BSK_hDFCGF_VCAM1_down | ✓ | ✓ | ||||||
| BSK_LPS_VCAM1_down | ✓ |
| Gene Name and Function | ToxCast Assay Name | TBHQ | FD&C Red 3 | PFOS | PFDA | PFUnDA |
|---|---|---|---|---|---|---|
| Coagulation factor III Involved in the innate immune response and host defense against infection, initiates the coagulation cascades | BSK_LPS_TissueFactor_down | ✓ | ||||
| Matrix metallopeptidase 1 Enzyme that breaks down extracellular matrix; involved in the immune response to infection | BSK_BE3C_MMP1_down | ✓ | ||||
| BSK_hDFCGF_MMP1_down | ✓ | ✓ | ✓ | |||
| Matrix metallopeptidase 9 Enzyme that breaks down extracellular matrix; involved in the immune response to infection | BSK_KF3CT_MMP9_down | ✓ | ✓ | ✓ | ✓ | |
| Tissue plasminogen activator Secreted serine protease that converts the plasminogen to plasmin, regulates innate immune response | BSK_BE3C_tPA_down | ✓ | ✓ | ✓ | ||
| Urokinase-type plasminogen activator Secreted serine protease that converts the plasminogen to plasmin, regulates innate immune response | BSK_BE3C_uPA_down | ✓ | ✓ | |||
| BSK_KF3CT_uPA_down | ✓ | ✓ | ||||
| Urokinase-type plasminogen activator receptor Regulates inflammatory, immune, and coagulation responses | BSK_3C_uPAR_down | ✓ | ||||
| BSK_BE3C_uPAR_down | ✓ | |||||
| BSK_CASM3C_uPAR_down | ✓ | |||||
| SERPINE1 (serpin peptidase inhibitor, clade E) Involved in the innate immune response and early host defense against infection, an inhibitor of fibrinolysis | BSK_BE3C_PAI1_down | ✓ | ✓ | ✓ | ||
| BSK_hDFCGF_PAI1_down | ✓ | ✓ | ✓ | |||
| Thrombomodulin Transmembrane receptor, suppresses coagulation and inflammation | BSK_CASM3C_Thrombomodulin_down | ✓ | ||||
| BSK_CASM3C_Thrombomodulin_up | ✓ | |||||
| TIMP metallopeptidase inhibitor 1 Inhibitor of the enzymes that break down extracellular matrix; regulates innate immune response | BSK_hDFCGF_TIMP1_down | ✓ | ✓ | ✓ | ||
| TIMP metallopeptidase inhibitor 2 Inhibitor of the enzymes that break down extracellular matrix; regulates innate immune response | BSK_KF3CT_TIMP2_down | ✓ | ✓ |
| Gene Name and Function | ToxCast Assay Name | TBHQ | FD&C Red 3 | 6:2 FTOH | PFOS | PFOA | PFUnDA |
|---|---|---|---|---|---|---|---|
| Aryl hydrocarbon receptor (AhR) Involved in xenobiotic response | ATG_Ahr_CIS_up | ✓ | |||||
| OX21_AhR_LUC_Agonist * | ✓ | ||||||
| Nuclear factor, erythroid 2-like 2 (NFE2L2, Nrf2) Involved in oxidative stress, inflammation, and injury | ATG_NRF2_ARE_CIS_up | ✓ | ✓ | ✓ | ✓ | ||
| TOX21_ARE_BLA_agonist_ratio | ✓ | ✓ | |||||
| Glucocorticoid receptor (nuclear receptor subfamily 3, group C, member 1) Involved in regulation of stress response, inflammation, and immune processes | NVS_NR_hGR | ✓ | ✓ | ||||
| TOX21_GR_BLA_Antagonist_ratio | ✓ |
| TBHQ Target Reported in Immunological Studies | Studies Reporting This Target | ToxCast Assay Direction |
|---|---|---|
| Increased activity of Nrf2 | [45,46,53] | up |
| Decreased activity of NFκB | [46,58] | down * |
| Decreased CD69 expression | [45,57] | down |
| CCL2 increase | [53] | down |
| TNFα decrease | [59] | down |
| IL-6 decrease | [59] | no activity |
| Chemical | ToxCast | Laboratory Animal Studies | Epidemiological Studies | Conclusion |
|---|---|---|---|---|
| FD&C Red 3 | Affects multiple immune parameters | No studies identified | No studies identified | Potential for immunotoxic effects, should be further investigated |
| TBHQ | Affects multiple immune parameters | Immune modulation, changes in the immune functions | No studies identified | Immunological and mechanistic studies point to risk for the immune system |
| PFUnDA | Affects multiple immune parameters | Some evidence of immune suppression | Immune suppression | Human data and mechanistic studies point to risk for the immune system |
| PFOA | Does not show strong activity in ToxCast assays with immune targets | Immune suppression | Immune suppression | Human data point to risk for the immune system with limited support from mechanistic studies |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naidenko, O.V.; Andrews, D.Q.; Temkin, A.M.; Stoiber, T.; Uche, U.I.; Evans, S.; Perrone-Gray, S. Investigating Molecular Mechanisms of Immunotoxicity and the Utility of ToxCast for Immunotoxicity Screening of Chemicals Added to Food. Int. J. Environ. Res. Public Health 2021, 18, 3332. https://doi.org/10.3390/ijerph18073332
Naidenko OV, Andrews DQ, Temkin AM, Stoiber T, Uche UI, Evans S, Perrone-Gray S. Investigating Molecular Mechanisms of Immunotoxicity and the Utility of ToxCast for Immunotoxicity Screening of Chemicals Added to Food. International Journal of Environmental Research and Public Health. 2021; 18(7):3332. https://doi.org/10.3390/ijerph18073332
Chicago/Turabian StyleNaidenko, Olga V., David Q. Andrews, Alexis M. Temkin, Tasha Stoiber, Uloma Igara Uche, Sydney Evans, and Sean Perrone-Gray. 2021. "Investigating Molecular Mechanisms of Immunotoxicity and the Utility of ToxCast for Immunotoxicity Screening of Chemicals Added to Food" International Journal of Environmental Research and Public Health 18, no. 7: 3332. https://doi.org/10.3390/ijerph18073332
APA StyleNaidenko, O. V., Andrews, D. Q., Temkin, A. M., Stoiber, T., Uche, U. I., Evans, S., & Perrone-Gray, S. (2021). Investigating Molecular Mechanisms of Immunotoxicity and the Utility of ToxCast for Immunotoxicity Screening of Chemicals Added to Food. International Journal of Environmental Research and Public Health, 18(7), 3332. https://doi.org/10.3390/ijerph18073332