
Impact of Food Additive Titanium Dioxide on Gut Microbiota Composition, Microbiota-Associated Functions, and Gut Barrier: A Systematic Review of In Vivo Animal Studies

 ,
,  , ,
, , 

Abstract
1. Introduction
2. Methods
2.1. Eligibility Criteria
2.2. Data Sources and Search Strategy
2.3. Study Selection
2.4. Data Extraction and Reporting
2.5. Quality Assessment
3. Results
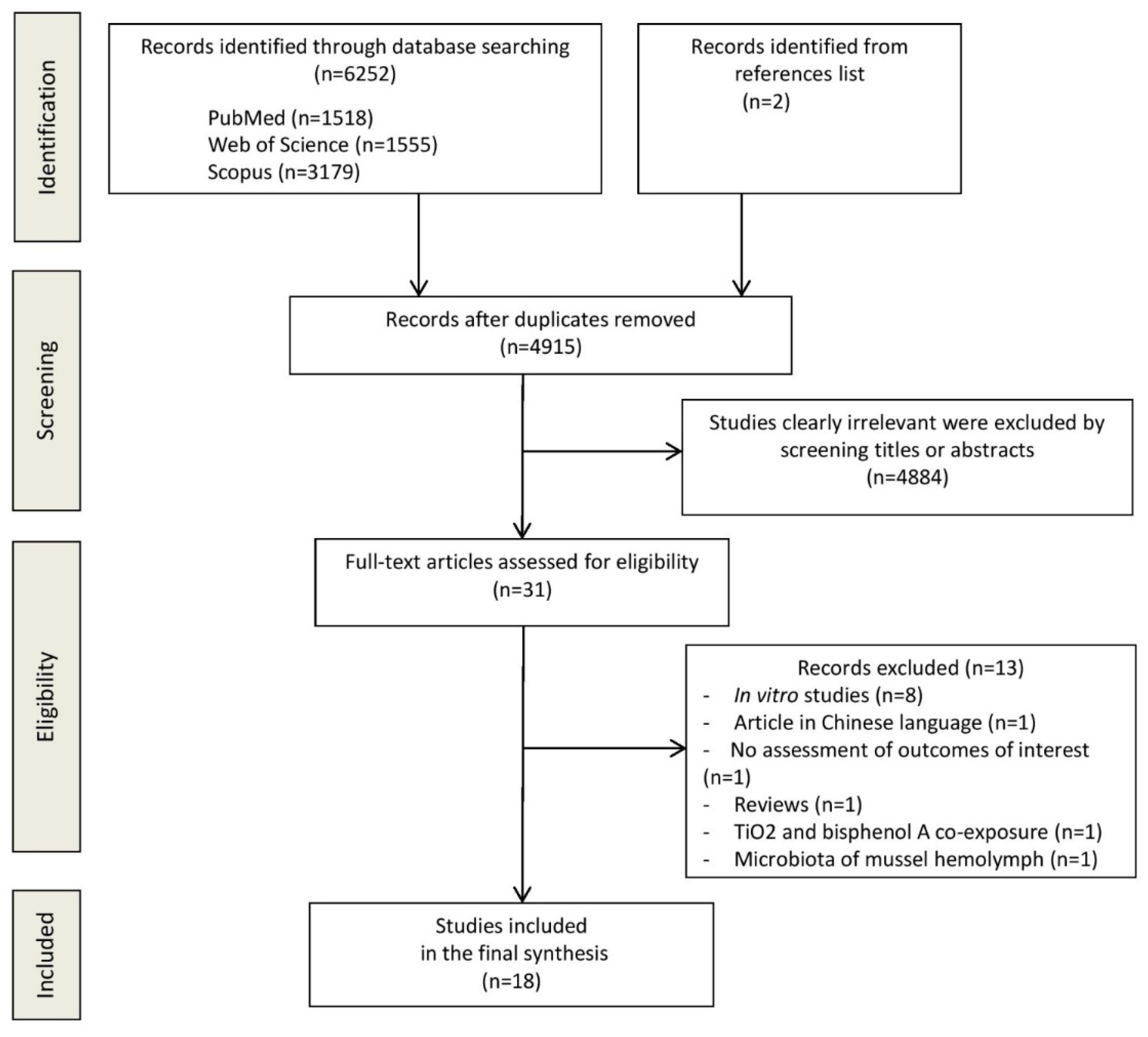
3.1. Study Selection
3.2. Study Characteristics
3.3. Quality Assessment
3.4. Results
3.4.1. TiO2 and gut Microbial Diversity
3.4.2. TiO2 and Abundance of Individual Microbial Species
3.4.3. TiO2 and SCFAs Levels
3.4.4. TiO2 and Metabolism
3.4.5. TiO2 and Gut Barrier Permeability
3.4.6. TiO2 and Inflammatory Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martirosyan, A.; Schneider, Y.-J. Engineered nanomaterials in food: Implications for food safety and consumer health. Int. J. Environ. Res. Public Health 2014, 11, 5720–5750. [Google Scholar] [CrossRef]
- Yang, Y.; Doudrick, K.; Bi, X.; Hristovski, K.; Herckes, P.; Westerhoff, P.; Kaegi, R. Characterization of food-grade titanium dioxide: The presence of nanosized particles. Environ. Sci. Technol. 2014, 48, 6391–6400. [Google Scholar] [CrossRef] [PubMed]
- Weir, A.; Westerhoff, P.; Fabricius, L.; Hristovski, K.; Von Goetz, N. Titanium dioxide nanoparticles in food and personal care products. Environ. Sci. Technol. 2012, 46, 2242–2250. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. CFR—Code of Federal Regulations Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfCFR/CFRSearch.cfm?fr=73.575 (accessed on 15 December 2020).
- EFSA ANS Panel (EFSA Panel on Food Additives and Nutrient Sources Added to Food). Scientific opinion on the re-evaluation of titanium dioxide (E 171) as a food additive. EFSA J. 2016, 14, 4545. [Google Scholar]
- NTP (National Toxicology Program). Bioassay of Titanium Dioxide for Possible Carcinogenicity; U.S. Department of Health, Education, and Welfare: Rockville, MD, USA, 1979.
- Heringa, M.B.; Geraets, L.; van Eijkeren, J.C.H.; Vandebriel, R.J.; deJong, W.; Oomen, A.G. Risk assessment of titanium dioxide nanoparticles via oral exposure, including toxicokinetic considerations. Nanotoxicology 2016, 10, 1515–1525. [Google Scholar] [CrossRef]
- Bettini, S.; Boutet-Robinet, E.; Cartier, C.; Coméra, C.; Gaultier, E.; Dupuy, J.; Naud, N.; Taché, S.; Grysan, P.; Reguer, S.; et al. Food-grade TiO2 impairs intestinal and systemic immune homeostasis, initiates preneoplastic lesions and promotes aberrant crypt development in the rat colon. Sci. Rep. 2017, 7, srep40373. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Martucci, N.J.; Moreno-Olivas, F.; Tako, E.; Mahler, G.J. Titanium dioxide nanoparticle ingestion alters nutrient absorption in an in vitro model of the small intestine. NanoImpact 2017, 5, 70–82. [Google Scholar] [CrossRef]
- Proquin, H.; Rodríguez-Ibarra, C.; Moonen, C.G.J.; Urrutia Ortega, I.M.; Briedé, J.J.; De Kok, T.M.; Van Loveren, H.; Chirino, Y.I. Titanium dioxide food additive (E171) induces ROS formation and genotoxicity: Contribution of micro and nano-sized fractions. Mutagenesis 2017, 32, 139–149. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; et al. Evaluation of four new studies on the potential toxicity of titanium dioxide used as a food additive (E 171). EFSA J. 2018, 16, e05366. [Google Scholar] [CrossRef]
- ANSES. Avis de l’Agence Nationale de Securit e Sanitaire de l’Alimentation, de l’environnement et du Travail Relatif au Risques Lies a la Ingestion de l’Additif Alimentaire E 171. Available online: https://www.anses.fr/fr/system/files/ERCA2019SA0036.pdf (accessed on 20 December 2020).
- EFSA (European Food Safety Authority). EFSA statement on the review of the risks related to the exposure to the food additive titanium dioxide (E 171) performed by the French Agency for Food, Environmental and Occupational Health and Safety (ANSES). EFSA J. 2019, 17, e05714. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? a changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Dorier, M.; Béal, D.; Marie-Desvergne, C.; Dubosson, M.; Barreau, F.; Houdeau, E.; Herlin-Boime, N.; Carriere, M. Continuous in vitro exposure of intestinal epithelial cells to E171 food additive causes oxidative stress, inducing oxidation of DNA bases but no endoplasmic reticulum stress. Nanotoxicology 2017, 11, 751–761. [Google Scholar] [CrossRef]
- Dorier, M.; Brun, E.; Veronesi, G.; Barreau, F.; Pernet-Gallay, K.; Desvergne, C.; Rabilloud, T.; Carapito, C.; Herlin-Boime, N.; Carrière, M. Impact of anatase and rutile titanium dioxide nanoparticles on uptake carriers and efflux pumps in Caco-2 gut epithelial cells. Nanoscale 2015, 7, 7352–7360. [Google Scholar] [CrossRef] [PubMed]
- Faust, J.J.; Masserano, B.M.; Mielke, A.H.; Abraham, A.; Capco, D.G. Engineered nanoparticles induced brush border disruption in a human model of the intestinal epithelium. Adv. Exp. Med. Biol. 2014, 811, 55–72. [Google Scholar] [PubMed]
- Song, B.; Liu, J.; Feng, X.; Wei, L.; Shao, L. A review on potential neurotoxicity of titanium dioxide nanoparticles. Nanoscale Res. Lett. 2015, 10, 342. [Google Scholar] [CrossRef] [PubMed]
- Pedata, P.; Ricci, G.; Malorni, L.; Venezia, A.; Cammarota, M.; Volpe, M.; Iannaccone, N.; Guida, V.; Schiraldi, C.; Romano, M.; et al. In vitro intestinal epithelium responses to titanium dioxide nanoparticles. Food Res. Int. 2019, 119, 634–642. [Google Scholar] [CrossRef]
- Kolba, N.; Guo, Z.; Olivas, F.M.; Mahler, G.J.; Tako, E. Intra-amniotic administration (Gallus gallus) of TiO2, SiO2, and ZnO nanoparticles affect brush border membrane functionality and alters gut microflora populations. Food Chem. Toxicol. 2020, 135, 110896. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and me-ta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Hooijmans, C.R.; Rovers, M.M.; De Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Limage, R.; Tako, E.; Kolba, N.; Guo, Z.; García-Rodríguez, A.; Marques, C.N.H.; Mahler, G.J. TiO2 nanoparticles and com-mensal bacteria alter mucus layer thickness and composition in a gastrointestinal tract model. Small 2020, 16, e2000601. [Google Scholar] [CrossRef]
- Dudefoi, W.; Moniz, K.; Allen-Vercoe, E.; Ropers, M.H.; Walker, V.K. Impact of food-grade and nano-TiO2 particles on a human intestinal community. Food Chem. Toxicol. 2017, 106, 242–249. [Google Scholar] [CrossRef]
- Agans, R.T.; Gordon, A.; Hussain, S.; Paliy, O. Titanium dioxide nanoparticles elicit lower direct inhibitory effect on human gut microbiota than silver nanoparticles. Toxicol. Sci. 2019, 172, 411–416. [Google Scholar] [CrossRef]
- Dorier, M.; Béal, D.; Tisseyre, C.; Marie-Desvergne, C.; Dubosson, M.; Barreau, F.; Houdeau, E.; Herlin-Boime, N.; Rabilloud, T.; Carriere, M. The food additive E171 and titanium dioxide nanoparticles indirectly alter the homeostasis of human intestinal epithelial cells in vitro. Environ. Sci. Nano 2019, 6, 1549–1561. [Google Scholar] [CrossRef]
- Taylor, A.A.; Marcus, I.M.; Guysi, R.L.; Walker, S.L. metal oxide nanoparticles induce minimal phenotypic changes in a model colon gut microbiota. Environ. Eng. Sci. 2015, 32, 602–612. [Google Scholar] [CrossRef]
- Waller, T.; Chen, C.; Walker, S.L. Food and industrial grade titanium dioxide impacts gut microbiota. Environ. Eng. Sci. 2017, 34, 537–550. [Google Scholar] [CrossRef]
- Hong, F.; Zhou, Y.; Zhao, X.; Sheng, L.; Wang, L. Maternal exposure to nanosized titanium dioxide suppresses embryonic development in mice. Int. J. Nanomed. 2017, 12, 6197–6204. [Google Scholar] [CrossRef] [PubMed]
- Auguste, M.; Lasa, A.; Pallavicini, A.; Gualdi, S.; Vezzulli, L.; Canesi, L. Exposure to TiO2 nanoparticles induces shifts in the microbiota composition of Mytilus galloprovincialis hemolymph. Sci. Total. Environ. 2019, 670, 129–137. [Google Scholar] [CrossRef]
- Mercier-Bonin, M.; Despax, B.; Raynaud, P.; Houdeau, E.; Thomas, M. Mucus and microbiota as emerging players in gut nanotoxicology: The example of dietary silver and titanium dioxide nanoparticles. Crit. Rev. Food Sci. Nutr. 2018, 58, 1023–1032. [Google Scholar] [CrossRef]
- Han, S.; Chen, Z.; Zhou, D.; Zheng, P.; Zhang, J.; Jia, G. Effects of titanium dioxide nanoparticles on fecal metabolome in rats after oral administration for 90 days. J. Peking Univ. 2020, 52, 457–463. [Google Scholar]
- Chen, L.; Guo, Y.; Hu, C.; Lam, P.K.; Lam, J.C.; Zhou, B. Dysbiosis of gut microbiota by chronic coexposure to titanium dioxide nanoparticles and bisphenol A: Implications for host health in zebrafish. Environ. Pollut. 2018, 234, 307–317. [Google Scholar] [CrossRef]
- Cao, X.; Han, Y.; Gu, M.; Du, H.; Song, M.; Zhu, X.; Ma, G.; Pan, C.; Wang, W.; Zhao, E.; et al. Foodborne titanium dioxide nanoparticles induce stronger adverse effects in obese mice than non-obese mice: Gut microbiota dysbiosis, colonic inflammation, and proteome alterations. Small 2020, 16, e2001858. [Google Scholar] [CrossRef]
- Chen, Z.; Han, S.; Zhou, D.; Zhou, S.; Jia, G. Effects of oral exposure to titanium dioxide nanoparticles on gut microbiota and gut-associated metabolism in vivo. Nanoscale 2019, 11, 22398–22412. [Google Scholar] [CrossRef]
- Chen, Z.; Han, S.; Zheng, P.; Zhou, D.; Zhou, S.; Jia, G. Effect of oral exposure to titanium dioxide nanoparticles on lipid me-tabolism in Sprague-Dawley rats. Nanoscale 2020, 12, 5973–5986. [Google Scholar] [CrossRef]
- Duan, Y.; Liu, J.; Ma, L.; Li, N.; Liu, H.; Wang, J.; Zheng, L.; Liu, C.; Wang, X.; Zhao, X.; et al. Toxicological characteristics of nanoparticulate anatase titanium dioxide in mice. Biomaterials 2010, 31, 894–899. [Google Scholar] [CrossRef]
- Kurtz, C.C.; Mitchell, S.; Nielsen, K.; Crawford, K.D.; Mueller-Spitz, S.R. Acute high-dose titanium dioxide nanoparticle ex-posure alters gastrointestinal homeostasis in mice. J. Appl. Toxicol. 2020, 40, 1384–1395. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, S.; Lei, R.; Gu, W.; Qin, Y.; Ma, S.; Chen, K.; Chang, Y.; Bai, X.; Xia, S.; et al. Oral administration of rutile and anatase TiO2 nanoparticles shifts mouse gut microbiota structure. Nanoscale 2018, 10, 7736–7745. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, F.; Lu, Z.; Fang, Y.; Qu, J.; Mao, T.; Wang, H.; Chen, J.; Li, B. Effects of TiO2 nanoparticles on intestinal microbial composition of silkworm, Bombyx mori. Sci. Total. Environ. 2020, 704, 135273. [Google Scholar] [CrossRef]
- Liu, L.-Y.; Sun, L.; Zhong, Z.-T.; Zhu, J.; Song, H.-Y. Effects of titanium dioxide nanoparticles on intestinal commensal bacteria. Nucl. Sci. Tech. 2016, 27, 1–5. [Google Scholar] [CrossRef]
- Mao, Z.; Li, Y.; Dong, T.; Zhang, L.; Zhang, Y.; Li, S.; Hu, H.; Sun, C.; Xia, Y. Exposure to titanium dioxide nanoparticles during pregnancy changed maternal gut microbiota and increased blood glucose of rat. Nanoscale Res. Lett. 2019, 14, 26. [Google Scholar] [CrossRef]
- Mu, W.; Wang, Y.; Huang, C.; Fu, Y.; Li, J.; Wang, H.; Jia, X.; Ba, Q. Effect of Long-Term Intake of Dietary Titanium Dioxide Nanoparticles on Intestine Inflammation in Mice. J. Agric. Food Chem. 2019, 67, 9382–9389. [Google Scholar] [CrossRef] [PubMed]
- Pinget, G.; Tan, J.; Janac, B.; Kaakoush, N.O.; Angelatos, A.S.; O’Sullivan, J.; Koay, Y.C.; Sierro, F.; Davis, J.; Divakarla, S.K.; et al. Impact of the food additive titanium dioxide (E171) on gut microbiota-host interaction. Front. Nutr. 2019, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.W.; Shull, G.M.; Fountain, J.H.; Guo, Z.; Musselman, L.P.; Fiumera, A.C.; Mahler, G.J. Titanium dioxide nanoparticle exposure alters metabolic homeostasis in a cell culture model of the intestinal epithelium and Drosophila melanogaster. Nanotoxicology 2018, 12, 390–406. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, P.A.; Morón, B.; Becker, H.M.; Lang, S.; Atrott, K.; Spalinger, M.R.; Scharl, M.; Wojtal, K.A.; Fischbeck-Terhalle, A.; Frey-Wagner, I.; et al. Titanium dioxide nanoparticles exacerbate DSS-induced colitis: Role of the NLRP3 inflammasome. Gut 2017, 66, 1216–1224. [Google Scholar] [CrossRef]
- Talbot, P.; Radziwill-Bienkowska, J.M.; Kamphuis, J.B.J.; Steenkeste, K.; Bettini, S.; Robert, V.; Noordine, M.-L.; Mayeur, C.; Gaultier, E.; Langella, P.; et al. Food-grade TiO2 is trapped by intestinal mucus in vitro but does not impair mucin O-glycosylation and short-chain fatty acid synthesis in vivo: Implications for gut barrier protection. J. Nanobiotechnol. 2018, 16, 1–14. [Google Scholar] [CrossRef]
- Yan, J.; Wang, D.; Li, K.; Chen, Q.; Lai, W.; Tian, L.; Lin, B.; Tan, Y.; Liu, X.; Xi, Z. Toxic effects of the food additives titanium dioxide and silica on the murine intestinal tract: Mechanisms related to intestinal barrier dysfunction involved by gut micro-biota. Environ. Toxicol. Pharmacol. 2020, 80, 103485. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, X.; Cheng, S.; Fan, J.; Qin, X.; Wang, T.; Zhang, Y.; Zhang, J.; Qiu, Y.; Qiu, J.; et al. Titanium dioxide nanoparticles via oral exposure leads to adverse disturbance of gut microecology and locomotor activity in adult mice. Arch. Toxicol. 2020, 94, 1173–1190. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, L.; Liu, Z.; Zhou, Q.; Zhu, Y.; Zhao, Y.; Yang, X. Long-term exposure to titanium dioxide nanoparticles promotes diet-induced obesity through exacerbating intestinal mucus layer damage and microbiota dysbiosis. Nano Res. 2020, 1–11. [Google Scholar] [CrossRef]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of diet on the gut microbiota: Rethinking intervention duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef]
- Lamas, B.; Martins Breyner, N.; Houdeau, E. Impacts of foodborne inorganic nanoparticles on the gut microbiota-immune axis: Potential consequences for host health. Part. Fibre Toxicol. 2020, 17, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Rodríguez-Piñeiro, A.M.; Schütte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Bäckhed, F.; Hansson, G.C.; Johansson, M.E.V. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015, 16, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.; Jakobsson, H.E.; Holmén-Larsson, J.; Schütte, A.; Ermund, A.; Rodríguez-Piñeiro, A.M.; Arike, L.; Wising, C.; Svensson, F.; Bäckhed, F.; et al. Normalization of host intestinal mucus layers requires long-term microbial colonization. Cell Host Microbe 2015, 18, 582–592. [Google Scholar] [CrossRef]
- Becker, S.; Oelschlaeger, T.A.; Wullaert, A.; Vlantis, K.; Pasparakis, M.; Wehkamp, J.; Stange, E.F.; Gersemann, M. Bacteria regulate intestinal epithelial cell differentiation factors both in vitro and in vivo. PLoS ONE 2013, 8, e55620. [Google Scholar]
- Brun, E.; Barreau, F.; Veronesi, G.; Fayard, B.; Sorieul, S.; Chanéac, C.; Carapito, C.; Rabilloud, T.; Mabondzo, A.; Herlin-Boime, N.; et al. Titanium dioxide nanoparticle impact and translocation through ex vivo, in vivo and in vitro gut epithelia. Part. Fibre Toxicol. 2014, 11, 13. [Google Scholar] [CrossRef]
- Coombes, J.L.; Maloy, K.J. Control of intestinal homeostasis by regulatory T cells and dendritic cells. Semin. Immunol. 2007, 19, 116–126. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Gasbarrini, A.; Mele, M.C. Food additives, gut microbiota, and irritable Bowel syndrome: A hidden track. Int. J. Environ. Res. Public Heal. 2020, 17, 8816. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-X.; Cheng, B.; Yang, Y.-X.; Cao, A.; Liu, J.-H.; Du, L.-J.; Liu, Y.; Zhao, Y.; Wang, H. Characterization and preliminary toxicity assay of nano-titanium dioxide additive in sugar-coated chewing gum. Small 2013, 9, 1765–1774. [Google Scholar] [CrossRef]
- Rompelberg, C.; Heringa, M.B.; Van Donkersgoed, G.; Drijvers, J.; Roos, A.; Westenbrink, S.; Peters, R.; Van Bemmel, G.; Brand, W.; Oomen, A.G. Oral intake of added titanium dioxide and its nanofraction from food products, food supplements and toothpaste by the Dutch population. Nanotoxicology 2016, 10, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Alvarez, A.-S.; De Vos, W.M. The gut microbiota in the first decade of life. Trends Microbiol. 2019, 27, 997–1010. [Google Scholar] [CrossRef]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Lopetuso, L.R.; Scaldaferri, F.; Pulcini, G.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. Food components and dietary habits: Keys for a healthy gut microbiota composition. Nutrients 2019, 11, 2393. [Google Scholar] [CrossRef] [PubMed]
- Bachler, G.; Von Goetz, N.; Hungerbuhler, K. Using physiologically based pharmacokinetic (PBPK) modeling for dietary risk assessment of titanium dioxide (TiO2) nanoparticles. Nanotoxicology 2014, 9, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Criteria | Definition |
|---|---|
| Participants | Adult animals |
| Exposure | TiO2 NPs (rutile or anastase forms, with any size of nanoparticles) |
| Comparator | Any comparator |
| Outcomes | Primary outcomes
|
| Study design | Peer-reviewed original animal experimental studies |
| Animal Species | First Author, Year | Sex | Age * | Weight * | Sample Size | TiO2 Particules Type and Size | Dose Exposure and Administration Route | Duration of Exposure | Significant Compositional Changes of Gut Microbiota (Compared with the Control Group) | Significant Effects on Microbiota-Associated Functions (Compared with the Control Group) |
|---|---|---|---|---|---|---|---|---|---|---|
| Mice Obese and non-obese C57BL/6 | Cao, 2020 [35] | M | 6 weeks | n.r. | N = 20 • low-fat diet (control) n = 5 • high fat diet n = 5 • high-fat diet + E171 n = 5 • high-fat diet + TiO2 NPs n = 5 | E171, 112 nm TiO2 NPs, 33 nm | 0.1 weight percent | 8 weeks | In mice treated with TiO2 NPs + high-fat diet • ↑ Firmicutes • ↓ Actinobacteria and Bacteroidetes • ↓ Bifidobacterium, Allobaculum and Lactobacillus • ↑ Oscillospira | In mice treated with TiO2 NPs + high-fat diet: • ↓ SCFAs production • Loss of goblet cells and crypts • ↑ IL-12 • ↑ IL-17 |
| Mice CD-1 (ICR) | Duan, 2010 [38] | F | n.r. | 22 ± 2 g | N = 80 • control group (n = 20) • 62.5 mg/kg bw TiO2 (n = 20) • 125 mg/kg bw TiO2 (n = 20) • 250 mg/kg bw TiO2 (n = 20) | Anatase TiO2 NPs 5 nm | 62.5, 125, 250 mg/ kg bw/ day TiO2 NPs via intragastric administration | 30 days | • ↑ IL-2 activity in the exposed groups in a dose-dependent manner • ↓ CD3, CD4, and CD8 in the group treated with 250 mg/kg bwTiO2, • ↓ B cells and NK cells in all exposure groups | |
| Mice C57/BL/6 | Kurtz, 2020 [39] | F | 7–8 weeks | n.r. | N = 48 • control group (n = 16) • TiO2 NPs (n = 32) | TiO2 NPs with irregular shapes | 1 g/kg bw TiO2 NPs by oral gavage | 24 h, 48 h, 7 days, and 14 days | • ↑ Firmicutes in the ileum at 14 days post-exposure • ↓ Lactobacillus spp. at 24 h | • ↑ SCFAs production in stools • Higher body weight • ↑ mucus production from 48 h post gavage to 7 days • ↑ IL-4 levels at 24 h |
| Mice C57BL/6 | Li, 2018 [40] | M | 8 weeks | 22–26 g | N = 30 • control group (n = 10) • rutile TiO2 NPs (n = 10) • anatase TiO2 NPs (n = 10) | Anatase NPs in water 20.13 ± 0.18 nm Rutile NPs in water 15.91 ± 0.05 nm | 100 mg/kg bw/day by oral gavage | 28 days | • No decrease in overall microbiota diversity (Chao1 index, Shannon index, Simpson diversity index) • Shift of microbiota composition in a time-dependent manner • ↑ Proteobacteria by rutile TiO2 NPs but not by anatase TiO2 NPs • ↓ Prevotella by both TiO2 NPs • ↑ Rhodococcus, Escherichia-Shigella by rutile TiO2 NPs • ↑ Bacteroides, Akkermansia by anatase TiO2 NPs | In rutile TiO2 NPs—exposed mice: • Longer intestinal villi • Irregular arrangement of villus epithelial cells. |
| Mice C57BL/6J | Mu, 2019 [44] | F | 3 weeks | n.r. | N = 20 • control group (n = 5) • NP10 (n = 5) • NP50 (n = 5) • NP100 (n = 5) | TiO2 NPs were added to the diet as an ingredient in the feed preparation process NP10: anatase; 10 nm NP50: anastase 50 nm NP100: anastase 100 nm | Diet containing 0.1% TiO2 NPs | 3 months | • No significant bacterial diversity changes • ↑ Bacteroidetes in NP10 and NP50 treatment groups • ↓ Actinobacteria in NP10 and NP50 treatment groups • ↓ Lactobacillus and Bifidobacterium in NP10 and NP50 treatment groups | • Lower body weight in mice fed with NP10 and NP50 for 3 months • ↑ LCN2 levels in stools (a marker for intestinal inflammation) in NP10 and NP50-treated groups • Aggravation of DSS-induced chronic colitis • Aggravation of immune response • ↓ CD4 + T cells, regulatory T cells, and macrophages |
| Mice C57BL/6Jausb | Pinget, 2019 [45] | M | 5–6 weeks | n.r. | N = 24 • control group (n = 6) • 2 mg TiO2/kg bw/day (n = 6) • 10 mg TiO2/kg bw/day (n = 6) • 50 mg TiO2/kg bw/day (n = 6) | Ti02 NPs, 28 to 1158 nm | 2, 10, 50 mg TiO2/kg bw/day orally administrated via drinking water | 3 weeks | • Limited variations of bacterial diversity (Simpson, Shannon analyses), bacterial richness, and evenness in all exposed groups (although these trended toward decrease with increasing dose of TiO2) • ↑ Parabacteroides in TiO2-treated mice, at a dose of 50 mg TiO2/kg bw/day • ↑ Lactobacillus and Allobaculum in all exposed groups • ↓ Adlercreutzia and unclassified Clostridiaceae in mice treated with TiO2 at the doses of 10 and 50 mg TiO2/kg bw/day | In mice treated with 50 mg TiO2/kg bw/day: • ↓ SCFAs • ↓ TMA • ↓ crypt length In mice treated with 10 and 50 mg TiO2/kg bw/day: • ↓ MUC2 gene expression • ↑ expression of the β defensin gene • Infiltration of CD8+ T cells • ↑ production of macrophages (CD45 + F4/80 + CD8−Ly6 g−Iab +CD11b+ CD103−) • ↑inflammatory cytokines (IL-6, TNF-α and IL-10) |
| Mice Wild-type (C57BL/6J) and NLRP3-deficient | Ruiz, 2016 [47] | F | 12–14 weeks | n.r. | N = 56 WT mice (n = 36) • water (n = 12) • 50 mg /day/kg body weight of TiO2 NPs (n = 12) • 500 mg /day/kg body weight of TiO2 NPs (n = 12) NLRP3-deficient mice (n = 20) • water (n = 10) • 500 mg /day/ kg body weight of TiO2 NPs (n = 10) | Suspension of TiO2 rutile NPs, 30–50 nm | 50 and 500 mg/day/kg bw of rutileTiO2 NPs by oral gavage | 8 days | • TiO2 NPs enhance intestinal inflammation in the DSS mouse model of colitis. • TiO2 proinflammatory effects required NLRP3 inflammasome activation when comparing WT with NLRP3-deficient mice | |
| Mice ICR | Yan, 2020 [49] | M | Adult | 18–20 g | N = 28 • control group (n = 7) • 10 mg/kg bw/day TiO2 NPs (n = 7) • 40 mg/kg bw/day TiO2 NPs (n = 7) • 160 mg/kg bw/day TiO2 NPs (n = 7) | Anatase TIO2 NPs, 20 nm | 10, 40, 160 mg/ kg bw/day by oral gavage | 28 days | • ↑ Firmicutes in all exposed groups • ↓ Verrucomicrobia in all exposed groups • ↓ Bacteroidetes at 160 mg/kg bw/day TiO2 • ↓ Barnesiella in all exposed groups in a dose-dependent manner • ↓ Akkermansia genus and Porphyromonadaceae family | • ↑ TG and glucose levels in the exposed group with 160 mg/kg bw/day TiO2 • ↑ LPS levels in all exposed groups. • ↑ IL-1α levels in the exposed group with 160 mg/kg bw/day TiO2 • ↑ IL-6 levels in all exposed groups • ↑ TNF-α levels in the exposed groups treated with 40 and 160 mg/kg/day TiO2 •↑ PKC protein at 40 mg/kg • Elevated TLR4 protein levels in the 40 and 160 mg/kg groups and P-P65 in all exposed groups • ↓ MUC2 expression at 160 mg/kg • ↓ intestinal mucus thickness in all exposed groups |
| Mice C57BL/6J | Zhang, 2020 [50] | M | 7 weeks | 20–24 g | N = 30 • control group (n = 15) • TiO2 NPs (n = 15) | TiO2 NPs, 21 nm | 150 mg/kg bw/day by intragastric administration | 30 days | • ↓ Richness and evenness of gut microbiota (decreased Shannon’s diversity, Chao, observed species and elevated Simpson’s diversity) • ↑ Proteobacteria | • No changes in body weight • Abnormal excitement on the enteric neurons • ↑ expression of TuJ1 (a neuronal marker of the peripheral nervous system) • No changes of IL-6 and IL-1β in the gut tissues |
| Mice C57BL/6J | Zhu, 2020 [51] | F | 4–5 weeks | n.r. | N = 24 • control fed with CHOW diet (n = 6) • TiO2 NPs fed with CHOW diet (n = 6) • control fed with HFD (n = 6) • TiO2 NPs fed with HFD (n = 6) | TiO2 NPs, 30 ± 7 nm | 10 μL/g bw/day via oral gavage | 8 weeks | • ↑ Firmicutes to Bacteroidetes ratio in TiO2 NPs treated mice fed with HFD compared with both CHOW group and controls. • ↑ Desulfovibrionaceae in TiO2 NPs treated mice fed with CHOW or HFD compared with controls • ↑ Ruminococcaceae in TiO2 NPs treated mice fed with CHOW diet or HFD compared with controls • ↑ Lachnospiraceae in TiO2 NPs treated mice fed with CHOW compared with CHOW controls. | • ↓ crypt length in TiO2 groups compared with controls • ↓ muc2 expression in TiO2 groups compared with controls • ↓ MUC2 proteins levels in TiO2 groups compared with controls • ↑ LPS levels in TiO2 groups compared with controls with a significant increase in TIO2 mice fed HFD compared with TIO2 mice fed CHOW. • ↑ IL-1β, IL-6, and TNFα in TiO2 groups compared with controls with a significant increase in TIO2 mice fed HFD compared with TIO2 mice fed CHOW |
| Rats Wistar | Bettini, 2017 [8] | M | adult | 175–200 g | First series of experiments N = 30 • control group (n = 10) • food-grade E171 (n = 10) • NM-105 (n = 10) Second series of experiments N = 34 • water (n = 11) treated with DMH • food-grade E171 (n = 11) treated with DMH • control (n = 12) water only | NM-105: Ti02 NPs, 105 nm | 10 mg/kg bw/day by intragastric gavage 10 mg/ kg bw/ day TiO2 NPs through drinking water | 7 days 100 days | • No changes in epithelium permeability • Accumulation of dendritic cells in the immune cells of Peyer’s patches regardless of the TiO2 treatment • ↓ regulatory T cells • ↑ IL8, IL10, TNFα in food-grade E171 group after 100 days • At 7 days, no intestinal inflammation in E171 and NM-105 groups • Initiation of colon inflammation and pre-neoplastic lesions in the 100-day E171 group. | |
| Rats Sprague-Dawley | Chen, 2019 [36] | M | 3 weeks | n.r. | N = 12 • control group (n = 6) • TiO2 NPs (n = 6) | Anatase TiO2 NPs, 29 ± 9 nm | 2, 10, 50 mg/ kg bw/ day TiO2 NPs via oral gavage | 30 days | • ↑ L. gasseri in the high-dose group • ↑ Turicibacter in the low-dose group • ↑ L. NK4A136_group in the medium-dose group • ↓ Veillonella in all exposure groups | • Increase of N-acetylhistamine, caprolactam, and glycerophosphocholine • ↓ 4-methyl-5-thiazoleethanol, L-histidine, and L-ornithine • No significant changes in SCFAs levels • ↑ LPS production • ↑ IL-6 in the high-dose group • ↑ intestinal oxidative stress and inflammatory response |
| Rats Sprague-Dawley | Chen, 2020 [37] | M | 3 weeks | n.r. | N = 12 • control group (n = 6) • TiO2 NPs (n = 6) | Anatase TiO2 NPs, 29 ± 9 nm | 2, 10, 50 mg/ kg bw/ day TiO2 NPs via oral gavage | 90 days | • No significant changes in SCFAs levels • ↓ TG levels in medium and high-dose groups • Significant serum lipophilic metabolites changes in the high-dose group with ↑ phosphatidylcholines and ↓ lysophosphatidylcholine and glycerophosphocholine levels • ↓ activity of the antioxidant enzyme SOD | |
| Rats Pregnant Sprague-Dawley | Mao, 2019 [43] | F | 12 weeks | n.r. | N = 8 • control group (n = 4) • TiO2 NPs (n = 4) | TiO2 NPs, 21 nm | 5 mg/kg bw/day of TiO2 NPs | from the 5th to 18th day after pregnancy | • No significant changes of alpha-diversity (although the increasing trend in Shannon, and a significant change in Simpson index but no difference in Chao1) • ↓ Ellin607 at GD 10 and GD 17 • Increase of Clostridiales at GD 10 • ↓ Dehalobacteriaceae at GD 17 | • ↑ fasting blood glucose levels at GD 10 and GD 17 after exposure. • Strengthened genes about type 2 diabetes mellitus related function and lipid biosynthesis in the exposure group • Weakened taurine and hypotaurine metabolism in the exposure group |
| Rats Wistar | Talbot, 2018 [48] | M | Adult | 175–200 g | First series of experiments N = 24 • control group (n = 8) • food-grade E171 (n = 8) • NM-105 (n = 8) Second series of experiments N = 30 • water (n = 10) • food-grade E171 (n = 10) • NM-105 (n = 10) | Food-grade E171, TiO2, 364 nm NM-105: TiO2 NPs, 105 nm | 0.1 mg/kg bw/ day intragastric gavage 10 mg/kg bw/day through the drinking water | 7 days 60 days | • No impact on the overall caecal composition of SCFAs (irrespectively of TiO2) • No effect on mucin O-glycosylation (irrespectively of TiO2) • Absence of mucus barrier impairment irrespectively of TiO2) | |
| Fruit flies Drosophila | Liu, 2016 [42] | F | n.r. | n.r. | N = 45 | 10, 50, and 100 nm TiO2 NPs | 1, 2 mg/mL and 200 mg/mL dietary TiO2 NPs of 3 different sizes | 5 days | • No inhibition of the growth of symbiotic bacteria in the gut of Drosophila larva or adults | • No alteration of pupation cycle • No alteration of weight and lipid levels |
| Fruit flies Drosophila Melanogaster | Richter, 2018 [46] | n.r. | 2 to 6 days | n.r. | N = 24 • control group (n = 6) • 5 ppM TiO2 NPs (n = 6) • 50 ppM TiO2 NPs (n = 6) • 500 ppM TiO2 NPs (n = 6) | TiO2 NPs, 30 nm | 5 ppm, 50 ppm, 500 ppm of TiO2 NPs suspended in the food during cooking | From first instar larvae to adulthood | • Alteration of metabolic gut homeostasis with significant changes in pupation, time to pupation, time to emergence, body size, and glucose content. | |
| Larvae of Bombyx mori | Li, 2020 [41] | n.r. | n.r. | n.r. | N = 16 • control group (n = 8) • TiO2 NPs (n = 8) | TiO2 NPs, 6–10 nm | Mulberry leaves soaked in 5 mg/L TiO2 NPs and naturally dried | From the 3rd day of fifth-instar larvae until morning | • ↑ Staphylococcus, Lachnospiraceae, Pseudomonas, Sphingomonas, Kineococcus, Norank_f_bacteroidales, • ↓ Methylobacterium and Serratia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinninella, E.; Cintoni, M.; Raoul, P.; Mora, V.; Gasbarrini, A.; Mele, M.C. Impact of Food Additive Titanium Dioxide on Gut Microbiota Composition, Microbiota-Associated Functions, and Gut Barrier: A Systematic Review of In Vivo Animal Studies. Int. J. Environ. Res. Public Health 2021, 18, 2008. https://doi.org/10.3390/ijerph18042008
Rinninella E, Cintoni M, Raoul P, Mora V, Gasbarrini A, Mele MC. Impact of Food Additive Titanium Dioxide on Gut Microbiota Composition, Microbiota-Associated Functions, and Gut Barrier: A Systematic Review of In Vivo Animal Studies. International Journal of Environmental Research and Public Health. 2021; 18(4):2008. https://doi.org/10.3390/ijerph18042008
Chicago/Turabian StyleRinninella, Emanuele, Marco Cintoni, Pauline Raoul, Vincenzina Mora, Antonio Gasbarrini, and Maria Cristina Mele. 2021. "Impact of Food Additive Titanium Dioxide on Gut Microbiota Composition, Microbiota-Associated Functions, and Gut Barrier: A Systematic Review of In Vivo Animal Studies" International Journal of Environmental Research and Public Health 18, no. 4: 2008. https://doi.org/10.3390/ijerph18042008
APA StyleRinninella, E., Cintoni, M., Raoul, P., Mora, V., Gasbarrini, A., & Mele, M. C. (2021). Impact of Food Additive Titanium Dioxide on Gut Microbiota Composition, Microbiota-Associated Functions, and Gut Barrier: A Systematic Review of In Vivo Animal Studies. International Journal of Environmental Research and Public Health, 18(4), 2008. https://doi.org/10.3390/ijerph18042008