
Partial Replacement of Dietary Fat with Polyunsaturated Fatty Acids Attenuates the Lipopolysaccharide-Induced Hepatic Inflammation in Sprague-Dawley Rats Fed a High-Fat Diet

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments and Diets
2.2. Fatty Acid Composition Analysis of Experimental Diets
2.3. Oral Glucose Tolerance Test (OGTT)
2.4. Fatty Acid Composition Analysis of Whole Blood
2.5. Biochemical Analyses of Serum
2.6. Determination of Lipid Contents in the Liver and Adipose Tissue
2.7. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Fatty Acid Profiles of Experimental Diets and Rat Whole Blood
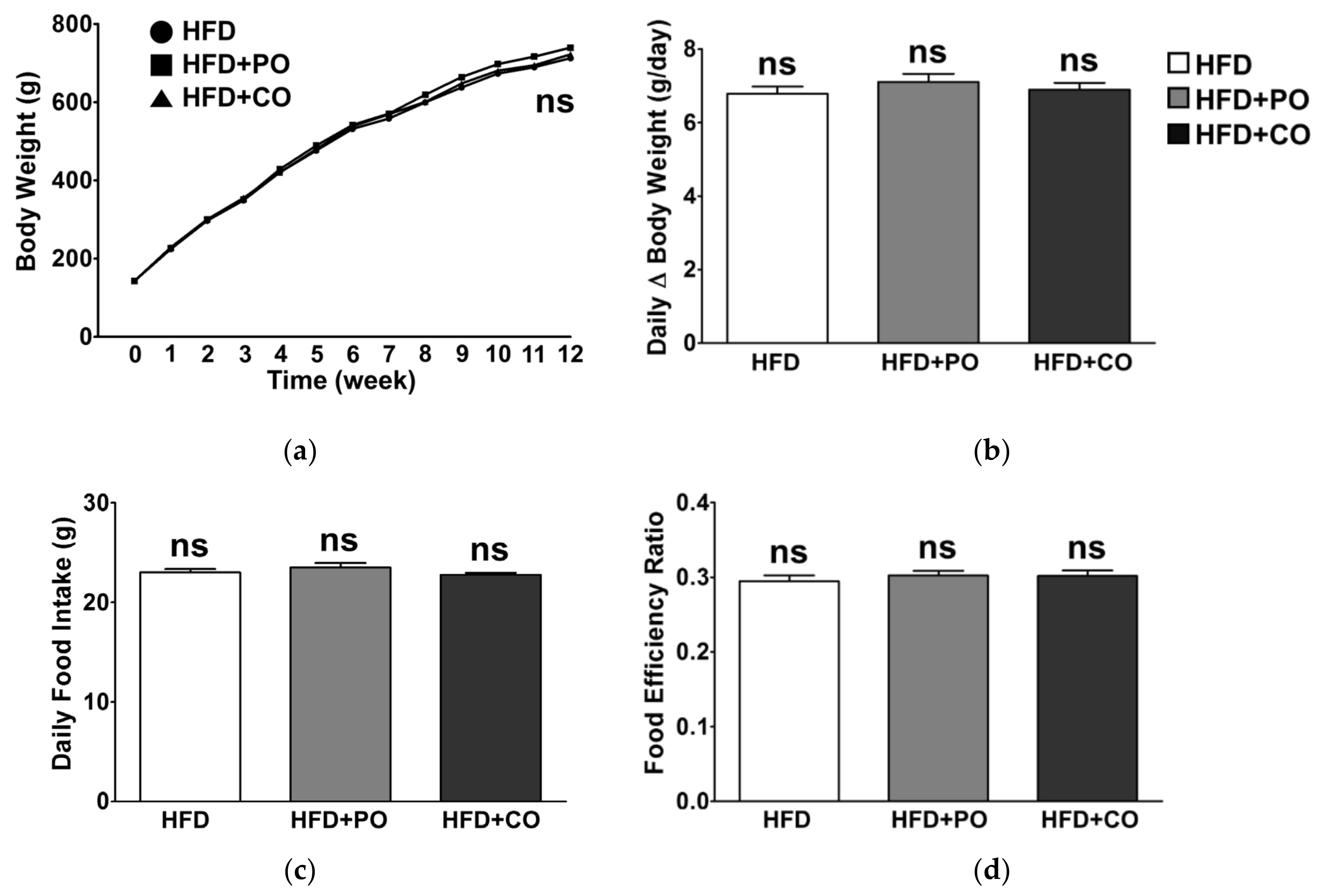
3.2. Partial Replacement of Dietary Fat with PO and CO Did Not Affect the Body Weight and Food Intake
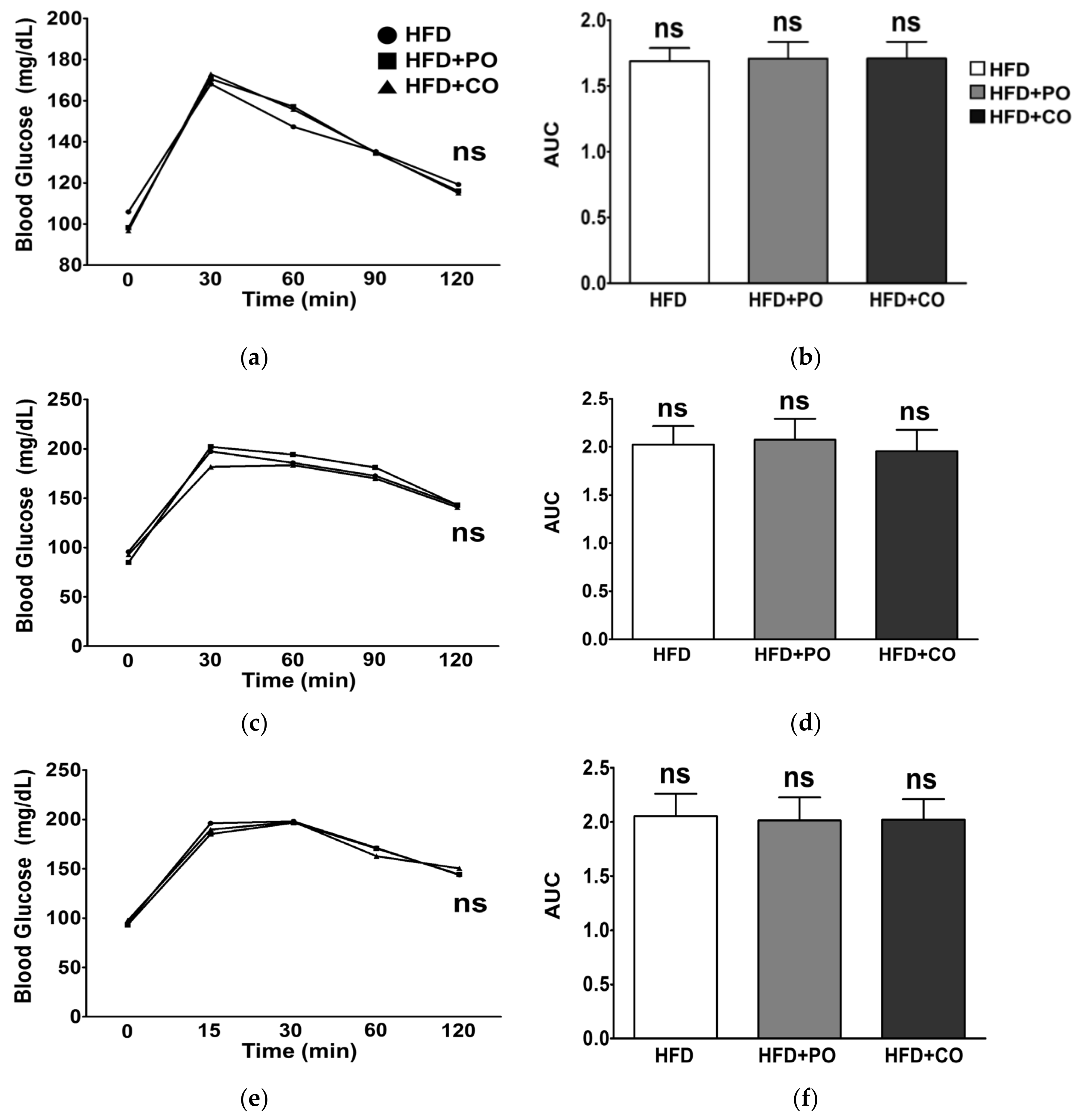
3.3. Partial Replacement of Dietary Fat with PO and CO Improved Glucose Metabolism
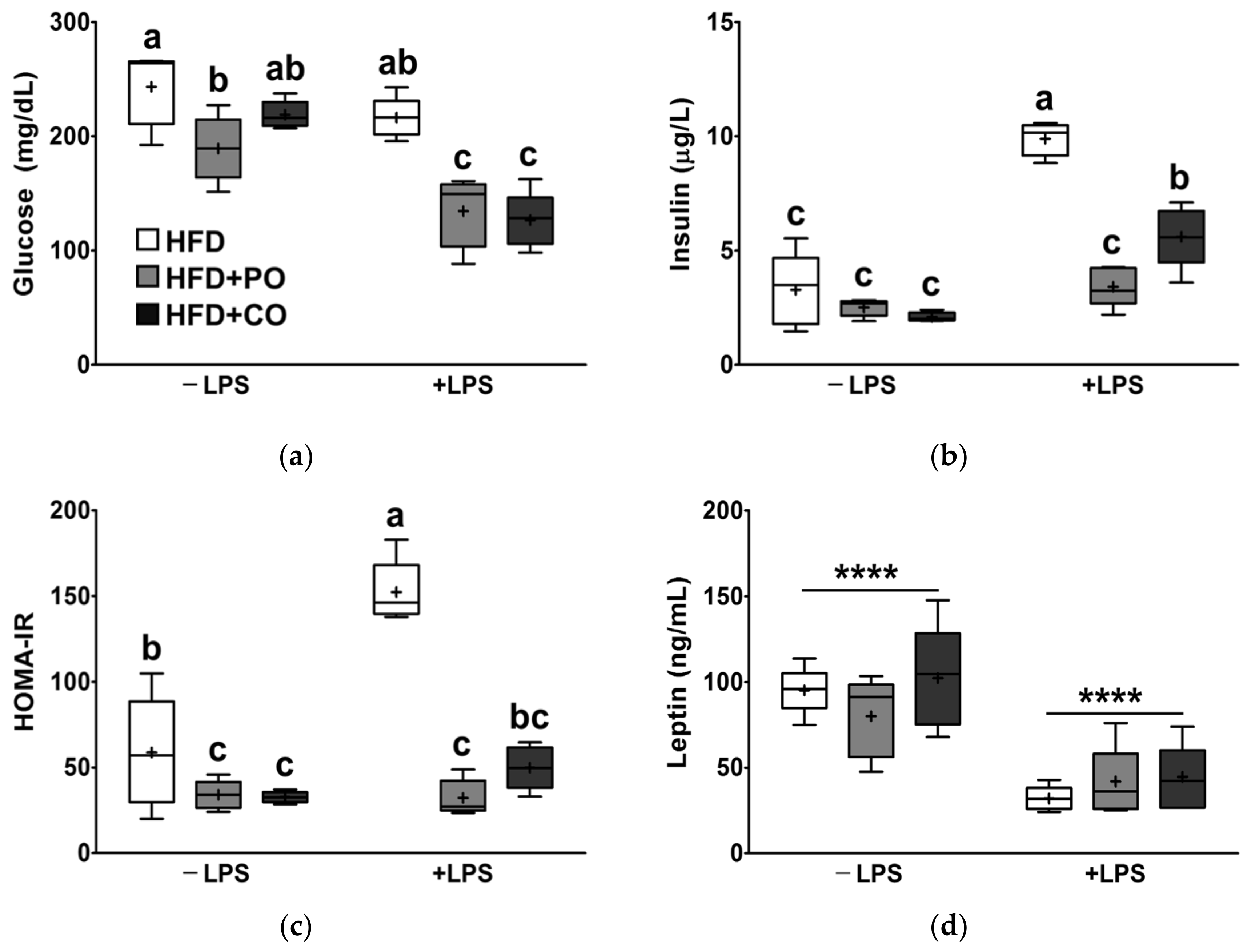
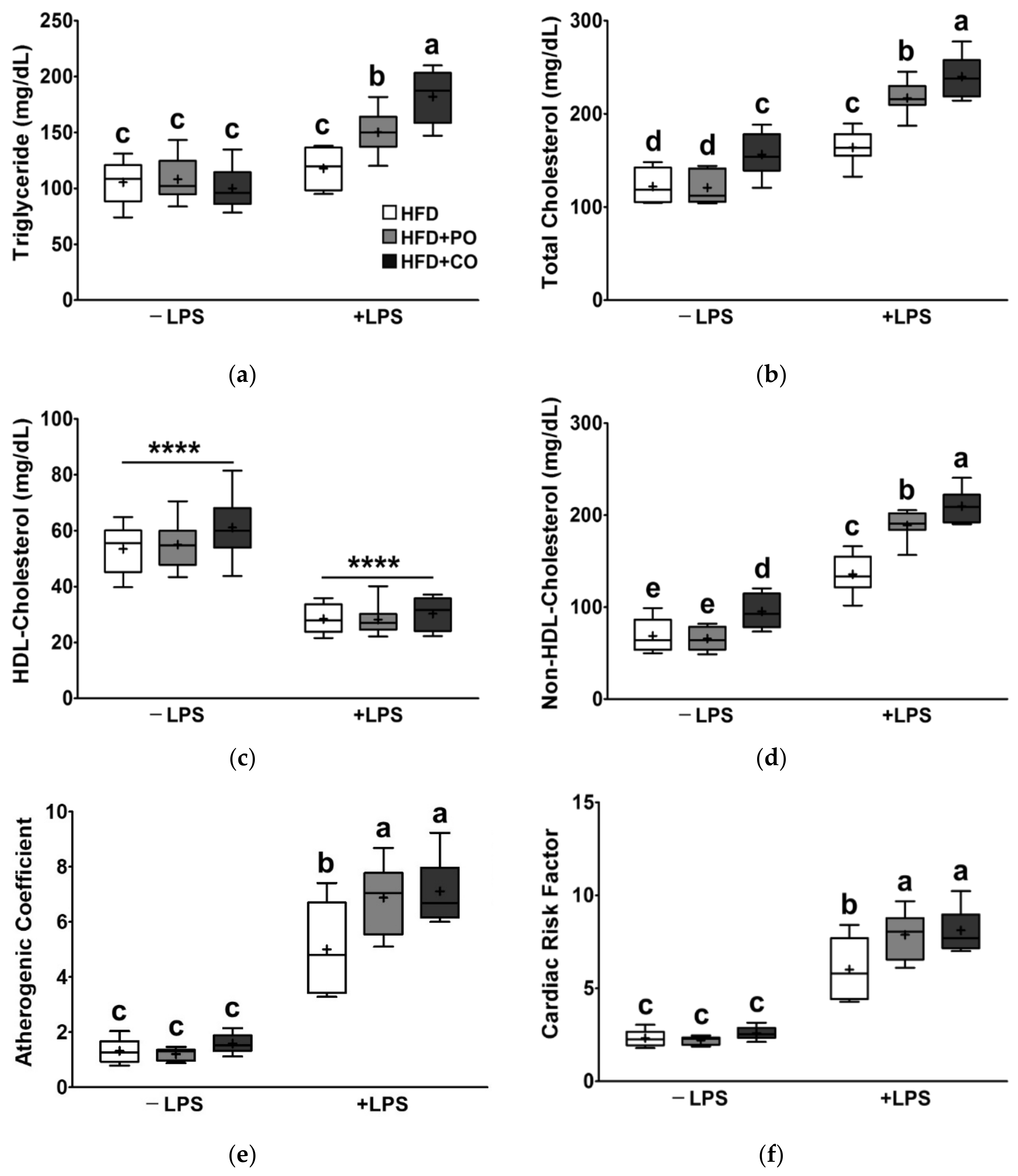
3.4. Partial Replacement of Dietary Fat with PO and CO Amplified the Changes in Serum Lipids Induced by LPS
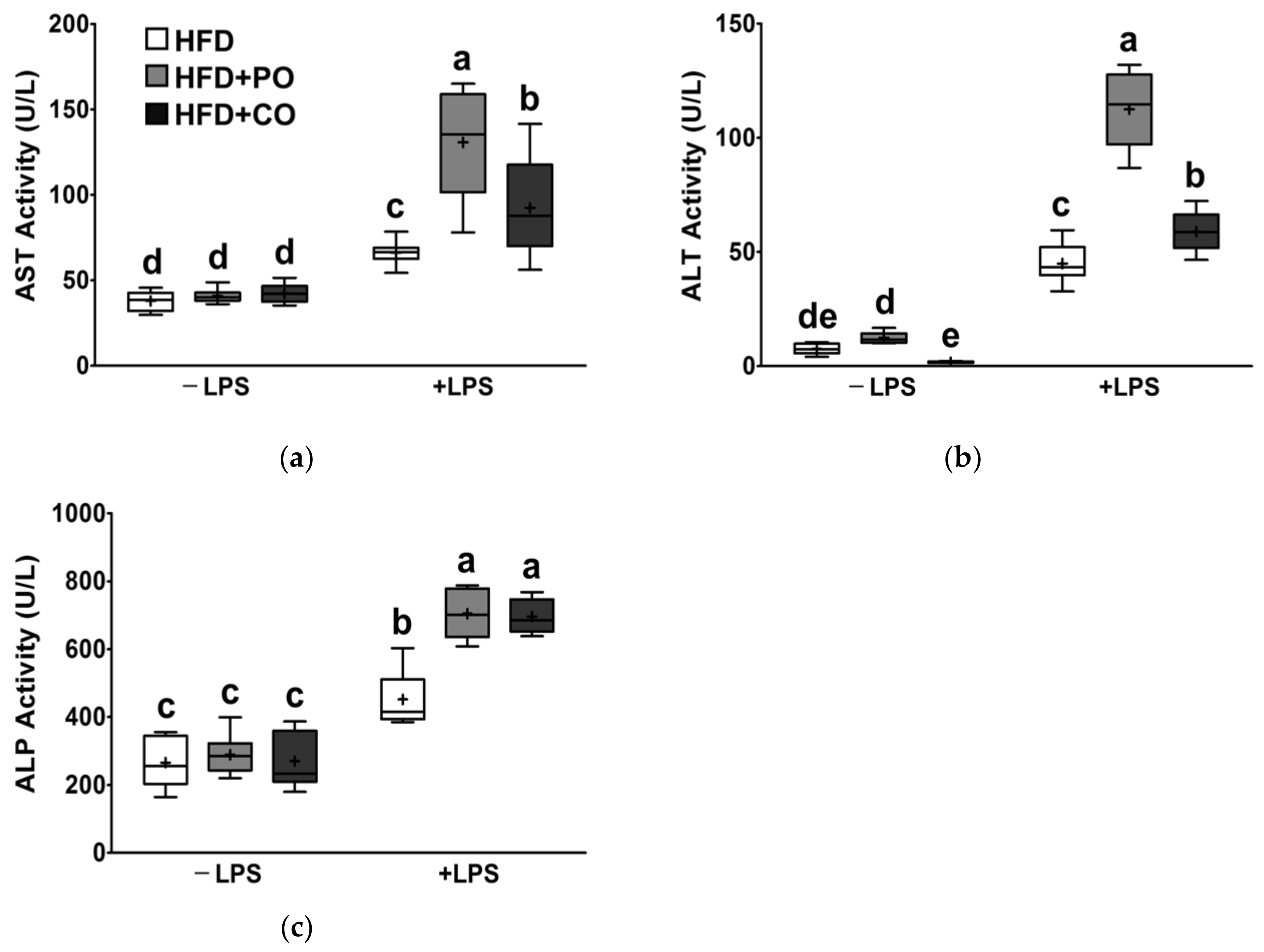
3.5. Partial Replacement of Dietary Fat with PO and CO Altered the Liver Function Parameter Changes Induced by LPS
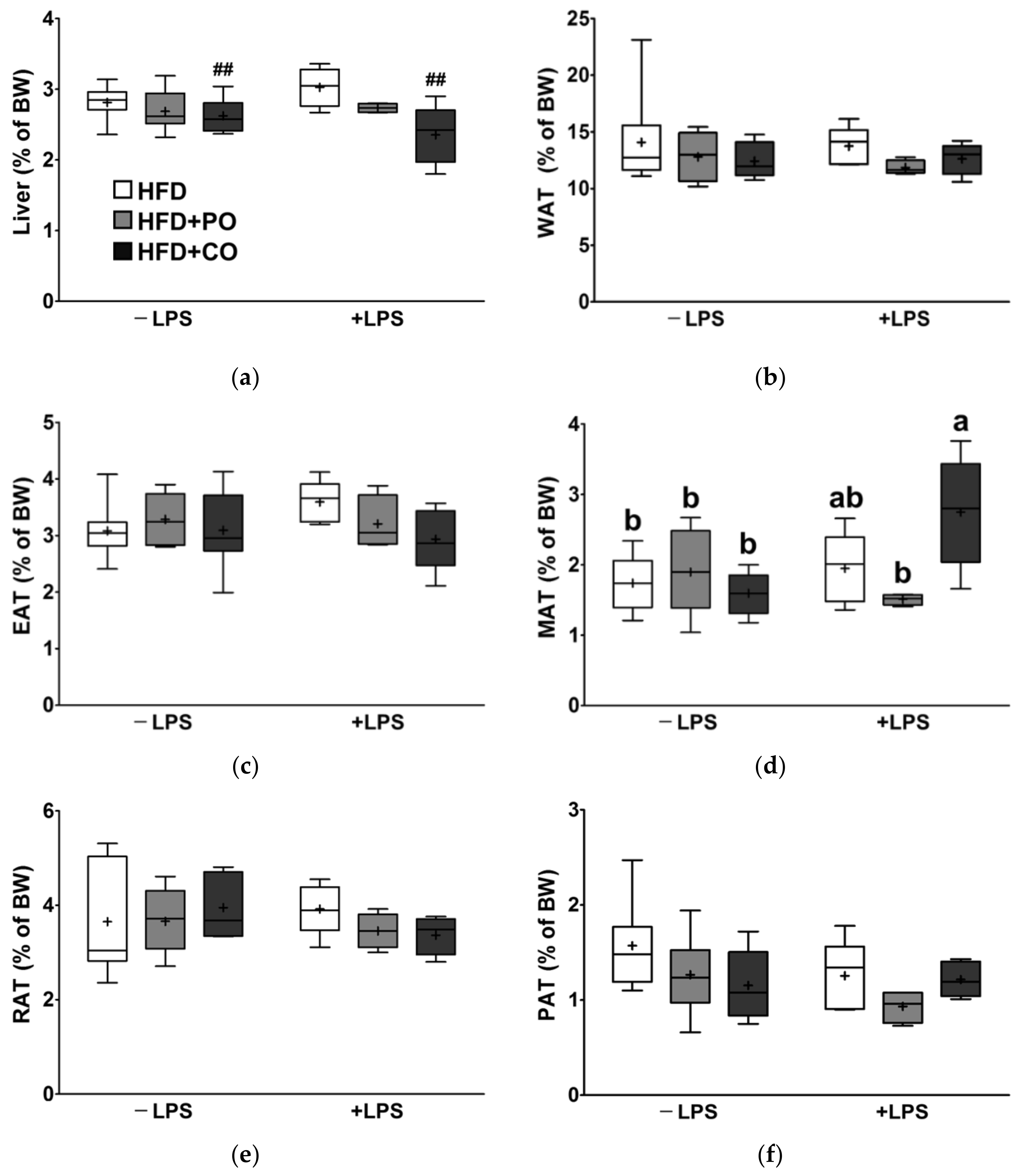
3.6. Partial Replacement of Dietary Fat with PO and CO Did Not Alter the Liver and White Adipose Tissue Weight (WAT)
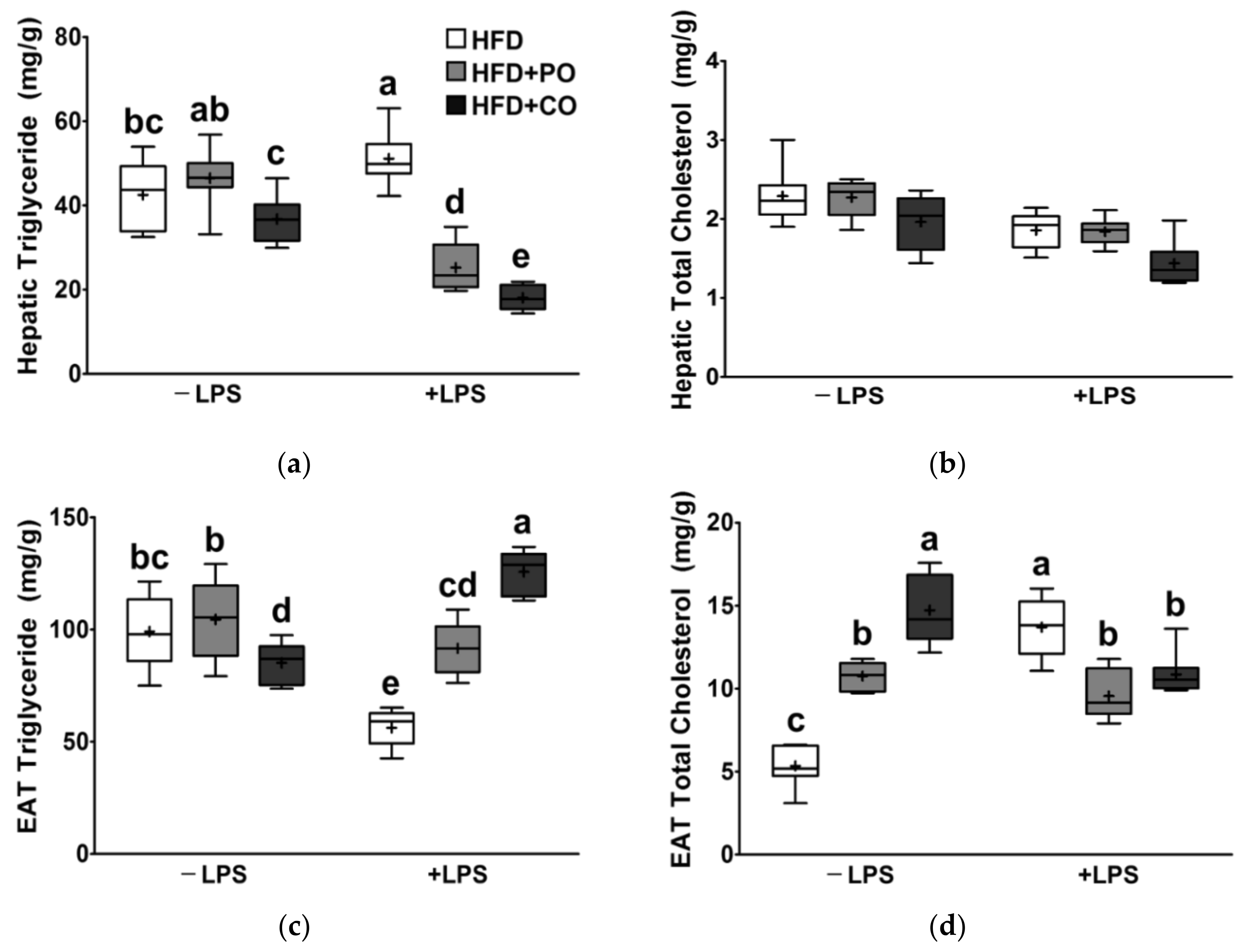
3.7. Partial Replacement of Dietary Fat with PO and CO Mitigated the LPS-Induced Lipid Changes in Liver and EAT
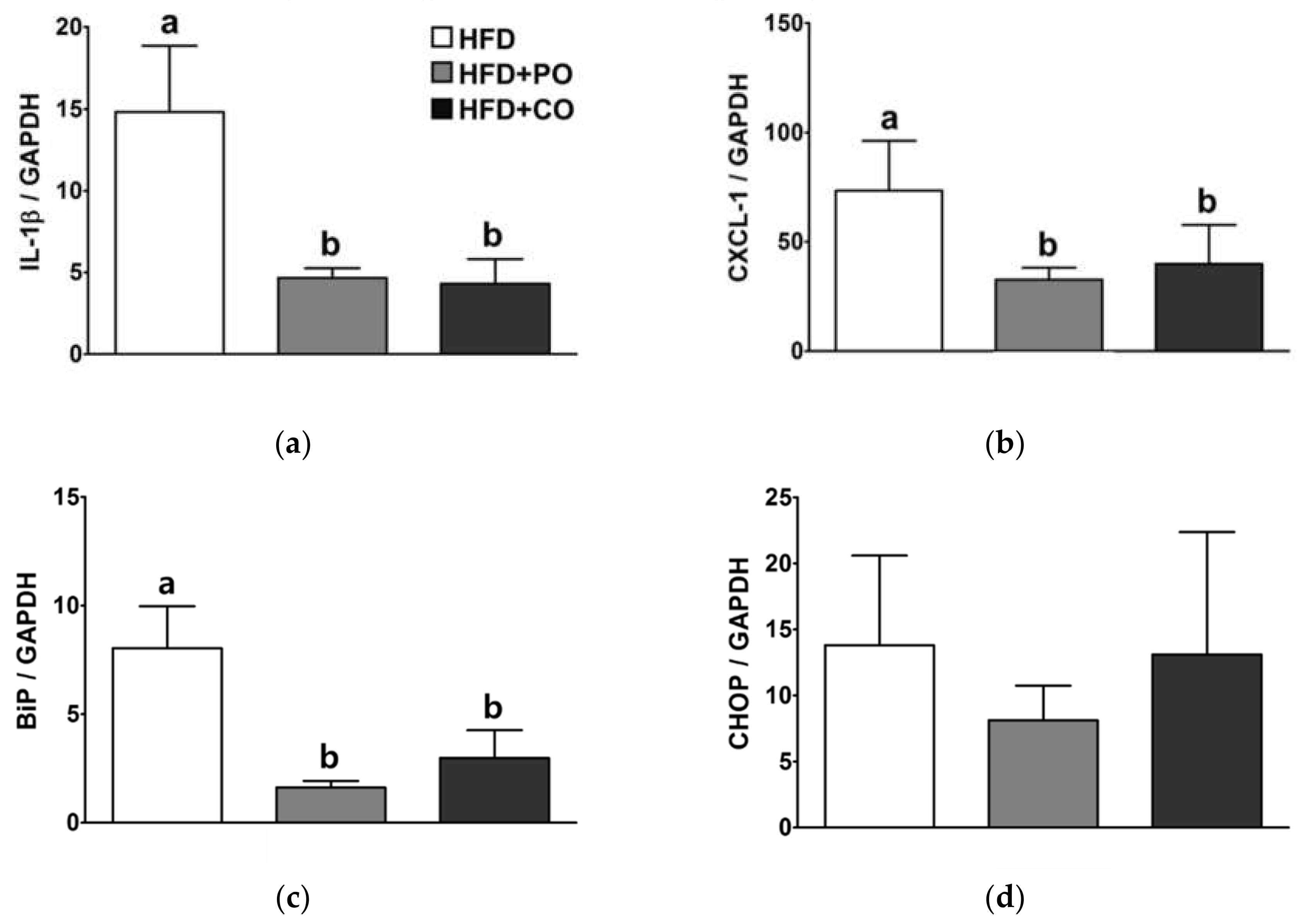
3.8. Partial Replacement of Dietary Fat with PO and CO Alleviated the mRNA Expression of Genes Related to Inflammation and ER Stress in the Liver of Rats Injected with LPS
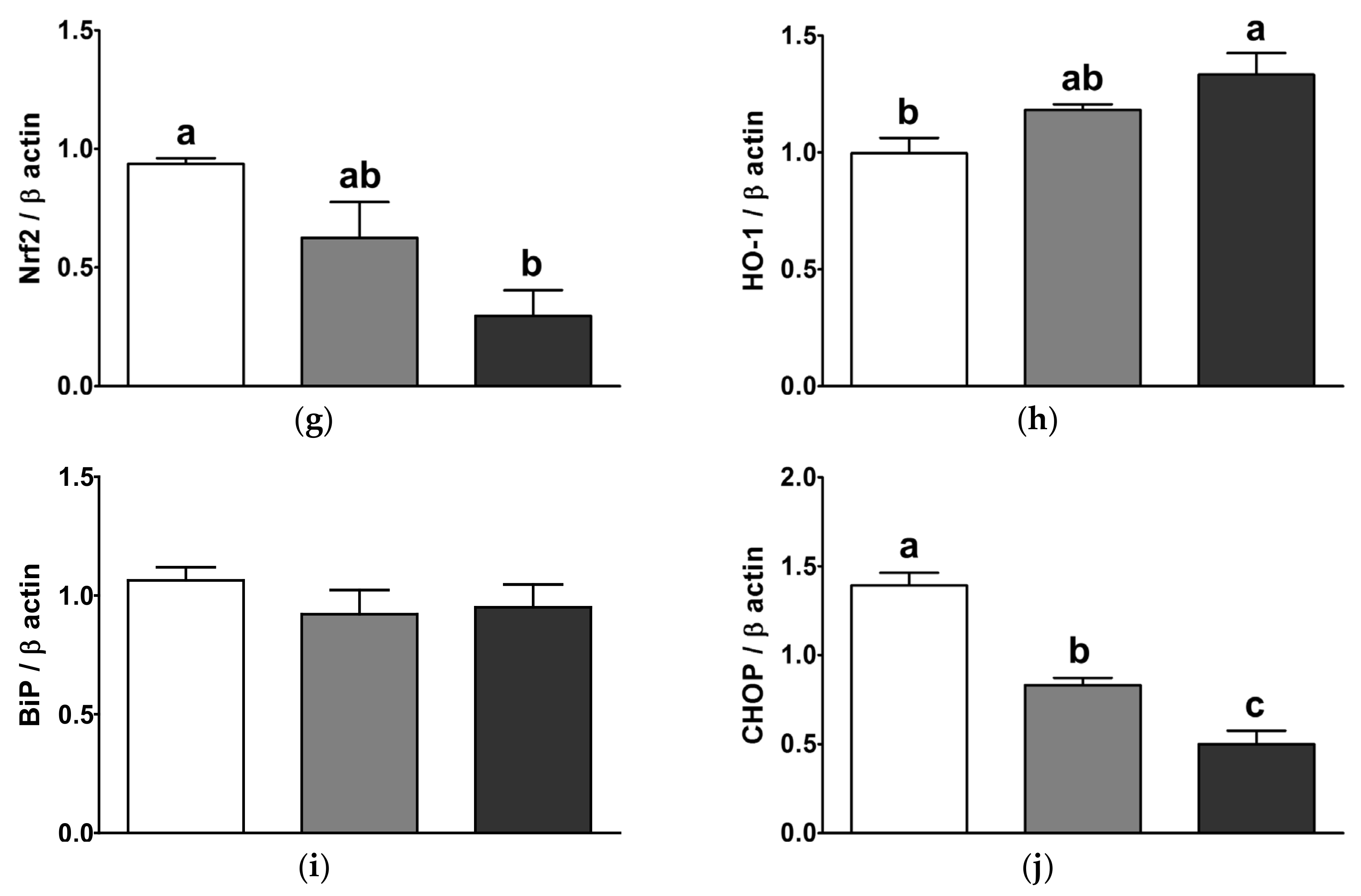
3.9. Partial Replacement of Dietary Fat with PO and CO Suppressed the Nuclear Factor-Kappa B (NF-κB) and Mitogen-Activated Protein Kinase (MAPK) Signaling Pathways and Increased the Antioxidant Enzyme Expression in the Liver of LPS-Injected Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jo, M.G.; Kim, M.W.; Jo, M.H.; Bin Abid, N.; Kim, M.O. Adiponectin homolog osmotin, a potential anti-obesity compound, suppresses abdominal fat accumulation in C57BL/6 mice on high-fat diet and in 3T3-L1 adipocytes. Int. J. Obes. 2019, 43, 2422–2433. [Google Scholar] [CrossRef]
- Tay, J.; Thompson, C.H.; Luscombe-Marsh, N.; Wycherley, T.; Noakes, M.; Buckley, J.; Wittert, G.; Yancy, W.S., Jr.; Brinkworth, G.D. Effects of an energy-restricted low-carbohydrate, high unsaturated fat/low saturated fat diet versus a high-carbohydrate, low-fat diet in type 2 diabetes: A 2-year randomized clinical trial. Diabetes Obes. Metab. 2017, 20, 858–871. [Google Scholar] [CrossRef] [PubMed]
- Wright, N.; Wilson, L.; Smith, M.; Duncan, B.; McHugh, P. The BROAD study: A randomised controlled trial using a whole food plant-based diet in the community for obesity, ischaemic heart disease or diabetes. Nutr. Diabetes 2017, 7, e256. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, R.; Pierce, N.; Koppe, S. Obesity and nonalcoholic fatty liver disease: Current perspectives. Diabetes Metab. Syndr. Obes. Targets Ther. 2018, 11, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-N.; Su, Y. Remdesivir attenuates high fat diet (HFD)-induced NAFLD by regulating hepatocyte dyslipidemia and inflammation via the suppression of STING. Biochem. Biophys. Res. Commun. 2020, 526, 381–388. [Google Scholar] [CrossRef]
- Imajo, K.; Fujita, K.; Yoneda, M.; Nozaki, Y.; Ogawa, Y.; Shinohara, Y.; Kato, S.; Mawatari, H.; Shibata, W.; Kitani, H.; et al. Hyperresponsivity to Low-Dose Endotoxin during Progression to Nonalcoholic Steatohepatitis Is Regulated by Leptin-Mediated Signaling. Cell Metab. 2012, 16, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Diao, N.; Yuan, R.; Chen, K.; Geng, S.; Li, M.; Li, L. Subclinical-Dose Endotoxin Sustains Low-Grade Inflammation and Exacerbates Steatohepatitis in High-Fat Diet–Fed Mice. J. Immunol. 2016, 196, 2300–2308. [Google Scholar] [CrossRef]
- Lawrence, C.B.; Brough, D.; Knight, E. Obese mice exhibit an altered behavioural and inflammatory response to lipopolysaccharide. Dis. Model. Mech. 2012, 5, 649–659. [Google Scholar] [CrossRef]
- Grundy, S.M. Multifactorial causation of obesity: Implications for prevention. Am. J. Clin. Nutr. 1998, 67, 563S–572S. [Google Scholar] [CrossRef]
- Lee, C.H.; Cheung, B.; Yi, G.-H.; Oh, B.; Oh, Y.H. Mobile health, physical activity, and obesity. Medicine 2018, 97, e12309. [Google Scholar] [CrossRef]
- Hendren, N.S.; De Lemos, J.A.; Ayers, C.; Das, S.R.; Rao, A.; Carter, S.; Rosenblatt, A.; Walchok, J.G.; Omar, W.; Khera, R.; et al. Association of Body Mass Index and Age With Morbidity and Mortality in Patients Hospitalized With COVID-19. Circulation 2021, 143, 135–144. [Google Scholar] [CrossRef]
- Wang, L.; Wang, H.; Zhang, B.; Popkin, B.M.; Du, S. Elevated Fat Intake Increases Body Weight and the Risk of Overweight and Obesity among Chinese Adults: 1991–2015 Trends. Nutrients 2020, 12, 3272. [Google Scholar] [CrossRef]
- FAO; WHO. Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation. FAO Food and Nutrition Paper. In Proceedings of the Joint FAO/WHO Expert Consultation, Geneva, Switzerland, 10–14 November 2008; FAO: Rome, Italy, 2010. [Google Scholar]
- Nishida, C.; Uauy, R. WHO Scientific Update on health consequences of trans fatty acids: Introduction. Eur. J. Clin. Nutr. 2009, 63, S1–S4. [Google Scholar] [CrossRef]
- Meyer, B.J.; Mann, N.J.; Lewis, J.L.; Milligan, G.C.; Sinclair, A.J.; Howe, P.R.C. Dietary intakes and food sources of omega-6 and omega-3 polyunsaturated fatty acids. Lipids 2003, 38, 391–398. [Google Scholar] [CrossRef]
- McGuire, S. U.S. Department of Agriculture and U.S. Department of Health and Human Services, Dietary Guidelines for Americans, 2010. 7th Edition, Washington, DC: U.S. Government Printing Office, January 2011. Adv. Nutr. 2011, 2, 293–294. [Google Scholar] [CrossRef]
- WHO; FAO. Diet, Nutrition and the Prevention of Chronic Diseases; WHO Technical Report Series 916; FAO: Rome, Italy, 2003; pp. 1–149. [Google Scholar]
- Zhuang, P.; Wang, W.; Wang, J.; Zhang, Y.; Jiao, J. Polyunsaturated fatty acids intake, omega-6/omega-3 ratio and mortality: Findings from two independent nationwide cohorts. Clin. Nutr. 2018, 38, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Is insulin resistance influenced by dietary linoleic acid and trans fatty acids? Free. Radic. Biol. Med. 1994, 17, 367–372. [Google Scholar] [CrossRef]
- Simopoulos, A. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Lands, B.; Bibus, D.; Stark, K. Dynamic interactions of n-3 and n-6 fatty acid nutrients. Prostaglandins Leukot. Essent. Fat. Acids 2017, 136, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Steffen, L.M.; Vessby, B.; Basu, S.; Steinberger, J.; Moran, A.; Jacobs, D.R., Jr.; Hong, C.-P.; Sinaiko, A.R. Obesity Modifies the Relations Between Serum Markers of Dairy Fats and Inflammation and Oxidative Stress Among Adolescents. Obesity 2011, 19, 2404–2410. [Google Scholar] [CrossRef] [PubMed]
- Herbert, D.; Franz, S.; Popkova, Y.; Anderegg, U.; Schiller, J.; Schwede, K.; Lorz, A.; Simon, J.C.; Saalbach, A. High-Fat Diet Exacerbates Early Psoriatic Skin Inflammation Independent of Obesity: Saturated Fatty Acids as Key Players. J. Investig. Dermatol. 2018, 138, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, Y.B.; Hein, G.; Chicco, A. Metabolic Syndrome: Effects of n-3 PUFAs on a Model of Dyslipidemia, Insulin Resistance and Adiposity. Lipids 2007, 42, 427–437. [Google Scholar] [CrossRef]
- Li, J.; Li, F.R.; Wei, D.; Jia, W.; Kang, J.X.; Stefanovic-Racic, M.; Dai, Y.; Zhao, A.Z. Endogenous ω-3 Polyunsaturated Fatty Acid Production Confers Resistance to Obesity, Dyslipidemia, and Diabetes in Mice. Mol. Endocrinol. 2014, 28, 1316–1328. [Google Scholar] [CrossRef] [PubMed]
- Mantzioris, E.; James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary substitution with an α-linolenic acid-rich vegetable oil increases eicosapentaenoic acid concentrations in tissues. Am. J. Clin. Nutr. 1994, 59, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Thota, R.; Acharya, S.H.; Garg, M.L. Curcumin and/or omega-3 polyunsaturated fatty acids supplementation reduces insulin resistance and blood lipids in individuals with high risk of type 2 diabetes: A randomised controlled trial. Lipids Health Dis. 2019, 18, 31. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-F.; Zhang, H.-M.; Li, Y.-Y.; Xia, S.; Wei, Y.; Yang, L.; Wang, D.; Ye, J.-J.; Li, H.-X.; Yuan, J.; et al. A combination of omega-3 and plant sterols regulate glucose and lipid metabolism in individuals with impaired glucose regulation: A randomized and controlled clinical trial. Lipids Health Dis. 2019, 18, 1–9. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, H.; Yuan, F.; Li, N.; Huang, Q.; He, L.; Wang, L.; Liu, Z. Perilla Oil Has Similar Protective Effects of Fish Oil on High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease and Gut Dysbiosis. BioMed Res. Int. 2016, 2016, 9462571. [Google Scholar] [CrossRef]
- Scaioli, E.; Sartini, A.; Bellanova, M.; Campieri, M.; Festi, D.; Bazzoli, F.; Belluzzi, A. Eicosapentaenoic Acid Reduces Fecal Levels of Calprotectin and Prevents Relapse in Patients With Ulcerative Colitis. Clin. Gastroenterol. Hepatol. 2018, 16, 1268–1275.e2. [Google Scholar] [CrossRef]
- Arbex, A.K.; Bizarro, V.R.; Santos, J.C.S.; Araújo, L.M.M.; de Jesus, A.L.C.; Fernandes, M.S.A.; Salles, M.M.; Rocha, D.R.T.W.; Marcadenti, A. The Impact of the Essential Fatty Acids (EFA) in Human Health. Open J. Endocr. Metab. Dis. 2015, 5, 98–104. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2007, 22, 659–661. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Prato, E.; Biandolino, F. Total lipid content and fatty acid composition of commercially important fish species from the Mediterranean, Mar Grande Sea. Food Chem. 2011, 131, 1233–1239. [Google Scholar] [CrossRef]
- Jackson, K.H.; Polreis, J.; Sanborn, L.; Chaima, D.; Harris, W.S. Analysis of breast milk fatty acid composition using dried milk samples. Int. Breastfeed. J. 2016, 11, 1–7. [Google Scholar] [CrossRef]
- Kang, N.-H.; Lee, W.K.; Yi, B.-R.; Lee, H.-R.; Park, M.-A.; Park, S.-K.; Park, H.K.; Choi, K.-C. Risk of cardiovascular disease is suppressed by dietary supplementation with protamine and chitooligosaccharide in Sprague-Dawley rats. Mol. Med. Rep. 2012, 7, 127–133. [Google Scholar] [CrossRef]
- Motamed, N.; Miresmail, S.J.H.; Rabiee, B.; Keyvani, H.; Farahani, B.; Maadi, M.; Zamani, F. Optimal cutoff points for HOMA-IR and QUICKI in the diagnosis of metabolic syndrome and non-alcoholic fatty liver disease: A population based study. J. Diabetes Complicat. 2015, 30, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-X.; Fan, J.; Ma, J.; Rao, Y.-S.; Zhang, L.; Yan, Y.-E. Selection of Suitable Reference Genes for Quantitative Real-Time PCR Normalization in Three Types of Rat Adipose Tissue. Int. J. Mol. Sci. 2016, 17, 968. [Google Scholar] [CrossRef]
- Lamping, K.G.; Nuno, D.W.; Coppey, L.J.; Holmes, A.J.; Hu, S.; Oltman, C.L.; Norris, A.W.; Yorek, M.A. Modification of high saturated fat diet with n-3 polyunsaturated fat improves glucose intolerance and vascular dysfunction. Diabetes Obes. Metab. 2012, 15, 144–152. [Google Scholar] [CrossRef]
- Imamura, F.; Micha, R.; Wu, J.; Otto, M.C.D.O.; Otite, F.O.; Abioye, A.I.; Mozaffarian, D. Effects of Saturated Fat, Polyunsaturated Fat, Monounsaturated Fat, and Carbohydrate on Glucose-Insulin Homeostasis: A Systematic Review and Meta-analysis of Randomised Controlled Feeding Trials. PLoS Med. 2016, 13, e1002087. [Google Scholar] [CrossRef]
- Eguchi, K.; Manabe, I.; Oishi-Tanaka, Y.; Ohsugi, M.; Kono, N.; Ogata, F.; Yagi, N.; Ohto, U.; Kimoto, M.; Miyake, K.; et al. Saturated Fatty Acid and TLR Signaling Link β Cell Dysfunction and Islet Inflammation. Cell Metab. 2012, 15, 518–533. [Google Scholar] [CrossRef]
- Hao, X.-Q.; Du, J.-X.; Li, Y.; Li, M.; Zhang, S. Prenatal Exposure to Lipopolysaccharide Combined with Pre- and Postnatal High-Fat Diet Result in Lowered Blood Pressure and Insulin Resistance in Offspring Rats. PLoS ONE 2014, 9, e88127. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.; Fava, F.; Tuohy, K.; Chabo, C.; et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef]
- Kim, H.-K.; Choi, S.-W.; Lee, H.-J.; Lee, J.-H.; Choi, H.-M. Suppression of fatty acid synthase by dietary polyunsaturated fatty acids is mediated by fat itself, not by peroxidative mechanism. J. Biochem. Mol. Biol. 2003, 36, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Jung, S.; Park, Y.; Lee, Y.; Jo, J.; Lee, S.; Oh, Y. Fatty acid composition and characterisation of commercial vegetable oils with chemometric approaches. Int. Food Res. J. 2020, 27, 270–279. [Google Scholar]
- TACONIC. Sprague Dawley® Rat. Available online: https://www.taconic.com/pdfs/sprague-dawley-rat.pdf (accessed on 23 July 2021).
- Lupton, J.R.; Brooks, J.; Butte, N.; Caballero, B.; Flatt, J.; Fried, S. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; National Academy Press: Washington, DC, USA, 2002; Volume 5, pp. 589–768. [Google Scholar]
- Buettner, R.; Parhofer, K.G.; Woenckhaus, M.; Wrede, C.E.; Kunz-Schughart, L.; Schölmerich, J.; Bollheimer, L.C. Defining high-fat-diet rat models: Metabolic and molecular effects of different fat types. J. Mol. Endocrinol. 2006, 36, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Liput, K.; Lepczyński, A.; Nawrocka, A.; Poławska, E.; Ogłuszka, M.; Jończy, A.; Grzybek, W.; Liput, M.; Szostak, A.; Urbański, P.; et al. Effects of Three-Month Administration of High-Saturated Fat Diet and High-Polyunsaturated Fat Diets with Different Linoleic Acid (LA, C18:2n–6) to α-Linolenic Acid (ALA, C18:3n–3) Ratio on the Mouse Liver Proteome. Nutrients 2021, 13, 1678. [Google Scholar] [CrossRef] [PubMed]
- Rockett, B.D.; Salameh, M.; Carraway, K.; Morrison, K.; Shaikh, S.R. n-3 PUFA improves fatty acid composition, prevents palmitate-induced apoptosis, and differentially modifies B cell cytokine secretion in vitro and ex vivo. J. Lipid Res. 2010, 51, 1284–1297. [Google Scholar] [CrossRef]
- Nagy, C.; Einwallner, E. Study of In Vivo Glucose Metabolism in High-fat Diet-fed Mice Using Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT). J. Vis. Exp. 2018, e56672. [Google Scholar] [CrossRef]
- Ibrahim, A.; Natarajan, S. Substituting dietary linoleic acid with α-linolenic acid improves insulin sensitivity in sucrose fed rats. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2005, 1733, 67–75. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, S.; Li, W.; Ma, L.; Ding, M.; Li, R.; Liu, Y. High-fat diet from perilla oil induces insulin resistance despite lower serum lipids and increases hepatic fatty acid oxidation in rats. Lipids Health Dis. 2014, 13, 15. [Google Scholar] [CrossRef]
- Giknis, M.L.A.; Clifford, C. Clinical Laboratory Parameters for Crl: WI (Han); Charles River Laboratories: Wilmington, MA, USA, 2008; pp. 1–14. Available online: https://www.criver.com/sites/default/files/resources/rm_rm_r_Wistar_Han_clin_lab_parameters_08.pdf (accessed on 24 September 2021).
- Raetzsch, C.F.; Brooks, N.L.; Alderman, J.M.; Moore, K.S.; Hosick, P.A.; Klebanov, S.; Akira, S.; Bear, J.E.; Baldwin, A.S.; Mackman, N.; et al. Lipopolysaccharide inhibition of glucose production through the Toll-like receptor-4, myeloid differentiation factor 88, and nuclear factor κb pathway. Hepatology 2009, 50, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, T.; Chuang, J.-C.; Sakata, I.; Ramadori, G.; Coppari, R. Leptin therapy improves insulin-deficient type 1 diabetes by CNS-dependent mechanisms in mice. Proc. Natl. Acad. Sci. USA 2010, 107, 17391–17396. [Google Scholar] [CrossRef]
- Zhao, S.; Zhu, Y.; Schultz, R.D.; Li, N.; He, Z.; Zhang, Z.; Caron, A.; Zhu, Q.; Sun, K.; Xiong, W.; et al. Partial Leptin Reduction as an Insulin Sensitization and Weight Loss Strategy. Cell Metab. 2019, 30, 706–719.e6. [Google Scholar] [CrossRef]
- Al-Lahham, S.; Jaradat, N.; Al-Qub, M.; Hamayel, A.; Assaassa, A.; Hammad, F.; Mosa, A.; Nafaa, F.; Barqawi, A.; Abdeen, Z. Lipopolysaccharide influence on leptin hormone and tumor necrosis factor-alpha release from human adipose tissue. Eur. J. Inflamm. 2018, 16, 2058739218774975. [Google Scholar] [CrossRef]
- Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef]
- Yu, S.; Wen, Y.; Li, J.; Zhang, H.; Liu, Y. Prenatal Lipopolysaccharide Exposure Promotes Dyslipidemia in the Male Offspring Rats. Front. Physiol. 2018, 9, 542. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-P.; Chen, Y.-H.; Li, H. Association between the levels of polyunsaturated fatty acids and blood lipids in healthy individuals. Exp. Ther. Med. 2012, 4, 1107–1111. [Google Scholar] [CrossRef][Green Version]
- Grenon, S.M.; Owens, C.D.; Nosova, E.V.; Hughes-Fulford, M.; Alley, H.F.; Chong, K.; Perez, S.; Yen, P.K.; Boscardin, J.; Hellmann, J.; et al. Short-Term, High-Dose Fish Oil Supplementation Increases the Production of Omega-3 Fatty Acid–Derived Mediators in Patients With Peripheral Artery Disease (the OMEGA-PAD I Trial). J. Am. Heart Assoc. 2015, 4, e002034. [Google Scholar] [CrossRef]
- Miller, E.R.; Juraschek, S.; Anderson, C.A.; Guallar, E.; Henoch-Ryugo, K.; Charleston, J.; Turban, S.; Bennett, M.R.; Appel, L.J. The Effects of n-3 Long-Chain Polyunsaturated Fatty Acid Supplementation on Biomarkers of Kidney Injury in Adults With Diabetes: Results of the GO-FISH trial. Diabetes Care 2012, 36, 1462–1469. [Google Scholar] [CrossRef]
- Jenkinson, A.M.; Collins, A.R.; Duthie, S.; Wahle, K.W.J.; Duthie, G.G. The effect of increased intakes of polyunsaturated fatty acids and vitamin E on DNA damage in human lymphocytes. FASEB J. 1999, 13, 2138–2142. [Google Scholar] [CrossRef]
- Emam, H.; Ahmed, E.; Abdel-Daim, M. Antioxidant capacity of omega-3-fatty acids and vitamin E against imidacloprid-induced hepatotoxicity in Japanese quails. Environ. Sci. Pollut. Res. 2018, 25, 11694–11702. [Google Scholar] [CrossRef]
- Johansen, O.; Seljeflot, I.; Høstmark, A.T.; Arnesen, H. The Effect of Supplementation With Omega-3 Fatty Acids on Soluble Markers of Endothelial Function in Patients With Coronary Heart Disease. Arter. Thromb. Vasc. Biol. 1999, 19, 1681–1686. [Google Scholar] [CrossRef]
- Wander, R.C.; Hall, J.A.; Gradin, J.L.; Du, S.-H.; Jewell, D.E. The ratio of dietary (n-6) to (n-3) fatty acids influences immune system function, eicosanoid metabolism, lipid peroxidation and vitamin E status in aged dogs. J. Nutr. 1997, 127, 1198–1205. [Google Scholar] [CrossRef]
- Chautan, M.; Calaf, R.; Léonardi, J.; Charbonnier, M.; Andre, M.; Portugal, H.; Pauli, A.; Lafont, H.; Nalbone, G. Inverse modifications of heart and liver alpha-tocopherol status by various dietary n-6/n-3 polyunsaturated fatty acid ratios. J. Lipid Res. 1990, 31, 2201–2208. [Google Scholar] [CrossRef]
- Rosqvist, F.; Iggman, D.; Kullberg, J.; Cedernaes, J.; Johansson, H.-E.; Larsson, A.; Johansson, L.; Ahlström, H.; Arner, P.; Dahlman, I.; et al. Overfeeding Polyunsaturated and Saturated Fat Causes Distinct Effects on Liver and Visceral Fat Accumulation in Humans. Diabetes 2014, 63, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Greaves, P. Chapter 9—Liver and Pancreas. In Histopathology of Preclinical Toxicity Studies, 4th ed.; Greaves, P., Ed.; Academic Press: Boston, MA, USA, 2012; pp. 433–535. [Google Scholar]
- Fukunishi, S.; Sujishi, T.; Takeshita, A.; Ohama, H.; Tsuchimoto, Y.; Asai, A.; Tsuda, Y.; Higuchi, K. Lipopolysaccharides accelerate hepatic steatosis in the development of nonalcoholic fatty liver disease in Zucker rats. J. Clin. Biochem. Nutr. 2014, 54, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Bjermo, H.; Iggman, D.; Kullberg, J.; Dahlman, I.; Johansson, L.; Persson, L.; Berglund, J.; Pulkki, K.; Basu, S.; Uusitupa, M.; et al. Effects of n-6 PUFAs compared with SFAs on liver fat, lipoproteins, and inflammation in abdominal obesity: A randomized controlled trial. Am. J. Clin. Nutr. 2012, 95, 1003–1012. [Google Scholar] [CrossRef]
- Kim, S.R.; Je, J.; Jeong, K.; Kim, S.J.; Lee, K.-Y.; Choi, S.-G.; Kim, H.; Park, S.W. Perilla Oil Decreases Aortic and Hepatic Lipid Accumulation by Modulating Lipogenesis and Lipolysis in High-Fat Diet-Fed Mice. J. Med. Food 2019, 22, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Duwaerts, C.C.; Maher, J.J. Macronutrients and the Adipose-Liver Axis in Obesity and Fatty Liver. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Balvers, M.G.; Verhoeckx, K.C.; Plastina, P.; Wortelboer, H.M.; Meijerink, J.; Witkamp, R.F. Docosahexaenoic acid and eicosapentaenoic acid are converted by 3T3-L1 adipocytes to N-acyl ethanolamines with anti-inflammatory properties. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2010, 1801, 1107–1114. [Google Scholar] [CrossRef]
- Hao, W.; Wong, O.Y.; Liu, X.; Lee, P.; Chen, Y.; Wong, K.K. ω-3 fatty acids suppress inflammatory cytokine production by macrophages and hepatocytes. J. Pediatr. Surg. 2010, 45, 2412–2418. [Google Scholar] [CrossRef]
- Ibrahim, A.; Mbodji, K.; Hassan, A.; Aziz, M.; Boukhettala, N.; Coëffier, M.; Savoye, G.; Déchelotte, P.; Marion-Letellier, R. Anti-inflammatory and anti-angiogenic effect of long chain n-3 polyunsaturated fatty acids in intestinal microvascular endothelium. Clin. Nutr. 2011, 30, 678–687. [Google Scholar] [CrossRef]
- Mullen, A.; Loscher, C.E.; Roche, H.M. Anti-inflammatory effects of EPA and DHA are dependent upon time and dose-response elements associated with LPS stimulation in THP-1-derived macrophages. J. Nutr. Biochem. 2010, 21, 444–450. [Google Scholar] [CrossRef]
- Ajuwon, K.M.; Spurlock, M.E. Palmitate Activates the NF-κB Transcription Factor and Induces IL-6 and TNFα Expression in 3T3-L1 Adipocytes. J. Nutr. 2005, 135, 1841–1846. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.F.; Chau, Y.P.; Ni Kung, H.; Lu, K.S. The lipopolysaccharide-induced pro-inflammatory response in RAW264.7 cells is attenuated by an unsaturated fatty acid–bovine serum albumin complex and enhanced by a saturated fatty acid–bovine serum albumin complex. Inflamm. Res. 2011, 61, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.A.; Walker, C.L.; Pavlina, T.M.; Xu, Z.; Zaloga, G.P.; Siddiqui, R.A. Long-chain saturated fatty acids induce pro-inflammatory responses and impact endothelial cell growth. Clin. Nutr. 2010, 29, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Fu, Q.; Ma, S. Perillaldehyde attenuates cerebral ischemia–reperfusion injury-triggered overexpression of inflammatory cytokines via modulating Akt/JNK pathway in the rat brain cortex. Biochem. Biophys. Res. Commun. 2014, 454, 65–70. [Google Scholar] [CrossRef]
- Inui, T.; Watanabe, M.; Nakamoto, K.; Sada, M.; Hirata, A.; Nakamura, M.; Honda, K.; Ogawa, Y.; Takata, S.; Yokoyama, T.; et al. Bronchial epithelial cells produce CXCL1 in response to LPS and TNFα: A potential role in the pathogenesis of COPD. Exp. Lung Res. 2018, 44, 323–331. [Google Scholar] [CrossRef]
- Lai, J.-L.; Liu, Y.-H.; Liu, C.; Zhou, Q.-G.; Liu, R.-N.; Zhu, X.-F.; Zhou, Q.-G.; Chen, Y.-Y.; Guo, A.-Z.; Hu, C.-M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-kB and MAPK Signaling Pathways. Inflammation 2016, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Sanjo, H.; Takeda, K.; Ninomiya-Tsuji, J.; Yamamoto, M.; Kawai, T.; Matsumoto, K.; Takeuchi, O.; Akira, S. Essential function for the kinase TAK1 in innate and adaptive immune responses. Nat. Immunol. 2005, 6, 1087–1095. [Google Scholar] [CrossRef]
- Chattree, V.; Khanna, N.; Rao, D. Alterations in T cell signal transduction by M. leprae antigens is associated with downregulation of second messengers PKC, calcium, calcineurin, MAPK and various transcription factors in leprosy patients. Mol. Immunol. 2007, 44, 2066–2077. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, J.; Li, J.-S.; Chen, H.; Huang, K.; Zheng, L. Accumulation of endoplasmic reticulum stress and lipogenesis in the liver through generational effects of high fat diets. J. Hepatol. 2012, 56, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.-R.; Ren, Y.-L.; Liu, W.-X.; Hu, Y.-J.; Zheng, J.-S.; Xu, Y.; Wang, G. Resveratrol prevents hepatic steatosis and endoplasmic reticulum stress and regulates the expression of genes involved in lipid metabolism, insulin resistance, and inflammation in rats. Nutr. Res. 2015, 35, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.J.; Kim, J.E.; Choi, H.J.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Seo, S.; Yang, S.Y.; Rapeanu, G.; Lee, H.S.; et al. α-Linolenic Acid-Enriched Cold-Pressed Perilla Oil Suppress High-Fat Diet-Induced Hepatic Steatosis through Amelioration of the ER Stress-Mediated Autophagy. Molecules 2020, 25, 2662. [Google Scholar] [CrossRef] [PubMed]
- Vasileva, L.V.; Savova, M.S.; Amirova, K.M.; Dinkova-Kostova, A.T.; Georgiev, M.I. Obesity and NRF2-mediated cytoprotection: Where is the missing link? Pharmacol. Res. 2020, 156, 104760. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg) | Amount |
|---|---|
| Casein | 220 |
| L-cysteine | 3.4 |
| Sucrose | 100 |
| Corn starch | 160 |
| Dextrose | 155 |
| Cellulose | 58 |
| Mineral mix 1 | 43 |
| Vitamin mix 2 | 19 |
| Choline bitartrate | 2.8 |
| Lard | 55 |
| tert-Butylhydroquinone | 0.034 |
| Energy (kcal/g) | 4.8 |
| Fat (%) | 23.9 |
| Fat (kcal%) | 45 |
| Ingredient (g/kg) | HFD | HFD + PO | HFD + CO |
|---|---|---|---|
| Butter 1 | 184 | 164 | 104 |
| Perilla oil | 0 | 20 | 0 |
| Corn oil | 0 | 0 | 80 |
| Gene | Primer Sequence (5′→3′) | |
|---|---|---|
| CXCL-1 | Forward Reverse | CCACACTCAAGAATGGTCGC GTTGTCAGAAGCCAGCGTTC |
| IL-1β | Forward Reverse | AAAAATGCCTCGTGCTGTCT TCGTTGCTTGTCTCTCCTTG |
| BiP | Forward Reverse | TGCCCACCAAGAAGTCTCAGA TCAAATGTACCCAGAAGGTGATTG |
| CHOP | Forward Reverse | GGAGAAGGAGCAGGAGAATG GAGACAGACAGGAGGTGATG |
| GAPDH | Forward Reverse | CTGTGTCTTTCCGCTGTTTTC TGTGCTGTGCTTATGGTCTCA |
| Antibody | Company | Catalog Number | Dilution |
|---|---|---|---|
| p-IκBα | Cell Signaling | 2859 | 1:1000 |
| t-IκBα | Cell Signaling | 4814 | 1:1000 |
| p-NFκB | Cell Signaling | 3033 | 1:1000 |
| t-NFκB | Cell Signaling | 8242 | 1:1000 |
| p-JNK | Cell Signaling | 9251 | 1:500 |
| t-JNK | Cell Signaling | 9252 | 1:1000 |
| p-ERK | Cell Signaling | 4370 | 1:3000 |
| t-ERK | Cell Signaling | 4695 | 1:1000 |
| p-p38 | Cell Signaling | 4511 | 1:1000 |
| t-p38 | Cell Signaling | 8690 | 1:1000 |
| Nrf2 | Cell Signaling | 12721S | 1:1000 |
| HO-1 | Cell Signaling | 5853S | 1:1000 |
| BiP | Cell Signaling | 3183 | 1:1000 |
| CHOP | Cell Signaling | 2895 | 1:1000 |
| β-actin | Santa Cruz | sc-47778 | 1:2000 |
| Anti-rabbit IgG | Cell Signaling | 7074 | 1:3000 |
| Anti-mouse IgG | Cell Signaling | 7076 | 1:1000 |
| Parameter | LPS Main Effect | Diet Main Effect | LPS X Diet Interaction |
|---|---|---|---|
| Serum glucose, insulin, and leptin levels | |||
| Glucose | **** p < 0.0001 | **** p < 0.0001 | * p < 0.05 |
| Insulin | **** p < 0.0001 | ** p < 0.01 | ** p < 0.01 |
| HOMA-IR | **** p < 0.0001 | **** p < 0.0001 | **** p < 0.0001 |
| Serum leptin | **** p < 0.0001 | ns p = 0.328 | ns p = 0.332 |
| Serum lipid profile | |||
| Triglyceride | **** p < 0.0001 | *** p < 0.001 | **** p < 0.0001 |
| Total cholesterol | **** p < 0.0001 | **** p < 0.0001 | *** p < 0.001 |
| HDL-cholesterol | **** p < 0.0001 | ns p = 0.191 | ns p = 0.573 |
| Non-HDL-cholesterol | **** p < 0.0001 | **** p < 0.0001 | **** p < 0.0001 |
| Atherogenic coefficient | **** p < 0.0001 | ** p < 0.01 | * p < 0.05 |
| Cardiac risk factor | **** p < 0.0001 | ** p < 0.01 | * p < 0.05 |
| Hepatic function parameters | |||
| Serum AST | **** p < 0.0001 | **** p < 0.0001 | **** p < 0.0001 |
| Serum ALT | **** p < 0.0001 | **** p < 0.0001 | **** p < 0.0001 |
| Serum ALP | **** p < 0.0001 | **** p < 0.0001 | **** p < 0.0001 |
| Relative tissue weights | |||
| Liver | ns p = 0.972 | ** p < 0.01 | ns p = 0.134 |
| WAT | ns p = 0.610 | ns p = 0.384 | ns p = 0.846 |
| EAT | ns p = 0.718 | ns p = 0.420 | ns p = 0.412 |
| MAT | ns p = 0.102 | ns p = 0.074 | ** p < 0.01 |
| RAT | ns p = 0.484 | ns p = 0.861 | ns p = 0.523 |
| PAT | ns p = 0.319 | ns p = 0.465 | ns p = 0.541 |
| Lipid contents in liver and epididymal adipose tissue | |||
| Hepatic triglyceride | **** p < 0.0001 | **** p < 0.0001 | **** p < 0.0001 |
| Hepatic total cholesterol | **** p < 0.0001 | *** p < 0.001 | ns p = 0.846 |
| EAT triglyceride | ns p = 0.160 | **** p < 0.0001 | **** p < 0.0001 |
| EAT total cholesterol | * p < 0.05 | **** p < 0.0001 | **** p < 0.0001 |
| Fatty Acid (%) | HFD | HFD + PO | HFD + CO |
|---|---|---|---|
| Palmitic acid (C16:0) | 59.64 | 35.27 | 25.46 |
| Stearic acid (C18:0) | 10.32 | 12.44 | 9.36 |
| Oleic acid (C18:1n-9c) | 23.77 | 33.21 | 33.53 |
| Elaidic acid (C18:1n-9t) | 1.45 | 2.8 | 1.44 |
| Linoleic acid (C18:2n-6c) | 4.82 | 7.58 | 28.08 |
| Linolelaidic acid (C18:2n-6t) | <LLOQ | 0.27 | 0.33 |
| α-linolenic acid (C18:3n-3) | <LLOQ | 8.12 | 0.38 |
| Eicosanoic acid (C20:0) | <LLOQ | 0.2 | <LLOQ |
| Eicosenoic acid (C20:1n-9) | <LLOQ | <LLOQ | 1.36 |
| Eicosatrienoic acid (C20:3n-3) | <LLOQ | 0.12 | 0.08 |
| SFA | 69.96 | 47.91 | 34.81 |
| MUFA | 25.23 | 36.01 | 36.33 |
| trans | 1.45 | 3.06 | 1.77 |
| PUFA | 4.82 | 16.09 | 28.86 |
| n-6 | 4.82 | 7.85 | 28.40 |
| n-3 | <LLOQ | 8.24 | 0.46 |
| n-6/n-3 | NC | 0.95 | 62.24 |
| Fatty Acid (%) | HFD | HFD + PO | HFD + CO |
|---|---|---|---|
| Myristic acid (C14:0) | 0.64 ± 0.32 ns | 0.70 ± 0.10 | 0.58 ± 0.11 |
| Palmitic acid (C16:0) | 26.40 ± 1.70 ns | 26.60 ± 0.20 | 25.73 ± 1.17 |
| Palmitoleic acid (C16:1n-7c) | 1.34 ± 0.29 a | 0.82 ± 0.22 ab | 0.42 ± 0.16 b |
| Palmitoleic acid (C16:1n-7t) | 0.39 ± 0.12 a | 0.19 ± 0.02 b | 0.17 ± 0.05 b |
| Stearic acid (C18:0) | 14.65 ± 0.35 b | 18.57 ± 0.31 a | 18.17 ± 1.67 a |
| Oleic acid (C18:1n-9c) | 19.50 ± 1.90 a | 12.93 ± 1.33 b | 10.28 ± 0.69 b |
| Oleic acid (C18:1n-9t) | 0.38 ± 0.26 ns | 0.47 ± 0.12 | 0.40 ± 0.22 |
| Linoleic acid (C18:2n-6c) | 11.47 ± 1.53 a | 7.23 ± 0.39 b | 9.91 ± 0.85 a |
| Linoleic acid (C18:2n-6t) | 0.07 ± 0.01 b | 0.06 ± 0.01 b | 0.10 ± 0.02 a |
| α-linolenic acid (C18:3n-3) | 0.25 ± 0.02 b | 0.37 ± 0.03 a | 0.04 ± 0.02 c |
| γ-linolenic acid (C18:3n-6) | 0.10 ± 0.07 ns | 0.03 ± 0.02 | 0.04 ± 0.02 |
| Eicosenoic acid (C20:1n-9) | 0.23 ± 0.06 ns | 0.11 ± 0.02 | 0.13 ± 0.08 |
| Eicosadienoic acid (C20:2n-6) | 0.31 ± 0.10 ns | 0.26 ± 0.10 | 0.35 ± 0.07 |
| Dihomo-y-linolenic acid (C20:3n-6) | 1.15 ± 0.57 ns | 0.91 ± 0.12 | 0.40± 0.10 |
| Arachidonic acid (C20:4n-6) | 16.70 ± 1.90 c | 20.63 ± 1.45 b | 26.33 ± 0.85 a |
| Eicosapentaenoic acid (C20:5n-3) | 0.34 ± 0.07 b | 1.89 ± 0.23 a | 0.23 ± 0.07 b |
| Docosatetraenoic acid (C22:4n-6) | 0.96 ± 0.09 b | 0.44 ± 0.06 c | 2.50 ± 0.22 a |
| Docosapentaenoic acid (C22:5n-3) | 0.84 ± 0.10 b | 2.98 ± 0.26 a | 0.65 ± 0.14 b |
| Docosahexaenoic acid (C22:6n-3) | 3.65 ± 0.88 a | 4.08 ± 0.83 a | 1.60 ± 0.26 b |
| Docosapentaenoic acid (C22:6n-6) | 0.20 ± 0.01 b | 0.11 ± 0.05 b | 1.34 ± 0.52 a |
| Lignoceric acid (C24:0) | 0.16 ± 0.11 b | 0.33 ± 0.01 a | 0.26 ± 0.04 ab |
| Nervonic acid (C24:1n-9) | 0.12 ± 0.08 ns | 0.14 ± 0.02 | 0.15 ± 0.05 |
| SFA | 41.84 ± 1.77 b | 46.19 ± 0.18 a | 44.74 ± 0.49 a |
| MUFA | 21.95 ± 2.35 a | 14.66 ± 1.55 b | 11.55 ± 0.73 b |
| TFA | 0.83 ± 0.14 ns | 0.72 ± 0.12 | 0.67 ± 0.16 |
| PUFA | 36.01 ± 4.09 b | 38.99 ± 1.58 ab | 43.49 ± 0.35 a |
| n-6 | 30.95 ± 3.26 b | 29.67 ± 1.34 b | 40.96 ± 0.28 a |
| n-3 | 5.07 ± 0.83 b | 9.32 ± 1.26 a | 2.52 ± 0.11 c |
| n-6/n-3 | 6.15 ± 0.37 b | 3.22 ± 0.47 c | 16.25 ± 0.66 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, H.-K.; Xiang, H.; Park, S.; Lee, J.; Lee, J.-J.; Jung, S.; Ha, J.-H. Partial Replacement of Dietary Fat with Polyunsaturated Fatty Acids Attenuates the Lipopolysaccharide-Induced Hepatic Inflammation in Sprague-Dawley Rats Fed a High-Fat Diet. Int. J. Environ. Res. Public Health 2021, 18, 10986. https://doi.org/10.3390/ijerph182010986
Son H-K, Xiang H, Park S, Lee J, Lee J-J, Jung S, Ha J-H. Partial Replacement of Dietary Fat with Polyunsaturated Fatty Acids Attenuates the Lipopolysaccharide-Induced Hepatic Inflammation in Sprague-Dawley Rats Fed a High-Fat Diet. International Journal of Environmental Research and Public Health. 2021; 18(20):10986. https://doi.org/10.3390/ijerph182010986
Chicago/Turabian StyleSon, Hee-Kyoung, Huo Xiang, Seohyun Park, Jisu Lee, Jae-Joon Lee, Sunyoon Jung, and Jung-Heun Ha. 2021. "Partial Replacement of Dietary Fat with Polyunsaturated Fatty Acids Attenuates the Lipopolysaccharide-Induced Hepatic Inflammation in Sprague-Dawley Rats Fed a High-Fat Diet" International Journal of Environmental Research and Public Health 18, no. 20: 10986. https://doi.org/10.3390/ijerph182010986
APA StyleSon, H.-K., Xiang, H., Park, S., Lee, J., Lee, J.-J., Jung, S., & Ha, J.-H. (2021). Partial Replacement of Dietary Fat with Polyunsaturated Fatty Acids Attenuates the Lipopolysaccharide-Induced Hepatic Inflammation in Sprague-Dawley Rats Fed a High-Fat Diet. International Journal of Environmental Research and Public Health, 18(20), 10986. https://doi.org/10.3390/ijerph182010986