
Estrogen and Progesterone Receptor Immunoexpression in Fallopian Tubes among Postmenopausal Women Based on Time since the Last Menstrual Period


 , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Histological Analysis
2.3. Immunohistochemistry
2.4. Quantitative Analysis of Immunoexpression of ERα and PR
2.5. Statistical Analysis
3. Results
3.1. Baseline Characteristics of the Patients
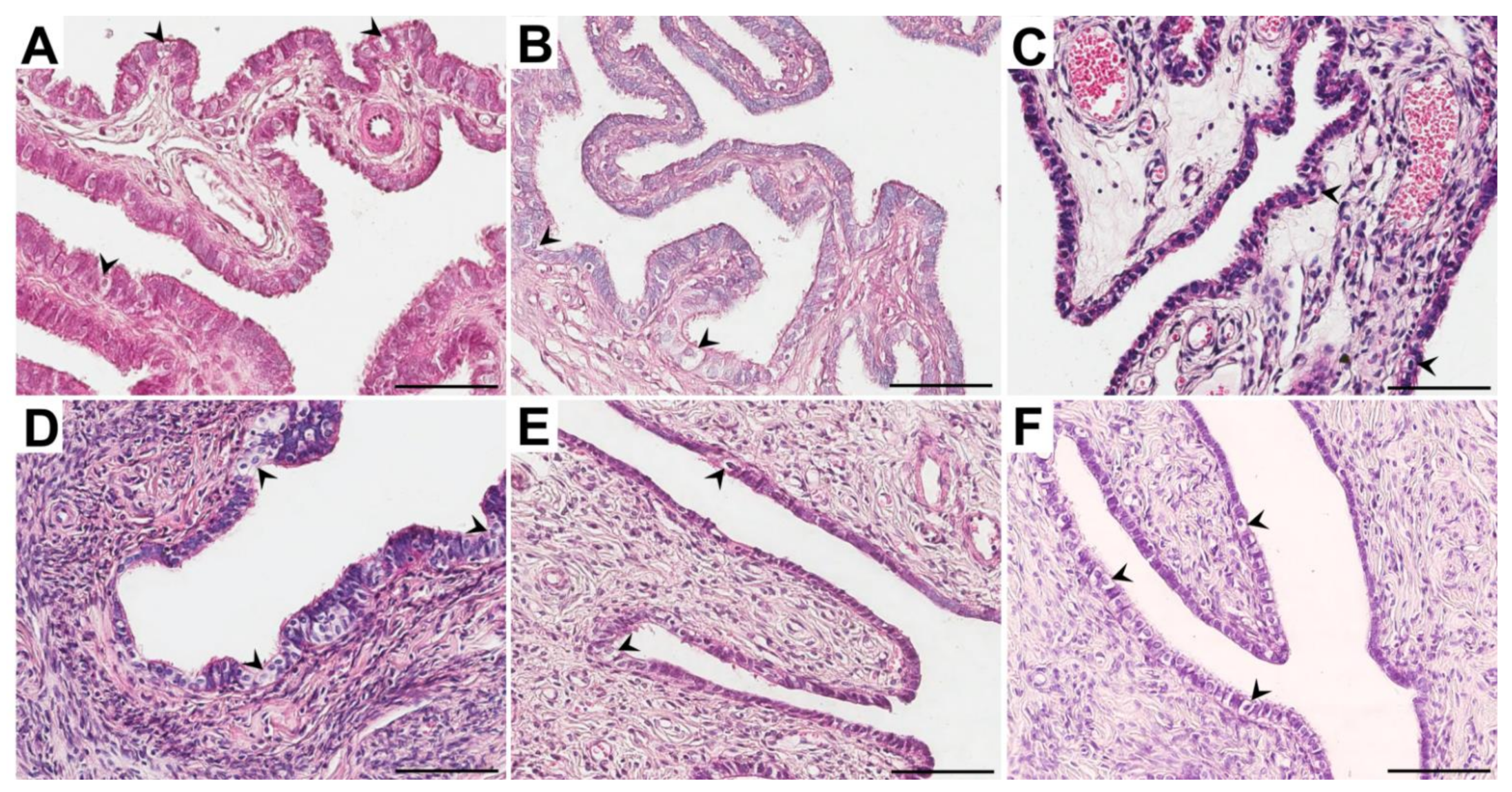
3.2. Morphological Studies
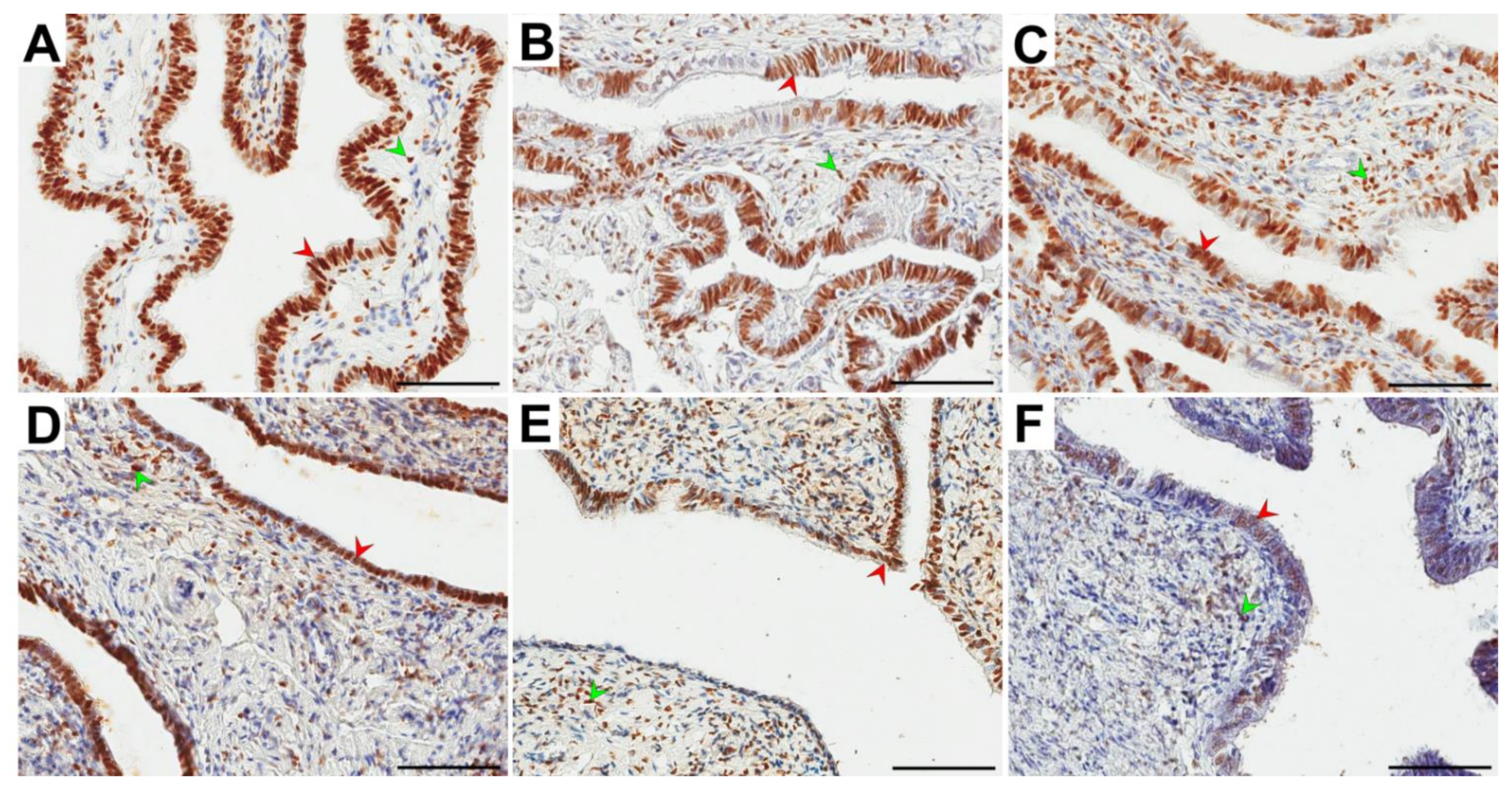
3.3. Immunolocalization and Immunoexpression of ERα
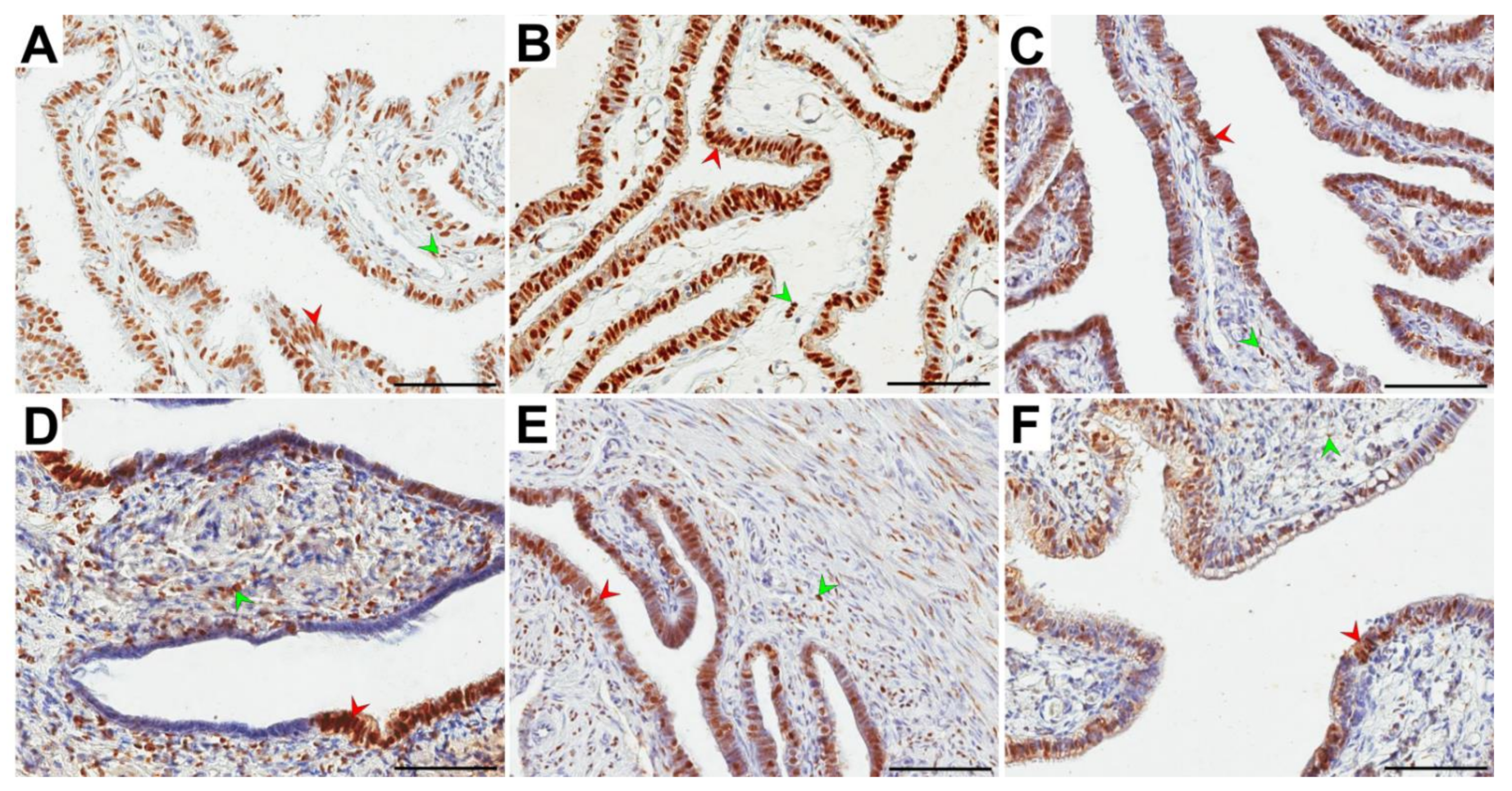
3.4. Immunolocalization and Immunoexpression of PR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Research on the Menopause in the 1990’s; World Health Technical Report Series; World Health Organization: Geneva, Switzerland, 1996.
- Paciuc, J. Hormone Therapy in Menopause. Adv. Exp. Med. Biol. 2020, 1242, 89–120. [Google Scholar]
- Sussman, M.; Trocio, J.; Best, C.; Mirkin, S.; Bushmakin, A.G.; Yood, R.; Friedman, M.; Menzin, J.; Louie, M. Prevalence of menopausal symptoms among mid-life women: Findings from electronic medical records. BMC Womens Health. 2015, 15, 58. [Google Scholar] [CrossRef]
- Kim, C.; Harlow, S.D.; Zheng, H.; McConnell, D.S.; Randolph, J.F., Jr. Changes in androstenedione, dehydroepiandrosterone, testosterone, estradiol, and estrone over the menopausal transition. Womens Midlife Health 2017, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Santoro, N.; Randolph, J.F., Jr. Reproductive hormones and the menopause transition. Obstet. Gynecol. Clin. North Am. 2011, 38, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Safwat, M.D.; Habib, F.A.; Oweiss, N.Y. Distribution of macrophages in the human fallopian tubes: An immunohistochemical and electron microscopic study. Folia Morphol. Warsz. 2008, 67, 43–52. [Google Scholar] [PubMed]
- Deroo, B.; Korach, K. Estrogen receptors and human disease. J. Clin. Investig. 2006, 116, 561–570. [Google Scholar] [CrossRef]
- Regidor, P.A. Progesterone in Peri- and Postmenopause: A Review. Geburtshilfe Frauenheilkd. 2014, 74, 995–1002. [Google Scholar] [CrossRef]
- Prior, J.C. Progesterone for the prevention and treatment of osteoporosis in women. Climacteric 2018, 21, 366–374. [Google Scholar] [CrossRef]
- Yaşar, P.; Ayaz, G.; User, S.D.; Güpür, G.; Muyan, M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod. Med. Biol. 2016, 16, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Paterni, I.; Granchi, C.; Katzenellenbogen, J.A.; Minutolo, F. Estrogen receptors alpha (ERα) and beta (ERβ): Subtype-selective ligands and clinical potential. Steroids 2014, 90, 13–29. [Google Scholar] [CrossRef]
- Lee, H.R.; Kim, T.H.; Choi, K.C. Functions and physiological roles of two types of estrogen receptors, ERα and ERβ, identified by estrogen receptor knockout mouse. Lab. Anim. Res. 2012, 28, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Eriksson, H.; Sahlin, L. Estrogen receptors alpha and beta in the female reproductive tract of the rat during the estrous cycle. Biol. Reprod. 2000, 63, 1331–1340. [Google Scholar] [CrossRef]
- Mowa, C.N.; Iwanaga, T. Developmental changes of the oestrogen receptor-alpha and -beta mRNAs in the female reproductive organ of the rat—an analysis by in situ hybridization. J. Endocrinol. 2000, 167, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Weijdegard, B.; Fernandez-Rodriguez, J.; Egecioglu, E.; Zhu, C.; Andersson, N.; Thurin-Kjellberg, A.; Bergh, C.; Billig, H. Ciliated epithelial-specific and regional-specific expression and regulation of the estrogen receptor-beta2 in the fallopian tubes of immature rats: A possible mechanism for estrogen-mediated transport process in vivo. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E147–E158. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shao, R.; Egecioglu, E.; Weijdegård, B.; Kopchick, J.J.; Fernandez-Rodriguez, J.; Andersson, N.; Billig, H. Dynamic regulation of estrogen receptor-alpha isoform expression in the mouse fallopian tube: Mechanistic insight into estrogen-dependent production and secretion of insulin-like growth factors. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1430–E1442. [Google Scholar] [CrossRef] [PubMed]
- Amso, N.N.; Crow, J.; Shaw, R.W. Comparative immunohistochemical study of oestrogen and progesterone receptors in the fallopian tube and uterus at different stages of the menstrual cycle and the menopause. Hum. Reprod. 1994, 9, 1027–1037. [Google Scholar] [CrossRef]
- Shah, A.; Nandedkar, T.D.; Raghavan, V.P.; Parulekar, S.V.; Natraj, U. Characterization and localization of estrogen and progesterone receptors of human fallopian tube. Indian J. Exp. Biol. 1999, 37, 893–899. [Google Scholar]
- Punnonen, R.; Lukola, A. Binding of estrogen and progestin in the human fallopian tube. Fertil. Steril. 1981, 36, 610–614. [Google Scholar] [CrossRef]
- Urabe, R.; Hachisuga, T.; Ueda, T.; Kawagoe, T.; Kurita, T.; Kagami, S.; Hisaoka, M.; Fujino, Y. Effect of menopause on hormonal receptors in ampullae of the fallopian tube with a special reference to the p53 signature. Int. J. Womens Health. 2017, 9, 163–169. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pollow, K.; Inthraphuvasak, J.; Manz, B.; Grill, H.J.; Pollow, B. A comparison of cytoplasmic and nuclear estradiol and progesterone receptors in human fallopian tube and endometrial tissue. Fertil. Steril. 1981, 36, 615–622. [Google Scholar] [CrossRef]
- Coppens, M.T.; de Boever, J.G.; Dhont, M.A.; Serreyn, R.F.; Vandekerckhove, D.A.; Roels, H.J. Topographical distribution of oestrogen and progesterone receptors in the human endometrium and fallopian tube. An immunocytochemical study. Histochemistry 1993, 99, 127–131. [Google Scholar] [CrossRef]
- Press, M.F.; Nousek-Goebl, N.A.; Bur, M.; Greene, G.L. Estrogen receptor localization in the female genital tract. Am. J. Pathol. 1986, 123, 280–292. [Google Scholar] [CrossRef][Green Version]
- Land, J.A.; Arends, J.W. Immunohistochemical analysis of estrogen and progesterone receptors in fallopian tubes during ectopic pregnancy. Fertil. Steril. 1992, 58, 335–337. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Bancroft, J.D., Gamble, M., Eds.; Churchill Livingstone: London, UK, 2002; pp. 135–136. [Google Scholar]
- Amso, N.N.; Crow, J.; Lewin, J.; Shaw, R.W. A comparative morphological and ultrastructural study of endometrial gland and fallopian tube epithelia at different stages of the menstrual cycle and the menopause. Hum. Reprod. 1994, 9, 2234–2241. [Google Scholar] [CrossRef]
- Crow, J.; Amso, N.N.; Lewin, J.; Shaw, R.W. Morphology and ultrastructure of fallopian tube epithelium at different stages of the menstrual cycle and menopause. Hum. Reprod. 1994, 9, 2224–2233. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Casanas-Roux, F.; Ferin, J.; Thomas, K. Changes in ciliation and cell height in human tubal epithelium in the fertile and post-fertile years. Maturitas 1983, 5, 39–45. [Google Scholar] [CrossRef]
- Correr, S.; Makabe, S.; Heyn, R.; Relucenti, M.; Naguro, T.; Familiari, G. Microplicae-like structures of the fallopian tube in postmenopausal women as shown by electron microscopy. Histol. Histopathol. 2006, 21, 219–226. [Google Scholar]
- Makabe, S.; Motta, P.M.; Naguro, T.; Vizza, E.; Perrone, G.; Zichella, L. Microanatomy of the female reproductive organs in postmenopause by scanning electron microscopy. Climacteric 1998, 1, 63–71. [Google Scholar] [CrossRef]
- Robertson, D.M.; Landgren, B.M. Oestradiol receptor levels in the human fallopian tube during the menstrual cycle and after menopause. J. Steroid Biochem. 1975, 6, 511–513. [Google Scholar] [CrossRef]
- Robertson, D.M.; Landgren, B.M.; Guerrero, R. Oestradiol receptor levels in the human fallopian tube during the menstrual cycle. Acta Endocrinol. 1975, 80, 705–718. [Google Scholar] [CrossRef]
- Pollow, K.; Inthraphuvasak, J.; Grill, H.J.; Manz, B. Estradiol and progesterone binding components in the cytosol of normal human fallopian tubes. J. Steroid Biochem. 1982, 16, 429–435. [Google Scholar] [CrossRef]
- Noe, G. Sex hormone binding globulin expression and colocalization with estrogen receptor in the human Fallopian tube. J. Steroid Biochem. Mol. Biol. 1999, 68, 111–117. [Google Scholar] [CrossRef]
- Teilmann, S.C.; Clement, C.A.; Thorup, J.; Byskov, A.G.; Christensen, S.T. Expression and localization of the progesterone receptor in mouse and human reproductive organs. J. Endocrinol. 2006, 191, 525–535. [Google Scholar] [CrossRef]
- Nutu, M.; Weijdegard, B.; Thomas, P.; Bergh, C.; Thurin-Kjellberg, A.; Pang, Y.; Billig, H.; Larsson, D.G. Membrane progesterone receptor gamma: Tissue distribution and expression in ciliated cells in the fallopian tube. Mol. Reprod. Dev. 2007, 74, 843–850. [Google Scholar] [CrossRef]
- Nutu, M.; Weijdegard, B.; Thomas, P.; Thurin-Kjellberg, A.; Billig, H.; Larsson, D.G. Distribution and hormonal regulation of membrane progesterone receptors beta and gamma in ciliated epithelial cells of mouse and human fallopian tubes. Reprod. Biol. Endocrinol. 2009, 7, 1477–7827. [Google Scholar] [CrossRef]
- Lowder, J.L.; Oliphant, S.S.; Ghetti, C.; Burrows, L.J.; Meyn, L.A.; Balk, J. Prophylactic bilateral oophorectomy or removal of remaining ovary at the time of hysterectomy in the United States, 1979-2004. Am. J. Obstet. Gynecol. 2010, 202, e1–e9. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, V.L.; Vittinghoff, E.; Nakagawa, S.; Jackson, R.; Richter, H.E.; Chan, J.; Kuppermann, M. Factors associated with undergoing bilateral salpingo-oophorectomy at the time of hysterectomy for benign conditions. Obstet. Gynecol. 2009, 113, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Dietl, J.; Wischhusen, J.; Häusler, S.F. The post-reproductive Fallopian tube: Better removed? Hum. Reprod. 2011, 26, 2918–2924. [Google Scholar] [CrossRef]
- McAlpine, J.N.; Hanley, G.E.; Woo, M.M.; Tone, A.A.; Rozenberg, N.; Swenerton, K.D.; Gilks, C.B.; Finlayson, S.J.; Huntsman, D.G.; Miller, D.M. Ovarian Cancer Research Program of British Columbia. Opportunistic salpingectomy: Uptake, risks, and complications of a regional initiative for ovarian cancer prevention. Am. J. Obstet. Gynecol. 2014, 210, 471.e1. [Google Scholar] [CrossRef]
- Hanley, G.E.; Kwon, J.S.; McAlpine, J.N.; Huntsman, D.G.; Finlayson, S.J.; Miller, D. Examining indicators of early menopause following opportunistic salpingectomy: A cohort study from British Columbia, Canada. Am. J. Obstet. Gynecol. 2020, 223, 211.e1–e221. [Google Scholar] [CrossRef]
- Lee, Y.; Miron, A.; Drapkin, R.; Nucci, M.R.; Medeiros, F.; Saleemuddin, A.; Garber, J.; Birch, C.; Mou, H.; Gordon, R.W.; et al. A candidate precursor to serous carcinoma that originates in the distal fallopian tube. J. Pathol. 2007, 211, 26–35. [Google Scholar] [CrossRef]
- Li, J.; Fadare, O.; Xiang, L.; Kong, B.; Zheng, W. Ovarian serous carcinoma: Recent concepts on its origin and carcinogenesis. J. Hematol. Oncol. 2012, 5, 1756–8722. [Google Scholar] [CrossRef] [PubMed]
- Hickey, M.; Ambekar, M.; Hammond, I. Should the ovaries be removed or retained at the time of hysterectomy for benign disease? Hum. Reprod. Update. 2010, 16, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, V.L.; Grady, D.; Wactawski-Wende, J.; Manson, J.E.; Allison, M.A.; Kuppermann, M.; Sarto, G.E.; Robbins, J.; Phillips, L.; Martin, L.W.; et al. Oophorectomy vs ovarian conservation with hysterectomy: Cardiovascular disease, hip fracture, and cancer in the Women’s Health Initiative Observational Study. Arch. Intern. Med. 2011, 171, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.H. Components of oviduct physiology in eutherian mammals. Biol. Rev. Camb. Philos. Soc. 2012, 87, 244–255. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Group (X ± SD) | ||
|---|---|---|---|
| A (n = 40) | B (n = 30) | C (n = 30) | |
| Calendar age (years) | 53.6 a ± 2.4 | 57.9 a ± 4.6 | 67.3 a ± 7.3 |
| Age at LMP (years) | 50.8 ± 4.4 | 50.2 ± 3.3 | 50.7 ± 3.9 |
| BMI (kg/m2) | 24.6 b ± 3.0 | 28.5 b ± 4.6 | 29.2 b ± 6.4 |
| Group | Glandular Epithelium Median (Range) X ± SD | Stroma Median (Range) X ± SD | |
|---|---|---|---|
| A | ampulla | 85.8 a,c (30.8−95.9) 81.2 ± 12.8 | 49.8 c (25.0−75.4) 52.4 ± 13.1 |
| B | 81.3 d (34.8−90.6) 76.6 ± 12.9 | 50.6 d (26.1−75.4) 49.5 ± 15.2 | |
| C | 73.4 a,e (34.1−89.7) 68.5 ± 13.9 | 51.5 e (22.0−71.1) 46.4 ± 14.4 | |
| A | isthmus | 84.2 b,f (68.2−94.0) 83.3 ± 6.9 | 57.0 f (25.2−72.9) 53.6 ± 13.2 |
| B | 83.9 g (35.1−91.6) 78.6 ± 14.8 | 56.4 g (24.8−72.9) 51.7 ± 15.5 | |
| C | 76.8 b,h (24.4−92.6) 68.9 ± 18.1 | 46.4 h (29.5−71.0) 51.1 ± 13.6 | |
| Group | Glandular Epithelium Median (Range) X ± SD | Stroma Median (Range) X ± SD | |
|---|---|---|---|
| A | ampulla | 86.4 a,c,f (68.3−98.8) 85.6 ± 7.5 | 53.6 f (37.3−81.4) 54.2 ± 12.2 |
| B | 86.0 d,g (56.2−94.8) 83.7 ± 8.6 | 52.0 g (34.0−80.8) 51.8 ± 8.1 | |
| C | 74.8 a,e,h (51.7−91.3) 72.2 ± 13.4 | 48.7 h (26.4−69.9) 49.4 ± 11.0 | |
| A | isthmus | 75.0 b,c,i (51.0−91.8) 73.2 ± 11.7 | 48.5 i (25.9−75.8) 50.4 ± 11.4 |
| B | 68.3 d,j (47.7−80.2) 65.7 ± 10.2 | 47.8 j (21.7−64.0) 47.2 ± 11.4 | |
| C | 55.9 b,e,k (41.1−80.1) 59.6 ± 10.1 | 45.0 k (21.5−65.4) 45.7 ± 11.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brodowska, A.; Grabowska, M.; Bittel, K.; Ciećwież, S.; Brodowski, J.; Szczuko, M.; Szydłowska, I.; Piasecka, M. Estrogen and Progesterone Receptor Immunoexpression in Fallopian Tubes among Postmenopausal Women Based on Time since the Last Menstrual Period. Int. J. Environ. Res. Public Health 2021, 18, 9195. https://doi.org/10.3390/ijerph18179195
Brodowska A, Grabowska M, Bittel K, Ciećwież S, Brodowski J, Szczuko M, Szydłowska I, Piasecka M. Estrogen and Progesterone Receptor Immunoexpression in Fallopian Tubes among Postmenopausal Women Based on Time since the Last Menstrual Period. International Journal of Environmental Research and Public Health. 2021; 18(17):9195. https://doi.org/10.3390/ijerph18179195
Chicago/Turabian StyleBrodowska, Agnieszka, Marta Grabowska, Katarzyna Bittel, Sylwester Ciećwież, Jacek Brodowski, Małgorzata Szczuko, Iwona Szydłowska, and Małgorzata Piasecka. 2021. "Estrogen and Progesterone Receptor Immunoexpression in Fallopian Tubes among Postmenopausal Women Based on Time since the Last Menstrual Period" International Journal of Environmental Research and Public Health 18, no. 17: 9195. https://doi.org/10.3390/ijerph18179195
APA StyleBrodowska, A., Grabowska, M., Bittel, K., Ciećwież, S., Brodowski, J., Szczuko, M., Szydłowska, I., & Piasecka, M. (2021). Estrogen and Progesterone Receptor Immunoexpression in Fallopian Tubes among Postmenopausal Women Based on Time since the Last Menstrual Period. International Journal of Environmental Research and Public Health, 18(17), 9195. https://doi.org/10.3390/ijerph18179195