
Effects of Sleep Deprivation by Olfactorily Induced Sexual Arousal Compared to Immobilization Stress and Manual Sleep Deprivation on Neuromessengers and Time Keeping Genes in the Suprachiasmatic Nuclei and Other Cerebral Entities of Syrian Hamsters—An Immunohistochemical Study


Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Animals
2.2. Pre-Tests Regarding the Animals’ Behavior
2.3. Sexual Stimulation, Immobilization Stress, and Gentle Manual Handling
- Group 1, sexual stimulation with ahrodisin and females: A hamster cage (Makrolon cage type III) was divided in its centre by a double layer of flywire. Two cellulose balls soaked with the aphrodisin-DMDS-mixture were placed under the bedding of one area, five adult female hamsters (age ≥ 3 months) in the other area. The female Syrian hamsters (purchased from Charles River, Sulzfeld, Germany) were kept under the same conditions as the males but completely separated in another room. According to literature, female hamsters are sexually sensible for one day each 4 to 7 days at median daily temperature of 15 °C [28]. When the experiment started, five male hamsters were placed in the pheromone-containing compartment. They were able to see, hear and smell the females through the wire-netting without being able to physically contact them. The aphrodisin-DMDS-mix was additionally applied to the cage with a vaporizer. This experiment was performed twice, resulting in materials from 10 hamsters.
- Group 2, sexual stimulation with aphrodisin solely: The experimental setting of the pheromone-only group consisting of five hamsters was the same as described for group 1 with the exception that no female animals were present.
- Group 3, manual sleep deprivation: The manual sleep deprivation group was deprived of sleep by gentle manual handling whenever the hamsters tended to fall asleep during the same period of the diurnal phase starting at ZT 00:00. This group was subdivided into 10 animals sleep deprived for 1 h (ZT 00:00 to ZT 01:00, later referred to as group 3a (group 3a represents the principle control group for groups 1, 2 and 4)), five hamsters sleep deprived for 6 h (ZT 23:00 to ZT 05:00, later referred to as group 3b) and nine animals sleep-deprived for 20 h (ZT 09:00 to ZT 05:00 of the following day, later referred to as group 3c). (Groups 3b and 3c were introduced for deciphering possible changes after different durations of sleep deprivation).
- Group 4, sleep deprivation by stress: Finally, the last comparison group was exposed to immobilization stress by nearly complete immobilization caused by wrapping them with extra strong scotch tape. The head remained unwrapped. Thus, ventilation was still guaranteed. All comparison groups for the sexual stimulation group were also sacrificed at zeitgeber time 01.00.
- Group 5, sleep control: Nine hamsters were sacrificed after undisturbed sleep for 5 h at ZT 05:00. A further group studied after 1 h of sleep was not used, since there are no significant changes in comparison to 5 h of sleep, however, the experimental setup to prevent the effects of disturbed sleep after 1 h is rather complicated as shown in [29].
2.4. Sample Acquisition and Preparation
2.5. Immunohistochemistry
2.6. Blood Analyses
2.7. Statistics
3. Results
3.1. Behavioral Changes
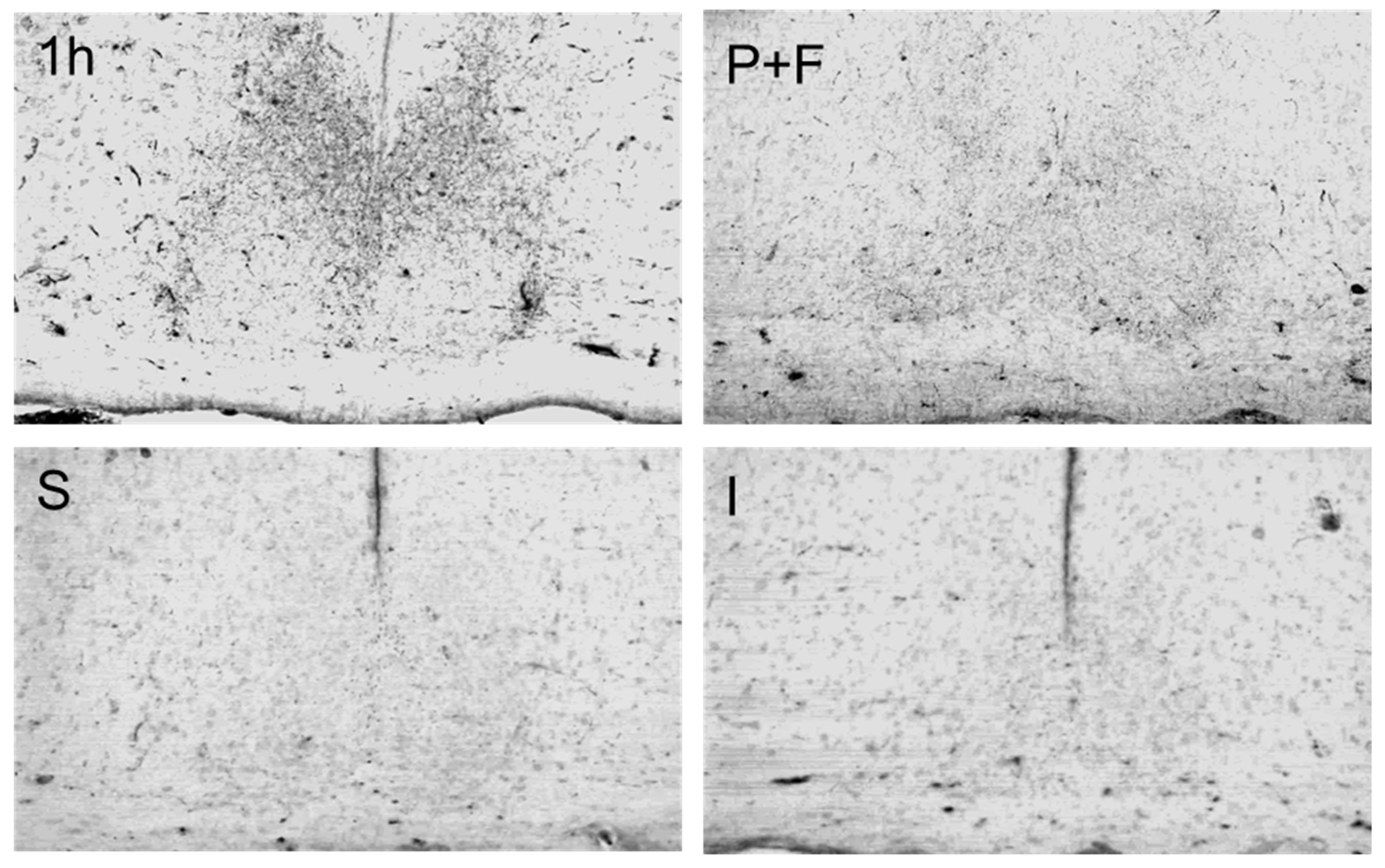
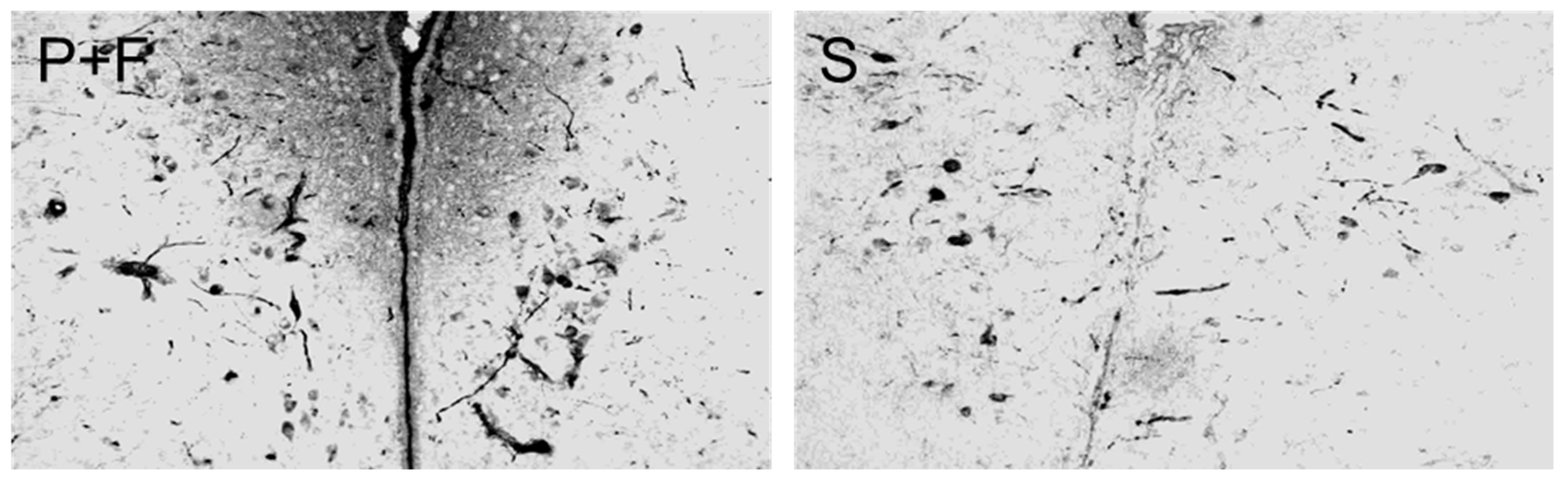
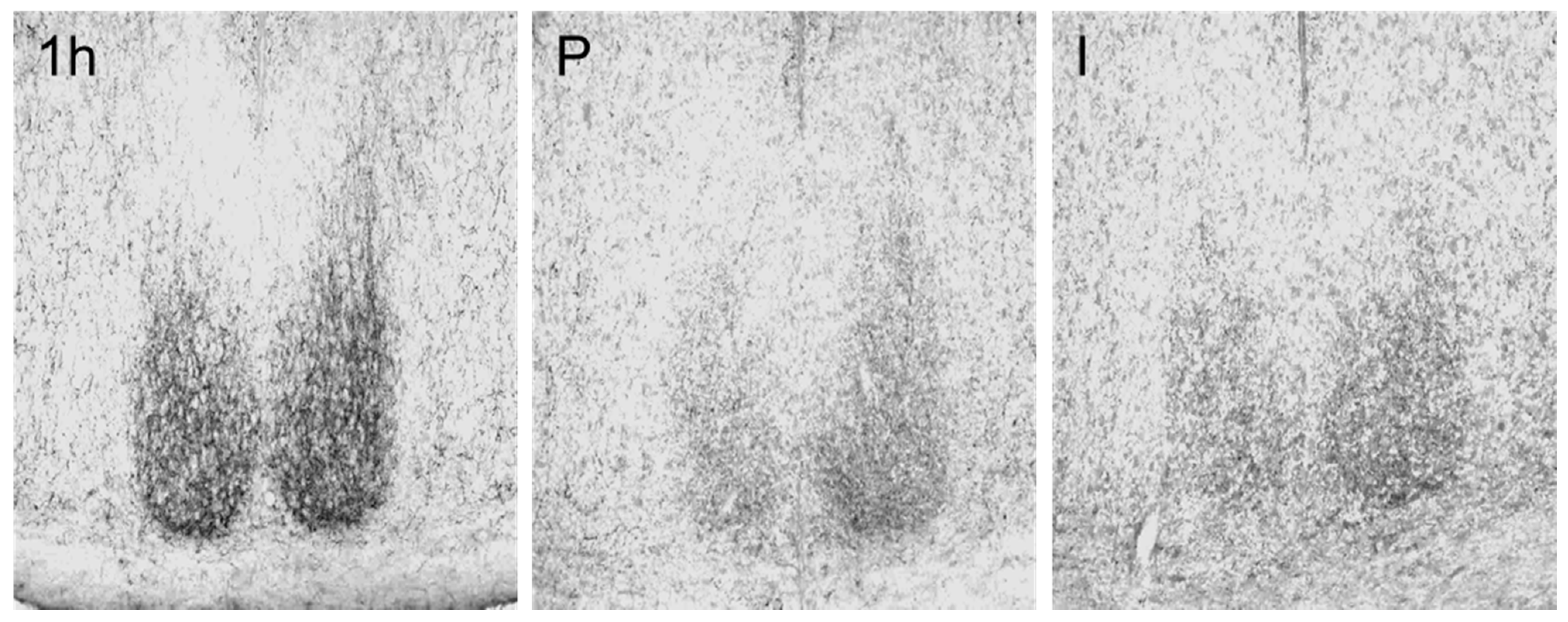
3.2. Changes of Immunoreactivity to Neuromessengers, Time Keeping Proteins and Steroid Receptors in Cerebral Entities as Well as Their Concentrations in Blood
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A















References
- Pedersen, C.A.; Boccia, M.L. Vasopressin interactions with oxytocin in the control of female sexual behavior. Neuroscience 2006, 139, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S. Oxytocin and sexual behavior. Neurosci. Biobehav. Rev. 1992, 16, 131–144. [Google Scholar] [CrossRef]
- Swaab, D.F. The Role of Hypothalamus and Endocrine System in Sexuality. In Biological Substrates of Human Sexuality; Hyde, J.S., Ed.; American Psychological Association: Washington, DC, USA, 2005; pp. 21–74. [Google Scholar] [CrossRef]
- Wood, R.I.; Williams, S.J. Steroidal control of male hamster sexual behaviour in Me and MPOA: Effects of androgen dose and tamoxifen. Physiol. Behav. 2001, 72, 727–733. [Google Scholar] [CrossRef]
- Rampin, O.; Jerome, N.; Suaudeau, C. Proerectile effects of apomorphine in mice. Life Sci. 2003, 72, 2329–2336. [Google Scholar] [CrossRef]
- Azizi, V.; Oryan, S.; Khazali, H.; Hosseini, A. Central injection of neuropeptide Y modulates sexual behavior in male rats: Interaction with GnRH and kisspeptin/neurokinin B/dynorphin. Int. J. Neurosci. 2020, 20, 780–788. [Google Scholar]
- Moreira, N.; Sandini, T.M.; Reis-Silva, T.M.; Navas-Suáresz, P.; Auada, A.V.V.; Lebrun, I.; Flório, J.C.; Bernardi, M.M.; Spinosa, H.S. Ivermectin reduces motor coordination, serum testosterone, and central neurotransmitter levels but does not affect sexual motivation in male rats. Reprod. Toxicol. 2017, 74, 195–203. [Google Scholar] [CrossRef] [PubMed]
- van Furth, W.R.; van Ree, J.M. Sexual motivation: Involvement of endogenous opioids in the ventral tegmental area. Brain Res. 1996, 279, 20–28. [Google Scholar] [CrossRef]
- Kapicioglu, S.; Mollamehmetoglu, M.; Kutlu, N.; Can, G.; Ozgur, G.K. Inhibition of penile erection in rats by a long-acting somatostatin analogue, octreotide (SMS 201-995). Br. J. Urol. 1998, 81, 142–145. [Google Scholar] [CrossRef]
- Hull, E.M.; Lorrain, D.S.; Du, J.; Matuszewich, L.; Lumley, L.A.; Putnam, S.K.; Moses, J. Hormone-neurotransmitter interactions in the control of sexual behavior. Behav. Brain Res. 1999, 105, 105–116. [Google Scholar] [CrossRef]
- Weaver, D.R. The suprachiasmatic nucleus: A 25-year retrospective. J. Biol. Rhythm. 1998, 13, 100–112. [Google Scholar] [CrossRef]
- Ottesen, B.; Ulrichsen, H.; Fahrenkrug, J.; Larsen, J.J.; Wagner, G.; Schierup, L.; Sondergaard, F. Vasoactive intestinal polypeptide and the female genital tract: Relationship to reproductive phase and delivery. Am. J. Obstet. Gynecol. 1982, 143, 414–420. [Google Scholar] [CrossRef]
- Yamada, K.; Wada, E.; Wada, K. Female gastrin-releasing peptide receptor (GRP-R)-deficient mice exhibit altered social preference for male conspecifics: Implications for GRP/GRP-R modulation of GABAergic function. Brain Res. 2001, 894, 281–287. [Google Scholar] [CrossRef]
- Sakamoto, H. Brain-spinal cord neural circuits controlling male sexual function and behavior. Neurosci. Res. 2012, 72, 103–116. [Google Scholar] [CrossRef]
- Vrang, N.; Larsen, P.J.; Mikkelsen, J.D. Direct projection from the suprachiasmatic nucleus to hypophysiotrophic corticotropin-releasing factor immunoreactive cells in the paraventricular nucleus of the hypothalamus demonstrated by means of Phaseolus vulgaris-leucoagglutinin tract tracing. Brain Res. 1995, 684, 61–69. [Google Scholar] [CrossRef]
- Cui, L.N.; Saeb-Parsy, K.; Dyball, R.E. Neurones in the supraoptic nucleus of the rat are regulated by a projection from the suprachiasmatic nucleus. J. Physiol. 1997, 502 Pt 1, 149–159. [Google Scholar] [CrossRef]
- Vincent, S.R.; McIntosh, C.H.; Buchan, A.M.; Brown, J.C. Central somatostatin systems revealed with monoclonal antibodies. J. Comp. Neurol. 1985, 238, 169–186. [Google Scholar] [CrossRef] [PubMed]
- Glass, J.D.; DiNardo, L.A.; Ehlen, J.C. Dorsal raphe nuclear stimulation of SCN serotonin release and circadian phase-resetting. Brain Res. 2000, 859, 224–232. [Google Scholar] [CrossRef]
- Glass, J.D.; Grossman, G.H.; Farnbauch, L.; DiNardo, L. Midbrain raphe modulation of nonphotic circadian clock resetting and 5-HT release in the mammalian suprachiasmatic nucleus. J. Neurosci. 2003, 23, 7451–7460. [Google Scholar] [CrossRef]
- Pudovkina, O.L.; Cremers, T.I.; Westerink, B.H. The interaction between the locus coeruleus and dorsal raphe nucleus studied with dual-probe microdialysis. Eur. J. Pharmacol. 2002, 445, 37–42. [Google Scholar] [CrossRef]
- González, M.M.; Aston-Jones, G. Circadian regulation of arousal: Role of the noradrenergic locus coeruleus system and light exposure. Sleep 2006, 29, 1327–1336. [Google Scholar] [CrossRef]
- Deboer, T.; Détári, L.; Meijer, J.H. Long term effects of sleep deprivation on the mammalian circadian pacemaker. Sleep 2007, 30, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Petrulis, A.; Johnston, R.E. A reevaluation of dimethyl disulfide as a sex attractant in golden hamsters. Physiol. Behav. 1995, 57, 779–784. [Google Scholar] [CrossRef]
- Briand, L.; Huet, J.C.; Perez, V.; Lenoir, G.; Nespoulous, C.; Boucher, Y.; Trotier, D.; Pernollet, J.C. Odorant and pheromone binding by aphrodisn, a hamster aphrodisiac protein. FEBS Lett. 2000, 476, 179–185. [Google Scholar] [CrossRef]
- Jang, T.; Singer, A.G.; O’Connell, R.J. Induction of c-fos in hamster accessory olfactory bulbs by natural and cloned aphrodisin. Neuroreport 2001, 12, 449. [Google Scholar] [CrossRef]
- Naylor, E.; Buxton, O.M.; Bergmann, B.M.; Easton, A.; Phyllis, C.; Zee, P.C.; Turek, F.W. Effects of Aging on Sleep in the Golden Hamster. Sleep 1998, 21, 687–693. [Google Scholar] [CrossRef]
- Mägert, H.J.; Cieslak, A.; Alkan, O.; Lüscher, B.; Kauffels, W.; Forssmann, W.G. The golden hamster aphrodisin gene. Structure, expression in parotid glands of female animals, and comparison with a similar murine gene. J. Biol. Chem. 1999, 274, 444–450. [Google Scholar] [CrossRef]
- Bielfeld, H. Der Goldhamster. 2. Auflage. Ulmer Stuttgart 1992, 72–74. Available online: http://d-nb.info/930466705 (accessed on 26 August 2021).
- Schilling, J.; Nürnberger, F. Dynamic changes in the immunoreactivity of neuropeptide systems of the suprachiasmatic nuclei in golden hamsters during the sleep-wake cycle. Cell Tissue Res. 1998, 294, 233–241. [Google Scholar] [CrossRef]
- Zamboni, L.; De Martino, C. Buffered picric acid-formaldehyde: A new rapid fixative for electron microscopy. J. Cell Biol. 1967, 35, 148A. [Google Scholar]
- Nürnberger, F.; Schindler, C.U.; Kriete, A. The serotonin-immunoreactive system of the suprachiasmatic nucleus in the hibernating ground squirrel, Spermophilus richardsonii. Cell Tissue Res. 1989, 256, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual immunohistochemistry staining of mouse tissues using the avidin-biotin complex (ABC) technique. Cold Spring Harb. Protoc. 2014, 2014, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Card, J.P.; Moore, R.Y. The suprachiasmatic nucleus of the golden hamster: Immunohistochemical analysis of cell and fiber distribution. Neuroscience 1984, 13, 415–431. [Google Scholar] [CrossRef]
- Moore, R.Y.; Speh, J.C.; Leak, R.K. Suprachiasmatic nucleus organization. Cell Tissue Res. 2002, 309, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Engel, S. Camera lucida tracings in medical research. Med. Biol. Illus. 1955, 5, 71–74. [Google Scholar] [PubMed]
- Forer, A. A camera-lucida procedure for low-contrast material. J. R. Microsc. Soc. 1968, 88, 611–613. [Google Scholar] [CrossRef]
- Andy, O.J.; Kurimoto, T.; Velamati, S.; Peeler, D. Limbic stimulation-induced hypersexuality. Levels of sexual drive. Pavlov. J. Biol. Sci. 1983, 18, 22–35. [Google Scholar]
- Tsuchiya, T.; Horii, I. Different effects of acute and chronic immobilization stress on plasma testosterone levels in male Syrian hamsters. Psychoneuroendocrinology 1995, 20, 95–102. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Taylor, M.R.; Mann, D.R. Effect of immobilization stress on plasma luteinizing hormone, testosterone, and corticosterone concentrations and on 3 beta-hydroxysteroid dehydrogenase activity in the testes of adult rats. Proc. Soc. Exp. Biol. Med. 1993, 204, 231–235. [Google Scholar] [CrossRef]
- Demura, R.; Suzuki, T.; Nakamura, S.; Komatsu, H.; Odagiri, E.; Demura, H. Effect of immobilization stress on testosterone and inhibin in male rats. J. Androl. 1989, 10, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; Teclemariam-Mesbah, R.; Pevet, P. Efferent projections of the suprachiasmatic nucleus in the golden hamster (Mesocricetus auratus). J. Comp. Neurol. 1993, 332, 293–314. [Google Scholar] [CrossRef]
- Jirikowski, G.F.; Caldwell, J.D.; Haussler, H.U.; Pedersen, C.A. Mating alters topography and content of oxytocin immunoreactivity in male mouse brain. Cell Tissue Res. 1991, 266, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Smock, T.; Albeck, D.; Stark, P. A peptidergic basis for sexual behaviour in mammals. Prog. Brain Res. 1998, 119, 467–481. [Google Scholar] [PubMed]
- Sodersten, P.; Boer, G.J.; De Vries, G.J.; Buijs, R.M.; Melin, P. Effects of vasopressin on female sexual behaviour in male rats. Neurosci. Lett. 1986, 69, 188–191. [Google Scholar] [CrossRef][Green Version]
- Jezova, D.; Skultetyova, I.; Tokarev, D.I.; Bakos, P.; Vigas, M. Vasopressin and oxytocin in stress. Ann. N. Y. Acad. Sci. 1995, 771, 192–203. [Google Scholar] [CrossRef]
- de Goeij, D.C.; Jezova, D.; Tilders, F.J. Repeated stress enhances vasopressin synthesis in corticotropin releasing factor neurons in the paraventricular nucleus. Brain Res. 1992, 577, 165–168. [Google Scholar] [CrossRef]
- Segraves, R.T. Effects of psychotropic drugs on human erection and ejaculation. Arch. Gen. Psychiatry 1989, 46, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Levin, R.J. VIP, vagina, clitoral and periurethral glans—an update on human female genital arousal. Exp. Clin. Endocrinol. 1991, 98, 61–69. [Google Scholar] [CrossRef]
- Gavalda, A.; Benyassi, A.; Arancibia, S.; Armario, A. Chronic but not acute exposure to stress is associated with hypothalamic vasoactive intestinal polypeptide (VIP) release into median eminence. J. Neuroendocrinol. 1993, 5, 421–425. [Google Scholar] [CrossRef]
- Iwanaga, T.; Ohtsuka, H.; Adachi, I.; Fujita, T. GRP (gastrin-releasing peptide)-containing nerves in the rat stomach and stress-induced depletion of their synaptic vesicles. An electron microscope study. Arch. Histol. Cytol. 1991, 54, 573–580. [Google Scholar] [CrossRef][Green Version]
- Bitran, D.; Miller, S.A.; McQuade, D.B.; Leipheimer, R.E.; Sachs, B.D. Inhibition of sexual reflexes by lumbosacral injection of a GABA-B agonist in the male rat. Pharmacol. Biochem. Behav. 1988, 31, 657–666. [Google Scholar] [CrossRef]
- Qureshi, G.A.; Bednar, I.; Forsberg, B.G.; Sodersten, P. GABA inhibits sexual behaviour in female rats. Neuroscience 1988, 27, 169–174. [Google Scholar] [CrossRef]
- Fernandez-Guasti, A.; Larsson, K.; Beyer, C. Effect of bicuculline on sexual activity in castrated male rats. Physiol. Behav. 1986, 36, 235–237. [Google Scholar] [CrossRef]
- Navarro Becerra, N.; Munaro, N.I. Gamma-Aminobutyric acid activity in the olfactory bulb of the rat during the sexual cycle and response to olfactory stimuli. Can. J. Physiol. Pharmacol. 1992, 70, 922–925. [Google Scholar] [CrossRef]
- Yoneda, Y.; Kanmori, K.; Ida, S.; Kuriyama, K. Stress-induced alterations in metabolism of gamma-aminobutyric acid in rat brain. J. Neurochem. 1983, 40, 350–356. [Google Scholar] [CrossRef]
- Otero Losada, M.E. Acute stress and GABAergic function in the rat brain. Br. J. Pharmacol. 1989, 96, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Curie, T.; Maret, S.; Emmenegger, Y.; Franken, P. In Vivo Imaging of the Central and Peripheral Effects of Sleep Deprivation and Suprachiasmatic Nuclei Lesion on PERIOD-2 Protein in Mice. Sleep 2015, 38, 1381–1394. [Google Scholar] [CrossRef] [PubMed]
- Mongrain, V.; La Spada, F.; Curie, T.; Franken, P. Sleep loss reduces the DNA-binding of BMAL1, CLOCK, and NPAS2 to specific clock genes in the mouse cerebral cortex. PLoS ONE 2011, 6, e26622. [Google Scholar] [CrossRef] [PubMed]
- Wisor, J.P.; O’Hara, B.F.; Terao, A.; Selby, C.P.; Kilduff, T.S.; Sancar, A.; Edgar, D.M.; Franken, P. A role for cryptochromes in sleep regulation. BMC Neurosci. 2002, 3, 20. [Google Scholar] [CrossRef]
- Yagita, K.; Yamaguchi, S.; Tamanini, F.; van Der Horst, G.T.; Hoeijmakers, J.H.; Yasui, A.; Loros, J.J.; Dunlap, J.C.; Okamura, H. Dimerization and nuclear entry of mPER proteins in mammalian cells. Genes Dev. 2000, 14, 1353–1363. [Google Scholar] [PubMed]
- Herzog, E.D.; Takahashi, J.S.; Block, G.D. Clock controls circadian period in isolated suprachiasmatic nucleus neurons. Nat. Neurosci. 1998, 1, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Varcoe, T.J.; Kennaway, D.J.; Voultsios, A. Activation of 5-HT2C receptors acutely induces Per gene expression in the rat suprachiasmatic nucleus at night. Brain Res. Mol. Brain Res. 2003, 119, 192–200. [Google Scholar] [CrossRef]
- Dornan, W.A.; Malsbury, C.W.; Penney, R.B. Facilitation of lordosis by injection of substance P into the midbrain central gray. Neuroendocrinology 1987, 45, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Takayama, H.; Ota, Z.; Ogawa, N. Effect of immobilization stress on neuropeptides and their receptors in rat central nervous system. Regul. Pept. 1986, 15, 239–248. [Google Scholar] [CrossRef]
- Iumatov, E.A.; Poppai, M.; Ratsak, R. Changes in the substance P content of the blood and hypothalamus during experimental emotional stress. Biull. Eksp. Biol. Med. 1985, 99, 397–399. [Google Scholar] [PubMed]
- Gray, T.S.; Morley, J.E. Neuropeptide Y: Anatomical distribution and possible function in the mammalian nervous system. Life Sci. 1986, 38, 389–401. [Google Scholar] [CrossRef]
- Corder, R.; Castagne, V.; Rivet, J.M.; Mormede, P.; Gaillard, R.C. Central and peripheral effects of repeated stress and high NaCl diet on neuropeptide Y. Physiol. Behav. 1992, 52, 205–210. [Google Scholar] [CrossRef]
- Pralong, F.P.; Corder, R.; Gaillard, R.C. The effects of chronic glucocorticoid excess, adrenalectomy and stress on neuropeptide Y in individual rat hypothalamic nuclei. Neuropeptides 1993, 25, 223–231. [Google Scholar] [CrossRef]
- Makino, S.; Baker, R.A.; Smith, M.A.; Gold, P.W. Differential regulation of neuropeptide Y mRNA expression in the arcuate nucleus and locus coeruleus by stress and antidepressants. J. Neuroendocrinol. 2000, 12, 387–395. [Google Scholar] [CrossRef]
- Grossman, G.H.; Mistlberger, R.E.; Antle, M.C.; Ehlen, J.C.; Glass, J.D. Sleep deprivation stimulates serotonin release in the suprachiasmatic nucleus. Neuroreport 2000, 11, 1929–1932. [Google Scholar] [CrossRef] [PubMed]
- Alfaro-Rodríguez, A.; González-Piña, R.; González-Maciel, A.; Arch-Tirado, E. Serotonin and 5-hydroxy-indole-acetic acid contents in dorsal raphe and suprachiasmatic nuclei in normal, malnourished and rehabilitated rats under 24 h of sleep deprivation. Brain Res. 2006, 1110, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Kvetnanský, R.; Palkovits, M.; Mitro, A.; Torda, T.; Mikulaj, L. Catecholamines in individual hypothalamic nuclei of acutely and repeatedly stressed rats. Neuroendocrinology 1977, 23, 257–267. [Google Scholar] [CrossRef]
- Carmichael, M.S.; Humbert, R.; Dixen, J.; Palmisano, G.; Greenleaf, W.; Davidson, J.M. Plasma oxytocin increases in the human sexual response. J. Clin. Endocrinol. Metab. 1987, 64, 27–31. [Google Scholar] [CrossRef]
- Hillegart, V.; Alster, P.; Uvnas-Moberg, K.; Ahlenius, S. Sexual motivation promotes oxytocin secretion in male rats. Peptides 1998, 19, 39–45. [Google Scholar] [CrossRef]
- Blaicher, W.; Gruber, D.; Bieglmayer, C.; Blaicher, A.M.; Knogler, W.; Huber, J.C. The role of oxytocin in relation to female sexual arousal. Gynecol. Obstet. Investig. 1999, 47, 125–126. [Google Scholar] [CrossRef]
- Melis, M.R.; Spano, M.S.; Succu, S.; Argiolas, A. The oxytocin antagonist d(CH2)5Tyr(Me)2-Orn8-vasotocin reduces non-contact penile erections in male rats. Neurosci. Lett. 1999, 265, 171–174. [Google Scholar] [CrossRef]
- Melis, M.R.; Argiolas, A. Central oxytocinergic neurotransmission: A drug target for the therapy of psychogenic erectile dysfunction. Curr. Drug Targets 2003, 4, 55–66. [Google Scholar] [CrossRef]
- Carmichael, M.S.; Warburton, V.L.; Dixen, J.; Davidson, J.M. Relationships among cardiovascular, muscular, and oxytocin responses during human sexual activity. Arch. Sex. Behav. 1994, 23, 59–79. [Google Scholar] [CrossRef] [PubMed]
- Jezova, D.; Michajlovskij, N.; Kvetnansky, R.; Makara, G.B. Paraventricular and supraoptic nuclei of the hypothalamus are not equally important for oxytocin release during stress. Neuroendocrinology 1993, 57, 776–781. [Google Scholar] [CrossRef]
- Kato, T.; Montplaisir, J.M.; Lavigne, G.J. Experimentally induced arousals during sleep: A matchimg paradigm. J. Sleep Res. 2004, 13, 229–238. [Google Scholar] [CrossRef]
- Zerbi, V.; Floriou-Servou, A.; Markicevic, M.; Vermieren, Y.; Sturman, O.; Privitera, M.; Ziegler, L.V.; Ferrari, K.D.; Weber, B.; DeDeyn, P.P.; et al. Rapid reconfiguration of the functional connectome after chemogenetic Locus coeruleus activation. Neuron 2019, 4, 702–718.e5. [Google Scholar] [CrossRef]
- Viau, V.; Chu, A.; Soriano, L.; Dallman, F. Independent and overlapping effects of corticosterone and testosterone on corticotropin-releasing hormone and arginine vasopressin mRNA expression in the paraventricular nucleus of the hypothalamus and stress-induced adrenocorticotropic hormone release. J. Neurosci. 1999, 19, 6648–6693. [Google Scholar] [CrossRef]
- Winsky-Sonnerer, R.; Yamanaka, A.; Diano, S.; Borok, E.; Roberts, A.J.; Sakurai, T.; Kilduff, T.S.; Horvath, T.L.; de Lecea, L. Interaction between the corticotropin-releasing factor system and hypocretins (Orexins): A novel circuit mediating stress response. J. Neurosci. 2004, 24, 11439–11448. [Google Scholar] [CrossRef] [PubMed]
- Pieper, D.R.; Newman, S.W. Neural pathway from the olfactory bulbs regulating tonic gonadotropin secretion. Neurosci. Biobehav. Rev. 1999, 23, 555–562. [Google Scholar] [CrossRef]
- Model, Z.; Butler, M.P.; LeSauter, J.; Silver, R. Suprachiasmatic nucleus as site of androgen action on circadian rhythms. Horm. Behav. 2015, 73, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.; Bingham, B.; Gray, M.; Innala, L.; Viau, V. The medial preoptic nucleus integrates the central influences of testosterone on the paraventricular nucleus of the hypothalamus and its extended circuitries. J. Neurosci. 2010, 30, 11762–11770. [Google Scholar] [CrossRef][Green Version]
- Staiger, J.F.; Nürnberger, F. The efferent connections of the lateral septal nucleus in the guinea pig: Projections to the diencephalon and brainstem. Cell Tissue Res. 1991, 264, 391–413. [Google Scholar] [CrossRef] [PubMed]
- Torterolo, P.; Sampogna, S.; Morales, F.R.; Chase, M.H. MCH-containing neurons in the hypothalamus of the cat: Searching for a role in the control of sleep and wakefulness. Brain Res. 2006, 1119, 101–114. [Google Scholar] [CrossRef][Green Version]
- Lin, L.; Faraco, J.; Li, R.; Kadotani, H.; Rogers, W.; Lin, X.; Qiu, X.; de Jong, P.J.; Nishino, S.; Mignot, E. The sleep disorder canine narcolepsy is caused by a mutation in the hypocretin (orexin) receptor 2 gene. Cell 1999, 98, 273–408. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody against | Working Dilution | Producer |
|---|---|---|
| AVP | 1:1000 | Own production [29] |
| VIP | 1:1000 | Sigma product, product number: V3508 |
| GRP | 1:500 | Affinity Res. Prod., Exeter, UK |
| GAD-65/67 | 1:1000 | Biotrend Chemikalien GmbH, Köln, Germany |
| 5-HT | 1:1000 | Diasorin, Stillwater, MN, USA |
| SP | 1:1000 | Chemicon International, Temecula, CA, USA |
| NPY | 1:1000 | Affinity Res. Prod., Exeter, UK |
| ME | 1:1000 | Chemicon International, Temecula, CA, USA |
| OXY | 1:1000 | Own production [32] |
| SOM | 1:1000 | Chemicon International, Temecula, CA, USA |
| AR | 1:50 | Novocastra Laboratories, Newcastle, UK |
| OR | 1:50 | Novocastra Laboratories, Newcastle, UK |
| TH | 1:1000 | Chemicon International, Temecula, CA, USA |
| DBH | 1:1000 | Chemicon International, Temecula, CA, USA |
| Clock | 1:60 | Santa Cruz Biotechnology, Inc., Dallas, TX, USA |
| Per 1 | 1:60 | Santa Cruz Biotechnology, Inc., Dallas, TX, USA |
| Per 2 | 1:60 | Alpha Diagnostics Ltd., Reinach, Switzerland |
| Cry 2 | 1:60 | Alpha Diagnostics Ltd., Reinach, Switzerland |
| Parameter | Entity | Group 1: 1 h Exposit. to Pheromones and Females, ZT 01.00, | Group 2: 1 h Exposit. to Pheromones, ZT 01.00, | Group 3a: 1 h Manual Sleep Deprivation, ZT 01.00, | Group 3b: 6 h Manual Sleep Deprivation, ZT 05.00, | Group 3c: 20 h Manual Sleep Deprivation, ZT 05.00, | Group 4: 1 h Exposit. to Immobiliza-tion Stress, ZT 01.00, | Group 5: Sleep Control: Sleeping Animals, ZT 05.00, | p (Significance over All Compared Groups) |
|---|---|---|---|---|---|---|---|---|---|
| AVP 1 | SCN | 0.45% (±0.22%) ↓ | 1.39% (±0.54%) | 5.59% (±2.32%) ↑ | 0.67% (±0.17%) | 1.62% (±0.39%) | 0.62% (±0.36%) ↓ | 0.36% (±0.47%) ↓ | 0.0002 |
| VIP 2 | SCN | 0.02% (±0.02%) ↓ | 0.30% (±0.11%) ↑ | 0.14% (±0.17%) | 0.04% (±0.05%) ↓ | 0.09% (±0.07%) | 0.15% (±0.04%) | 0.43% (±0.33%) | 0.0027 |
| GRP 3 | SCN | 1.22% (±0.93%) | 1.19% (±1.15%) | 7.52% (±5.94%) ↑ | 1.37% (±0.41%) | 0.06% (±0.02%) ↓ | 1.22% (±0.84%) | 2.36% (±1.22%) ↑ | 0.0115 |
| GAD65/67 | SCN | 7.26% (±2.94%) | 7.64% (±4.77%) | 7.64% (±2.43%) | 12.15% (±3.30%) | 9.37% (±4.48%) | 4.05% (±1.94%) | 14.35% (±7.89%) | 0.0572 |
| 5-HT 4 | SCN | 481.60 pi (±155.91 pi) ↓ | - | 1625.20 pi. (±155.91 pi) ↑ | 520.00 pi. (±96.75 pi.) | 959.40 pi. (±237.6 pi) ↑ | - | 292.75 pi. (±59.83 pi) ↓ | 0.0004 |
| SP 5 | SCN | 0.04% (±0.01%) ↓ | 0.73% (±0.14%) | 1.58% (±0.49%) ↑ | - | - | 0.63% (±0.25%) | - | 0.0039 |
| NPY 6 | SCN | 11.67% (±3.63%) | 8.52% (±7.79%) ↓ | 28.67% (±4.95%) ↑ | - | - | 7.79% (±3.49%) ↓ | - | 0.0087 |
| ME 5 | SCN | 0.38% (±0.15%) ↓ | - | 2.85% (±0.4%) ↑ | - | - | - | - | 0.0159 |
| Clock 7 | SCN | - | - | - | - | 10.47% (±1.02%) ↑ | - | 8.13% (±0.81%) ↓ | 0.0286 |
| Per 1 | SCN | - | - | - | - | 6.61% (±0.50%) | - | 4.83% (±0.71%) | 0.571 |
| Per 2 7 | SCN | - | - | - | - | 5.09% (±0.64%) ↑ | - | 3,92% (±0.15%) ↓ | 0.0286 |
| Cry 2 | SCN | - | - | - | - | 6.85% (±1.76%) | - | 6.61% (±0.58%) | >0.9999 |
| AVP 8 | PVN | 7.01% (±0.9%) ↑ | 8.35% (±1.35%) | 11.64% (±8.96%) | 6.22% (±2.10%) | 4.20% (±0.79%) | 6.06% (±2.52%) | 3.93% (±1.22%) ↓ | 0.0098 |
| OXY 9 | PVN | 9.97% (±1.75%) | 4.81% (±1.75%) | 13.04% (±2.93%) ↑ | 3.31% (±3.36%) ↓ | 2.96% (±1.27%) | 4.54% (±2.20%) | 2.04% (±0.36%) ↓ | 0.0022 |
| AR 5 | PVN | 0.53% (±0.39%) ↓ | - | 1.91% (±0.82%) ↑ | - | - | - | - | 0.0317 |
| OR 5 | PVN | 0.70% (±0.37%) ↓ | - | 2.17% (±0.78%) ↑ | - | - | - | - | 0.0159 |
| Clock | PVN | - | - | - | - | 6.13% (±0.24%) | - | 4.13% (±0.83%) | 0.0571 |
| Per 1 | PVN | - | - | - | - | 5.58% (±0.84%) | - | 5.77% (±0.59%) | 0.8571 |
| Per 2 | PVN | - | - | - | - | 2.77% (±0.82%) | - | 3.16% (±0.28%) * | >0.9999 |
| Cry 2 | PVN | - | - | - | - | 4.17% (±0.80%) | - | 4.59% (±1.22%) | 0.6286 |
| AVP 10 | SON | 22.56% (±3.26%) | 10.18% (±3.09%) ↓ | 66.60% (±4.53%) ↑ | 46.23% (±3.20%) ↑ | 47.19% (±3.46%) ↑ | 6.85% (±1.54%) ↓ | 36.03% (±7.75%) | <0.0001 |
| OXY 11 | SON | 17.30% (±4.05%) ↑ | 3.97% (±1.21%) ↓ | 15.39% (±1.89%) ↑ | - | - | 6.20% (±0.82%) | - | 0.0018 |
| Clock | SON | - | - | - | - | 6.17% (±0.72%) | - | 3.03% (±0.55%) | 0.0571 |
| Per 1 | SON | - | - | - | - | 5.37% (±0.64%) | - | 3.66% (±1.01%) | 0.1143 |
| Per 2 | SON | - | - | - | - | 2.86% (±0.58%) | - | 3.22% (±0.57%) | 0.4000 |
| Cry 2 | SON | - | - | - | - | 4.27% (±0.70%) | - | 4.24% (±0.51%) | 0.8571 |
| AR | MPOA | 0.55% (±0.65%) | - | 12.29% (±8.2%) | 9.62% (±1.50%) | 7.74% (n.e.) * | - | 12.38% (±4.89%) | 0.0493 |
| OR | MPOA | 12.65% (±8.67%) | - | 21.35% (±4.44%) | 22.21% (±3.30%) | 16.55% (n.e.) * | - | 23.32% (±13.75%) | 0.4540 |
| SOM 5 | MPVN | 2.45% (±0.24%) ↓ | - | 8.63% (±2.67%) ↑ | - | - | - | - | 0.0159 |
| TH 12 | IN | 9.15% (±1.52%) ↑ | 6.83% (±4.99%) | 7.84% (±0.41%) ↑ | 3.67% (±0.69%) | 4.96% (±0.45%) | 3.37% (±2.01%) ↓ | 2.30% (±0.25%) ↓ | 0.0005 |
| 5-HT 13 | DRN | 7.22% (±4.8%) | 2.16% (±0.96%) ↓ | 11.45% (±2.19%) ↑ | 11.33% (±4.06%) | 10.20% (±2.36%) | 3.56% (±0.89%) | 8.47% (±2.62%) | 0.0077 |
| DBH 14 | A7 | 5.04% (±0.82%) ↑ | - | 2.83% (±1.04%) | 2.35% (±0.65%) | 5.33% (±1.20%) ↑ | - | 2.17% (±0.55%) ↓ | 0.0029 |
| AVP 15 | Blood | 5.84 pg/mL (±0.86 pg/mL) | 18.00 pg/mL (±25.51 pg/mL) | 3.94 pg/mL (±1.56 pg/mL) | 3.00 pg/mL (±0.79 pg/mL) ↓ | 4.00 pg/mL (±0.96 pg/mL) | 111.0 pg/mL (±87.7 pg/mL) ↑ | 3.34 pg/mL (±2.14 pg/mL) ↓ | 0.0153 |
| Testo-sterone 16 | Blood | 9.42 ng/mL (±1.73 ng/mL) ↑ | 4.12 ng/mL (±1.20 ng/mL) | 3.26 ng/mL (±0.72 ng/mL) | 8.25 ng/mL (±3.61 ng/mL) ↑ | 4.38 ng/mL (±3.72 ng/mL) | 1.63 ng/mL (±0.46 ng/mL) ↓ | 5.40 ng/mL (±2.62 ng/mL) | 0.0017 |
| Experimental Group | Sacrifice Time (ZT) | Relative Increase Compared to at Least One Other Group | Relative Decrease Compared to at Least One Other Group |
|---|---|---|---|
| Intermediate-term sleep deprivation (20 h) | 05.00 |
|
|
| Short-term sleep deprivation (6 h) | 05.00 |
|
|
| Short-term sleep deprivation (1 h) | 01.00 |
| |
| Sleep | 05.00 |
|
|
| Exposure to pheromone (1 h) | 01.00 |
|
|
| Exposure to pheromone and females (1 h) | 01.00 |
|
|
| Immobilization stress (1 h) | 01.00 |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knöchel, C.; Frickmann, H.; Nürnberger, F. Effects of Sleep Deprivation by Olfactorily Induced Sexual Arousal Compared to Immobilization Stress and Manual Sleep Deprivation on Neuromessengers and Time Keeping Genes in the Suprachiasmatic Nuclei and Other Cerebral Entities of Syrian Hamsters—An Immunohistochemical Study. Int. J. Environ. Res. Public Health 2021, 18, 9169. https://doi.org/10.3390/ijerph18179169
Knöchel C, Frickmann H, Nürnberger F. Effects of Sleep Deprivation by Olfactorily Induced Sexual Arousal Compared to Immobilization Stress and Manual Sleep Deprivation on Neuromessengers and Time Keeping Genes in the Suprachiasmatic Nuclei and Other Cerebral Entities of Syrian Hamsters—An Immunohistochemical Study. International Journal of Environmental Research and Public Health. 2021; 18(17):9169. https://doi.org/10.3390/ijerph18179169
Chicago/Turabian StyleKnöchel, Christian, Hagen Frickmann, and Frank Nürnberger. 2021. "Effects of Sleep Deprivation by Olfactorily Induced Sexual Arousal Compared to Immobilization Stress and Manual Sleep Deprivation on Neuromessengers and Time Keeping Genes in the Suprachiasmatic Nuclei and Other Cerebral Entities of Syrian Hamsters—An Immunohistochemical Study" International Journal of Environmental Research and Public Health 18, no. 17: 9169. https://doi.org/10.3390/ijerph18179169
APA StyleKnöchel, C., Frickmann, H., & Nürnberger, F. (2021). Effects of Sleep Deprivation by Olfactorily Induced Sexual Arousal Compared to Immobilization Stress and Manual Sleep Deprivation on Neuromessengers and Time Keeping Genes in the Suprachiasmatic Nuclei and Other Cerebral Entities of Syrian Hamsters—An Immunohistochemical Study. International Journal of Environmental Research and Public Health, 18(17), 9169. https://doi.org/10.3390/ijerph18179169