
Interaction of Smoking and Lead Exposure among Carriers of Genetic Variants Associated with a Higher Level of Oxidative Stress Indicators




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Health Examination
2.2. Genotyping Methods for Glutathione Peroxidase-1 (GPx-1) Gene (rs1050450) and Cytochrome b Light Chain (CYBA) Gene (rs4673)
2.3. Measurement of TBARS and Oxidized LDL
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kianoush, S.; Balali-Mood, M.; Mousavi, S.R.; Shakeri, M.T.; Dadpour, B.; Moradi, V.; Sadeghi, M. Clinical, toxicological, biochemical, and hematologic parameters in lead exposed workers of a car battery industry. Iran. J. Med. Sci. 2013, 38, 30–37. [Google Scholar]
- Rocha, A.; Trujillo, K.A. Neurotoxicity of low-level lead exposure: History, mechanisms of action, and behavioral effects in humans and preclinical models. Neurotoxicology 2019, 73, 58–80. [Google Scholar] [CrossRef]
- Harari, F.; Sallsten, G.; Christensson, A.; Petkovic, M.; Hedblad, B.; Forsgard, N.; Melander, O.; Nilsson, P.M.; Borné, Y.; Engström, G.; et al. Blood Lead Levels and Decreased Kidney Function in a Population-Based Cohort. Am. J. Kidney Dis. 2018, 72, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Obeng-Gyasi, E.; Obeng-Gyasi, B. Chronic Stress and Cardiovascular Disease among Individuals Exposed to Lead: A Pilot Study. Diseases 2020, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Gurer, H.; Ercal, N. Can antioxidants be beneficial in the treatment of lead poisoning? Free Radic. Biol. Med. 2000, 29, 927–945. [Google Scholar] [CrossRef]
- Farmand, F.; Ehdaie, A.; Roberts, C.K.; Sindhu, R.K. Lead-induced dysregulation of superoxide dismutases, catalase, glutathione peroxidase, and guanylate cyclase. Environ. Res. 2005, 98, 33–39. [Google Scholar] [CrossRef]
- Pech-Amsellem, M.A.; Myara, I.; Storogenko, M.; Demuth, K.; Proust, A.; Moatti, N. Enhanced modifications of low-density lipoproteins (LDL) by endothelial cells from smokers: A possible mechanism of smoking-related atherosclerosis. Cardiovasc. Res. 1996, 31, 975–983. [Google Scholar] [CrossRef]
- Flora, S.J. Arsenic-induced oxidative stress and its reversibility. Free Radic. Biol. Med. 2011, 51, 257–281. [Google Scholar] [CrossRef]
- Ushio-Fukai, M.; Zafari, A.M.; Fukui, T.; Ishizaka, N.; Griendling, K.K. p22phox is a critical component of the superoxide-generating NADH/NADPH oxidase system and regulates angiotensin II-induced hypertrophy in vascular smooth muscle cells. J. Biol. Chem. 1996, 271, 23317–23321. [Google Scholar] [CrossRef] [Green Version]
- Inoue, N.; Kawashima, S.; Kanazawa, K.; Yamada, S.; Akita, H.; Yokoyama, M. Polymorphism of the NADH/NADPH oxidase p22 phox gene in patients with coronary artery disease. Circulation 1998, 97, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Fortuno, A.; San Jose, G.; Moreno, M.U.; Beloqui, O.; Diez, J.; Zalba, G. Phagocytic NADPH oxidase overactivity underlies oxidative stress in metabolic syndrome. Diabetes 2006, 55, 209–215. [Google Scholar] [CrossRef]
- Fortuno, A.; Olivan, S.; Beloqui, O.; San Jose, G.; Moreno, M.U.; Diez, J.; Zalba, G. Association of increased phagocytic NADPH oxidase-dependent superoxide production with diminished nitric oxide generation in essential hypertension. J. Hypertens. 2004, 22, 2169–2175. [Google Scholar] [CrossRef]
- Guzik, T.J.; Mussa, S.; Gastaldi, D.; Sadowski, J.; Ratnatunga, C.; Pillai, R.; Channon, K.M. Mechanisms of increased vascular superoxide production in human diabetes mellitus: Role of NAD(P)H oxidase and endothelial nitric oxide synthase. Circulation 2002, 105, 1656–1662. [Google Scholar] [CrossRef] [Green Version]
- Zalba, G.; Fortuño, A.; Díez, J. Oxidative stress and atherosclerosis in early chronic kidney disease. Nephrol. Dial. Transplant. 2006, 21, 2686–2690. [Google Scholar] [CrossRef] [Green Version]
- Sorescu, D.; Weiss, D.; Lassegue, B.; Clempus, R.E.; Szocs, K.; Sorescu, G.P.; Valppu, L.; Quinn, M.T.; Lambeth, J.D.; Vega, J.D.; et al. Superoxide production and expression of nox family proteins in human atherosclerosis. Circulation 2002, 105, 1429–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.A.; Drummond, G.R.; Sobey, C.G. Novel isoforms of NADPH-oxidase in cerebral vascular control. Pharmacol. Ther. 2006, 111, 928–948. [Google Scholar] [CrossRef] [PubMed]
- Dinauer, M.C.; Pierce, E.A.; Bruns, G.A.; Curnutte, J.T.; Orkin, S.H. Human neutrophil cytochrome b light chain (p22-phox). Gene structure, chromosomal location, and mutations in cytochrome-negative autosomal recessive chronic granulomatous disease. J. Clin. Investig. 1990, 86, 1729–1737. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga-Irie, S.; Maruyama, T.; Yamamoto, Y.; Motohashi, Y.; Hirose, H.; Shimada, A.; Murata, M.; Saruta, T. Relation between development of nephropathy and the p22phox C242T and receptor for advanced glycation end product G1704T gene polymorphisms in type 2 diabetic patients. Diabetes Care 2004, 27, 303–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Lou, Y.; Jin, W.; Liu, Y.; Lu, L.; Chen, Q.; Xie, Y.; Lu, G. Relationship of the p22phox (CYBA) gene polymorphism C242T with risk of coronary artery disease: A meta-analysis. PLoS ONE 2013, 8, e70885. [Google Scholar] [CrossRef]
- Men, T.; Zhang, X.; Yang, J.; Shen, B.; Li, X.; Chen, D.; Wang, J. The rs1050450 C > T polymorphism of GPX1 is associated with the risk of bladder but not prostate cancer: Evidence from a meta-analysis. Tumor Biol. 2014, 35, 269–275. [Google Scholar] [CrossRef]
- Forgione, M.A.; Weiss, N.; Heydrick, S.; Cap, A.; Klings, E.S.; Bierl, C.; Eberhardt, R.T.; Farber, H.W.; Loscalzo, J. Cellular glutathione peroxidase deficiency and endothelial dysfunction. Am. J. Physiol. Circ. Physiol. 2002, 282, H1255–H1261. [Google Scholar] [CrossRef]
- Hauser, F.; Rossmann, H.; Laubert-Reh, D.; Wild, P.S.; Zeller, T.; Muller, C.; Neuwirth, S.; Blankenberg, S.; Lackner, K.J. Inflammatory bowel disease (IBD) locus 12: Is glutathione peroxidase-1 (GPX1) the relevant gene? Genes Immun. 2015, 16, 571–575. [Google Scholar] [CrossRef]
- Arthur, J.R. The glutathione peroxidases. Cell. Mol. Life Sci. 2001, 57, 1825–1835. [Google Scholar] [CrossRef]
- Hamanishi, T.; Furuta, H.; Kato, H.; Doi, A.; Tamai, M.; Shimomura, H.; Sakagashira, S.; Nishi, M.; Sasaki, H.; Sanke, T.; et al. Functional variants in the glutathione peroxidase-1 (GPx-1) gene are associated with increased intima-media thickness of carotid arteries and risk of macrovascular diseases in Japanese type 2 diabetic patients. Diabetes 2004, 53, 2455–2460. [Google Scholar] [CrossRef] [Green Version]
- Forgione, M.A.; Cap, A.; Liao, R.; Moldovan, N.I.; Eberhardt, R.T.; Lim, C.C.; Jones, J.; Goldschmidt-Clermont, P.J.; Loscalzo, J. Heterozygous cellular glutathione peroxidase deficiency in the mouse: Abnormalities in vascular and cardiac function and structure. Circulation 2002, 106, 1154–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Z.; Tian, C.; Zhang, X. GPX1 gene Pro200Leu polymorphism, erythrocyte GPX activity, and cancer risk. Mol. Biol. Rep. 2013, 40, 1801–1812. [Google Scholar] [CrossRef]
- Tang, T.S.; Prior, S.L.; Li, K.W.; Ireland, H.A.; Bain, S.C.; Hurel, S.J.; Cooper, J.A.; Humphries, S.E.; Stephens, J.W. Association between the rs1050450 glutathione peroxidase-1 (C > T) gene variant and peripheral neuropathy in two independent samples of subjects with diabetes mellitus. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.P.; Wang, L.S.; Yang, L.; Gu, H.J.; Sun, Q.M.; Cong, R.H.; Zhou, B.; Zhu, H.J.; Wang, B. Genetic variant in glutathione peroxidase 1 gene is associated with an increased risk of coronary artery disease in a Chinese population. Clin. Chim. Acta 2008, 395, 89–93. [Google Scholar] [CrossRef]
- Kuzuya, M.; Ando, F.; Iguchi, A.; Shimokata, H. Glutathione peroxidase 1 Pro198Leu variant contributes to the metabolic syndrome in men in a large Japanese cohort. Am. J. Clin. Nutr. 2008, 87, 1939–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, E.; Hughes, G.R.; Khamashta, M.A. Oxidation of LDL and its clinical implication. Autoimmun. Rev. 2008, 7, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Girona, J.; Manzanares, J.M.; Marimon, F.; Cabre, A.; Heras, M.; Guardiola, M.; Ribalta, J.; Masana, L. Oxidized to non-oxidized lipoprotein ratios are associated with arteriosclerosis and the metabolic syndrome in diabetic patients. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, S.; Raghavamenon, A.; Garelnabi, M.O.; Santanam, N. Oxidized low-density lipoprotein. Methods Mol. Biol. 2010, 610, 403–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, D.; Parthasarathy, S.; Carew, T.E.; Khoo, J.C.; Witztum, J.L. Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 1989, 320, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Halliwell, B.; Chirico, S. Lipid peroxidation: Its mechanism, measurement, and significance. Am. J. Clin. Nutr. 1993, 57, 715S–725S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, E.; Karimi Galougahi, K.; Liu, C.C.; Bhindi, R.; Figtree, G.A. Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, M.; Browne, R.; Ram, M.; Muti, P.; Freudenheim, J.; Carosella, A.M.; Armstrong, D. Correlates of markers of oxidative status in the general population. Am. J. Epidemiol. 2001, 154, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Schisterman, E.F.; Faraggi, D.; Browne, R.; Freudenheim, J.; Dorn, J.; Muti, P.; Armstrong, D.; Reiser, B.; Trevisan, M. TBARS and cardiovascular disease in a population-based sample. J. Cardiovasc. Risk 2001, 8, 219–225. [Google Scholar] [CrossRef]
- Schisterman, E.F.; Faraggi, D.; Browne, R.; Freudenheim, J.; Dorn, J.; Muti, P.; Armstrong, D.; Reiser, B.; Trevisan, M. Minimal and best linear combination of oxidative stress and antioxidant biomarkers to discriminate cardiovascular disease. Nutr. Metab. Cardiovasc. Dis. 2002, 12, 259–266. [Google Scholar]
- Seet, R.C.; Lee, C.Y.; Loke, W.M.; Huang, S.H.; Huang, H.; Looi, W.F.; Chew, E.S.; Quek, A.M.; Lim, E.C.; Halliwell, B. Biomarkers of oxidative damage in cigarette smokers: Which biomarkers might reflect acute versus chronic oxidative stress? Free Radic. Biol. Med. 2011, 50, 1787–1793. [Google Scholar] [CrossRef] [PubMed]
- Morrow, J.D.; Frei, B.; Longmire, A.W.; Gaziano, J.M.; Lynch, S.M.; Shyr, Y.; Strauss, W.E.; Oates, J.A.; Roberts, L.J., 2nd. Increase in circulating products of lipid peroxidation (F2-isoprostanes) in smokers. Smoking as a cause of oxidative damage. N. Engl. J. Med. 1995, 332, 1198–1203. [Google Scholar] [CrossRef]
- Lykkesfeldt, J. Malondialdehyde as biomarker of oxidative damage to lipids caused by smoking. Clin. Chim. Acta 2007, 380, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Ellman, G.; Lysko, H. A precise method for the determination of whole blood and plasma sulfhydryl groups. Anal. Biochem. 1979, 93, 98–102. [Google Scholar] [CrossRef]
- Aviram, M.; Fuhrman, B. LDL oxidation by arterial wall macrophages depends on the oxidative status in the lipoprotein and in the cells: Role of prooxidants vs. antioxidants. Mol. Cell. Biochem. 1998, 188, 149–159. [Google Scholar] [CrossRef]
- Pan, W.H.; Flegal, K.M.; Chang, H.Y.; Yeh, W.T.; Yeh, C.J.; Lee, W.C. Body mass index and obesity-related metabolic disorders in Taiwanese and US whites and blacks: Implications for definitions of overweight and obesity for Asians. Am. J. Clin. Nutr. 2004, 79, 31–39. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity: Preventing and managing the global epidemic. Report of a WHO consultation. World Health Organ. Tech. Rep. Ser. 2000, 894, 1–253. [Google Scholar]
- National Institutes of Health. Clinical Guidelines on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults—The Evidence Report. Obes. Res. 1998, 6, 51s–209s. [Google Scholar]
- Deurenberg, P.; Yap, M.; van Staveren, W.A. Body mass index and percent body fat: A meta analysis among different ethnic groups. Int. J. Obes. 1998, 22, 1164–1171. [Google Scholar] [CrossRef] [Green Version]
- Chu, N.F. Prevalence of obesity in Taiwan. Obes. Rev. 2005, 6, 271–274. [Google Scholar] [CrossRef]
- Stephens, J.W.; Gable, D.R.; Hurel, S.J.; Miller, G.J.; Cooper, J.A.; Humphries, S.E. Increased plasma markers of oxidative stress are associated with coronary heart disease in males with diabetes mellitus and with 10-year risk in a prospective sample of males. Clin. Chem. 2006, 52, 446–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alya, A.; Ines, D.B.; Montassar, L.; Najoua, G.; Saloua, E.F. Oxidative stress, biochemical alterations, and hyperlipidemia in female rats induced by lead chronic toxicity during puberty and post puberty periods. Iran. J. Basic Med Sci. 2015, 18, 1034–1043. [Google Scholar] [PubMed]
- Agrawal, S.; Flora, G.; Bhatnagar, P.; Flora, S.J. Comparative oxidative stress, metallothionein induction and organ toxicity following chronic exposure to arsenic, lead and mercury in rats. Cell. Mol. Biol. 2014, 60, 13–21. [Google Scholar] [PubMed]
- Miller, E.R., 3rd; Appel, L.J.; Jiang, L.; Risby, T.H. Association between cigarette smoking and lipid peroxidation in a controlled feeding study. Circulation 1997, 96, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Panagiotakos, D.B.; Pitsavos, C.; Chrysohoou, C.; Skoumas, J.; Masoura, C.; Toutouzas, P.; Stefanadis, C. Effect of exposure to secondhand smoke on markers of inflammation: The ATTICA study. Am. J. Med. 2004, 116, 145–150. [Google Scholar] [CrossRef]
- Forsberg, L.; de Faire, U.; Morgenstern, R. Low yield of polymorphisms from EST blast searching: Analysis of genes related to oxidative stress and verification of the P197L polymorphism in GPX1. Hum. Mutat. 1999, 13, 294–300. [Google Scholar] [CrossRef]
- Ravn-Haren, G.; Olsen, A.; Tjonneland, A.; Dragsted, L.O.; Nexo, B.A.; Wallin, H.; Overvad, K.; Raaschou-Nielsen, O.; Vogel, U. Associations between GPX1 Pro198Leu polymorphism, erythrocyte GPX activity, alcohol consumption and breast cancer risk in a prospective cohort study. Carcinogenesis 2006, 27, 820–825. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.J.; Diamond, A.M. Role of glutathione peroxidase 1 in breast cancer: Loss of heterozygosity and allelic differences in the response to selenium. Cancer Res. 2003, 63, 3347–3351. [Google Scholar]
- Ichimura, Y.; Habuchi, T.; Tsuchiya, N.; Wang, L.; Oyama, C.; Sato, K.; Nishiyama, H.; Ogawa, O.; Kato, T. Increased risk of bladder cancer associated with a glutathione peroxidase 1 codon 198 variant. J. Urol. 2004, 172, 728–732. [Google Scholar] [CrossRef]
- Zarbock, R.; Hendig, D.; Szliska, C.; Kleesiek, K.; Gotting, C. Pseudoxanthoma elasticum: Genetic variations in antioxidant genes are risk factors for early disease onset. Clin. Chem. 2007, 53, 1734–1740. [Google Scholar] [CrossRef] [Green Version]
- Raaschou-Nielsen, O.; Sorensen, M.; Hansen, R.D.; Frederiksen, K.; Tjonneland, A.; Overvad, K.; Vogel, U. GPX1 Pro198Leu polymorphism, interactions with smoking and alcohol consumption, and risk for lung cancer. Cancer Lett. 2007, 247, 293–300. [Google Scholar] [CrossRef]
- Mostowska, A.; Hozyasz, K.K.; Lianeri, M.; Piwowar, W.; Jagodzinski, P.P. Polymorphic variants of genes encoding main antioxidant enzymes and the risk of CL/P-affected pregnancies. Clin. Biochem. 2007, 40, 416–419. [Google Scholar] [CrossRef]
- Choi, J.Y.; Neuhouser, M.L.; Barnett, M.; Hudson, M.; Kristal, A.R.; Thornquist, M.; King, I.B.; Goodman, G.E.; Ambrosone, C.B. Polymorphisms in oxidative stress-related genes are not associated with prostate cancer risk in heavy smokers. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1115–1120. [Google Scholar] [CrossRef] [Green Version]
- Lightfoot, T.J.; Skibola, C.F.; Smith, A.G.; Forrest, M.S.; Adamson, P.J.; Morgan, G.J.; Bracci, P.M.; Roman, E.; Smith, M.T.; Holly, E.A. Polymorphisms in the oxidative stress genes, superoxide dismutase, glutathione peroxidase and catalase and risk of non-Hodgkin’s lymphoma. Haematol. 2006, 91, 1222–1227. [Google Scholar]
- Knight, J.A.; Onay, U.V.; Wells, S.; Li, H.; Shi, E.J.; Andrulis, I.L.; Ozcelik, H. Genetic variants of GPX1 and SOD2 and breast cancer risk at the Ontario site of the Breast Cancer Family Registry. Cancer Epidemiol. Biomark. Prev. 2004, 13, 146–149. [Google Scholar] [CrossRef] [Green Version]
- Ratnasinghe, D.; Tangrea, J.A.; Andersen, M.R.; Barrett, M.J.; Virtamo, J.; Taylor, P.R.; Albanes, D. Glutathione peroxidase codon 198 polymorphism variant increases lung cancer risk. Cancer Res. 2000, 60, 6381–6383. [Google Scholar]
- Forsberg, L.; de Faire, U.; Marklund, S.L.; Andersson, P.M.; Stegmayr, B.; Morgenstern, R. Phenotype determination of a common Pro-Leu polymorphism in human glutathione peroxidase 1. Blood Cells Mol. Dis. 2000, 26, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Sorescu, D.; Ushio-Fukai, M. NAD(P)H oxidase: Role in cardiovascular biology and disease. Circ. Res. 2000, 86, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, A.S.; FitzGerald, G.A. Twenty-first century phox: Not yet ready for widespread screening. Circulation 2001, 103, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Matsunaga, S.; Nagata, A.; Maruyama, T. NAD(P)H oxidase p22phox Gene C242T polymorphism and lipoprotein oxidation. Clin. Chim. Acta 2003, 335, 101–107. [Google Scholar] [CrossRef]
- Taiwan Tobacco Control Annual Report; Health Promotion Administration, Ministry of Health and Welfare: Taipei, Taiwan, 2019. Available online: https://www.hpa.gov.tw/EngPages/Detail.aspx?nodeid=1069&pid=12873 (accessed on 8 May 2021).
- Tsai, Y.W.; Chang, L.C.; Sung, H.Y.; Hu, T.W.; Chiou, S.T. The impact of smoke-free legislation on reducing exposure to secondhand smoke: Differences across gender and socioeconomic groups. Tob. Control. 2015, 24, 62–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannino, D.M.; Homa, D.M.; Matte, T.; Hernandez-Avila, M. Active and passive smoking and blood lead levels in U.S. adults: Data from the Third National Health and Nutrition Examination Survey. Nicotine Tob. Res. 2005, 7, 557–564. [Google Scholar] [CrossRef] [PubMed]
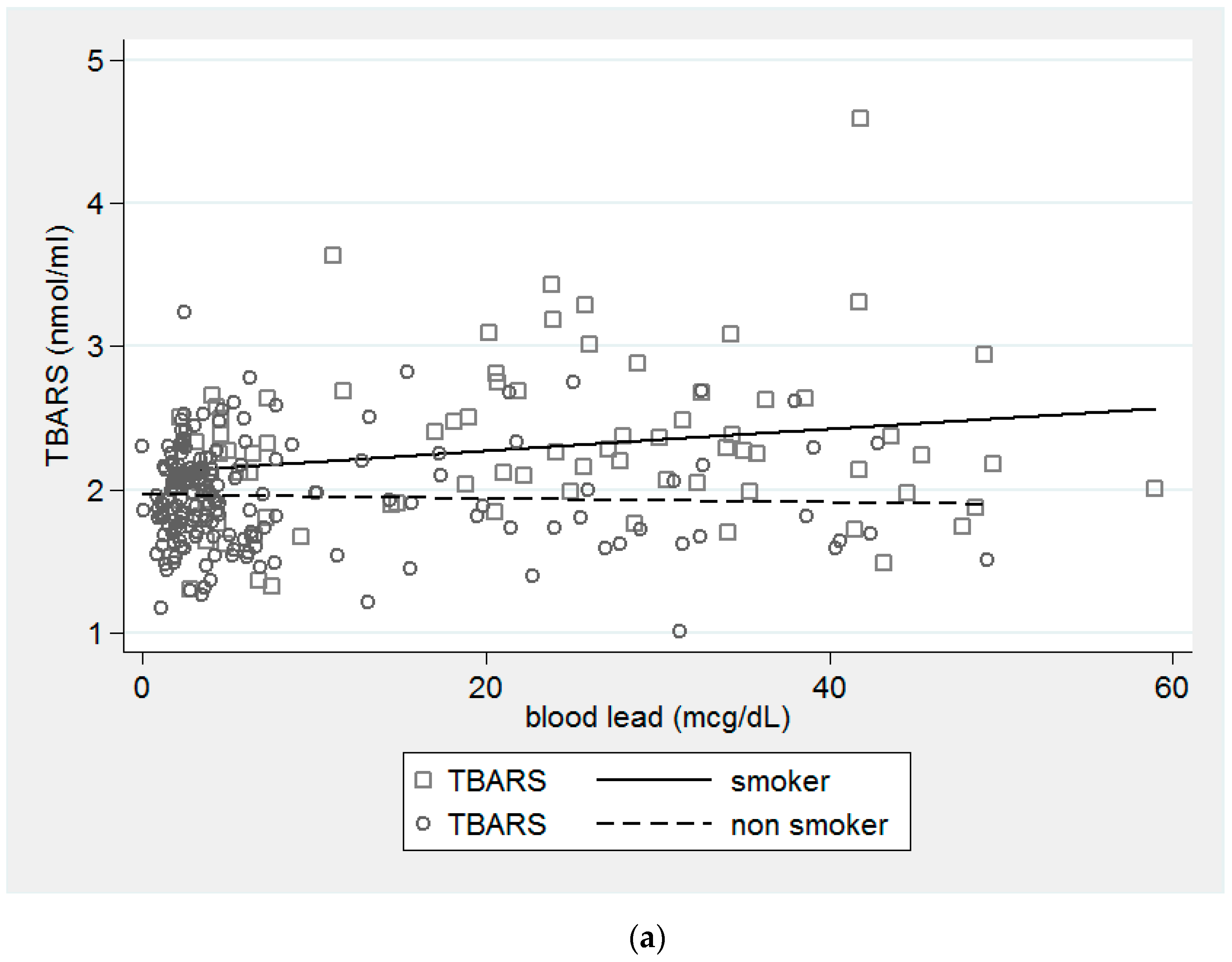
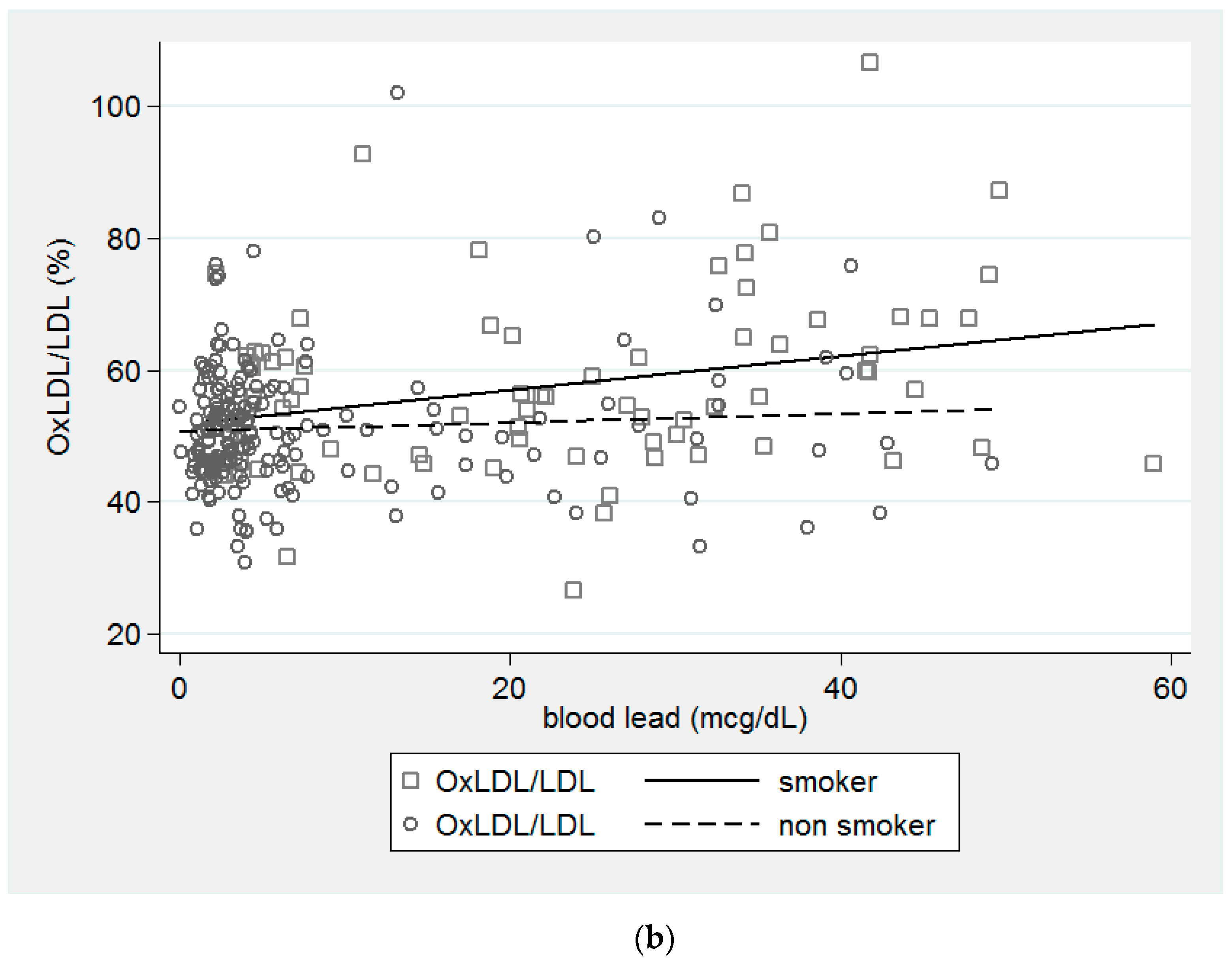

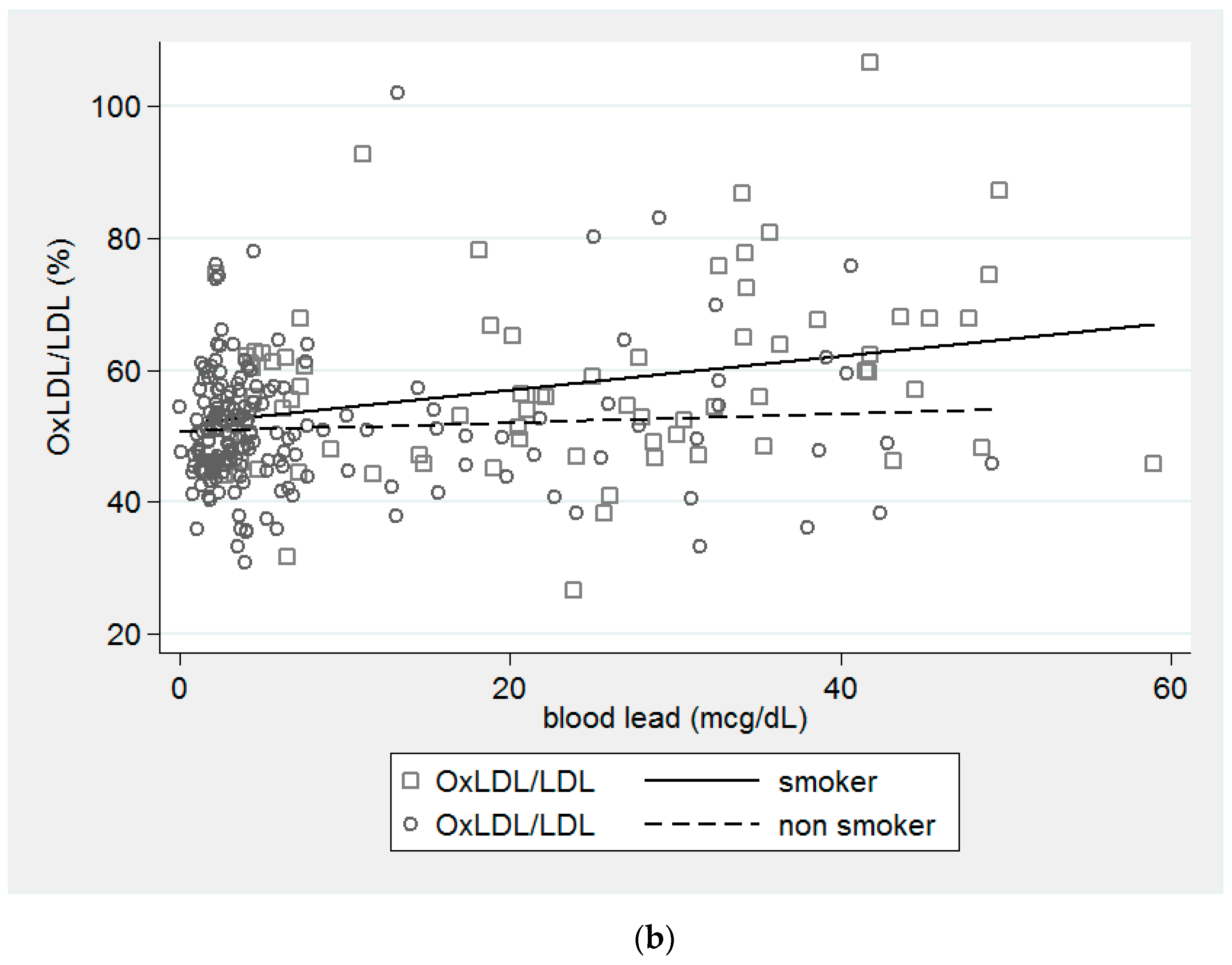
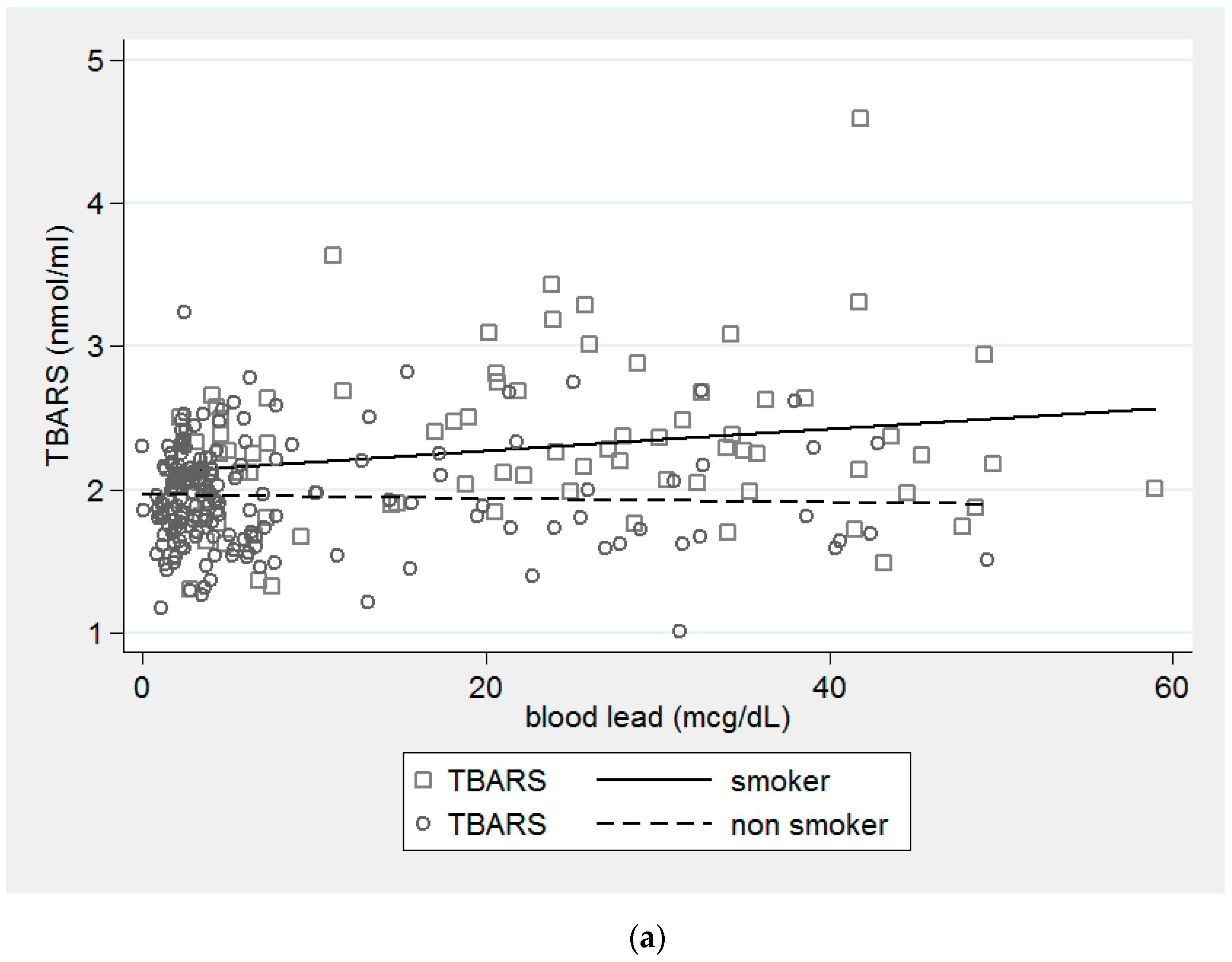{kind=link}
{kind=link}
| Variables | Smokers (n = 81) | Nonsmokers (n = 186) | p Value |
|---|---|---|---|
| Blood Pb (μg/dL) | 22.11 ± 15.19 | 7.97 ± 10.42 | <0.001 ** |
| Age (years) | 43.7 ± 9.7 | 41.1 ± 10.4 | 0.058 |
| Sex (male) | 80 (98.8%) | 113 (60.8%) | <0.001 ** |
| Alcohol (>3 times/week) | 28 (42.4%) | 13 (7.3%) | <0.001 ** |
| BMI (kg/m2) | 25.69 ± 3.52 | 23.88 ± 3.71 | <0.001 ** |
| BMI < 18.5 | 3 (3.7%) | 7 (3.8%) | 0.981 |
| BMI ≥ 24 | 56 (69.1%) | 83 (44.6%) | 0.001 ** |
| Creatinine (mg/dL) | 1.22 ± 0.14 | 1.15 ± 0.63 | 0.357 |
| ALT (U/L) | 22.85 ± 14.42 | 18.19 ± 16.14 | 0.027 * |
| AC sugar (mg/dL) | 102.92 ± 33.41 | 99.54 ± 31.08 | 0.433 |
| TBARS (nmol/mL) | 2.289 ± 0.559 | 1.953 ± 0.351 | <0.001 ** |
| OxLDL (mg/dL) | 59.857 ± 14.567 | 53.76 ± 11.252 | 0.001 ** |
| LDL (mg/dL) | 104.621 ± 18.304 | 104.457 ± 11.625 | 0.942 |
| CYBA gene (rs4673) | 0.625 | ||
| CC | 60 (74.1%) | 145 (78.0%) | |
| CT | 19 (23.5%) | 39 (21.0%) | |
| TT | 2 (2.5%) | 2 (1.1%) | |
| GPX1 gene (rs1050450) | 0.157 | ||
| CC | 75 (92.6%) | 161 (86.6%) | |
| CT | 6 (7.4%) | 25 (13.4%) |
| Variables | CYBA rs4673 | GPx-1 rs1050450 | |||||
|---|---|---|---|---|---|---|---|
| Mean ± SD or n (%) | CC (n = 205) | CT (n = 58) | TT (n = 4) | p Value | CC (n = 236) | CT (n = 31) | p Value |
| Blood Pb (μg/dL) | 11.82 ± 13.74 | 13.09 ± 12.75 | 23.04 ± 22.31 | 0.234 | 12.87 ± 14.03 | 7.58 ± 9.61 | 0.009 * |
| Age (years) | 42.1 ± 10.1 | 41.6 ± 11.2 | 35.6 ± 4.4 | 0.477 | 42.0 ± 10.3 | 40.9 ± 9.8 | 0.556 |
| Sex (male) | 148 (72.2%) | 41 (70.7%) | 4 (100%) | 0.448 | 171 (72.5%) | 22 (71.0%) | 0.862 |
| Current smoking | 60 (29.3%) | 19 (32.8%) | 2 (50%) | 0.605 | 75 (31.8%) | 6 (19.4%) | 0.157 |
| Alcohol (>3 times/week) | 32 (16.9%) | 9 (17.3%) | 0 (0%) | 0.734 | 38 (17.6%) | 3 (10.7%) | 0.36 |
| BMI (kg/m2) | 24.43 ± 3.72 | 24.48 ± 3.91 | 24.48 ± 2.55 | 0.875 | 24.51 ± 3.81 | 23.83 ± 3.14 | 0.343 |
| BMI < 18.5 | 8 (3.9%) | 2 (3.4%) | 0 (0%) | 0.912 | 9 (3.8%) | 1 (3.2%) | 0.871 |
| BMI ≥ 24 | 106 (51.7%) | 32 (55.1%) | 1 (25%) | 0.494 | 123 (52.1%) | 16 (51.6%) | 0.958 |
| Creatinine (mg/dL) | 1.18 ± 0.60 | 1.15 ± 0.18 | 1.17 ± 0.53 | 0.954 | 1.18 ± 0.59 | 1.10 ± 0.17 | 0.438 |
| ALT (U/L) | 19.7 ± 15.1 | 20.1 ± 18.3 | 11.0 ± 6.3 | 0.624 | 19.9 ± 16.2 | 17.8 ± 11.7 | 0.501 |
| AC sugar (mg/dL) | 98.9 ± 25.1 | 107.3 ± 49.0 | 90.2 ± 5.4 | 0.192 | 101.3 ± 33.3 | 94.8 ± 14.0 | 0.308 |
| TBARS(nmol/mL) | 2.06 ± 0.46 | 2.06 ± 0.41 | 1.72 ± 0.27 | 0.332 | 2.053 ± 0.464 | 2.078 ± 0.344 | 0.776 |
| OxLDL (mg/dL) | 55.98 ± 13.02 | 55.04 ± 11.26 | 45.87 ± 10.74 | 0.265 | 55.64 ± 12.92 | 55.50 ± 10.53 | 0.955 |
| LDL (mg/dL) | 105.31 ± 13.53 | 102.54 ± 14.89 | 93.07 ± 16.5 | 0.105 | 104.00 ± 14.10 | 108.39 ± 12.23 | 0.105 |
| Models | TBARS | OxLDL/LDL |
|---|---|---|
| β (SE) | β (SE) | |
| Model 1 | ||
| Blood Pb | −0.001 (0.003) | 0.078 (0.08) |
| smoking | 0.115 (0.101) | −0.704 (2.447) |
| Blood Pb x smoke | 0.009 (0.005) * | 0.223 (0.11) * |
| Model 2 | ||
| Blood Pb | 0.004 (0.003) | 0.193 (0.062) * |
| rs4673 | 0.003 (0.092) | 0.336 (2.219) |
| Blood Pb x rs4673 | −0.003 (0.005) | 0 (0.114) |
| Model 3 | ||
| Blood Pb | 0.004 (0.002) | 0.212 (0.058) ** |
| rs1050450 | 0.137 (0.114) | 2.054 (2.746) |
| Blood Pb x rs1050450 | −0.007 (0.009) | −0.317 (0.205) |
| Model 4 | ||
| Blood Pb | 0.004 (0.002) | 0.191 (0.057) ** |
| rs4673 | −0.055 (0.069) | 0.115 (1.67) |
| rs1050450 | 0.051 (0.104) | −1.278 (2.514) |
| rs4673 x rs1050450 | 0.1 (0.193) | 2.037 (4.66) |
| Model 5 | ||
| Blood Pb | 0.004 (0.002) | 0.192 (0.057) ** |
| rs4673 | −0.038 (0.067) | 0.433 (1.619) |
| rs1050450 | 0.09 (0.098) | −0.559 (2.352) |
| Blood Pb x rs4673 x rs1050450 | −0.003 (0.012) | −0.038 (0.285) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, K.-J.; Chen, T.-H.; Yang, C.-C.; Chuang, Y.-C.; Chuang, H.-Y. Interaction of Smoking and Lead Exposure among Carriers of Genetic Variants Associated with a Higher Level of Oxidative Stress Indicators. Int. J. Environ. Res. Public Health 2021, 18, 8325. https://doi.org/10.3390/ijerph18168325
Ho K-J, Chen T-H, Yang C-C, Chuang Y-C, Chuang H-Y. Interaction of Smoking and Lead Exposure among Carriers of Genetic Variants Associated with a Higher Level of Oxidative Stress Indicators. International Journal of Environmental Research and Public Health. 2021; 18(16):8325. https://doi.org/10.3390/ijerph18168325
Chicago/Turabian StyleHo, Kuo-Jung, Tzu-Hua Chen, Chen-Cheng Yang, Yao-Chung Chuang, and Hung-Yi Chuang. 2021. "Interaction of Smoking and Lead Exposure among Carriers of Genetic Variants Associated with a Higher Level of Oxidative Stress Indicators" International Journal of Environmental Research and Public Health 18, no. 16: 8325. https://doi.org/10.3390/ijerph18168325
APA StyleHo, K.-J., Chen, T.-H., Yang, C.-C., Chuang, Y.-C., & Chuang, H.-Y. (2021). Interaction of Smoking and Lead Exposure among Carriers of Genetic Variants Associated with a Higher Level of Oxidative Stress Indicators. International Journal of Environmental Research and Public Health, 18(16), 8325. https://doi.org/10.3390/ijerph18168325