
Comparative Analysis of Metabolite Profiling of Momordica charantia Leaf and the Anti-Obesity Effect through Regulating Lipid Metabolism

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Extraction and Preparation of Sample
2.2. Antioxidant Activity Analysis
2.3. UPLC-Q-TOF MS Investigation of the Fragmentation Patterns
2.4. Cell Culture, Differentiation, and Treatments
2.5. Determination of Lipid Accumulation by Oil-Red O Staining
2.6. Experimental Animals, Diet, and Treatments
2.7. Biochemical Analysis
2.8. Histological Analysis
2.9. MRNA Expression Analysis
2.10. Protein Expression Analysis
2.11. Statistical Analysis
3. Results
3.1. Effects of M. charantia on Antioxidant Activity and Lipid Accumulation
3.2. Identification of the Bioactive Components in M. charantia
3.3. Effects of MCLW on Body Composition and Blood Biochemical Parameters
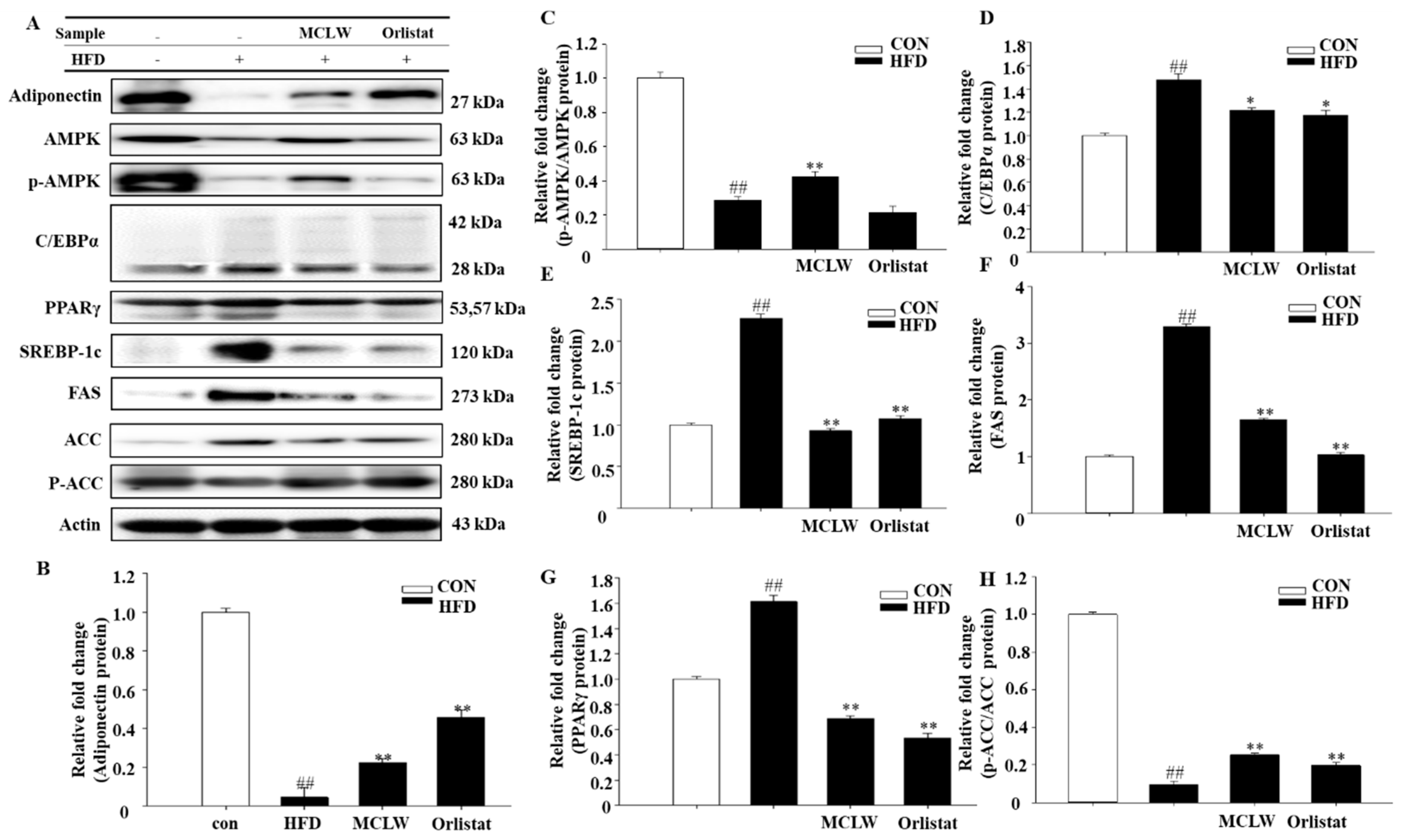
3.4. Effects of MCLW on Fat Metabolic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mokdad, A.H. Burden of Obesity in the Eastern Mediterranean Region: Findings from the Global Burden of Disease 2015 Study. Int. J. Environ. Res. Public Health 2018, 63, 165–176. [Google Scholar]
- Bhadoria, A.S.; Sahoo, K.; Sahoo, B.; Choudhury, A.K.; Sofi, N.Y.; Kumar, C.A. Childhood Obesity: Causes and Consequences. J. Fam. Med. Prim. Care 2015, 4, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Serrano, A.; Arola-Arnal, A.; Suárez-García, S.; Bravo, F.I.; Suárez, M.; Arola, L.; Bladé, C. Grape Seed Proanthocya-nidin Supplementation Reduces Adipocyte Size and Increases Adipocyte Number in Obese Rats. Int. J. Obes. 2017, 41, 1246–1255. [Google Scholar] [CrossRef]
- Krentz, A.J.; Fujioka, K.; Hompesch, M. Evolution of Pharmacological Obesity Treatments: Focus on Adverse Side-effect Profiles. Diabetes Obes. Metab. 2016, 18, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, S.; Mohanam, L.; Alwin, D.; Prabhu, V. Effect of Combination Therapy of Melatonin and Orlistat on High Fat Diet Induced Changes in Lipid Profiles and Liver Function Parameters in Serum of Rats. Obes. Med. 2016, 2, 41–45. [Google Scholar] [CrossRef]
- Wang, L.-C.; Pan, T.-M.; Tsai, T.-Y. Lactic Acid Bacteria-fermented Product of Green Tea and Houttuynia Cordata Leaves Exerts anti-adipogenic and Anti-obesity Effects. J. Food Drug Anal. 2018, 26, 973–984. [Google Scholar] [CrossRef]
- Bai, J.; Zhu, Y.; Dong, Y. Response of Gut Microbiota and Inflammatory Status to Bitter Melon (Momordica charantia L.) in High Fat Diet Induced Obese Rats. J. Ethnopharmacol. 2016, 194, 717–726. [Google Scholar] [CrossRef]
- Daniel, P.; Supe, U.; Roymon, M.G. A Review on Phytochemical Analysis of Momordica charantia. Int. J. Adv. Pharm. Biol. Chem. 2014, 3, 214–220. [Google Scholar]
- Jia, S.; Shen, M.; Zhang, F.; Xie, J. Recent Advances in Momordica charantia: Functional Components and Biological Activities. Int. J. Mol. Sci. 2017, 18, 2555. [Google Scholar] [CrossRef] [PubMed]
- Annapoorani, C.A.; Manimegalai, K. Screening of Medicinal Plant Momordica charantia Leaf for Secondary Metabolites. Int. J. Pharm. Res. Dev. 2013, 5, 1–6. [Google Scholar]
- Shivanagoudra, S.R.; Perera, W.H.; Perez, J.L.; Athrey, G.; Sun, Y.; Jayaprakasha, G.; Patil, B.S. Cucurbitane-type Compounds from Momordica charantia: Isolation, In Vitro Antidiabetic, Anti-inflammatory Activities and in Silico Modeling Approaches. Bioorganic Chem. 2019, 87, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Farah, N.; Bukhari, S.A.; Ali, M.; Naqvi, S.A.R.; Mahmood, S. Phenolic Acid Profiling and Antiglycation Studies of Leaf and Fruit Extracts of Tyrosine Primed Momordica charantia Seeds for Possible Treatment of Diabetes Mellitus. Pak. J. Pharm. Sci. 2018, 31, 2667–2672. [Google Scholar]
- Krawinkel, M.B.; Keding, G.B. Bitter Gourd (Momordica charantia): A Dietary Approach to Hyperglycemia. Nutr. Rev. 2006, 64, 331–337. [Google Scholar] [CrossRef]
- Yoon, N.A.; Park, J.; Lee, J.; Jeong, J.Y.; Kim, H.-K.; Lee, H.S.; Hwang, I.G.; Roh, G.S.; Kim, H.J.; Cho, G.J.; et al. Anti-diabetic Effects of Ethanol Extract from Bitter Melon in Mice Fed a High-fat Diet. Dev. Reprod. 2017, 21, 259–267. [Google Scholar] [CrossRef]
- Grover, J.K.; Yadav, S.P. Pharmacological Actions and Potential Uses of Momordica charantia: A Review. J. Ethnopharmacol. 2004, 93, 123–132. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nat. Cell Biol. 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on Products of Browning Reaction Antioxidative Activities of Products of Browning Reaction Prepared from Glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Rudich, A.; Tirosh, A.; Bashan, N. Prolonged Oxidative Stress Impairs Insulin-induced GLUT4 Translocation in 3T3-L1 Adipo-cytes. Diabetes 1998, 47, 1562–1569. [Google Scholar] [CrossRef]
- Fan, M.; Choi, Y.-J.; Tang, Y.; Bae, S.M.; Yang, H.P.; Kim, E.-K. Efficacy and Mechanism of Polymerized Anthocyanin from Grape-Skin Extract on High-Fat-Diet-Induced Nonalcoholic Fatty Liver Disease. Nutrients 2019, 11, 2586. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Fan, M.; Tang, Y.; Yang, H.P.; Hwang, J.-Y.; Kim, E.-K. In Vivo Effects of Polymerized Anthocyanin from Grape Skin on Benign Prostatic Hyperplasia. Nutrients 2019, 11, 2444. [Google Scholar] [CrossRef]
- Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity: Implications for Metabolic Syndrome, Diabetes, Hypertension, Dyslipidemia, Atherosclerosis, and Cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, Oxidative Stress, and Obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef]
- Verdile, G.; Keane, K.N.; Cruzat, V.F.; Medic, S.; Sabale, M.; Rowles, J.; Wijesekara, N.; Martins, R.N.; Fraser, P.E.; Newsholme, P. Inflammation and Oxidative Stress: The Molecular Connectivity between Insulin Resistance, Obesity, and Alzheimer’s Disease. Mediat. Inflamm. 2015, 2015, 1–17. [Google Scholar] [CrossRef]
- Nijhawan, P.; Arora, S.; Behl, T. Intricate role of oxidative stress in the progression of obesity. Obes. Med. 2019, 15, 100125. [Google Scholar] [CrossRef]
- Amin, M.N.; Siddiqui, S.A.; Uddin, G.; Ibrahim, M.; Uddin, S.M.N.; Adnan, T.; Rahaman, Z.; Kar, A.; Islam, M.S. Increased Oxidative Stress, Altered Trace Elements, and Macro-Minerals Are Associated with Female Obesity. Biol. Trace Elem. Res. 2020, 197, 384–393. [Google Scholar] [CrossRef]
- Di Domenico, M.; Pinto, F.; Quagliuolo, L.; Contaldo, M.; Settembre, G.; Romano, A.; Nicoletti, G.F. The Role of Oxidative Stress and Hormones in Obesity. Front. Endocrinol. 2019, 10, 540. [Google Scholar] [CrossRef]
- Shafazila, T.S.; Lee, P.M.; Hung, L.K. Radical Scavenging Activities of Extract and Solvent-solvent Partition Fractions from Dendrobium Sonia “Red Bom” Flower. In Proceedings of the 2010 International Conference on Science and Social Research, CSSR 2010, Kuala Lumpur, Malaysia, 5–8 December 2010; pp. 762–765. [Google Scholar]
- Sun, Y.; Yang, Y.; Qin, Z.; Cai, J.; Guo, X.; Tang, Y.; Wan, J.; Su, D.-F.; Liu, X. The Acute-Phase Protein Orosomucoid Regulates Food Intake and Energy Homeostasis via Leptin Receptor Signaling Pathway. Diabetes 2016, 65, 1630–1641. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory Mechanisms Linking Obesity and Metabolic Disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef]
- Chimin, P.; Andrade, M.L.; Belchior, T.; Paschoal, V.A.; Magdalon, J.; Yamashita, A.S.; Castro, É.; Castoldi, A.; Chaves-Filho, A.B.; Yoshinaga, M.Y.; et al. Adipocyte mTORC1 Deficiency Promotes Adipose Tissue Inflammation and NLRP3 Inflammasome Activation via Oxidative Stress and de Novo Ceramide Synthesis. J. Lipid Res. 2017, 58, 1797–1807. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Stanislas, G.; Douraguia, E.; Gonthier, M.-P. Evaluation of Nutritional and Antioxidant Properties of the Tropical Fruits Banana, Litchi, Mango, Papaya, Passion Fruit and Pineapple Cultivated in Réunion French Island. Food Chem. 2016, 212, 225–233. [Google Scholar] [CrossRef]
- Fan, M.; Kim, E.-K.; Choi, Y.-J.; Tang, Y.; Moon, S.-H. The Role of Momordica charantia in Resisting Obesity. Int. J. Environ. Res. Public Health 2019, 16, 3251. [Google Scholar] [CrossRef]
- Kalaivani, A.; Uddandrao, V.V.S.; Brahmanaidu, P.; Saravanan, G.; Nivedha, P.R.; Tamilmani, P.; Swapna, K.; Vadivukkarasi, S. Anti Obese Potential of Cucurbita Maxima Seeds Oil: Effect on Lipid Profile and Histoarchitecture in High Fat Diet Induced Obese Rats. Nat. Prod. Res. 2018, 32, 2950–2953. [Google Scholar] [CrossRef]
- Brusaferro, A.; Cozzali, R.; Orabona, C.; Biscarini, A.; Farinelli, E.; Cavalli, E.; Grohmann, U.; Principi, N.; Esposito, S. Is It Time to Use Probiotics to Prevent or Treat Obesity? Nutrients 2018, 10, 1613. [Google Scholar] [CrossRef]
- Ghaedi, E.; Varkaneh, H.K.; Rahmani, J.; Mousavi, S.M.; Mohammadi, H.; Fatahi, S.; Pantovic, A.; Mofrad, M.D.; Zhang, Y. Possible Anti-obesity Effects of Phytosterols and Phytostanols Supplementation in Humans: A Systematic Review and Dose–Response Meta-analysis of Randomized Controlled Trials. Phytother. Res. 2019, 33, 1246–1257. [Google Scholar] [CrossRef]
- Hossain, M.A.; Weli, A.M.; Ahmed, S.H.I. Comparison of Total Phenols, Flavonoids and Antioxidant Activity of Various Crude Extracts of Hyoscyamus Gallagheri Traditionally Used for the Treatment of Epilepsy. Clin. Phytoscience 2019, 5, 20. [Google Scholar] [CrossRef]
- Jiang, Y.; Ding, S.; Li, F.; Zhang, C.; Sun-Waterhouse, D.; Chen, Y.; Li, D. Effects of (+)-catechin on the Differentiation and Lipid Metabolism of 3T3-L1 Adipocytes. J. Funct. Foods 2019, 62, 103558. [Google Scholar] [CrossRef]
- M Calderon-Montano, J.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A Review on the Dietary Flavonoid Kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Zhou, W.; Shan, J.; Meng, M. A Two-step Ultra-high-performance Liquid Chromatography-Quadrupole/Time of Flight Mass Spectrometry with Mass Defect Filtering Method for Rapid Identification of Analogues from Known Components of Different Chemical Structure Types in Fructus Gardeniae-Fructus Forsythiae Herb Pair Extract and in Rat’s Blood. J. Chromatogr. A 2018, 1563, 99–123. [Google Scholar] [PubMed]
- Baeza, G.; Bachmair, E.M.; Wood, S.; Mateos, R.; Bravo, L.; De Roos, B. The Colonic Metabolites Dihydrocaffeic Acid and Di-hydroferulic Acid are more Effective Inhibitors of In Vitro Platelet Activation than their Phenolic Precursors. Food Funct. 2017, 8, 1333–1342. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A Pharmacological Review and Call for Further Research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Wang, Q.; Lu, K.; Li, F.; Lei, L.; Zhao, J.; Wu, S.; Yin, R.; Ming, J. Polyphenols from Morchella Angusticepes Peck Attenuate D-galactosamine/Lipopolysaccharide-induced Acute Hepatic Failture in Mice. J. Funct. Foods 2019, 58, 248–254. [Google Scholar] [CrossRef]
- Wang, Z.; Lam, K.L.; Hu, J.; Ge, S.; Zhou, A.; Zheng, B.; Lin, S. Chlorogenic Acid Alleviates Obesity and Modulates Gut Microbiota in High-fat-fed Mice. Food Sci. Nutr. 2019, 7, 579–588. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Cao, H. Chlorogenic Acid and Caffeic Acid from Sonchus Oleraceus Linn Synergistically Attenuate Insulin Resistance and Modulate Glucose Uptake in HepG2 Cells. Food Chem. Toxicol. 2019, 127, 182–187. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and Metabolic Health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Abulizi, A.; Camporez, J.P.; Jurczak, M.J.; Høyer, K.F.; Zhang, D.; Cline, G.W.; Samuel, V.T.; Shulman, G.I.; Vatner, D.F. Ad-ipose Glucocorticoid Action Influences Whole-body Metabolism via Modulation of Hepatic Insulin Action. FASEB J. 2019, 33, 8174–8185. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Fernández-Real, J.M. Adipocyte Differentiation. In Adipose Tissue Biology; Metzler, J.B., Ed.; Springer: Cham, Switzerland, 2017; pp. 69–90. [Google Scholar]
- Choi, S.K.; Park, S.; Jang, S.; Cho, H.H.; Lee, S.; You, S.; Kim, S.-H.; Moon, H.-S. Cascade Regulation of PPARγ2 and C/EBPα Signaling Pathways by Celastrol Impairs Adipocyte Differentiation and Stimulates Lipolysis in 3T3-L1 Adipocytes. Metabolism 2016, 65, 646–654. [Google Scholar] [CrossRef]
- Kim, N.H.; Kim, H.J.; Im, J.Y.; Kwak, W.R.; Kim, Y.H.; Kim, D.K.; Lim, S.; Lee, Y. Pulsatilla Koreana Nakai Downregulates C/EBPs/PPARγ and Suppresses Fatty Acid Synthase via Activation of AMPKα in 3T3-L1 Cells. Indian J. Pharm. Sci. 2019, 81, 219–225. [Google Scholar] [CrossRef]
- Zhang, J.-M.; Wang, X.-H.; Hao, L.-H.; Wang, H.; Zhang, X.-Y.; Muhammad, I.; Qi, Y.; Li, G.-L.; Sun, X.-Q. Nrf2 is Crucial for the Down-regulation of Cyp7a1 Induced by Arachidonic Acid in Hepg2 Cells. Environ. Toxicol. Pharmacol. 2017, 52, 21–26. [Google Scholar] [CrossRef]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef]
- Ghadge, A.A.; Khaire, A.A.; Kuvalekar, A.A. Adiponectin: A Potential Therapeutic Target for Metabolic Syndrome. Cytokine Growth Factor Rev. 2018, 39, 151–158. [Google Scholar] [CrossRef]
- Goo, Y.H.; Son, S.H.; Paul, A. Lipid Droplet-associated Hydrolase Promotes Lipid Droplet Fusion and Enhances ATGL Degradation and Triglyceride Accumulation. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Chen, X.; Xue, H.; Zhang, P.; Fang, W.; Chen, X.; Ling, W. Coenzyme Q10 Attenuates High-fat Diet-induced Non-alcoholic Fatty Liver Disease through Activation of the AMPK Pathway. Food Funct. 2019, 10, 814–823. [Google Scholar] [CrossRef]
- Lin, K.-T.; Hsu, S.-W.; Lai, F.-Y.; Chang, T.-C.; Shi, L.-S.; Lee, S.-Y. Rhodiola Crenulata Extract Regulates Hepatic Glycogen and Lipid Metabolism via Activation of the AMPK Pathway. BMC Complement. Altern. Med. 2016, 16, 127. [Google Scholar] [CrossRef] [PubMed]
- Angeles, T.S.; Hudkins, R.L. Recent Advances in Targeting the Fatty Acid Biosynthetic Pathway Using Fatty Acid Synthase In-hibitors. Expert Opin. Drug Dis. 2016, 11, 1187–1199. [Google Scholar] [CrossRef]
- Harriman, G.; Greenwood, J.; Bhat, S.; Huang, X.; Wang, R.; Paul, D.; Tong, L.; Saha, A.K.; Westlin, W.F.; Kapeller, R.; et al. Acetyl-CoA Carboxylase Inhibition by ND-630 Reduces Hepatic Steatosis, Improves Insulin Sensitivity, and Modulates Dyslipidemia in Rats. Proc. Natl. Acad. Sci. USA 2016, 113, E1796–E1805. [Google Scholar] [CrossRef]
- Saponaro, C.; Gaggini, M.; Carli, F.; Gastaldelli, A. The Subtle Balance between Lipolysis and Lipogenesis: A Critical Point in Metabolic Homeostasis. Nutrients 2015, 7, 9453–9474. [Google Scholar] [CrossRef] [PubMed]
- Galic, S.; Loh, K.; Murray-Segal, L.; Steinberg, G.R.; Andrews, Z.B.; Kemp, B.E. AMPK Signaling to Acetyl-CoA Carboxylase is Required for Fasting-and Cold-induced Appetite but not Thermogenesis. Elife 2018, 7, 1–22. [Google Scholar] [CrossRef]
- Morak, M.; Schmidinger, H.; Riesenhuber, G.; Rechberger, G.N.; Kollroser, M.; Haemmerle, G.; Zechner, R.; Kronenberg, F.; Hermetter, A. Adipose Triglyceride Lipase (ATGL) and Hormone-Sensitive Lipase (HSL) Deficiencies Affect Expression of Lipolytic Activities in Mouse Adipose Tissues. Mol. Cell. Proteom. 2012, 11, 1777–1789. [Google Scholar] [CrossRef]
- Kim, S.J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK Phosphorylates Desnutrin/ATGL and Hor-mone-sensitive Lipase to Regulate Lipolysis and Fatty Acid Oxidation within Adipose Tissue. Mol. Cell Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef]
- Schulze, R.J.; Sathyanarayan, A.; Mashek, D.G. Breaking fat: The Regulation and Mechanisms of Lipophagy. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 1178–1187. [Google Scholar] [CrossRef]
- Christian, P.; Sacco, J.; Adeli, K. Autophagy: Emerging Roles in Lipid Homeostasis and Metabolic Control. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Busiello, R.A.; Savarese, S.; Lombardi, A. Mitochondrial Uncoupling Proteins and Energy Metabolism. Front. Physiol. 2015, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Chunjuan, L.; He, X.; Shi, Z.; Li, C.; Guo, F.; Li, S.; Li, Y.; Na, L.; Sun, C. Ursolic Acid Increases Energy Expenditure through Enhancing Free Fatty Acid Uptake and β-oxidation via an UCP3/AMPK-dependent Pathway in Skeletal Muscle. Mol. Nutr. Food Res. 2015, 59, 1491–1503. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time (min) | Identification (MCLW) | Exact Mass (m/z) | Fragment Ions (m/z) |
|---|---|---|---|---|
| 1 | 0.68 | Hydroferulic acid | 195.08 | 165, 133 |
| 2 | 0.91 | N-(1-Deoxy-1-fructosyl)leucine | 292.14 | 274 |
| 3 | 1.06 | Chlorogenic acid fragments | 353.08 | 323, 191, 129 |
| 4 | 1.37 | Abscisic acid fragments | 263.13 | 188, 144 |
| 5 | 1.43 | Hyperforin fragments | 535.37 | 257, 243 |
| 6 | 1.70 | Rosmaricine | 344.09 | 150 |
| 7 | 1.85 | Biotin fragments | 243.08 | 200, 156 |
| 8 | 2.18 | N-[2-Hydroxy-2-(4-hydroxyphenyl)ethyl]cinnamide | 282.12 | 150, 133 |
| 9 | 2.73 | Kaempferol 7-arabinoside | 417.16 | 357, 215, 135 |
| 10 | 2.91 | alpha,alpha-Trehalose | 341.14 | 135, 119 |
| 11 | 3.19 | Kaempferol 3-glucuronide | 461.14 | 399, 285 |
| 12 | 3.36 | Orientin 7,3′-dimethyl ether | 475.16 | 445, 299 |
| 13 | 3.53 | Momordicinin | 437.26 | 421, 409, 205 |
| 14 | 3.67 | 5-(6-Hydroxy-3,7-dimethyl-2,7-octadienyloxy)-7-methoxycoumarin | 343.26 | 327, 301 |
| 15 | 3.76 | Phytolaccasaponin G | 663.47 | 645, 601, 501 |
| 16 | 4.06 | Phytolaccasaponin G | 663.47 | 645, 631, 485 |
| 17 | 4.21 | (±)-3′,4′-Methylenedioxy-5,7-dimethylepicatechin | 329.28 | 229 |
| 18 | 4.34 | Myrianthic acid | 503.41 | 487, 485 |
| 19 | 4.73 | Momordicine III | 647.47 | 485, 99 |
| 20 | 4.93 | Goyaglycoside b | 647.47 | 485 |
| 21 | 4.96 | 5beta,19-Epoxy-19-methoxycucurbita-6,23(E)-diene-3beta,25-diol | 485.40 | 469, 455 |
| 22 | 5.20 | Momordicin I | 471.42 | 99 |
| 23 | 5.48 | 5beta,19-Epoxy-19-methoxycucurbita-6,23(E)-diene-3beta,25-diol | 485.40 | 455, 325 |
| 24 | 5.54 | Madasiatic acid | 487.42 | 471 |
| 25 | 5.69 | Vernolic acid | 295.27 | 277, 195, 99 |
| Peak No. | Retention Time (min) | Identification (MCLE) | Exact Mass (m/z) | Fragment Ions (m/z) |
| 1 | 0.83 | Isorhamnetin 7-rhamnoside | 461.13 | 357, 285, 283 |
| 2 | 0.98 | Diosmetin 7-glucuroside | 475.15 | 445, 299, 269 |
| 3 | 1.48 | 6-Hydroxyluteolin 7,3′,4′-trimethyl ether | 343.25 | 327, 283, 243 |
| 4 | 1.58 | Apigenin 6-C-arabinosyl-8-C-glucoside | 695.49 | 663, 519 |
| 5 | 1.69 | Luteolin | 285.08 | 269, 135 |
| 6 | 1.87 | ent-Epiafzelechin(2a->7,4a->8)epiafzelechin 3-(4-hydroxybenzoic acid) | 663.46 | 645 |
| 7 | 2.13 | 3,3′,5-Trihydroxy-4′-methoxy-6,7-methylenedioxyflavone 3-glucuronide | 519.40 | 501, 343, 327 |
| 8 | 2.33 | Apigenin | 269.07 | 227, 151, 117 |
| 9 | 2.41 | 5-Hydroxy-7,3′,4′-trimethoxy-8-methylisoflavone 5-neohesperidoside | 649.48 | 631, 487 |
| 10 | 2.46 | Diosmetin | 299.06 | 284, 256 |
| 11 | 2.81 | 5,7-Dihydroxy-8,4′-dimethoxyisoflavone | 329.27 | 229, 139 |
| 12 | 3.04 | Phytolaccoside E | 825.57 | 617, 487 |
| 13 | 3.36 | 5beta,19-Epoxy-19-methoxycucurbita-6,23(E)-diene-3beta,25-diol | 485.39 | 455 |
| 14 | 3.80 | Goyaglycoside b | 647.46 | 631, 485, 455 |
| 15 | 3.89 | Momordicoside F1 | 631.47 | 469 |
| 16 | 4.05 | Goyaglycoside c | 661.45 | 645, 631, 499 |
| 17 | 4.14 | Momordicin II | 633.45 | 615, 471, 455 |
| 18 | 4.34 | Momordicine III | 647.46 | 617, 485 |
| 19 | 4.45 | Momordicoside K | 647.46 | 617 |
| 20 | 4.61 | Dihydroxystearic acid | 315.29 | 297 |
| 21 | 4.76 | Momordicoside F2 | 617.45 | 527, 455 |
| 22 | 4.90 | Momoridin B | 631.47 | 469, 455 |
| 23 | 5.33 | Nonadecanoic acid | 297.28 | 279 |
| 24 | 6.21 | 1,17-Heptadecanediol | 271.26 | 225 |
| 25 | 6.82 | Retinoic acid | 299.30 | 253, 99 |
| Peak No. | Retention Time (min) | Identification (MCLH) | Exact Mass (m/z) | Fragment Ions (m/z) |
| 1 | 1.17 | Biotin | 243.15 | 225, 199 |
| 2 | 1.48 | Biotin | 243.15 | 225, 197 |
| 3 | 1.97 | Cetyl alcohol | 241.13 | 197, 127, 125, 99 |
| 4 | 2.81 | 5,7-Dihydroxy-8,4′-dimethoxyisoflavone | 329.27 | 229 |
| 5 | 3.36 | 3,3′,4′,5,6,8-Hexamethoxyflavone | 401.23 | 375, 329, 313, 209 |
| 6 | 3.96 | 5,7-Dihydroxy-3′,4′-dimethoxy-5′-prenylflavanone | 383.22 | 353, 311 |
| 7 | 4.04 | 2′,3,4′,5-Tetrahydroxy-4-prenylstilbene | 311.26 | 293, 265 |
| 8 | 4.18 | Cycloheterophyllin | 501.38 | 461, 311 |
| 9 | 4.46 | Medicarpin 3-O-(6′-malonylglucoside) | 517.38 | 499, 487, 471, 311 |
| 10 | 4.66 | Limocitrin 3-rhamnoside | 491.40 | 475, 371, 313 |
| 11 | 4.98 | Phytal | 293.24 | 277, 263 |
| 12 | 5.36 | Vernolic acid | 295.26 | 279 |
| 13 | 5.86 | Madasiatic acid | 487.40 | 471, 425 |
| 14 | 5.99 | 5beta,19-Epoxy-19-methoxycucurbita-6,23(E)-diene-3beta,25-diol | 485.38 | 455, 427, 423 |
| 15 | 6.23 | 5beta,19-Epoxy-19-methoxycucurbita-6,23(E)-diene-3beta,26-diol | 485.38 | 455 |
| 16 | 6.54 | Myrianthic acid | 503.40 | 487 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, M.; Lee, J.-I.; Ryu, Y.-B.; Choi, Y.-J.; Tang, Y.; Oh, M.; Moon, S.-H.; Lee, B.; Kim, E.-K. Comparative Analysis of Metabolite Profiling of Momordica charantia Leaf and the Anti-Obesity Effect through Regulating Lipid Metabolism. Int. J. Environ. Res. Public Health 2021, 18, 5584. https://doi.org/10.3390/ijerph18115584
Fan M, Lee J-I, Ryu Y-B, Choi Y-J, Tang Y, Oh M, Moon S-H, Lee B, Kim E-K. Comparative Analysis of Metabolite Profiling of Momordica charantia Leaf and the Anti-Obesity Effect through Regulating Lipid Metabolism. International Journal of Environmental Research and Public Health. 2021; 18(11):5584. https://doi.org/10.3390/ijerph18115584
Chicago/Turabian StyleFan, Meiqi, Jae-In Lee, Young-Bae Ryu, Young-Jin Choi, Yujiao Tang, Mirae Oh, Sang-Ho Moon, Bokyung Lee, and Eun-Kyung Kim. 2021. "Comparative Analysis of Metabolite Profiling of Momordica charantia Leaf and the Anti-Obesity Effect through Regulating Lipid Metabolism" International Journal of Environmental Research and Public Health 18, no. 11: 5584. https://doi.org/10.3390/ijerph18115584
APA StyleFan, M., Lee, J.-I., Ryu, Y.-B., Choi, Y.-J., Tang, Y., Oh, M., Moon, S.-H., Lee, B., & Kim, E.-K. (2021). Comparative Analysis of Metabolite Profiling of Momordica charantia Leaf and the Anti-Obesity Effect through Regulating Lipid Metabolism. International Journal of Environmental Research and Public Health, 18(11), 5584. https://doi.org/10.3390/ijerph18115584