Abstract
Polycyclic aromatic hydrocarbons (PAHs) are organic compounds that are widely distributed in the air, water, and soil. Recently, the amount of PAHs derived from fuels and from incomplete combustion processes is increasing. In the aquatic environment, oil spills directly cause PAH pollution and affect marine organisms. Oil spills correlate very well with the major shipping routes. Furthermore, accidental oil spills can seriously impact the marine environment toxicologically. Here, we describe PAH toxicities and related bioaccumulation properties in aquatic animals, including invertebrates. Recent studies have revealed the toxicity of PAHs, including endocrine disruption and tissue-specific toxicity, although researchers have mainly focused on the carcinogenic toxicity of PAHs. We summarize the toxicity of PAHs regarding these aspects. Additionally, the bioaccumulation properties of PAHs for organisms, including invertebrates, are important factors when considering PAH toxicity. In this review, we describe the bioaccumulation properties of PAHs in aquatic animals. Recently, microplastics have been the most concerning environmental problem in the aquatic ecosystem, and the vector effect of microplastics for lipophilic compounds is an emerging environmental issue. Here, we describe the correlation between PAHs and microplastics. Thus, we concluded that PAHs have a toxicity for aquatic animals, indicating that we should emphasize the prevention of aquatic PAH pollution.
1. Introduction
Polycyclic aromatic hydrocarbons (PAHs), a chemical group that has two or more condensed aromatic rings, are ubiquitous compounds in air, water, and soil [1,2,3,4,5], and are categorized as general environmentally harmful pollutants. PAHs are especially widely detected in the aquatic environment, including water, sediment, fish, benthic invertebrates, sea birds, and sea mammals [6,7,8,9,10,11,12]. PAHs in the aquatic environment are mainly considered to be of four types: derived from fuels (petrogenic), derived from an incomplete combustion process (pyrogenic), generated by organic metabolism (biogenic), and generated by the transformation process in sediment (diagenetic) [13]. Of these four types of sources, petrogenic and pyrogenic sources are mainly artificial and are important contributors of environmental PAH pollution in aquatic ecosystems.
Regarding PAH pollution in aquatic environments, oil spill accidents are among the most concerning exposure events [14,15,16,17,18,19]. Hydrocarbon chemicals are major components of crude oil and are classified as PAHs, aliphatic saturated hydrocarbons, aliphatic unsaturated hydrocarbons, and alicyclic saturated hydrocarbons [14]. The impact of these four categories on the ecosystem from PAHs is especially concerning because of their specific toxicity. In the last several decades, several oil spill accidents have happened all over the world, and enormous amounts of crude oil have been released into the aquatic environment. The most infamous oil spill of the decade was the Deepwater Horizon Oil Spill, in which approximately 4.9 million barrels of crude oil was discharged into the Gulf of Mexico between 20 April and 15 July 2010 [20]. In this accident, discharged crude oil expanded over a wide area of the ecosystem and negatively affected the Gulf of Mexico. Several researchers found that crude oil contained PAHs that had toxic effects, such as immunotoxicity, embryonic abnormalities, and cardiotoxicity, for wildlife including fish, benthic organisms, and marine vertebrates [21,22,23].
The most concerning toxicity of PAHs is their carcinogenicity [24,25,26,27]. Briefly, PAHs are transported into cells because of their hydrophobicity and induce gene expression of the cytochrome P450 (CYP) enzyme group [28,29,30,31]. Expressed CYP enzymes metabolize PAHs into additional metabolites. It is important to note that several intermediates in this metabolic pathway can bind to DNA and become mutagenic/carcinogenic. Because of their carcinogenicity, the International Agency for Research on Cancer (IARC) classified three PAHs: benzo[a]anthracene (BaA), benzo[a]pyrene (BaP), and dibenz[a,h]anthracene, as being probably carcinogenic chemicals (group 2A). Additionally, as per the United States Environmental Protection Agency (US EPA), the emissions to the environment of 16 representative PAHs are monitored (Figure 1). PAHs are considered carcinogenic chemicals and are concerning as they are important organic pollutants in the environment and human society (Figure 1). Moreover, additional toxicological studies have revealed other types of toxicities from PAHs: developmental toxicity, genotoxicity, immunotoxicity, oxidative stress, and endocrine disruption [32,33,34,35,36]. Because of their ubiquity in the natural environment and various harmful effects on organisms, PAHs are among the most concerning organic pollutants.
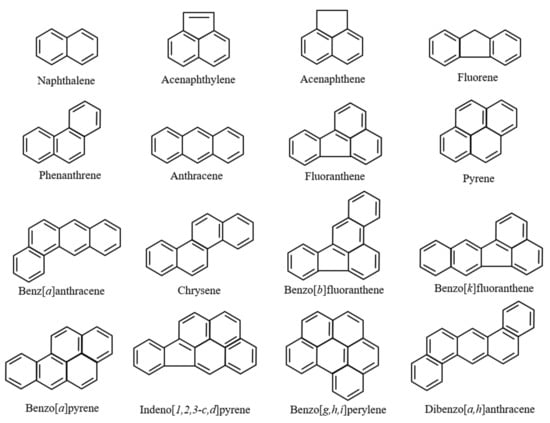
Figure 1.
Chemical structure of the 16 representative polycyclic aromatic hydrocarbons (PAHs) as decided upon by the United States Environmental Protection Agency (US EPA).
Recently, microplastics have emerged as one of the most concerning environmental problems in the aquatic ecosystem [37,38,39,40]. Even though toxicological studies of microplastics are occurring, their toxic effects on organisms are still unclear [41,42]. The vector effect of microplastics on lipophilic compounds is known to be an indirect effect of microplastics on the ecosystem [43,44], and is considered an emerging environmental issue. In the same way, it is hypothesized that PAHs are absorbed, transported, and exposed to organisms via microplastics [45,46,47]. Therefore, it is essential to describe not only general exposure pathways, such as via water or sediment, but also the vector effect via microplastics on the PAH exposure to organisms.
This review describes traditional and current studies of PAH toxicities and the related bioaccumulation properties in aquatic animals. Generally, researchers have mainly focused on the carcinogenic toxicity of PAHs; however, recent studies have revealed additional toxicities, including endocrine disruptions and tissue-specific toxicity. Additionally, the bioaccumulation properties of PAHs for organisms are important factors to consider regarding PAH toxicity. Finally, the correlation of PAHs and microplastics is additionally described here.
2. Toxicities of PAHs in Aquatic Animals
2.1. Carcinogenic Properties of PAHs in Mammals and Fish
Researchers have paid attention to the carcinogenicity of PAHs to mammals, including humans. Eight PAHs—BaA, chrysene, benzo[b]fluoranthene, benzo[k]fluoranthene, BaP, dibenz[a,h]anthracene, indeno[1,2,3-c,d]pyrene, and benzo[g,h,i]perylene—are considered possible carcinogens [48]. In particular, BaP has been identified as highly carcinogenic [49,50]. As there is 20–40 ng of BaP per cigarette [51], the relationship between mutations caused by BaP and lung cancer has been investigated. It has been reported that 60% of lung cancer cases were due to mutations caused by BaP and a few other PAHs [52]. Furthermore, it is known that BaP induces several carcinogenic responses in the cervix, bladder, breast, and prostate [53].
In aquatic animals, such as fish, epizootic neoplasia is strongly associated with environmental chemical pollution, which has increased exponentially since the 1940s with the growth of synthetic organic chemical-producing industries [54]. Certain fish species (e.g., rainbow trout and medaka) are well-established sensitive models for evaluating the effects of exogenous and endogenous factors on chemical carcinogenesis [55,56]. In feral fish, carcinogenic properties of PAHs have also been examined in English sole (Parophrys vetulus) and flounder (Platichthys stellatus). The level of BaP binding to hepatic DNA was 10 times higher in juvenile sole compared with adult sole and 90 times higher in juvenile sole than in Sprague Dawley rats, a species that is resistant to BaP-induced hepatocarcinogenesis [56]. Furthermore, the level of chemical modification of hepatic DNA in juvenile flounder was two to four times lower than that in juvenile sole, and the concentration of BaP 7,8-diol glucuronide in the bile of sole was significantly higher than that in flounder bile [56].
In fish, as well as mammals, the carcinogenic properties of PAHs have been reported. In fish, however, there have been more toxicological than carcinogenesis studies of PAHs. Thus, in this review, we mainly describe toxicological studies of early development, bone metabolism, liver metabolism, and reproduction in fish. Actually, the toxicological bioassay, using fish such as medaka (Oryzias latipes) and zebrafish (Danio rerio), has been adopted in the Organisation for Economic Co-operation and Development (OECD) guidelines. In addition, we describe PAHs attached to microplastics because plastic pollution is a worldwide problem in marine environments.
2.1.1. Toxicity of PAHs on the Early Development of Fish
The teleost fish embryo is particularly sensitive to PAHs at two distinct stages of development [33]. The first is early during the cleavage stages when PAHs alter the normal signaling associated with the establishment of the dorsal–ventral axis. This disruption involves the Wnt/β-catenin pathway and results in hyperdorsalized embryos that do not survive to hatching. The second, more sensitive, period is during the development of the heart. The heart is susceptible to chemical contaminants, including PAHs in aquatic and marine habitats, and the disruption in cardiac function impacts fish survival at all life stages [57]. The cardiovascular system is important for extracting oxygen from the atmosphere, or more specifically, for delivering oxygen to cell mitochondria and modulating cardiac output to meet the metabolic demands of active tissue [58]. In fish and other vertebrates, swim performance is dependent on increases in cardiac output [58], indicating that the disruption of cardiac function by pollutants is a major threat to fish. In crude oil from the Deepwater Horizon oil spill, which included three-ring PAH congeners (i.e., phenanthrenes), the mechanism of embryonic heart failure was demonstrated through two pathways: (1) the inhibition of the inwardly rectifying potassium channel, which drives the repolarization of cardiac action potentials; and (2) a disruption of intracellular calcium cycling in cardiomyocytes, either by blocking the ryanodine receptor or the sarcoplasmic reticulum calcium pump [59]. In addition to impacting cardiac function, PAHs contained in crude oil have been shown to cause the dysregulation of genes important in eye development and function, as well as morphological abnormalities of the eye [60]. The mean diameters of retinal layers and optomotor response were significantly reduced in oil-exposed larvae [60]. Embryos particularly sensitive to dispersed crude oil have been reported [61]. The embryos of Atlantic haddock (Melanogrammus aeglefinus) were fouled by crude oil droplets adhering to the chorion when exposed to concentrations of more than 0.7 μg/L tPAH [61]. This correlated with an increase in toxicological responses (malformations and cardiotoxicity). The early development of fish is influenced by PAHs though several phenomena.
The Japanese medaka (Oryzias latipes) is a model fish used in the OECD guidelines for testing chemicals. In mammals, oxygenated PAHs, including monohydroxylated PAHs (OHPAHs), have been noted to be toxic substances [62]. In medaka embryos, as well as mammals, OHPAHs were toxic for embryogenesis [63].
As OHPAHs, especially 3-hydroxybenzo[c]phenanthrene (3-OHBcP), may possess a strong toxic effect on the endocrine system of vertebrates [64], we examined the influence of 3-OHBcP on fish embryogenesis using an in ovo nanoinjection method. Nanoinjection uses a special glass micropipette to inject a nanolevel volume of liquid solution into a living cell under a microscope by using a micromanipulator. This method is widely known in transgenic experiments [65].
By injecting 3-OHBcP (1 nM) in ovo, the development of medaka embryos on the first, fourth, and sixth days post fertilization (dpf) was promoted. On the fifth dpf after injecting 3-OHBcP, the heart rates of embryos in the 1 nM 3-OHBcP exposure group were significantly higher than those in the control and solvent control groups [63]. Using mRNA-Seq data analysis, the detailed mechanisms of these phenomena were investigated. The 780 genes between the solvent-control (four replicates) and the 3-OHBcP-exposure (three replicates) groups had significant expression differences. The mRNA-Seq analysis indicated that many genes related to heart development in exposed embryos significantly increased compared with those in control embryos. These results indicate that an abnormal development of the heart in the 3-OHBcP-exposed medaka embryo had occurred. Also, the expression of genes related to eye development (lens, beaded filament, and crystalline) increased due to 3-OHBcP exposure, as shown above [60]. Furthermore, the expression of genes related to muscle development, energy supply, and stress-response proteins significantly changed during early development in medaka. Thus, 3-OHBcP, which is a metabolite of benzo[c]phenanthrene, acts on several organs and is toxic to fish embryogenesis.
2.1.2. Toxicity of PAHs on the Bone Metabolism of Fish
BaP and 7,12-dimethylbenz[a]anthracene, including cigarette smoke, induced a loss of bone mass and bone strength [66]. BaP was shown to inhibit osteogenesis in rat bone marrow cells [67]. Furthermore, in humans, associations between the contents of urinary PAHs and bone mass density were stronger for postmenopausal women when compared with the premenopausal group [68]. Therefore, atmospheric PAHs influence mammalian bone metabolism. In fish, as well as mammals, PAH exposure induces bone disruption in Pacific herring, pink salmon, and medaka [69,70,71], suggesting that more attention should be given to fish bone metabolism. However, the direct effect of PAHs on osteoclasts and osteoblasts has not been investigated in fish because of the lack of a suitable bioassay system for analyzing bone metabolism.
A teleost scale is a calcified tissue in which osteoblasts (Figure 2a), osteoclasts (Figure 2b), and a calcified bone matrix coexist [72,73,74,75,76]. The bone matrix, which includes type I collagen [77], osteocalcin [78], osteonectin [79], and hydroxyapatite [80], is present in scales, as well as in mammalian bone. Teleost scales have an important function in regulating blood calcium levels. Teleost scales are known to function as internal calcium reservoirs similar to those in the endoskeletons of mammals [81,82,83].
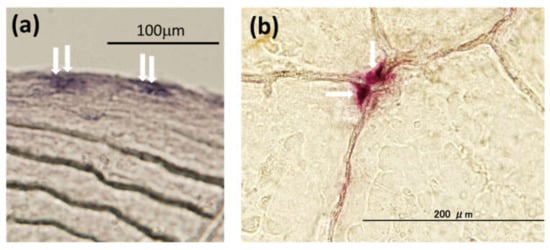
Figure 2.
Typical osteoblasts (a) and osteoclasts (b) in goldfish scales: (a) alkaline phosphatase staining for osteoblasts (arrows), and (b) tartrate-resistant acid phosphatase staining for osteoclasts (arrows).
Using teleost scales, we developed a novel in vitro assay system [81,84]. This system can simultaneously detect the activities of both scale osteoblasts and osteoclasts with alkaline phosphatase (ALP) and tartrate-resistant acid phosphatase (TRAP) as respective markers because, in mammals, the effects of bioactive substances, such as hormones, on osteoclasts and osteoblasts have been investigated using ALP and TRAP as respective markers [85,86,87]. Using the scale assay system, we demonstrated that calcemic hormones, such as parathyroid hormone (PTH) and calcitonin (CT), function in osteoblasts and osteoclasts. In the scales of goldfish, PTH, a hypercalcemic hormone, acts on osteoblasts, and then stimulates osteoclastogenesis via receptor activators of nuclear factor-κB/receptor activators of the nuclear factor-κB ligand (RANKL) pathway, just as PTH does in mammalian osteoblasts and osteoclasts [82]. CT, which is well known as a hypocalcemic hormone, suppresses osteoclastic activity in the scales of goldfish, a freshwater teleost [81,88,89], and nibbler fish, a marine teleost [81]. In addition to calcemic hormones, our bioassay was sensitive to pollutants. The concentrations of cadmium and gadolinium (even at 10−13 M) functioned in osteoclasts in the scales of goldfish [90,91]. Also, even 10−10 M tributyltin, a kind of marine environmental pollutant, significantly inhibited osteoblastic activity in goldfish [92].
Oil spills correlate very well with major shipping routes [93]. Oil contains several kinds of PAHs [64]. Furthermore, spinal deformities were observed in fish inhabiting sea areas polluted by crude and heavy oil resulting from tanker accidents [94]. Worldwide, polluted areas exist, even in the absence of oil tanker accidents. The Suez Canal in Egypt links the Mediterranean Sea to the Red Sea. Since its inauguration in November 1869, many ships and oil tankers have used this Suez Canal. Furthermore, Alexandria, located at the mouth of the Nile, is a very important port of the Mediterranean Sea route and is used as a fishing port with several kinds of marine resources. At both sites, crude oil is often contained in the ballast water thrown away by ships, and the marine pollution of the Mediterranean Sea coast and the Suez Canal worsens even if there is no ship or oil tanker accident. We have reported that the concentrations of PAHs, including the PAHs shown in Figure 1, in these areas (Suez Canal: 992.56 ng/L; Alexandria port: 1364.59 ng/L) were remarkably high, around 100 times that of the Sea of Japan [95]. Furthermore, we demonstrated that they were more likely caused by PAHs included in crude oil [95]. Each sample of polluted seawater was added into culture medium at dilution rates of 50, 100, and 500 times and incubated with goldfish scales for 6 h. Thereafter, ALP and TRAP activities in the scales of goldfish were measured. The results showed that ALP activity in the scales was significantly suppressed by both polluted seawater samples, even if seawater was diluted to 500 times, although TRAP activity did not change. The mRNA expressions of osteoblastic markers (ALP, osteocalcin, and RANKL) were also significantly suppressed by polluted seawater. Furthermore, at both the Alexandria site on the Mediterranean Sea and the Suez Canal site on the Red Sea, highly concentrated PAHs (naphthalene and acenaphthene) were investigated. The influence of these chemicals on ALP activity in scales was examined to confirm the toxicity of PAHs on fish bone metabolism. The concentrations of PAHs (naphthalene and acenaphthene) were each 6 ng/L. With the addition of acenaphthene, the ALP activity in the scales of goldfish decreased significantly (Figure 3). Naphthalene tended to decrease the activity (Figure 3). Thus, polluted seawater suppressed osteoblastic activity in the scales of goldfish through the additive and/or synergistic actions of these PAHs and was toxic to bone metabolism in teleosts.
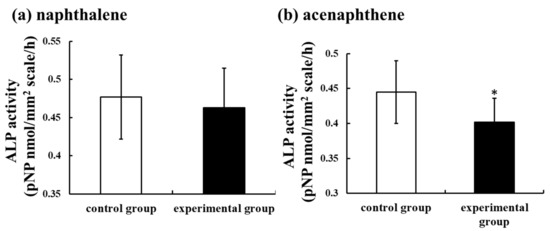
Figure 3.
Effect of naphthalene (a) and acenaphthene (b) (each 6 ng/L) on alkaline phosphatase (ALP) activity in cultured scales incubated for 6 h. The results are expressed as the means ± SE. The statistical significance between the control and experimental groups was assessed using a paired t-test. In all cases, the significance level was selected at p < 0.05. *: p < 0.05; n = 9 samples; one sample per fish. Data from Suzuki et al. [95].
2.1.3. Toxicity of PAHs on the Liver Metabolism of Fish
The liver is one target organ for PAHs because the bioaccumulation of PAHs occurs in the fish liver [96]. Most cases for PAH bioaccumulation in fish have involved benthic or bottom-feeding fish living in habitats with sediment contaminated by PAHs [97]. In fact, PAH levels were measured in Solea solea tissue and in marine sediments collected from three areas of the northern Adriatic Sea characterized by different anthropic impacts (Venetian Lagoon, Po Delta, and fishing grounds off Chioggia) [98]. As a result, the concentration of PAHs in sediment was related to PAH bioaccumulation in fish [98]. In fish, isolated hepatocytes or sliced livers have been used for experimental materials for toxicological bioassay of PAHs [99,100]. Using hepatocytes and sliced livers, carcinogenic actions [99,101] and endocrine disruptive actions [100] were investigated. Furthermore, in the liver of Chinese rare minnows (Gobiocypris rarus), data indicated that BaP may induce apoptosis [102]. Namely, BaP exposure significantly upregulated the mRNA levels of apoptosis-related genes, such as p53, bax, bcl-2, and caspase-9, as well as causing elevated caspase 3 and caspase 8 activities [102].
We recently examined the influence of BaA on liver metabolism in marine fish (nibbler fish, Girella punctate) [103]. BaA (1 or 10 ng/g body weight) was intraperitoneally injected (four times) into nibbler fish during breeding for 10 days. Thereafter, we analyzed the plasma marker of liver diseases in BaA-treated fish. We found that total protein, metabolic enzyme (alkaline phosphatase and lactate dehydrogenase) activities in liver, total cholesterol, free cholesterol, and high-density lipoprotein levels significantly decreased in BaA-injected fish. It has been reported that BaP showed a strong repression of genes involved in cholesterol and fatty acid biosynthesis [104]. These results support our results. In addition, there is an association between endoplasmic reticulum dysfunction and lipid metabolism induced by BaP exposure [105]. Therefore, PAHs function in the liver and disrupt lipid metabolism in fish. However, studies on PAH and glucose metabolism in fish have been limited. Administering BaP to flounder increases cortisol and glucose levels [106] and may be related to stress.
2.1.4. Toxicity and Endocrine-Disruptive Action of PAHs on Fish Reproduction
PAHs are toxic not only to the liver, but also to the gonads. BaP exposure induced important changes in the gene expression patterns in the liver and testes [107]. Alterations that were shared by both the liver and testes included arachidonic acid metabolism, androgen receptor to prostate-specific antigen signaling, and insulin-associated effects on lipogenesis [107]. In the case of testis-specific actions, BaP is toxic to immune system functions, inflammatory responses, and estrogen and androgen metabolic pathways [107]. These endocrine-disruptive actions may be related to OHPAHs, which are metabolites of PAHs.
A common feature of the structure of estrogenic compounds is a phenol group with a hydrophobic moiety at the para position without a bulky group at the ortho position [28]. Therefore, the structural similarity of several OHPAHs to 17β-estradiol induces the potency of estrogenic or antiestrogenic activities.
Using a yeast two-hybrid assay, OHPAHs have been demonstrated to bind to human estrogen receptors (ERs), while PAHs did not [108]. Several OHPAHs with four aromatic rings, such as 3-hydroxybenz[a]anthracene (3-OHBaA), 4-hydroxybenz[a]anthracene (4-OHBaA), and 3-OHBcP, bound to human ERs and possessed estrogenic and antiestrogenic activity [108]. Furthermore, in rat cytosol, 2-hydroxybenz[a]anthracene bound strongly to ERs [109]. In the ERα reporter assay with a human breast cancer cell line (MCF-7), 3-OHBaA and 9-hydroxybenz[a]anthracene indicated binding activity to ERα [110,111].
OHPAHs are also generated in animal bodies. After entering the body, PAHs bind to one of the nuclear receptors, the aryl hydrocarbon receptor (AhR), and then activate cytochrome P450 drug-metabolizing enzymes, such as Cyp1A1, Cyp1A2, and Cyp1B1, which metabolize PAHs into various PAH derivatives, including OHPAHs [28,112]. In teleost species, as well as in mammals, both AhR and Cyp1A1 are present [70,113]. Therefore, endocrine disruption may be caused by OHPAHs but not by PAHs.
2.1.5. Possible Toxicity of PAHs Attached to Microplastics
Recently, plastic pollution of the marine environment has been increasing. The annual global production of plastics was estimated to be approximately 322 million tons in 2015 [114]. The widespread use of plastic products causes a big problem in the marine environment. In particular, microplastic contaminants, small plastic particles with a diameter of less than 5 mm, are vectors for the transport and accumulation of pollutants, such as PAHs [45]. The PAH contents in microplastics are indicated in Table 1.

Table 1.
Attached PAH contents in microplastic.
Beach sediments in Spain and Brazil contained microplastic pellets and fragments [46,47]. The content of PAHs was extremely high, although fluctuations in the quantities of PAHs were observed. Microplastics were detected in surface water [114]. High levels of PAHs were attached to microplastics [114]. Therefore, the attached PAHs may display toxicity to aquatic animals. However, BaP eluted from microplastics did not reach sufficiently high concentrations to induce morphological effects in the fish embryo toxicity test [115]. Furthermore, juveniles (18 days after hatching) were exposed to microplastics, or pyrene (100 nM), or a combination of both, and the feeding rates and foraging activities (swimming) were examined [116]. Exposure to only microplastics did not significantly affect feeding performance in the juvenile fish, while pyrene showed a strong influence on fish behavior when concentrations were above 100 nM. The test combining pyrene with microplastics had no effect on feeding, while swimming speed decreased significantly.
Considering these facts, there are many unclear points regarding the toxicity of attached PAHs on microplastics. Further studies are needed to elucidate the toxicity of microplastics in fish.
2.2. Toxicities of PAHs in Invertebrates
2.2.1. Lethal Concentration 50% (LC50) in Invertebrates
Toxicological studies of invertebrates have been performed, and LC50 has been measured based on OECD guidelines. Sese et al. [117] reported the toxicity of acenaphthene, phenanthrene, anthracene, fluoranthene, pyrene, and BaP to Caenorhabditis elegans compared with other crustaceans, Daphnia magna, Artemia salina, and Chironomus tentans. The values of LC50 are summarized in Table 2 [117,118,119,120,121,122,123]. The sensitivities of Caenorhabditis elegans to PAHs: acenaphthene, phenanthrene, anthracene, and fluoranthene were less than those of Artemia salina and Chironomus tentans. However, Caenorhabditis elegans was sensitive to BaP. Daphnia magna was the most sensitive to fluoranthene. Both Daphnia magna and Artemia salina were very sensitive to pyrene. In addition, the toxicity of PAHs was examined using the earthworm (Eisenia fetida) [124] and was compared with other invertebrates (Table 2). The LC50 value after 72 h of exposure to phenanthrene was 114 μg/L. However, other PAHs, such as anthracene, fluoranthene, and pyrene, did not exhibit lethal toxicity to earthworms. Therefore, it was concluded that different animal species among invertebrates have different toxicities to the same PAHs, suggesting that we need to evaluate the toxicity of PAHs using many species rather than just one.
Table 2.
Lethal concentration 50% (LC50) values (μg/L) of PAHs to Caenorhabditis elegans, Daphnia magna, Artemia salina, Chironomus tentans, and Eisenia fetida.
2.2.2. Toxicity of OHPAHs to Sea Urchins
Until now, the sea urchin has been used in ecotoxicological studies [125,126]. The effect of various chemicals, including PAHs, on the development of sea urchins has been evaluated [127,128,129]. In the marine environment, the lipids and organic carbons of invertebrates have been exposed to and accumulated PAHs [130]. However, the influence of OHPAHs on invertebrates has not been reported yet. Thus, we have noted that the sea urchin is an established experimental animal for toxicological studies in invertebrates, and we examined the effect of both PAHs and OHPAHs on the embryogenesis of sea urchins. The results were described in Suzuki et al. [131]. Adult sea urchins (Hemicentrotus pulcherrimus) were collected from the shore of the Toyama Bay side of the Noto Peninsula. Spawning was induced via the intracoelomic injection of KCl (0.5 M). Eggs and sperm from spawning animals were collected in 50 mL beakers containing filtered seawater (FSW). Prior to fertilization, the eggs were washed twice with FSW. Eggs that reached at least 95% fertilization within 10 min postinsemination were used. The eggs were divided into control and experimental groups. After fertilization, BaA and 4-OHBaA were added to seawater at concentrations of 10−8 and 10−7 M, respectively, and kept at 18 °C while mixing lightly. There were no differences in the external features of the control and experimental groups in the blastula and prism stages. In the pluteus stage, morphological features changed. Spicule lengths (arrows in Figure 4) were measured using embryos crushed by a cover glass. Spicule lengths were significantly suppressed by 4-OHBaA (10−8 and 10−7 M). Figure 4 indicates the influence of 4-OHBaA on the early development of sea urchins compared with the control. BaA (10−7 M) suppressed the spicule length significantly, while the length did not change with BaA (10−8 M). The mRNA expression of the Hemicentrotus pulcherrimus spicule matrix protein 50 gene, which is a kind of spicule matrix protein, decreased significantly with 4-OHBaA treatment. Hemicentrotus pulcherrimus E26 transformation-specific gene 1 and Hemicentrotus pulcherrimus Aristaless-like homeobox gene 1, which are important transcription factors related to spicule formation, were significantly inhibited with 4-OHBaA. To determine the 4-OHBaA in BaA-treated embryos, pluteus-stage embryos treated with BaA (10−7 M) were analyzed using high-performance liquid chromatography with fluorescence detection. As a result, 4-OHBaA (1.55 pmol) was detected in the BaA-treated embryos, although 4-OHBaA was not detected in the control embryos. In addition, our further study indicated that BaA and 4-OHBaA treatment significantly inhibited the expression of vascular endothelial growth factor (VEGF) and heparan sulfate 6-O endosulfatase [132], suggesting that BaA and 4-OHBaA suppress spicule formation via disturbing the VEGF signaling pathway. Considering these facts, we believe that OHPAHs converted from PAHs are toxic substances that inhibit early embryogenesis in sea urchins and fish.
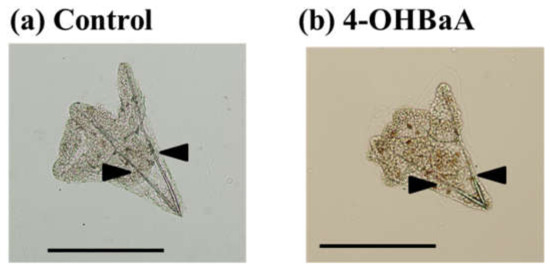
Figure 4.
Influences on spicule formation in (a) control and (b) 4-hydroxybenz[a]anthracene (4-OHBaA)-treated (10−7 M) embryos. Spicule length (arrows) was measured using embryos crushed with a cover glass. Bar: 200 µm.
3. Bioaccumulation of PAHs
3.1. General Trend of the Bioaccumulation of PAHs in Aquatic Organisms
The bioaccumulation of PAHs in aquatic animals has affected several factors, such as the octanol/water partition coefficient (Kow) of each PAH congener, concentration in environmental media, bioavailability, and depuration/excretion of PAHs [133,134,135]. PAHs are hydrophobic chemicals that have a high affinity with organic matter in water and sediment compared to the water phase. This trend is more predominant in high-molecular-weight PAHs (more than five-ring) than in low-molecular-weight PAHs because of high Kow values. Typical persistent organic pollutants, such as polychlorinated biphenyls, have the same trend, and high Kow values generally suggest a high bioaccumulation factor [135]. However, this bioaccumulation trend in aquatic animals is rarely observed in several trophic biomagnification studies [134,136,137]. For example, fish are considered to have a higher metabolism capacity and can metabolize/depure PAHs quickly; therefore, a generally positive correlation between the concentration of PAHs in the body and the Kow value is not observed in higher trophic-level fish [134]. Additionally, several previous studies suggested that species differences in the metabolism capacity of PAHs are strongly suggested for fish and invertebrates [138,139,140]. These differences may be caused by species differences in intake pathway and efficiency, capacity of xebiotics to metabolize, and ability of depuration/excretion.
The pathways of PAH accumulation in organisms are also varied in aquatic animals. Exposure pathways in aquatic organisms are considered to occur via respiration, the ingestion of food, sediments, suspended particles, and dermal absorption from the surrounding water (especially through gills) [141,142]. Compared with highly mobile animals, such as fish, benthic invertebrates are more affected by sediment and suspended particles regarding accumulation patterns that depend on their habitat [135,143,144]. In one case, bivalves that are commonly used as environmental monitoring species in coastal areas accumulated PAHs into their soft bodies via the suspension of organic matter because of their food habitats [145]. Because of huge species differences in these bioavailabilities and/or habitats, it is difficult to discuss general trends of PAH accumulation in vertebrates/invertebrates. To consider the bioaccumulation patterns of PAHs, it is necessary to discuss each organism separately, as these patterns depend on organisms’ metabolism capacities and habitats.
3.2. Bioaccumulation of PAHs in Fish
Many studies have found varied and detectable concentrations of PAHs in fish and other marine vertebrates worldwide [9,98,146,147,148,149,150]. Compared with other environmental pollutants, such as dichlorodiphenyltrichloroethanes [151], the half-lives of PAHs in organisms are relatively short and are considered to be metabolized/excreted quickly [148]. However, even with this background, detectable concentrations of PAHs are reported in many studies. Thus, this phenomenon suggests that continuous exposure to and contamination by PAHs are occurring worldwide. Because of the quick metabolism, it is not considered that the biomagnification of PAHs is occurring on the trophic level in the food chain [136,137,152]. Huang et al. [148] studied PAH concentrations in the Great Lakes and found lower concentrations of PAHs in lake trout (carnivorous fish) compared with omnivorous fish studied previously including invertebrates (Table 3) [153,154,155,156,157,158]. Additionally, higher trophic-level fish (carnivorous) generally have a higher capacity to metabolize PAHs and lower concentrations of PAHs compared with lower trophic-level fish (herbivorous, omnivorous) aquatic ecosystems [152]. However, some other studies have suggested the biomagnification of PAHs in fish. For example, Cheung et al. [146] detected higher concentrations of PAHs in the carnivorous fish golden threadfin bream Nemipterus virgatus and catfish Clarias fuscus compared with herbivorous/omnivorous fish. It is difficult to obtain a consensus on the PAH-accumulation trend in fish among trophic levels due to their huge differences in PAH bioavailability and habitat between species [140,150,159,160].
Table 3.
Total concentrations of PAHs reported for Great Lakes biota, modified from Huang et al. [148].
On the other hand, it is worth describing several trends of PAH accumulation in fish. Low-molecular-weight compounds (naphthalene and three-ring PAHs) are dominant among PAHs [133,139,140,161] due to their bioavailability, including relatively high water solubility. This bioavailability can cause higher uptake rates compared with high-molecular-weight PAHs via the surface area, especially the gill. At the same time, it indicates that the Kow values of PAHs are negatively correlated with accumulation levels [141]. Since PAHs are lipophilic compounds, tissue distributions of PAHs are correlated with lipid contents. Jafarabadi et al. [139] and Yu et al. [162] detected positive correlations between lipid contents and total PAH concentrations in marine fish, which reflected that lipid content was the important factor for tissue-specific accumulation. However, Frapiccini et al. [98], Soltani et al. [163], and Zhao et al. [142] detected extremely weak positive correlations or no correlations between lipid content and PAH concentrations in the tissues of fish. Thus, this may indicate that lipid content was not the key factor for tissue-specific distribution/accumulation in these fish species. Additionally, metabolized PAHs are excreted into bile; thus, bile tends to contain high concentrations of PAHs [142,164]. Generally, marine fish were contaminated with higher concentrations of PAHs compared with freshwater fish [146] because they were living near marine sediment that can store/accumulate PAHs [165]. The fish have a relatively higher metabolism capacity and excretion pathway for PAHs; therefore, PAH concentrations in fish are relatively low compared with those of invertebrates [134].
3.3. Bioaccumulation in Aquatic Invertebrates
It is worth mentioning that invertebrates have a lower metabolism capacity and relatively higher PAH concentrations in the body compared with fish [134]. Therefore, invertebrates are well studied regarding accumulation and pollution surveys for the biomonitoring of PAHs [7,144,163,166,167,168,169]. Biomonitoring species in coastal areas requires several special biological properties, such as wide distribution and settlement, easy sampling, high salinity tolerance capacity, and bioaccumulation properties for target chemicals [170,171]. Based on these requirements, bivalves, such as oysters and mussels, are most commonly used as “environmental indicators” on the mussel watch project that aims to monitor various contaminants in coastal areas [145,172,173,174,175,176] and, additionally, monitor PAH derivatives such as nitro PAHs and hydroxy PAHs [139,177]. For example, Tanaka and Onduka [178] collected a total of 1725 of seven species of bivalves—Mytilus galloprovincialis, Septifer virgatus, Crassostrea gigas, Perna viridis, Hormomya mutabilis, Crenomytilus grayanus, Modiolus philippinarum—from 64 sampling sites in coastal areas around the entire area of Japan and surveyed the background levels of 17 PAHs. They detected 1.6–140 ng/g-wet wt (range) and 19 ng/g-wet wt (median) of total PAH concentrations. These environmental studies were conducted not only to survey the background level, but also to monitor the accidental release of PAHs, especially via oil spills [12,15,179].
As with fish species, invertebrate species have huge differences in PAH accumulation, even within the category of shellfish [145], and deposit feeders tend to highly accumulate PAHs. Hicheky et al. [180] investigated species differences in PAH bioaccumulation among Macomona liliana (deposit feeder), Austrovenus stutchburyi (suspension feeder), and Crassostrea gigas (suspension feeder) and found significantly higher bioaccumulations in M. liliana, but a much lower lipid content, compared to the other two shellfish. PAH accumulation was dependent on the feeding habitat. Additionally, PAH kinetics between sediment and pore water are important for bioaccumulation for benthic organisms. Meador et al. [181] revealed that Amandia brevis (deposit feeder) accumulates higher Kow PAH (log Kow > 5.5) than Rhepoxynius abronius (non-deposit feeder). These results indicate that lower Kow PAH (log Kow < 5.5) can allow exposure via pore water, and higher Kow PAH (log Kow > 5.5) tends to be exposed via sediment.
The accumulation of low-molecular-weight PAHs is higher than that of high-molecular-weight PAHs in both fish and invertebrates [182], and this trend is more prominent in invertebrates. This phenomenon would be caused not only by bioavailability, such as the higher water solubility of low-molecular PAHs, but also by other biological factors. Thomann and Komlos [183] studied a model of biota-sediment accumulation factor for PAHs using sunfish and crayfish and found the high-metabolism capacity of PAHs (especially log Kow > 5) and slow absorption in the intestines while digesting. Additionally, they suggested that fish had a higher metabolism capacity of high Kow PAH compared to invertebrates, which indicates that differences in PAH bioaccumulation between fish and invertebrates may be induced by differences in their metabolisms. It is known that the CYP1A family has an important role in metabolizing PAHs [184], and CYP1A homologues are very consistent in vertebrates. However, although some studies indicate that the CYP family contributes to PAH metabolism, characteristics of CYP1A for PAH metabolism in invertebrates are still unclear.
4. Conclusions
Oil spills correlated very well with major shipping routes. Oil contains several kinds of PAHs. Worldwide, polluted areas exist even in the absence of oil tanker accidents. Actually, low-molecular-weight compounds, such as naphthalene and three-ring PAHs, are accumulated in both fish and invertebrates. The PAHs derived from the aquatic environment are accumulated and are toxic to fish and invertebrates. Additionally, we described the toxicity of OHPAHs, metabolites of PAHs. The toxicity of OHPAHs is stronger than that of PAHs, at least in fish and sea urchins. OHPAHs that occur with accumulated PAHs may have a toxic influence on aquatic animals, even if PAH levels in the aquatic environments are low (Figure 5). Thus, we should emphasize the prevention of aquatic PAH pollution.
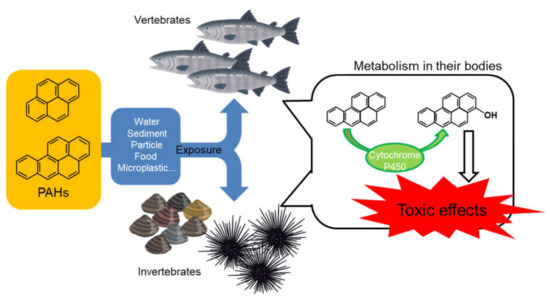
Figure 5.
General environmental fate and toxic mechanism of PAHs in the aquatic ecosystem.
Author Contributions
M.H. and N.S. planned this review and wrote the manuscript, including graph and table preparation. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by a Sasakawa Scientific Research Grant (grant number: 2019-6037) and by the cooperative research program of the Institute of Nature and Environmental Technology, Kanazawa University (19024).
Acknowledgments
We would like to thank Kitani (Kanazawa University) and Yachiguchi (Kanazawa University) for providing the beautiful photographs.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Honda, K.; Mizukami, M.; Ueda, Y.; Hamada, N.; Seike, N. Residue level of polycyclic aromatic hydrocarbons in Japanese paddy soils from 1959 to 2002. Chemosphere 2007, 68, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Nakata, H.; Uehara, K.; Goto, Y.; Fukumura, M.; Shimasaki, H.; Takikawa, K.; Miyawaki, T. Polycyclic aromatic hydrocarbons in oysters and sediments from the Yatsushiro Sea, Japan: Comparison of potential risks among PAHs, dioxins and dioxin-like compounds in benthic organisms. Ecotoxicol. Environ. Saf. 2014, 99, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Pies, C.; Hoffmann, B.; Petrowsky, J.; Yang, Y.; Ternes, T.A.; Hofmann, T. Characterization and source identification of polycyclic aromatic hydrocarbons (PAHs) in river bank soils. Chemosphere 2008, 72, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Slezakova, K.; Pires, J.C.M.; Castro, D.; Alvim-Ferraz, M.D.C.M.; Delerue-Matos, C.; Morais, S.; Pereira, M.D.C. PAH air pollution at a Portuguese urban area: Carcinogenic risks and sources identification. Environ. Sci. Pollut. Res. 2013, 20, 3932–3945. [Google Scholar] [CrossRef] [PubMed]
- Tamamura, S.; Sato, T.; Ota, Y.; Wang, X.; Tang, N.; Hayakawa, K. Long-range transport of polycyclic aromatic hydrocarbons (PAHs) from the eastern Asian continent to Kanazawa, Japan with Asian dust. Atmos. Environ. 2007, 41, 2580–2593. [Google Scholar] [CrossRef]
- Chizhova, T.; Hayakawa, K.; Tishchenko, P.; Nakase, H.; Koudryashova, Y. Distribution of PAHs in the northwestern part of the Japan Sea. Deep Sea Res. Part II 2013, 86, 19–24. [Google Scholar] [CrossRef]
- Honda, M.; Qiu, X.; Koyama, J.; Uno, S.; Undap, S.L.; Shimasaki, Y.; Oshima, Y. The wharf roach, Ligia sp., A novel indicator of polycyclic aromatic hydrocarbon contamination in coastal areas. Int. J. Environ. Res. 2018, 12, 1–11. [Google Scholar] [CrossRef]
- Hu, G.; Sun, C.; Li, J.; Zhao, Y.; Wang, H.; Li, Y. POPs accumulated in fish and benthos bodies taken from Yangtze River in Jiangsu area. Ecotoxicology 2009, 18, 647–651. [Google Scholar] [CrossRef]
- Kannan, K.; Perrotta, E. Polycyclic aromatic hydrocarbons (PAHs) in livers of California sea otters. Chemosphere 2008, 71, 649–655. [Google Scholar] [CrossRef]
- Miki, S.; Uno, S.; Ito, K.; Koyama, J.; Tanaka, H. Distributions of polycyclic aromatic hydrocarbons and alkylated polycyclic aromatic hydrocarbons in Osaka Bay, Japan. Mar. Pollut. Bull. 2014, 85, 558–565. [Google Scholar] [CrossRef]
- Pereira, M.G.; Walker, L.A.; Wright, J.; Best, J.; Shore, R.F. Concentrations of polycyclic aromatic hydrocarbons (PAHs) in the eggs of predatory birds in Britain. Environ. Sci. Technol. 2009, 43, 9010–9015. [Google Scholar] [CrossRef] [PubMed]
- Uno, S.; Koyama, J.; Kokushi, E.; Monteclaro, H.; Santander, S.; Cheikyula, J.O.; Miki, S.; Anasco, N.; Pahila, I.G.; Taberna, H.S., Jr.; et al. Monitoring of PAHs and alkylated PAHs in aquatic organisms after 1 month from the Solar I oil spill off the coast of Guimaras Island, Philippines. Environ. Monit. Assess. 2010, 165, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Hylland, K. Polycyclic aromatic hydrocarbon (PAH) ecotoxicology in marine ecosystems. J. Toxicol. Environ. Health A 2006, 69, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K. Oil spills and polycyclic aromatic hydrocarbons. In Polycyclic Aromatic Hydrocarbons; Hayakawa, K., Ed.; Springer: Singapore, 2018; pp. 213–223. ISBN 978-981-10-6775-4. [Google Scholar] [CrossRef]
- Koyama, J.; Uno, S.; Kohno, K. Polycyclic aromatic hydrocarbon contamination and recovery characteristics in some organisms after the Nakhodka oil spill. Mar. Pollut. Bull. 2004, 49, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Koyama, J.; Uno, S.; Nagai, Y.; Anukorn, B. Early monitoring of spilled oil contamination in Rayong, Thailand. Jpn. J. Environ. Toxicol. 2016, 19, 25–33. [Google Scholar] [CrossRef]
- Ladwani, K.D.; Ladwani, K.D.; Ramteke, D.S. Assessment of poly aromatic hydrocarbon (PAH) dispersion in the near shore environment of Mumbai, India after a large scale oil spill. Bull. Environ. Contam. Toxicol. 2013, 90, 515–520. [Google Scholar] [CrossRef][Green Version]
- Tronczyński, J.; Munschy, C.; Héas-Moisan, K.; Guiot, N.; Truquet, I.; Olivier, N.; Men, S.; Furaut, A. Contamination of the Bay of Biscay by polycyclic aromatic hydrocarbons (PAHs) following the T/V “Erika” oil spill. Aquat. Living Resour. 2004, 17, 243–259. [Google Scholar] [CrossRef]
- Uno, S.; Kokushi, E.; Añasco, N.C.; Iwai, T.; Ito, K.; Koyama, J. Oil spill off the coast of Guimaras Island, Philippines: Distributions and changes of polycyclic aromatic hydrocarbons in shellfish. Mar. Pollut. Bull. 2017, 124, 962–973. [Google Scholar] [CrossRef]
- McNutt, M.K.; Camilli, R.; Crone, T.J.; Guthrie, G.D.; Hsieh, P.A.; Ryerson, T.B.; Savas, O.; Shaffer, F. Review of flow rate estimates of the Deepwater Horizon oil spill. Proc. Natl. Acad. Sci. USA 2012, 109, 20260–20267. [Google Scholar] [CrossRef]
- Barron, M.G. Ecological impacts of the Deepwater Horizon oil spill: Implications for immunotoxicity. Toxicol. Pathol. 2012, 40, 315–320. [Google Scholar] [CrossRef]
- Romero, I.C.; Sutton, T.; Carr, B.; Quintana-Rizzo, E.; Ross, S.W.; Hollander, D.J.; Torres, J.J. Decadal assessment of polycyclic aromatic hydrocarbons in mesopelagic fishes from the Gulf of Mexico reveals exposure to oil-derived sources. Environ. Sci. Technol. 2018, 52, 10985–10996. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.M.; Pulster, E.L.; Wetzel, D.L.; Murawski, S.A. PAH exposure in Gulf of Mexico demersal fishes, post-Deepwater Horizon. Environ. Sci. Technol. 2015, 49, 8786–8795. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.F.; Brown, J.P.; Alexeeff, G.V.; Salmon, A.G. Potency equivalency factors for some polycyclic aromatic hydrocarbons and polycyclic aromatic hydrocarbon derivatives. Regul. Toxicol. Pharmacol. 1998, 28, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Devi, N.L.; Yadav, I.C.; Shihua, Q.; Dan, Y.; Zhang, G.; Raha, P. Environmental carcinogenic polycyclic aromatic hydrocarbons in soil from Himalayas, India: Implications for spatial distribution, sources apportionment and risk assessment. Chemosphere 2016, 144, 493–502. [Google Scholar] [CrossRef]
- Kuo, C.-Y.; Cheng, Y.-W.; Chen, Y.-W.; Lee, H. Correlation between the amounts of polycyclic aromatic hydrocarbons and mutagenicity of airborne particulate samples from Taichung City, Taiwan. Environ. Res. 1998, 78, 43–49. [Google Scholar] [CrossRef]
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Lokeshkumar, B.; Rajendran, P.; Nishigaki, I. Exposure to polycyclic aromatic hydrocarbons with special focus on cancer. Asian Pac. J. Trop. Biomed. 2015, 5, 182–189. [Google Scholar] [CrossRef]
- Bekki, K.; Toriba, A.; Tang, N.; Kameda, T.; Hayakawa, K. Biological effects of polycyclic aromatic hydrocarbon derivatives. J. UOEH 2013, 35, 17–24. [Google Scholar] [CrossRef]
- Ikenaka, Y.; Oguri, M.; Saengtienchai, A.; Nakayama, S.M.; Ijiri, S.; Ishizuka, M. Characterization of phase-II conjugation reaction of polycyclic aromatic hydrocarbons in fish species: Unique pyrene metabolism and species specificity observed in fish species. Environ. Toxicol. Pharmacol. 2013, 36, 567–578. [Google Scholar] [CrossRef]
- Jacob, J. The significance of polycyclic aromatic hydrocarbons as environmental carcinogens. 35 years research on PAH—A retrospective. Polycycl. Aromat. Compd. 2008, 28, 242–272. [Google Scholar] [CrossRef]
- Jørgensen, A.; Giessing, A.M.; Rasmussen, L.J.; Andersen, O. Biotransformation of polycyclic aromatic hydrocarbons in marine polychaetes. Mar. Environ. Res. 2008, 65, 171–186. [Google Scholar] [CrossRef]
- Bekki, K.; Takigami, H.; Suzuki, G.; Tang, N.; Hayakawa, K. Evaluation of toxic activities of polycyclic aromatic hydrocarbon derivatives using in vitro bioassays. J. Health Sci. 2009, 55, 601–610. [Google Scholar] [CrossRef]
- Cherr, G.N.; Fairbairn, E.; Whitehead, A. Impacts of petroleum-derived pollutants on fish development. Annu. Rev. Anim. Biosci. 2017, 5, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Hannam, M.L.; Bamber, S.D.; Moody, A.J.; Galloway, T.S.; Jones, M.B. Immunotoxicity and oxidative stress in the Arctic scallop Chlamys islandica: Effects of acute oil exposure. Ecotoxicol. Environ. Saf. 2010, 73, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Shim, W.J.; Lee, J.; Kim, G.B. Temporal and geographical trends in the genotoxic effects of marine sediments after accidental oil spill on the blood cells of striped beakperch (Oplegnathus fasciatus). Mar. Pollut. Bull. 2011, 62, 2264–2268. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, G.Z.; Hogan, N.S.; Köllner, B.; Thorpe, K.L.; Phalen, L.J.; Wagner, B.D.; Van Den Heuvel, M.R. Immunotoxic effects of oil sands-derived naphthenic acids to rainbow trout. Aquat. Toxicol. 2013, 126, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, M.; Wirzberger, V.; Krumpen, T.; Lorenz, C.; Primpke, S.; Tekman, M.B.; Gerdts, G. High quantities of microplastic in Arctic deep-sea sediments from the hausgarten observatory. Environ. Sci. Technol. 2017, 51, 11000–11010. [Google Scholar] [CrossRef]
- Cincinelli, A.; Scopetani, C.; Chelazzi, D.; Lombardini, E.; Martellini, T.; Katsoyiannis, A.; Fossi, M.C.; Corsolini, S. Microplastic in the surface waters of the Ross Sea (Antarctica): Occurrence, distribution and characterization by FTIR. Chemosphere 2017, 175, 391–400. [Google Scholar] [CrossRef]
- Collignon, A.; Hecq, J.H.; Glagani, F.; Voisin, P.; Collard, F.; Goffart, A. Neustonic microplastic and zooplankton in the North Western Mediterranean Sea. Mar. Pollut. Bull. 2012, 64, 861–864. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, S.; Wang, J.; Wang, Y.; Mu, J.; Wang, P.; Lin, X.; Ma, D. Microplastic pollution in the surface waters of the Bohai Sea, China. Environ. Pollut. 2017, 231, 541–548. [Google Scholar] [CrossRef]
- Nobre, C.R.; Santana, M.F.M.; Maluf, A.; Cortez, F.S.; Cesar, A.; Pereira, C.D.S.; Turra, A. Assessment of microplastic toxicity to embryonic development of the sea urchin Lytechinus variegatus (Echinodermata: Echinoidea). Mar. Pollut. Bull. 2015, 92, 99–104. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, X.; Wang, J.; Tan, L. Toxic effects of microplastic on marine microalgae Skeletonema costatum: Interactions between microplastic and algae. Environ. Pollut. 2017, 220, 1282–1288. [Google Scholar] [CrossRef]
- Koelmans, A.A.; Bakir, A.; Burton, G.A.; Janssen, C.R. Microplastic as a vector for chemicals in the aquatic environment: Critical review and model-supported reinterpretation of empirical studies. Environ. Sci. Technol. 2016, 50, 3315–3326. [Google Scholar] [CrossRef] [PubMed]
- Zarfl, C.; Matthies, M. Are marine plastic particles transport vectors for organic pollutants to the Arctic? Mar. Pollut. Bull. 2010, 60, 1810–1814. [Google Scholar] [CrossRef] [PubMed]
- Fisner, M.; Majer, A.; Taniguchi, S.; Bícego, M.; Turra, A.; Gorman, D. Colour spectrum and resin-type determine the concentration and composition of polycyclic aromatic hydrocarbons (PAHs) in plastic pellets. Mar. Pollut. Bull. 2017, 122, 323–330. [Google Scholar] [CrossRef]
- Camacho, M.; Herrera, A.; Gómez, M.; Acosta-Dacal, A.; Martínez, I.; Henríquez-Hernández, L.A.; Luzardo, O.P. Organic pollutants in marine plastic debris from Canary Islands beaches. Sci. Total Environ. 2019, 662, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Gorman, D.; Moreira, F.T.; Turra, A.; Fontenelle, F.R.; Combi, T.; Bícego, M.C.; de Castro Martins, C. Organic contamination of beached plastic pellets in the South Atlantic: Risk assessments can benefit by considering spatial gradients. Chemosphere 2019, 223, 608–615. [Google Scholar] [CrossRef]
- Menzie, C.A.; Potocki, B.B.; Santodonato, J. Exposure to carcinogenic PAHs in the environment. Environ. Sci. Technol. 1992, 26, 1278–1284. [Google Scholar] [CrossRef]
- Moorthy, B.; Chu, C.; Carlin, D.J. Polycyclic aromatic hydrocarbons: From metabolism to lung cancer. Toxicol. Sci. 2015, 145, 5–15. [Google Scholar] [CrossRef]
- David, R.M.; Gooderham, N.J. Dose-dependent synergistic and antagonistic mutation responses of binary mixtures of the environmental carcinogen benzo[a]pyrene with food-derived carcinogens. Arch. Toxicol. 2018, 92, 3459–3469. [Google Scholar] [CrossRef]
- Rodgman, A.; Smith, C.J.; Perfetti, T.A. The composition of cigarette smoke: A retrospective, with emphasis on polycyclic components. Hum. Exp. Toxicol. 2000, 19, 573–595. [Google Scholar] [CrossRef]
- Denissenko, M.F.; Pao, A.; Tang, M.S.; Pfeifer, G.P. Preferential formation of benzo[a]pyrene adducts at lung cancer mutational hotspots in P53. Science 1996, 274, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Pink, M.; Rettenmeier, A.W.; Schmitz-Spanke, S. Review on proteomic analyses of benzo[a]pyrene toxicity. Proteomics 2012, 12, 1731–1755. [Google Scholar] [CrossRef] [PubMed]
- Bunton, T.E. Experimental chemical carcinogenesis in fish. Toxicol. Pathol. 1996, 24, 603–618. [Google Scholar] [CrossRef] [PubMed]
- Bailey, G.S.; Hendricks, J.D.; Nixon, J.E.; Pawlowski, N.E. The sensitivity of rainbow trout and other fish species to carcinogens. Drug Metab. Rev. 1984, 15, 725–750. [Google Scholar] [CrossRef] [PubMed]
- Varanasi, U.; Stein, J.E.; Nishimoto, M.; Reichert, W.L.; Collier, T.K. Chemical carcinogenesis in feral fish: Uptake, activation, and detoxication of organic xenobiotics. Environ. Health. Perspect. 1987, 71, 155–1570. [Google Scholar] [CrossRef] [PubMed]
- Incardona, J.P.; Scholz, N.L. The influence of heart developmental anatomy on cardiotoxicity-based adverse outcome pathways in fish. Aquat. Toxicol. 2016, 177, 515–525. [Google Scholar] [CrossRef]
- Farrell, A.P. From hagfish to tuna: A perspective on cardiac function in fish. Physiol. Zool. 1991, 64, 1137–1164. [Google Scholar] [CrossRef]
- Brette, F.; Machado, B.; Cros, C.; Incardona, J.P.; Scholz, N.L.; Block, B.A. Crude oil impairs cardiac excitation-contraction coupling in fish. Science 2014, 343, 772–776. [Google Scholar] [CrossRef]
- Magnuson, J.T.; Khursigara, A.J.; Allmon, E.B.; Esbaugh, A.J.; Roberts, A.P. Effects of Deepwater Horizon crude oil on ocular development in two estuarine fish species, red drum (Sciaenops ocellatus) and sheepshead minnow (Cyprinodon variegatus). Ecotoxicol. Environ. Saf. 2018, 166, 186–191. [Google Scholar] [CrossRef]
- Sørensen, L.; Sørhus, E.; Nordtug, T.; Incardona, J.P.; Linbo, T.L.; Giovanetti, L.; Karlsen, Ø.; Meier, S. Oil droplet fouling and differential toxicokinetics of polycyclic aromatic hydrocarbons in embryos of Atlantic haddock and cod. PLoS ONE 2017, 12, e0180048. [Google Scholar] [CrossRef]
- Idowu, O.; Semple, K.T.; Ramadass, K.; O’Connor, W.; Hansbro, P.; Thavamani, P. Beyond the obvious: Environmental health implications of polar polycyclic aromatic hydrocarbons. Environ. Int. 2019, 123, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Tsutsumi, Y.; Yoshitake, S.; Qiu, X.; Xu, H.; Hashiguchi, Y.; Honda, M.; Tashiro, K.; Nakayama, K.; Hano, T.; et al. Alteration of development and gene expression induced by in ovo-nanoinjection of 3-hydroxybenzo[c]phenanthrene into Japanese medaka (Oryzias latipes) embryos. Aquatic. Toxicol. 2017, 182, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Nomura, M.; Nakagawa, T.; Oguri, S.; Kawanishi, T.; Toriba, A.; Kizu, R.; Sakaguchi, T.; Tamiya, E. Damage to and recovery of coastlines polluted with C-heavy oil spilled from the Nakhodka. Water Res. 2006, 40, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Porazinski, S.R.; Wang, H.; Furutani-Seiki, M. Microinjection of medaka embryos for use as a model genetic organism. J. Vis. Exp. 2010, 46. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.L.; Lee, J.S.C.; Waldman, S.D.; Casper, R.F.; Grynpas, M.D. Polycyclic aromatic hydrocarbons present in cigarette smoke cause bone loss in an ovariectomized rat model. Bone 2002, 30, 917–923. [Google Scholar] [CrossRef]
- Andreou, V.; D’Addario, M.; Zohar, R.; Sukhu, B.; Casper, R.F.; Ellen, R.P.; Tenenbaum, H.C. Inhibition of osteogenesis in vitro by a cigarette smoke-associated hydrocarbon combined with Porphyromonas gingivalis lipopolysaccharide: Reversal by resveratrol. J. Periodontol. 2004, 75, 939–948. [Google Scholar] [CrossRef]
- Guo, J.; Huang, Y.; Bian, S.; Zhao, C.; Jin, Y.; Yu, D.; Wu, X.; Zhang, D.; Cao, W.; Jing, F.; et al. Associations of urinary polycyclic aromatic hydrocarbons with bone mass density and osteoporosis in U.S. adults, NHANES 2005-2010. Environ. Pollut. 2018, 240, 209–218. [Google Scholar] [CrossRef]
- Barron, M.G.; Carls, M.G.; Heintz, R.; Rice, S.D. Evaluation of fish early life-stage toxicity models of chronic embryonic exposures to complex polycyclic aromatic hydrocarbon. Toxicol. Sci. 2004, 78, 60–67. [Google Scholar] [CrossRef]
- Billiard, S.M.; Timme-Laragy, A.R.; Wassenberg, D.M.; Cockman, C.; Giulio, R.T.D. The role of the aryl hydrocarbon receptor pathway in mediating synergistic developmental toxicity of polycyclic aromatic hydrocarbons to zebrafish. Toxicol. Sci. 2006, 92, 526–536. [Google Scholar] [CrossRef]
- Farwell, A.; Nero, V.; Croft, M.; Bal, P.; Dixon, D.G. Modified Japanese medaka embryo-larval bioassay for rapid determination of developmental abnormalities. Arch. Environ. Contam. Toxicol. 2006, 51, 600–607. [Google Scholar] [CrossRef]
- Bereiter-Hahn, J.; Zylberberg, L. Regeneration of teleost fish scale. Comp. Biochem. Physiol. Part A 1993, 105, 625–641. [Google Scholar] [CrossRef]
- Yoshikubo, H.; Suzuki, N.; Takemura, K.; Hoso, M.; Yashima, S.; Iwamuro, S.; Takagi, Y.; Tabata, M.J.; Hattori, A. Osteoblastic activity and estrogenic response in the regenerating scale of goldfish, a good model of osteogenesis. Life Sci. 2005, 76, 2699–2709. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Kitamura, K.; Nemoto, T.; Shimizu, N.; Wada, S.; Kondo, T.; Tabata, M.J.; Sodeyama, F.; Ijiri, K.; Hattori, A. Effect of vibration on osteoblastic and osteoclastic activities: Analysis of bone metabolism using goldfish scale as a model for bone. Adv. Space Res. 2007, 40, 1711–1721. [Google Scholar] [CrossRef]
- Ohira, Y.; Shimizu, M.; Ura, K.; Takagi, Y. Scale regeneration and calcification in goldfish Carassius auratus: Quantitative and morphological process. Fish. Sci. 2007, 73, 46–54. [Google Scholar] [CrossRef]
- Ikegame, M.; Hattori, A.; Tabata, M.J.; Kitamura, K.; Tabuchi, Y.; Furusawa, Y.; Maruyama, Y.; Yamamoto, T.; Sekiguchi, T.; Matsuoka, R.; et al. Melatonin is a potential drug for the prevention of bone loss during space flight. J. Pineal Res. 2019, 67, e12594. [Google Scholar] [CrossRef]
- Zylberberg, L.; Bonaventure, J.; Cohen-Solal, L.; Hartmann, D.J.; Bereiter-Hahn, J. Organization and characterization of fibrillar collagens in fish scales in situ and in vitro. J. Cell Sci. 1992, 103, 273–285. [Google Scholar]
- Nishimoto, S.K.; Araki, N.; Robinson, F.D.; Waite, J.H. Discovery of bone γ−carboxyglutamic acid protein in mineralized scales. J. Biol. Chem. 1992, 267, 11600–11605. [Google Scholar]
- Redruello, B.; Estevao, M.D.; Rotllant, J.; Guerreiro, P.M.; Anjos, L.I.; Canario, A.V.M.; Power, D.M. Isolation and characterization of piscine osteonectin and down regulation of its expression by PTH-related protein. J. Bone Miner. Res. 2005, 20, 682–692. [Google Scholar] [CrossRef]
- Onozato, H.; Watabe, N. Studies on fish scale formation and resorption III: Fine structure and calcification of the fibrillary plates of the scales in Carassius auratus (Cypriniformes: Cyprinidae). Cell Tissue Res. 1979, 201, 409–422. [Google Scholar] [CrossRef]
- Suzuki, N.; Suzuki, T.; Kurokawa, T. Suppression of osteoclastic activities by calcitonin in the scales of goldfish (freshwater teleost) and nibbler fish (seawater teleost). Peptides 2000, 21, 115–124. [Google Scholar] [CrossRef]
- Suzuki, N.; Danks, J.A.; Maruyama, Y.; Ikegame, M.; Sasayama, Y.; Hattori, A.; Nakamura, M.; Tabata, M.J.; Yamamoto, T.; Furuya, R.; et al. Parathyroid hormone 1 (1-34) acts on the scales and involves calcium metabolism in goldfish. Bone 2011, 48, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Kitamura, K.; Hattori, A. Fish scale is a suitable model for analyzing determinants of skeletal fragility in type 2 diabetes. Endocrine 2016, 54, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Hattori, A. Melatonin suppresses osteoclastic and osteoblastic activities in the scales of goldfish. J. Pineal Res. 2002, 33, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Vaes, G. Cellular biology and biochemical mechanism of bone resorption. Clin. Orthop. Relat. Res. 1988, 231, 239–271. [Google Scholar] [CrossRef]
- Dimai, H.P.; Linkhart, T.A.; Linkhart, S.G.; Donahue, L.R.; Beamer, W.G.; Rosen, C.J.; Farley, J.R.; Baylink, D.J. Alkaline phosphatase levels and osteoprogenitor cell numbers suggest bone formation may contribute to peak bone density differences between two inbred strains of mice. Bone 1998, 22, 211–216. [Google Scholar] [CrossRef]
- Suda, T.; Takahashi, N.; Udagawa, N.; Jimi, E.; Gillespie, M.T.; Martin, T.J. Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr. Rev. 1999, 20, 345–357. [Google Scholar] [CrossRef]
- Sekiguchi, T.; Suzuki, N.; Fujiwara, N.; Aoyama, M.; Kawada, T.; Sugase, K.; Murata, Y.; Sasayama, Y.; Ogasawara, M.; Satake, H. Calcitonin in a protochordate, Ciona intestinalis: The prototype of the vertebrate Calcitonin/Calcitonin gene related peptide superfamily. FEBS J. 2007, 276, 4437–4447. [Google Scholar] [CrossRef]
- Kase, Y.; Ikari, T.; Sekiguchi, T.; Sato, M.; Ogiso, S.; Kawada, T.; Matsubara, S.; Satake, H.; Sasayama, Y.; Endo, M.; et al. Sardine procalcitonin amino-terminal cleavage peptide has a different action from calcitonin and promotes osteoblastic activity in the scales of goldfish. Comp. Biochem. Physiol. Part A 2017, 211, 77–83. [Google Scholar] [CrossRef]
- Suzuki, N.; Yamamoto, M.; Watanabe, K.; Kambegawa, A.; Hattori, A. Both mercury and cadmium directly influence calcium homeostasis resulting from the suppression of scale bone cells: The scale is a good model for the evaluation of heavy metals in bone metabolism. J. Bone Miner. Metab. 2004, 22, 439–446. [Google Scholar] [CrossRef]
- Suzuki, N.; Watanabe, K.; Sekimoto, A.; Urata, M.; Zanaty, M.I.; Sekiguchi, T.; Kitani, Y.; Matsubara, H.; Srivastav, A.K.; Hattori, A. Gadolinium at low concentration suppresses both osteoclastic and osteoblastic activities in the scales of goldfish. Am. J. Environ. Sci. 2019, 15, 137–144. [Google Scholar] [CrossRef][Green Version]
- Suzuki, N.; Tabata, M.J.; Kambegawa, A.; Srivastav, A.K.; Shimada, A.; Takeda, H.; Kobayashi, M.; Wada, S.; Katsumata, T.; Hattori, A. Tributyltin inhibits osteoblastic activity and disrupts calcium metabolism through an increase in plasma calcium and calcitonin levels in teleosts. Life Sci. 2006, 78, 2533–2541. [Google Scholar] [CrossRef] [PubMed]
- Brekke, C.; Solberg, A.H.S. Oil spill detection by satellite remote sensing. Remote Sens. Environ. 2005, 95, 1–13. [Google Scholar] [CrossRef]
- de Soysa, T.Y.; Ulrich, A.; Friedrich, T.; Pite, D.; Compton, S.L.; Ok, D.; Bernardos, R.L.; Downes, G.B.; Hsieh, S.; Stein, R.; et al. Macondo crude oil from the Deepwater Horizon oil spill disrupts specific developmental processes during zebrafish embryogenesis. BMC Biol. 2012, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Sato, M.; Nassar, F.H.; Abdel-gawad, F.K.; Bassem, S.M.; Yachiguchi, K.; Tabuchi, Y.; Endo, M.; Sekiguchi, T.; Urata, M.; et al. Seawater polluted with highly concentrated polycyclic aromatic hydrocarbons suppresses osteoblastic activity in the scales of goldfish, Carassius auratus. Zool. Sci. 2016, 33, 407–413. [Google Scholar] [CrossRef]
- Rahmanpour, S.; Farzaneh Ghorghani, N.; Lotfi Ashtiyani, S.M. Polycyclic aromatic hydrocarbon (PAH) in four fish species from different trophic levels in the Persian Gulf. Environ. Monit. Assess. 2014, 186, 7047–7053. [Google Scholar] [CrossRef]
- Baumann, P.C. Epizootics of cancer in fish associated with genotoxins in sediment and water. Mutat. Res. 1998, 411, 227–233. [Google Scholar] [CrossRef]
- Frapiccini, E.; Annibaldi, A.; Betti, M.; Polidori, P.; Truzzi, C.; Marini, M. Polycyclic aromatic hydrocarbon (PAH) accumulation in different common sole (Solea solea) tissues from the North Adriatic Sea peculiar impacted area. Mar. Pollut. Bull. 2018, 137, 61–68. [Google Scholar] [CrossRef]
- Brinkmann, M.; Blenkle, H.; Salowsky, H.; Bluhm, K.; Schiwy, S.; Tiehm, A.; Hollert, H. Genotoxicity of heterocyclic PAHs in the micronucleus assay with the fish liver cell line RTL-W1. PLoS ONE 2014, 9, e85692. [Google Scholar] [CrossRef]
- Yadetie, F.; Zhang, X.; Hanna, E.M.; Aranguren-Abadía, L.; Eide, M.; Blaser, N.; Brun, M.; Jonassen, I.; Goksøyr, A.; Karlsen, O.A. RNA-Seq analysis of transcriptome responses in Atlantic cod (Gadus morhua) precision-cut liver slices exposed to benzo[a]pyrene and 17α-ethynylestradiol. Aquat. Toxicol. 2018, 201, 174–186. [Google Scholar] [CrossRef]
- Tsuji, P.A.; Walle, T. Benzo[a]pyrene-induced cytochrome P450 1A and DNA binding in cultured trout hepatocytes—nhibition by plant polyphenols. Chem. Biol. Interact. 2007, 169, 25–31. [Google Scholar] [CrossRef][Green Version]
- Zha, J.; Hong, X.; Rao, H.; Yuan, L.; Wang, Z.; Kumaran, S.S. Benzo[a]pyrene-induced a mitochondria-independent apoptosis of liver in juvenile Chinese rare minnows (Gobiocypris rarus). Environ. Pollut. 2017, 231, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Zanaty, M.I.; Sawada, N.; Kitani, Y.; Nassar, H.F.; Mahmoud, H.M.; Hayakawa, K.; Sekiguchi, T.; Ogiso, S.; Tabuchi, Y.; Urata, M.; et al. Influence of benz[a]anthracene on bone metabolism and on liver metabolism in nibbler fish, Girella punctate. Int. J. Environ. Res. Public Health. (in press).
- Souza, T.; Jennen, D.; van Delft, J.; van Herwijnen, M.; Kyrtoupolos, S.; Kleinjans, J. New insights into BaP-induced toxicity: Role of major metabolites in transcriptomics and contribution to hepatocarcinogenesis. Arch. Toxicol. 2016, 90, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Li, J.; Yu, L.; Wei, Y.; Miao, Z.; Chen, M.; Huang, R. Characterization of transcriptional responses mediated by benzo[a]pyrene stress in a new marine fish model of goby, Mugilogobius chulae. Genes Genom. 2019, 41, 113–123. [Google Scholar] [CrossRef]
- An, K.W.; Shin, H.S.; Choi, C.Y. Physiological responses and expression of metallothionein (MT) and superoxide dismutase (SOD) mRNAs in olive flounder, Paralichthys olivaceus exposed to benzo[a]pyrene. Comp. Biochem. Physiol. Part B 2008, 149, 534–539. [Google Scholar] [CrossRef]
- Colli-Dula, R.C.; Fang, X.; Moraga-Amador, D.; Albornoz-Abud, N.; Zamora-Bustillos, R.; Conesa, A.; Zapata-Perez, O.; Moreno, D.; Hernandez-Nuñez, E. Transcriptome analysis reveals novel insights into the response of low-dose benzo[a]pyrene exposure in male tilapia. Aquat. Toxicol. 2018, 201, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Onoda, Y.; Tachikawa, C.; Hosoi, S.; Yoshita, M.; Chung, S.W.; Kizu, R.; Toriba, A.; Kameda, T.; Tang, N. Estrogenic/antiestrogenic activities of polycyclic aromatic hydrocarbons and their monohydroxylated derivatives by yeast two-hybrid assay. J. Health Sci. 2007, 53, 562–570. [Google Scholar] [CrossRef]
- Ebright, R.H.; Wong, J.R.; Chen, L.B. Binding of 2-hydroxybenzo[a]pyrene to estrogen receptors in rat cytosol. Cancer Res. 1986, 46, 2349–2351. [Google Scholar]
- Charles, G.D.; Bartels, M.J.; Zacharewski, T.R.; Gollapudi, B.B.; Freshour, N.L.; Carney, E.W. Activity of benzo[a]pyrene and its hydroxylated metabolites in an estrogen receptor-α reporter gene assay. Toxicol. Sci. 2000, 55, 320–326. [Google Scholar] [CrossRef]
- Jaruchotikamol, A.; Jarukamjorn, K.; Sirisangtrakul, W.; Sakuma, T.; Kawasaki, Y.; Nemoto, N. Strong synergistic induction of CYP1A1 expression by andrographolide plus typical CYP1A inducers in mouse hepatocytes. Toxicol. Appl. Pharmacol. 2007, 224, 156–162. [Google Scholar] [CrossRef]
- Suzuki, N.; Ikari, T.; Sato, M.; Toriba, A.; Sekiguchi, T.; Kitani, Y.; Ogiso, S.; Yachiguchi, K.; Hattori, A.; Oshima, Y.; et al. Chapter 19. Toxicities of polycyclic aromatic hydrocarbons in fish and marine invertebrates. In Polycyclic Aromatic Hydrocarbons: Environmental Behavior and Toxicity in East Asia; Hayakawa, K., Ed.; Springer: Singapore, 2018; pp. 245–259. ISBN 978-981-10-6775-4. [Google Scholar] [CrossRef]
- Brown, D.R.; Clark, B.W.; Garner, L.V.T.; Di Giulio, R.T. Zebrafish cardiotoxicity: The effects of CYP1A inhibition and AHR2 knockdown following exposure to weak aryl hydrocarbon receptor agonists. Environ. Sci. Pollut. Res. Int. 2015, 22, 8329–8338. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Yu, X.; Cai, L.; Wang, J.; Peng, J. Microplastics and associated PAHs in surface water from the Feilaixia Reservoir in the Beijiang River, China. Chemosphere 2019, 221, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Batel, A.; Borchert, F.; Reinwald, H.; Erdinger, L.; Braunbeck, T. Microplastic accumulation patterns and transfer of benzo[a]pyrene to adult zebrafish (Danio rerio) gills and zebrafish embryos. Environ. Pollut. 2018, 235, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Guven, O.; Bach, L.; Munk, P.; Dinh, K.V.; Mariani, P.; Nielsen, T.G. Microplastic does not magnify the acute effect of PAH pyrene on predatory performance of a tropical fish (Lates calcarifer). Aquat. Toxicol. 2018, 198, 287–293. [Google Scholar] [CrossRef]
- Sese, B.; Grant, A.; Reid, B.J. Toxicity of polycyclic aromatic hydrocarbons to the nematode Caenorhabditis elegans. J. Toxicol. Environ. Health Part A 2009, 72, 1–13. [Google Scholar] [CrossRef]
- Atienzar, F.A.; Conradi, M.; Evenden, A.J.; Jha, A.N.; Depledge, M.H. Qualitative assessment of genotoxicity using random amplified polymorphic DNA: Comparison of genomic template stability with key fitness parameters in Daphnia magna exposed to benzo[a]pyrene. Environ. Toxicol. Chem. 1999, 18, 2275–2282. [Google Scholar] [CrossRef]
- Kagan, J.; Kagan, E.D.; Kagan, I.A.; Kagan, P.A.; Quigley, S. The phototoxicity of non-carcinogenic polycyclic aromatic hydrocarbons in aquatic organisms. Chemosphere 1985, 14, 1829–1834. [Google Scholar] [CrossRef]
- Eastmond, D.A.; Booth, G.M.; Lee, M.I. Toxicity, accumulation, and elimination of polycyclic aromatic sulfur heterocycles in Daphnia magna. Arch. Environ. Contam. Toxicol. 1984, 13, 105–111. [Google Scholar] [CrossRef]
- LeBlanc, G.A. Acute toxicity of priority pollutants to water flea (Daphnia magna). Bull. Environ. Contam. Toxicol. 1980, 24, 684–691. [Google Scholar] [CrossRef]
- Suedel, B.C.; Rodgers, J.H., Jr. Toxicity of fluoranthene to Daphnia magna, Hyalella azteca, Chironomus tentans, and Stylaria lacustris in water-only and whole sediment exposures. Bull. Environ. Contam. Toxicol. 1996, 57, 132–138. [Google Scholar] [CrossRef]
- Millemann, R.E.; Birge, W.J.; Black, J.A.; Cushman, R.M.; Daniels, K.L.; Franco, P.J.; Giddings, J.M. Comparative acute toxicity to aquatic organisms of components of coal-derived synthetic fuels. Trans. Am. Fish. Soc. 1984, 113, 74–85. [Google Scholar] [CrossRef]
- Nam, T.H.; Jeon, H.J.; Mo, H.H.; Cho, K.; Ok, Y.S.; Lee, S.E. Determination of biomarkers for polycyclic aromatic hydrocarbons (PAHs) toxicity to earthworm (Eisenia fetida). Environ. Geochem. Health. 2015, 37, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Dinnel, P.A.; Stober, Q.J.; DiJulio, D.H. Sea urchin sperm bioassay for sewage and chlorinated seawater and its relation to fish bioassays. Mar. Environ. Res. 1981, 5, 29–39. [Google Scholar] [CrossRef]
- Kobayashi, N. Marine pollution bioassay by sea urchin eggs, an attempt to enhance sensitivity. Public Seto Mar. Biol. Lab. 1990, 34, 225–237. [Google Scholar] [CrossRef][Green Version]
- Beiras, R.; Vazquez, E.; Bellas, J.; Lorenzo, J.; Fernandez, N.; Macho, G.; Marino, J.; Casas, L. Sea-urchin embryo bioassay for in situ evaluation of the biological quality of coastal seawater. Estuar. Coast. Shelf Sci. 2001, 52, 29–32. [Google Scholar] [CrossRef]
- Bellas, J.; Nieto, Ó.; Beiras, R. Integrative assessment of coastal pollution: Development and evaluation of sediment quality criteria from chemical contamination and ecotoxicological data. Cont. Shelf Res. 2011, 31, 448–456. [Google Scholar] [CrossRef]
- Alexander, F.J.; King, C.K.; Reichelt-Brushett, A.J.; Harrison, P.L. Fuel oil and dispersant toxicity to the Antarctic sea urchin (Sterechinus neumayeri). Environ. Toxicol. Chem. 2017, 36, 1563–1571. [Google Scholar] [CrossRef]
- Meador, J.P.; Stein, J.E.; Reichert, W.L.; Varanasi, U. Bioaccumulation of polycyclic aromatic hydrocarbons by marine organisms. Rev. Environ. Contam. Toxicol. 1995, 143, 79–165. [Google Scholar] [CrossRef]
- Suzuki, N.; Ogiso, S.; Yachiguchi, K.; Kawabe, K.; Makino, F.; Toriba, A.; Kiyomoto, M.; Sekiguchi, T.; Tabuchi, Y.; Kondo, T.; et al. Monohydroxylated polycyclic aromatic hydrocarbons influence spicule formation in the early development of sea urchins (Hemicentrotus pulcherrimus). Comp. Biochem. Physiol. Part C 2015, 171, 55–60. [Google Scholar] [CrossRef]
- Sekiguchi, T.; Yachiguchi, K.; Kiyomoto, M.; Ogiso, S.; Wada, S.; Tabuchi, Y.; Hong, C.S.; Srivastav, A.K.; Archer, S.D.J.; Pointing, S.B.; et al. Molecular mechanism of the suppression of larval skeleton by polycyclic aromatic hydrocarbons in early development of sea urchin Hemicentrotus pulcherrimus. Fish. Sci. 2018, 84, 1073–1079. [Google Scholar] [CrossRef]
- Bandowe, B.A.M.; Bigalke, M.; Boamah, L.; Nyarko, E.; Saalia, F.K.; Wilcke, W. Polycyclic aromatic compounds (PAHs and oxygenated PAHs) and trace metals in fish species from Ghana (West Africa): Bioaccumulation and health risk assessment. Environ. Int. 2014, 65, 135–146. [Google Scholar] [CrossRef]
- Dsikowitzky, L.; Nordhaus, I.; Andarwulan, N.; Irianto, H.E.; Lioe, H.N.; Ariyani, F.; Kleinertz, S.; Schwarzbauer, J. Accumulation patterns of lipophilic organic contaminants in surface sediments and in economic important mussel and fish species from Jakarta Bay, Indonesia. Mar. Pollut. Bull. 2016, 110, 767–777. [Google Scholar] [CrossRef]
- Gewurtz, S.B.; Lazar, R.; Haffner, G.D. Comparison of polycyclic aromatic hydrocarbon and polychlorinated biphenyl dynamics in benthic invertebrates of Lake Erie, USA. Environ. Toxicol. Chem. Int. J. 2000, 19, 2943–2950. [Google Scholar] [CrossRef]
- Poliakova, O.V.; Lebedev, A.T.; Petrosyan, V.S.; Hanninen, O.; Renzoni, A.; Sawa, D.; Walker, C. Accumulation of persistent organic pollutants in the food chain of Lake Baikal. Toxicol. Environ. Chem. 2000, 75, 235–243. [Google Scholar] [CrossRef]
- Wan, Y.; Jin, X.; Hu, J.; Jin, F. Trophic dilution of polycyclic aromatic hydrocarbons (PAHs) in a marine food web from Bohai Bay, North China. Environ. Sci. Technol. 2007, 41, 3109–3114. [Google Scholar] [CrossRef] [PubMed]
- Cheikyula, J.O.; Koyama, J.; Uno, S. Comparative study of bioconcentration and EROD activity induction in the Japanese flounder, red sea bream, and Java medaka exposed to polycyclic aromatic hydrocarbons. Environ. Toxicol. Int. J. 2008, 23, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Jafarabadi, A.R.; Bakhtiari, A.R.; Yaghoobi, Z.; Yap, C.K.; Maisano, M.; Cappello, T. Distributions and compositional patterns of polycyclic aromatic hydrocarbons (PAHs) and their derivatives in three edible fishes from Kharg coral Island, Persian Gulf, Iran. Chemosphere 2019, 215, 835–845. [Google Scholar] [CrossRef]
- Oliva, A.L.; La Colla, N.S.; Arias, A.H.; Blasina, G.E.; Cazorla, A.L.; Marcovecchio, J.E. Distribution and human health risk assessment of PAHs in four fish species from a SW Atlantic estuary. Environ. Sci. Pollut. Res. 2017, 24, 18979–18990. [Google Scholar] [CrossRef]
- Logan, D.T. Perspective on ecotoxicology of PAHs to fish. Hum. Ecol. Risk Assess. 2007, 13, 302–316. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, L.; Cai, Y.; Chen, Y. Distribution of polycyclic aromatic hydrocarbon (PAH) residues in several tissues of edible fishes from the largest freshwater lake in China, Poyang Lake, and associated human health risk assessment. Ecotoxicol. Environ. Saf. 2014, 104, 323–331. [Google Scholar] [CrossRef]
- Kaag, N.H.; Foekema, E.M.; Scholten, M.C.T.; van Straalen, N.M. Comparison of contaminant accumulation in three species of marine invertebrates with different feeding habits. Environ. Toxicol. Chem. Int. J. 1997, 16, 837–842. [Google Scholar] [CrossRef]
- Monikh, F.A.; Hosseini, M.; Rahmanpour, S. The effect of size and sex on PCB and PAH concentrations in crab Portunus pelagicus. Environ. Monit. Assess. 2014, 186, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Baumard, P.; Budzinski, H.; Garrigues, P. Polycyclic aromatic hydrocarbons in sediments and mussels of the western Mediterranean Sea. Environ. Toxicol. Chem. Int. J. 1998, 17, 765–776. [Google Scholar] [CrossRef]
- Cheung, K.C.; Leung, H.M.; Kong, K.Y.; Wong, M.H. Residual levels of DDTs and PAHs in freshwater and marine fish from Hong Kong markets and their health risk assessment. Chemosphere 2007, 66, 460–468. [Google Scholar] [CrossRef]
- Grung, M.; Petersen, K.; Fjeld, E.; Allan, I.; Christensen, J.H.; Malmqvist, L.M.; Meland, S.; Ranneklev, S. PAH related effects on fish in sedimentation ponds for road runoff and potential transfer of PAHs from sediment to biota. Sci. Total Environ. 2016, 566, 1309–1317. [Google Scholar] [CrossRef]
- Huang, L.; Chernyak, S.M.; Batterman, S.A. PAHs, nitro-PAHs, hopanes, and steranes in lake trout from Lake Michigan. Environ. Toxicol. Chem. 2014, 33, 1792–1801. [Google Scholar] [CrossRef]
- Moon, H.B.; An, Y.R.; Choi, S.G.; Choi, M.; Choi, H.G. Accumulation of PAHs and synthetic musk compound in minke whales (Balanoptera acutorostrata) and long-beaked common dolphins (Delphinus capensis) from Korean coastal waters. Environ. Toxicol. Chem. 2012, 31, 477–485. [Google Scholar] [CrossRef]
- Net, S.; Henry, F.; Rabodonirina, S.; Diop, M.; Merhaby, D.; Mahfouz, C.; Amara, R.; Ouddane, B. Accumulation of PAHs, Me-PAHs, PCBs and total mercury in sediments and marine species in coastal areas of Dakar, Senegal: Contamination level and impact. Int. J. Environ. Res. 2015, 9, 419–432. [Google Scholar] [CrossRef]
- Macek, K.J.; Rodgers, C.R.; Stalling, D.L.; Korn, S. The uptake, distribution and elimination of dietary 14C-DDT and 14C-dieldrin in rainbow trout. Trans. Am. Fish. Soc. 1970, 99, 689–695. [Google Scholar] [CrossRef]
- Zhang, G.; Pan, Z.; Wang, X.; Mo, X.; Li, X. Distribution and accumulation of polycyclic aromatic hydrocarbons (PAHs) in the food web of Nansi Lake, China. Environ. Monit. Assess. 2015, 187, 173. [Google Scholar] [CrossRef]
- Baumann, P.C.; Mac, M.J.; Smith, S.B.; Harshbarger, J.C. Tumor frequencies in walleye (Stizostedion vitreum) and brown bullhead (Ictalurus nebulosus) and sediment contaminants in tributaries of the Laurentian Great Lakes. Can. J. Fish. Aquat. Sci. 1991, 48, 1804–1810. [Google Scholar] [CrossRef]
- Eadie, B.J.; Landrum, P.F.; Faust, W. Polycyclic aromatic hydrocarbons in sediments, pore water and the amphipod Pontoporeia hoyi from Lake Michigan. Chemosphere 1982, 11, 847–858. [Google Scholar] [CrossRef]
- Levengood, J.M.; Schaeffer, D.J. Polycyclic aromatic hydrocarbons in fish and crayfish from the Calumet region of southwestern Lake Michigan. Ecotoxicology 2011, 20, 1411–1421. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, C.D.; Metcalfe, T.L.; Riddle, G.; Haffner, G.D. Aromatic hydrocarbons in biota from the Detroit River and western Lake Erie. J. Great Lakes Res. 1997, 23, 160–168. [Google Scholar] [CrossRef]
- Ridgway, L.L.; Chapleau, F.; Comba, M.E.; Backus, S.M. Population characteristics and contaminant burdens of the white sucker (Catostomus commersoni) from the St. Lawrence River near Cornwall, Ontario and Massena, New York. J. Great Lakes Res. 1999, 25, 567–582. [Google Scholar] [CrossRef]
- Zabik, M.E.; Booren, A.; Zabik, M.J.; Welch, R.; Humphrey, H. Pesticide residues, PCBs and PAHs in baked, charbroiled, salt boiled and smoked Great Lakes lake trout. Food Chem. 1996, 55, 231–239. [Google Scholar] [CrossRef]
- Landrum, P.F.; Fisher, S.W. Influence of lipids on the bioaccumulation and trophic transfer of organic contaminants in aquatic organisms. In Lipids in Freshwater Ecosystems; Arts, M.T., Wainman, B.C., Eds.; Springer: New York, NY, USA, 1999; pp. 203–234. ISBN 978-1-4612-0547-0. [Google Scholar] [CrossRef]
- Liang, Y.; Tse, M.F.; Young, L.; Wong, M.H. Distribution patterns of polycyclic aromatic hydrocarbons (PAHs) in the sediments and fish at Mai Po Marshes Nature Reserve, Hong Kong. Water Res. 2007, 41, 1303–1311. [Google Scholar] [CrossRef]
- Mashroofeh, A.; Bakhtiari, A.R.; Pourkazemi, M. Distribution and composition pattern of polycyclic aromatic hydrocarbons in different tissues of sturgeons collected from Iranian coastline of the Caspian Sea. Chemosphere 2015, 120, 575–583. [Google Scholar] [CrossRef]
- Yu, Z.; Lin, Q.; Gu, Y.; Du, F.; Wang, X.; Shi, F.; Ke, C.; Xiang, M.; Yu, Y. Bioaccumulation of polycyclic aromatic hydrocarbons (PAHs) in wild marine fish from the coastal waters of the northern South China Sea: Risk assessment for human health. Ecotoxicol. Environ. Saf. 2019, 180, 742–748. [Google Scholar] [CrossRef]
- Soltani, N.; Moore, F.; Keshavarzi, B.; Sorooshian, A.; Javid, R. Potentially toxic elements (PTEs) and polycyclic aromatic hydrocarbons (PAHs) in fish and prawn in the Persian Gulf, Iran. Ecotoxicol. Environ. Saf. 2019, 173, 251–265. [Google Scholar] [CrossRef]
- Beyer, J.; Jonsson, G.; Porte, C.; Krahn, M.M.; Ariese, F. Analytical methods for determining metabolites of polycyclic aromatic hydrocarbon (PAH) pollutants in fish bile: A review. Environ. Toxicol. Pharmacol. 2010, 30, 224–244. [Google Scholar] [CrossRef] [PubMed]
- Tolosa, I.; Bayona, J.M.; Albaigés, J. Aliphatic and polycyclic aromatic hydrocarbons and sulfur/oxygen derivatives in northwestern Mediterranean sediments: Spatial and temporal variability, fluxes, and budgets. Environ. Sci. Technol. 1996, 30, 2495–2503. [Google Scholar] [CrossRef]
- Batista, D.; Tellini, K.; Nudi, A.H.; Massone, T.P.; Scofield, A.D.L.; de LR Wagener, A. Marine sponges as bioindicators of oil and combustion derived PAH in coastal waters. Mar. Environ. Res. 2013, 92, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Bourgeault, A.; Gourlay-Francé, C. Monitoring PAH contamination in water: Comparison of biological and physico-chemical tools. Sci. Total Environ. 2013, 454, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Ghaeni, M.; Pour, N.A.; Hosseini, M. Bioaccumulation of polychlorinated biphenyl (PCB), polycyclic aromatic hydrocarbon (PAH), mercury, methyl mercury, and arsenic in blue crab Portunus segnis from Persian Gulf. Environ. Monit. Assess. 2015, 187, 253. [Google Scholar] [CrossRef]
- Rust, A.J.; Burgess, R.M.; Brownawell, B.J.; McElroy, A.E. Relationship between metabolism and bioaccumulation of benzo [α] pyrene in benthic invertebrates. Environ. Toxicol. Chem. Int. J. 2004, 23, 2587–2593. [Google Scholar] [CrossRef]
- Chandurvelan, R.; Marsden, I.D.; Glover, C.N.; Gaw, S. Assessment of a mussel as a metal bioindicator of coastal contamination: Relationships between metal bioaccumulation and multiple biomarker responses. Sci. Total Environ. 2015, 511, 663–675. [Google Scholar] [CrossRef]
- Monirith, I.; Ueno, D.; Takahashi, S.; Nakata, H.; Sudaryanto, A.; Subramanian, A. Asia-Pacific mussel watch: Monitoring contamination of persistent organochlorine compounds in coastal waters of Asian countries. Mar. Pollut. Bull. 2003, 46, 281–300. [Google Scholar] [CrossRef]
- Bouzas, A.; Aguado, D.; Martí, N.; Pastor, J.M.; Herráez, R.; Campins, P.; Seco, A. Alkylphenols and polycyclic aromatic hydrocarbons in eastern Mediterranean Spanish coastal marine bivalves. Environ. Monit. Assess. 2011, 176, 169–181. [Google Scholar] [CrossRef]
- Eduok, S.I.; Ebong, G.A.; Udoinyang, E.P.; Njoku, J.N.; Eyen, E.A. Bacteriological and polycyclic aromatic hydrocarbon accumulation in mangrove oyster (Crassostrea tulipa) from Douglas Creek, Nigeria. Pak. J. Nutr. 2010, 9, 35–42. [Google Scholar] [CrossRef][Green Version]
- Ito, K.; Uno, S.; Tanaka, H.; Miki, S.; Kokushi, E.; Yamamoto, M.; Koyama, J. Distribution of parent and alkylated PAHs in bivalves collected from Osaka Bay, Japan. Jpn. J. Environ. Toxicol. 2015, 18, 11–24. [Google Scholar] [CrossRef]
- Ramdine, G.; Fichet, D.; Louis, M.; Lemoine, S. Polycyclic aromatic hydrocarbons (PAHs) in surface sediment and oysters (Crassostrea rhizophorae) from mangrove of Guadeloupe: Levels, bioavailability, and effects. Ecotoxicol. Environ. Saf. 2012, 79, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, Y.; Suarez, P.; Alonso, A.; Longo, E.; Villaverde, A.; San Juan, F. Environmental quality of mussel farms in the Vigo estuary: Pollution by PAHs, origin and effects on reproduction. Environ. Pollut. 2011, 159, 250–265. [Google Scholar] [CrossRef] [PubMed]
- Uno, S.; Tanaka, H.; Miki, S.; Kokushi, E.; Ito, K.; Yamamoto, M.; Koyama, J. Bioaccumulation of nitroarenes in bivalves at Osaka Bay, Japan. Mar. Pollut. Bull. 2011, 63, 477–481. [Google Scholar] [CrossRef]
- Tanaka, H.; Onduka, T. Background Levels of PAHs in the coastal waters of Japan based on residual concentrations of bivalves. J. Environ. Chem. 2010, 20, 137–148. [Google Scholar] [CrossRef][Green Version]
- Beyer, J.; Trannum, H.C.; Bakke, T.; Hodson, P.V.; Collier, T.K. Environmental effects of the Deepwater Horizon oil spill: A review. Mar. Pollut. Bull. 2016, 110, 28–51. [Google Scholar] [CrossRef]
- Hickey, C.W.; Roper, D.S.; Holland, P.T.; Trower, T.M. Accumulation of organic contaminants in two sediment-dwelling shellfish with contrasting feeding modes: Deposit-(Macomona liliana) and filter-feeding (Austrovenus stutchburyi). Arch. Environ. Contam. Toxicol. 1995, 29, 221–231. [Google Scholar] [CrossRef]
- Meador, J.P.; Casillas, E.; Sloan, C.A.; Varanasi, U. Comparative bioaccumulation of polycyclic aromatic hydrocarbons from sediment by two infaunal invertebrates. Mar. Ecol. Prog. Ser. 1995, 123, 107–124. [Google Scholar] [CrossRef]
- Ikenaka, Y.; Ito, Y.; Eun, H.; Watanabe, E.; Miyabara, Y. Characteristics of accumulation patterns of polycyclic aromatic hydrocarbons in the organisms inhabited in Lake Suwa. J. Environ. Chem. 2008, 18, 341–352. [Google Scholar] [CrossRef]
- Thomann, R.V.; Komlos, J. Model of biota-sediment accumulation factor for polycyclic aromatic hydrocarbons. Environ. Toxicol. Chem. Int. J. 1999, 18, 1060–1068. [Google Scholar] [CrossRef]
- Incardona, J.P.; Collier, T.K.; Scholz, N.L. Defects in cardiac function precede morphological abnormalities in fish embryos exposed to polycyclic aromatic hydrocarbons. Toxicol. Appl. Pharmacol. 2004, 196, 191–205. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).