Exogenous Glutathione Alleviation of Cd Toxicity in Italian Ryegrass (Lolium multiflorum) by Modulation of the Cd Absorption, Subcellular Distribution, and Chemical Form


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Experiment Design
2.3. Sampling and Subcellular Fractions Fractionation
2.4. Extraction of Cd in Different Chemical Forms
2.5. Cd Concentration, Translocation Factor (TF), and Tolerance Index (TI)
2.6. Statistical Analysis
3. Results
3.1. Effect of GSH on the Growth of Italian Ryegrass Seedlings under Cd Stress
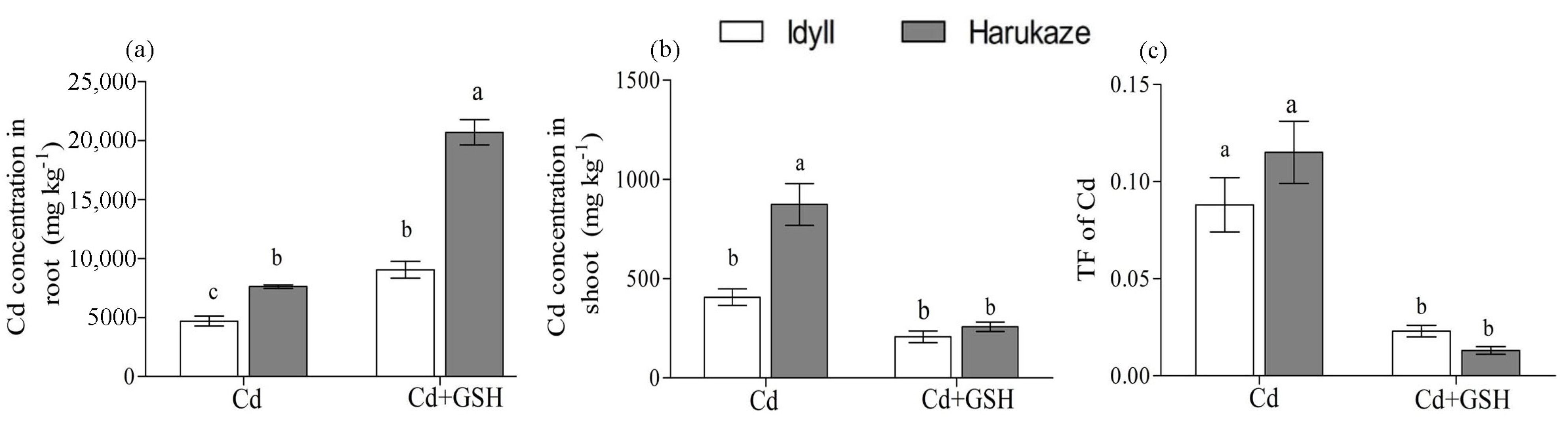
3.2. Effect of GSH on Cd Uptake and Translocation in Italian Ryegrass Seedlings
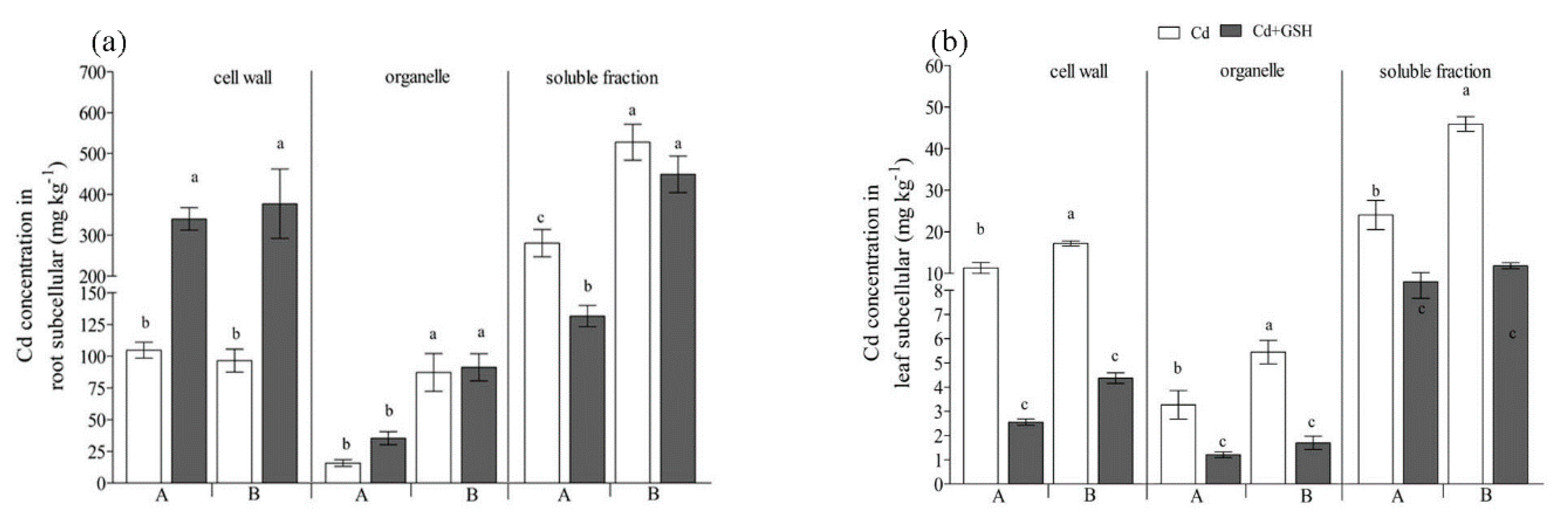
3.3. Effect of GSH on Subcellular Distribution of Cd in Italian Ryegrass Roots and Shoots
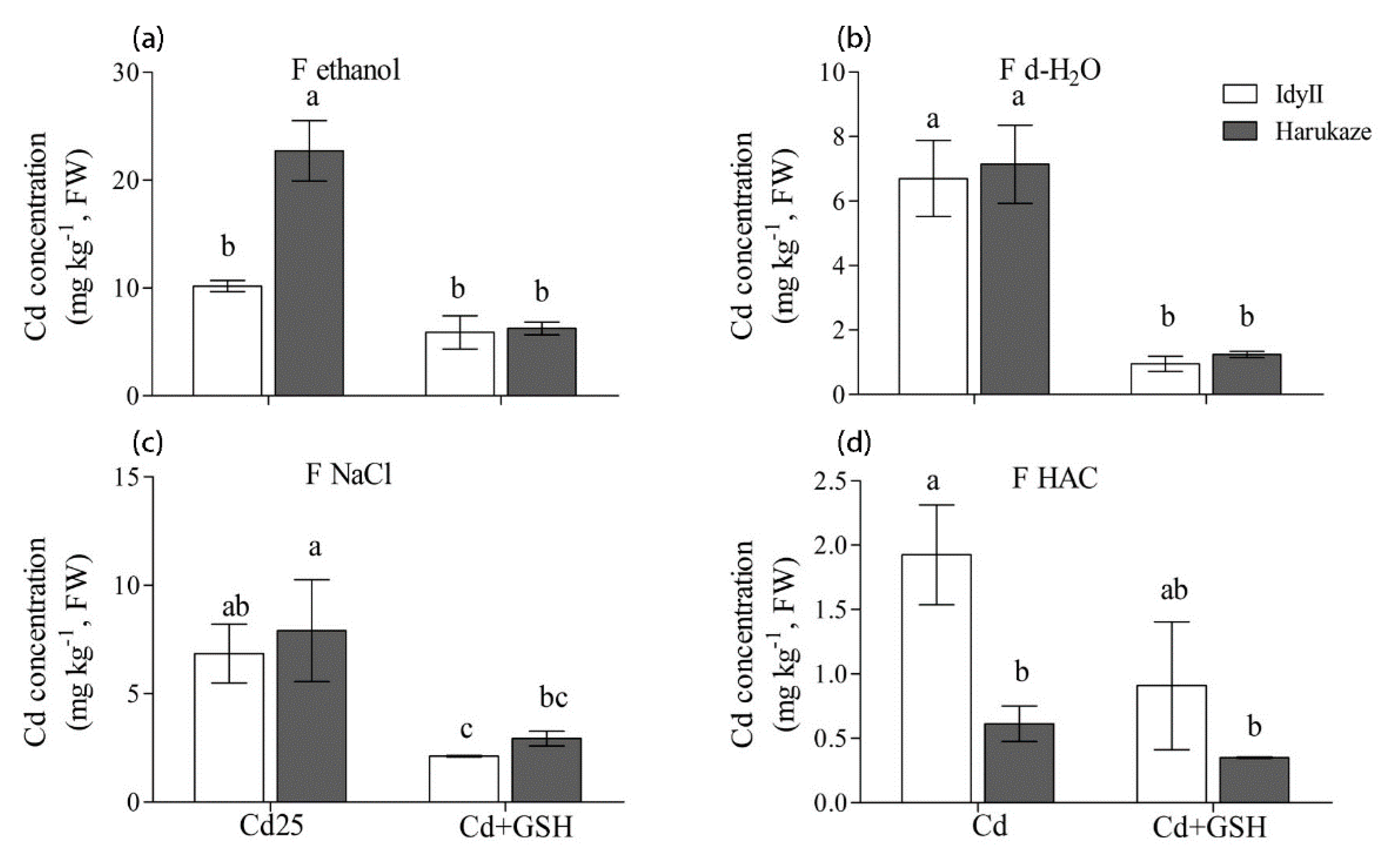
3.4. Effect of GSH on Chemical Forms of Cd in Italian Ryegrass Roots and Shoots
3.5. Correlations Analysis among TI, the Proportion of Cd in the Root Cell Wall and the Proportion of the Cd Form in the Roots
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Guan, M.; Zhang, H.; Pan, W.; Jin, C.; Lin, X. Sulfide alleviates cadmium toxicity in Arabidopsis plants by altering the chemical form and the subcellular distribution of cadmium. Sci. Total. Environ. 2018, 627, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Metwally, A.; Safronova, V.I.; Belimov, A.A.; Dietz, K.J. Genotypic variation of the response to cadmium toxicity in Pisum sativum L. J. Exp. Bot. 2005, 56, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Azzi, V.S.; Kanso, A.; Kobeissi, A.; Kazpard, V.; Lartiges, B.; El Samrani, A. Effect of Cadmium on Lactuca sativa Grown in Hydroponic Culture Enriched with Phosphate Fertilizer. J. Environ. Prot. 2015, 6, 1337–1346. [Google Scholar] [CrossRef]
- Bai, X.; Dong, Y.; Kong, J.; Xu, L.; Liu, S. Effects of application of salicylic acid alleviates cadmium toxicity in perennial ryegrass. Plant Growth Regul. 2014, 75, 695–706. [Google Scholar] [CrossRef]
- Zhao, H.; Jin, Q.; Wang, Y.; Chu, L.; Li, X.; Xu, Y. Effects of nitric oxide on alleviating cadmium stress in Typha angustifolia. Plant Growth Regul. 2016, 78, 243–251. [Google Scholar] [CrossRef]
- Shi, G.; Zhang, Z.; Liu, C. Silicon influences cadmium translocation by altering subcellular distribution and chemical forms of cadmium in peanut roots. Archi. Agron. Soil Sci. 2017, 63, 117–123. [Google Scholar] [CrossRef]
- Zhao, L.; Li, T.; Yu, H.; Chen, G.; Zhang, X.; Zheng, Z.; Li, J. Changes in chemical forms, subcellular distribution, and thiol compounds involved in Pb accumulation and detoxification in Athyrium wardii (Hook.). Environ. Sci. Pollut. Res. Int. 2015, 22, 12676–12688. [Google Scholar] [CrossRef]
- Xiao, Y.T.; Du, Z.J.; Busso, C.A.; Qi, X.B.; Wu, H.Q.; Guo, W.; Wu, D.F. Differences in root surface adsorption, root uptake, subcellular distribution, and chemical forms of Cd between low-and high-Cd-accumulating wheat cultivars. Environ. Sci. Pollut. Res. Int. 2020, 27, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Bora, M.S.; Gogoi, N.; Sarma, K.P. Tolerance mechanism of cadmium in Ceratopteris pteridoides: Translocation and subcellular distribution. Ecotoxicol. Environ. Saf. 2020, 197, 110599. [Google Scholar] [CrossRef]
- Ramos, I.; Esteban, E.; Lucena, J.J.; Gárate, A.N. Cadmium uptake and subcellular distribution in plants of Lactuca sp. Cd–Mn interaction. Plant Sci. 2002, 162, 761–767. [Google Scholar] [CrossRef]
- Hu, P.-J.; Qiu, R.-L.; Senthilkumar, P.; Jiang, D.; Chen, Z.W.; Tang, Y.-T.; Liu, F.J. Tolerance, accumulation and distribution of zinc and cadmium in hyperaccumulator Potentilla griffithii. Environ. Exp. Bot. 2009, 66, 317–325. [Google Scholar] [CrossRef]
- Lam, C.M.; Chen, K.S.; Lai, H.Y. Chemical Forms and Health Risk of Cadmium in Water Spinach Grown in Contaminated Soil with an Increased Level of Phosphorus. Int. J. Environ. Res. Public Health 2019, 16, 3322. [Google Scholar] [CrossRef]
- Fu, X.; Dou, C.; Chen, Y.; Chen, X.; Shi, J.; Yu, M.; Xu, J. Subcellular distribution and chemical forms of cadmium in Phytolacca americana L. J. Hazard. Mate. 2011, 186, 103–107. [Google Scholar] [CrossRef]
- Zhao, Y.; Wu, J.; Shang, D.; Ning, J.; Zhai, Y.; Sheng, X.; Ding, H. Subcellular distribution and chemical forms of cadmium in the edible seaweed, Porphyra yezoensis. Food Chem. 2015, 168, 48–54. [Google Scholar] [CrossRef]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione is a key player in metal-induced oxidative stress defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef]
- Hasan, M.K.; Liu, C.; Wang, F.; Ahammed, G.J.; Zhou, J.; Xu, M.X.; Yu, J.Q.; Xia, X.J. Glutathione-mediated regulation of nitric oxide, S-nitrosothiol and redox homeostasis confers cadmium tolerance by inducing transcription factors and stress response genes in tomato. Chemosphere 2016, 161, 536–545. [Google Scholar] [CrossRef]
- Jacquart, A.; Brayner, R.; El Hage Chahine, J.M.; Ha-Duong, N.T. Cd2+ and Pb2+ complexation by glutathione and the phytochelatins. Chem. Biol. Interact. 2017, 267, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Wojas, S.; Ruszczynska, A.; Bulska, E.; Clemens, S.; Antosiewicz, D.M. The role of subcellular distribution of cadmium and phytochelatins in the generation of distinct phenotypes of AtPCS1- and CePCS3-expressing tobacco. J. Plant Physiol. 2010, 167, 981–988. [Google Scholar]
- Wang, F.; Chen, F.; Cai, Y.; Zhang, G.; Wu, F. Modulation of exogenous glutathione in ultrastructure and photosynthetic performance against Cd stress in the two barley genotypes differing in Cd tolerance. Biol. Trace. Elem. Res. 2011, 144, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Daud, M.K.; Mei, L.; Azizullah, A.; Dawood, M.; Ali, I.; Mahmood, Q.; Ullah, W.; Jamil, M.; Zhu, S.J. Leaf-based physiological, metabolic, and ultrastructural changes in cultivated cotton cultivars under cadmium stress mediated by glutathione. Environ. Sci. Pollut. Res. Int. 2016, 23, 15551–15564. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhu, Z.; Wu, X.; Liu, Z.; Zou, J.; Chen, Y.; Su, N.; Cui, J. Lower cadmium accumulation and higher antioxidative capacity in edible parts of Brassica campestris L. seedlings applied with glutathione under cadmium toxicity. Environ. Sci. Pollut. Res. Int. 2019, 26, 13235–13245. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, J.; Lu, Z.; Wang, X.; Zhang, Z.; Shi, G. Effects of iron deficiency on subcellular distribution and chemical forms of cadmium in peanut roots in relation to its translocation. Environ. Exp. Bot. 2014, 97, 40–48. [Google Scholar] [CrossRef]
- Ye, S.; Yang, Y.; Xin, G.; Wang, Y.; Ruan, L.; Ye, G. Studies of the Italian ryegrass–rice rotation system in southern China: Arbuscular mycorrhizal symbiosis affects soil microorganisms and enzyme activities in the Lolium mutiflorum L. rhizosphere. Appl. Soil Ecol. 2015, 90, 26–34. [Google Scholar] [CrossRef]
- Mugica-Alvarez, V.; Cortés-Jiménez, V.; Vaca-Mier, M.; Domínguez-Soria, V. Phytoremediation of Mine Tailings Using Lolium Multiflorum. Int. J. Environ. Sci. Dev. 2015, 6, 246–251. [Google Scholar] [CrossRef]
- Yasuda, M.; Takenouchi, Y.; Nitta, Y.; Ishii, Y.; Ohta, K. Italian ryegrass (Lolium multiflorum Lam.) as a High-Potential Bio-Ethanol Resource. BioEnergy Res. 2015, 8, 1303–1309. [Google Scholar] [CrossRef]
- El Hamiani, O.; El Khalil, H.; Lounate, K.; Sirguey, C.; Hafidi, M.; Bitton, G.; Schwartz, C.; Boularbah, A. Toxicity assessment of garden soils in the vicinity of mining areas in Southern Morocco. J. Hazard Mater. 2010, 177, 755–761. [Google Scholar] [CrossRef]
- Fang, Z.; Lou, L.; Tai, Z.; Wang, Y.; Yang, L.; Hu, Z.; Cai, Q. Comparative study of Cd uptake and tolerance of two Italian ryegrass (Lolium multiflorum) cultivars. PeerJ 2017, 5, e3621. [Google Scholar] [CrossRef]
- Qiu, B.; Zeng, F.; Cai, S.; Wu, X.; Haider, S.I.; Wu, F.; Zhang, G. Alleviation of chromium toxicity in rice seedlings by applying exogenous glutathione. J. Plant Physiol. 2013, 170, 772–779. [Google Scholar] [CrossRef]
- Weigel, H.J.; JaGer, H.J. Subcellular distribution and chemical form of cadmium in bean plants. Plant Physiol. 1980, 65, 480–482. [Google Scholar] [CrossRef]
- Mwamba, T.M.; Li, L.; Gill, R.A.; Islam, F.; Nawaz, A.; Ali, B.; Farooq, M.A.; Lwalaba, J.L.; Zhou, W. Differential subcellular distribution and chemical forms of cadmium and copper in Brassica napus. Ecotoxicol. Environ. Saf. 2016, 134, 239–249. [Google Scholar] [CrossRef]
- Chen, L.; Long, X.-H.; Zhang, Z.-H.; Zheng, X.-T.; Rengel, Z.; Liu, Z.-P. Cadmium Accumulation and Translocation in Two Jerusalem Artichoke (Helianthus tuberosus L.) Cultivars. Pedosphere 2011, 21, 573–580. [Google Scholar] [CrossRef]
- Shi, G.; Liu, C.; Cui, M.; Ma, Y.; Cai, Q. Cadmium tolerance and bioaccumulation of 18 hemp accessions. Appl. Biochem. Biotechnol. 2012, 168, 163–173. [Google Scholar] [CrossRef]
- Henson, T.M.; Cory, W.; Rutter, M.T. Extensive variation in cadmium tolerance and accumulation among populations of Chamaecrista fasciculata. PLoS ONE 2013, 8, e63200. [Google Scholar] [CrossRef]
- Fang, Z.; Hu, Z.; Zhao, H.; Yang, L.; Ding, C.; Lou, L.; Cai, Q. Screening for cadmium tolerance of 21 cultivars from Italian ryegrass (Lolium multiflorum Lam) during germination. Grassl. Sci. 2017, 63, 36–45. [Google Scholar] [CrossRef]
- Cai, Y.; Cao, F.; Wei, K.; Zhang, G.; Wu, F. Genotypic dependent effect of exogenous glutathione on Cd-induced changes in proteins, ultrastructure and antioxidant defense enzymes in rice seedlings. J. Hazard. Mate. 2011, 192, 1056–1066. [Google Scholar] [CrossRef]
- Ding, S.; Ma, C.; Shi, W.; Liu, W.; Lu, Y.; Liu, Q.; Luo, Z.B. Exogenous glutathione enhances cadmium accumulation and alleviates its toxicity in Populus x canescens. Tree Physiol. 2017, 37, 1697–1712. [Google Scholar] [CrossRef]
- Wang, J.; Huang, B.; Chen, X.; Shi, Y. Alleviation of cadmium toxicity to Cole (Brassica campestris L. Cruciferae) by exogenous glutathione. IOP Conf. Ser. Earth Environ. Sci. 2017, 61, 012080. [Google Scholar] [CrossRef]
- Tong, B. Low molecular weight organic acids in root exudates and cadmium accumulation in cadmium hyperaccumulator Solanum nigrum L. and non-hyperaccumulator Solanum lycopersicum L. Afr. J. Biotechnol. 2011, 10, 75. [Google Scholar] [CrossRef]
- Fu, H.; Yu, H.; Li, T.; Zhang, X. Influence of cadmium stress on root exudates of high cadmium accumulating rice line (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2018, 150, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Pourghasemian, N.; Ehsanzadeh, P.; Greger, M. Genotypic variation in safflower (Carthamus spp.) cadmium accumulation and tolerance affected by temperature and cadmium levels. Environ. Exp. Bot. 2013, 87, 218–226. [Google Scholar] [CrossRef]
- Yang, J.; Guo, J.; Yang, J. Cadmium accumulation and subcellular distribution in populations of Hylotelephium spectabile (Boreau) H. Ohba. Environ. Sci. Pollut. Res. Int. 2018, 25, 30917–30927. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Deng, X.; Huang, Y.; Fang, X.; Zhang, J.; Wan, H.; Yang, C. Comparison of subcellular distribution and chemical forms of cadmium among four soybean cultivars at young seedlings. Environ. Sci. Pollut. Res. Int. 2015, 22, 19584–19595. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.I.; Suzui, N.; Yin, Y.G.; Ishii, S.; Fujimaki, S.; Kawachi, N.; Rai, H.; Matsumoto, T.; Sato-Izawa, K.; Ohkama-Ohtsu, N. Effects of enhancing endogenous and exogenous glutathione in roots on cadmium movement in Arabidopsis thaliana. Plant Sci. 2020, 290, 110304. [Google Scholar] [CrossRef]
- Nakamura, S.-I.; Suzui, N.; Ito-Tanabata, S.; Ishii, S.; Kawachi, N.; Rai, H.; Hattori, H.; Fujimaki, S. Application of glutathione and dithiothreitol to oil seed rape (Brassica napusL.) roots affects cadmium distribution in roots and inhibits Cd translocation to shoots. Soil Sci. Plant Nutr. 2016, 62, 379–385. [Google Scholar] [CrossRef]
- Son, K.-H.; Kim, D.-Y.; Koo, N.; Kim, K.-R.; Kim, J.-G.; Owens, G. Detoxification through phytochelatin synthesis in Oenothera odorata exposed to Cd solutions. Environ. Exp. Bot. 2012, 75, 9–15. [Google Scholar] [CrossRef]
- Li, T.; Tao, Q.; Shohag, M.J.I.; Yang, X.; Sparks, D.L.; Liang, Y. Root cell wall polysaccharides are involved in cadmium hyperaccumulation in Sedum alfredii. Plant Soil 2014, 389, 387–399. [Google Scholar] [CrossRef]
- Nakamura, S.I.; Wongkaew, A.; Nakai, Y.; Rai, H.; Ohkama-Ohtsu, N. Foliar-applied glutathione activates zinc transport from roots to shoots in oilseed rape. Plant Sci. 2019, 283, 424–434. [Google Scholar] [CrossRef]
- Hao, X.; Li, T.; Yu, H.; Zhang, X.; Zheng, Z.; Chen, G.; Zhang, S.; Zhao, L.; Pu, Y. Cd accumulation and subcellular distribution in two ecotypes of Kyllinga brevifolia Rottb as affected by Cd treatments. Environ. Sci. Pollut. Res. 2015, 22, 7461–7469. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Y.; Zeng, G.; Chai, L.; Song, X.; Min, Z.; Xiao, X. Subcellular distribution and chemical forms of cadmium in Bechmeria nivea (L.) Gaud. Environ. Exp. Bot. 2008, 62, 389–395. [Google Scholar] [CrossRef]
- Zhan, F.; He, Y.; Li, Y.; Li, T.; Yang, Y.Y.; Toor, G.S.; Zhao, Z. Subcellular distribution and chemical forms of cadmium in a dark septate endophyte (DSE), Exophiala pisciphila. Environ. Sci. Pollut. Res. 2015, 22, 17897–17905. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatment | Root DW/mg | Root Tl | Shoot DW/mg | Shoot TI |
|---|---|---|---|---|---|
| IdyII | CK | 22.51 ± 2.77 a | 1.00 | 123.98 ± 10.4 a | 1.00 |
| Cd | 13.30 ± 1.57 c | 0.59 | 80.48 ± 8.41 c | 0.65 | |
| Cd + GSH | 20.54 ± 1.71 ab | 0.91 | 104.42 ± 5.65 ab | 0.84 | |
| GSH | 24.79 ± 1.87 a | 1.10 | 122.38 ± 5.55 a | 0.99 | |
| Harukaze | CK | 15.29 ± 0.99 bc | 1.00 | 84.00 ± 5.05 bc | 1.00 |
| Cd | 5.46 ± 0.58 d | 0.36 | 35.20 ± 3.62 e | 0.42 | |
| Cd + GSH | 13.02 ± 2.43 c | 0.85 | 59.41 ± 6.82 d | 0.71 | |
| GSH | 14.72 ± 1.47 c | 0.96 | 94.83 ± 3.90 bc | 1.12 |
| Cultivar | Treatment | Root | Leaf | ||||
|---|---|---|---|---|---|---|---|
| Cell Wall% | Organelle% | Soluble Fraction% | Cell Wall% | Organelle% | Soluble Fraction% | ||
| IdyII | Cd | 26.28 ± 1.01 c | 4.06 ± 0.93 c | 69.66 ± 1.87 a | 29.50 ± 1.53 a | 8.53 ± 1.31 a | 61.97 ± 2.81 b |
| Cd + GSH | 66.98 ± 2.50 a | 6.94 ± 0.79 bc | 26.08 ± 1.95 c | 20.58 ± 2.53 b | 9.67 ± 1.28 a | 69.75 ± 3.22 a | |
| Harukaze | Cd | 13.56 ± 0.22 d | 12.11 ± 1.38 a | 74.33 ± 1.58 a | 25.14 ± 0.21 ab | 7.92 ± 0.47 a | 66.95 ± 0.26 ab |
| Cd + GSH | 40.21 ± 5.49 b | 10.27 ± 2.04 ab | 49.52 ± 5.06 b | 24.46 ± 0.87 ab | 9.38 ± 0.87 a | 66.16 ± 0.67 ab | |
| Tissue | Cultivar | Treatment | F Ethanol% | F d-H2O% | F NaCl% | F HAC% | F HCl% | F Residue% |
|---|---|---|---|---|---|---|---|---|
| Root | IdyII | Cd50 | 59.25 ± 5.42 a | 14.45 ± 3.03 b | 23.31 ± 1.91 a | 2.79 ± 0.49 c | 0.20 ± 0.04 c | 0 c |
| Cd50 + GSH | 30.96 ± 1.00 b | 22.07 ± 0.95 a | 14.51 ± 1.12 b | 25.36 ± 1.01 b | 7.05 ± 0.45 a | 0.2 ± 0.06 b | ||
| Harukaze | Cd50 | 69.72 ± 2.52 a | 11.59 ± 1.1 b | 16.18 ± 3.92 ab | 1.15 ± 0.26 c | 1.36 ± 0.47 b | 0 c | |
| Cd50 + GSH | 29.69 ± 3.56 b | 17.11 ± 2.25 ab | 15.49 ± 0.44 b | 30.92 ± 1.5 a | 6.65 ± 0.27 a | 0.77 ± 0.13 a | ||
| Leaf | IdyII | Cd50 | 40.84 ± 6.29 a | 25.69 ± 2.32 a | 26.13 ± 3.14 a | 7.34 ± 0.88 a | - | - |
| Cd50 + GSH | 58.25 ± 9.64 a | 9.86 ± 2.67 c | 22.36 ± 3.14 a | 9.67 ± 2.03 a | - | - | ||
| Harukaze | Cd50 | 59.46 ± 7.17 a | 18.36 ± 2.06 b | 20.51 ± 5.97 a | 1.67 ± 0.49 b | - | - | |
| Cd50 + GSH | 57.93 ± 0.58 a | 11.71 ± 1.22 bc | 27.05 ± 1.07 a | 3.31 ± 0.33 b | - | - |
| Index | Tl | RCW% | F Ethanol% | F H2O% | F NaCl% | F HAC% | F HCl% | F Residue% |
|---|---|---|---|---|---|---|---|---|
| Tl | 1 | |||||||
| RCW% | 0.860 ** | 1 | ||||||
| F ethanol% | –0.774 ** | –0.812 ** | 1 | |||||
| F H2O% | 0.545 | 0.776 ** | –0.805 ** | 1 | ||||
| F NaCl% | –0.205 | –0.316 | 0.254 | –0.255 | 1 | |||
| F HAC% | 0.767 ** | 0.753 ** | –0.954 ** | 0.663 * | –0.460 | 1 | ||
| F HCl% | 0.698 * | 0.791 ** | –0.891 ** | 0.700 * | –0.637 * | 0.954 ** | 1 | |
| F Residue% | 0.747 ** | 0.550 | –0.823 ** | 0.439 | –0.404 | 0.913 ** | 0.822 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Z.; Hu, Z.; Yin, X.; Song, G.; Cai, Q. Exogenous Glutathione Alleviation of Cd Toxicity in Italian Ryegrass (Lolium multiflorum) by Modulation of the Cd Absorption, Subcellular Distribution, and Chemical Form. Int. J. Environ. Res. Public Health 2020, 17, 8143. https://doi.org/10.3390/ijerph17218143
Fang Z, Hu Z, Yin X, Song G, Cai Q. Exogenous Glutathione Alleviation of Cd Toxicity in Italian Ryegrass (Lolium multiflorum) by Modulation of the Cd Absorption, Subcellular Distribution, and Chemical Form. International Journal of Environmental Research and Public Health. 2020; 17(21):8143. https://doi.org/10.3390/ijerph17218143
Chicago/Turabian StyleFang, Zhigang, Zhaoyang Hu, Xinqiang Yin, Gang Song, and Qingsheng Cai. 2020. "Exogenous Glutathione Alleviation of Cd Toxicity in Italian Ryegrass (Lolium multiflorum) by Modulation of the Cd Absorption, Subcellular Distribution, and Chemical Form" International Journal of Environmental Research and Public Health 17, no. 21: 8143. https://doi.org/10.3390/ijerph17218143
APA StyleFang, Z., Hu, Z., Yin, X., Song, G., & Cai, Q. (2020). Exogenous Glutathione Alleviation of Cd Toxicity in Italian Ryegrass (Lolium multiflorum) by Modulation of the Cd Absorption, Subcellular Distribution, and Chemical Form. International Journal of Environmental Research and Public Health, 17(21), 8143. https://doi.org/10.3390/ijerph17218143