Herbicide Glyphosate: Toxicity and Microbial Degradation

 ,
,  , ,
, ,  ,
,  , ,
, , 
Abstract
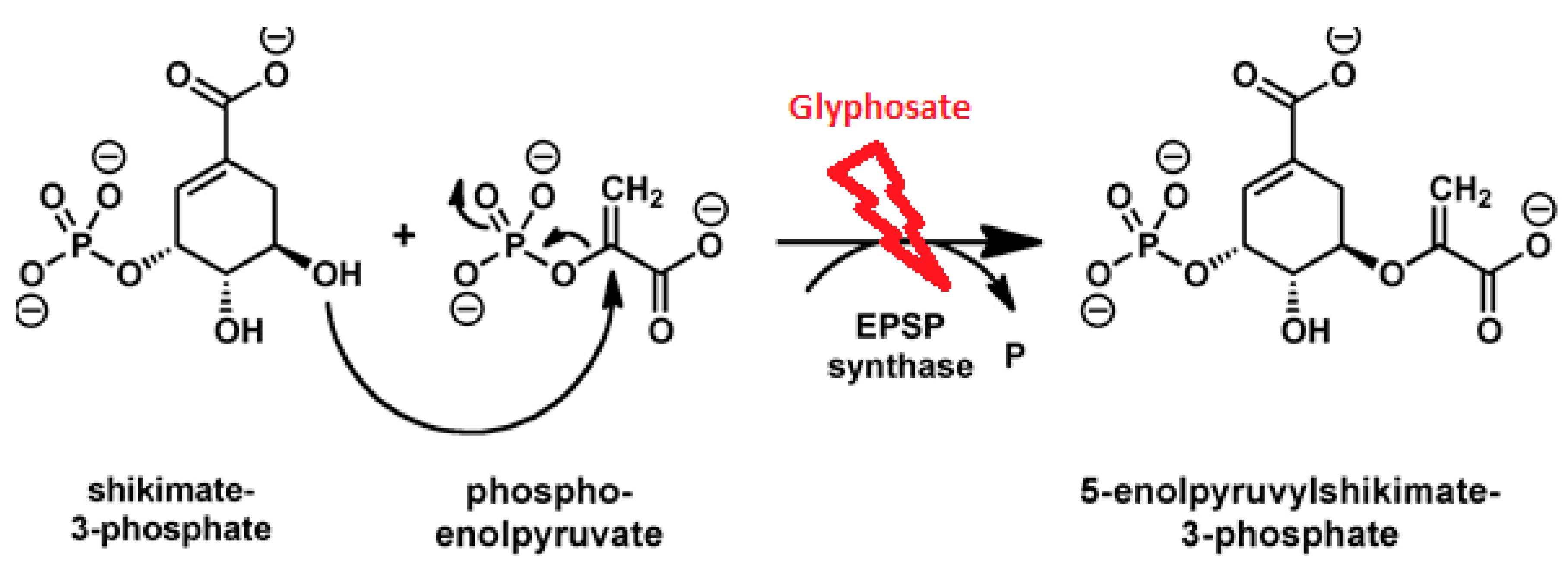
1. Introduction
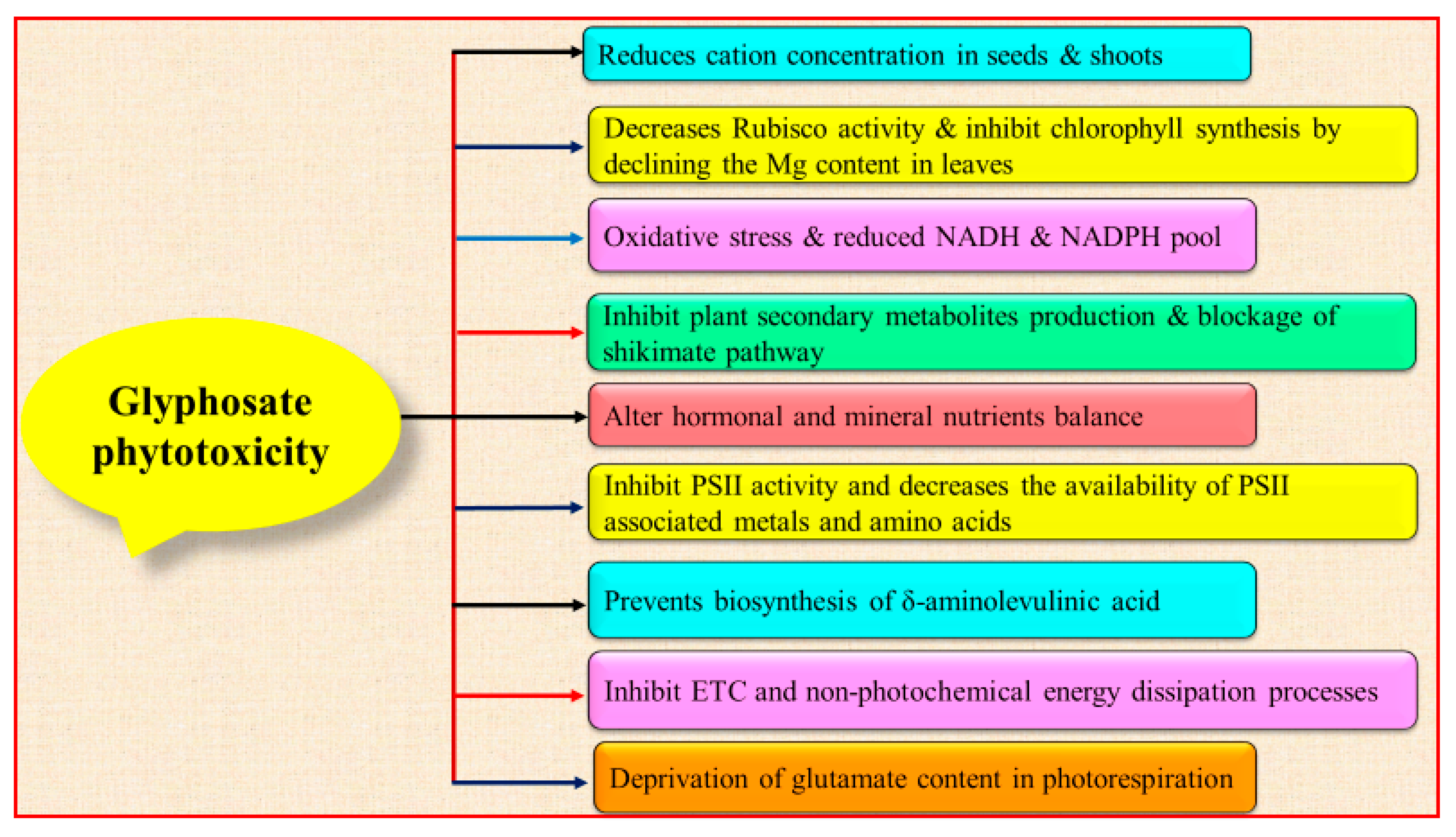
2. Glyphosate Toxicity
2.1. Ecotoxicity
2.2. Cytotoxicity and Genotoxicity
3. Microbial Degradation of Glyphosate
4. Degradation Kinetics and Glyphosate Residues
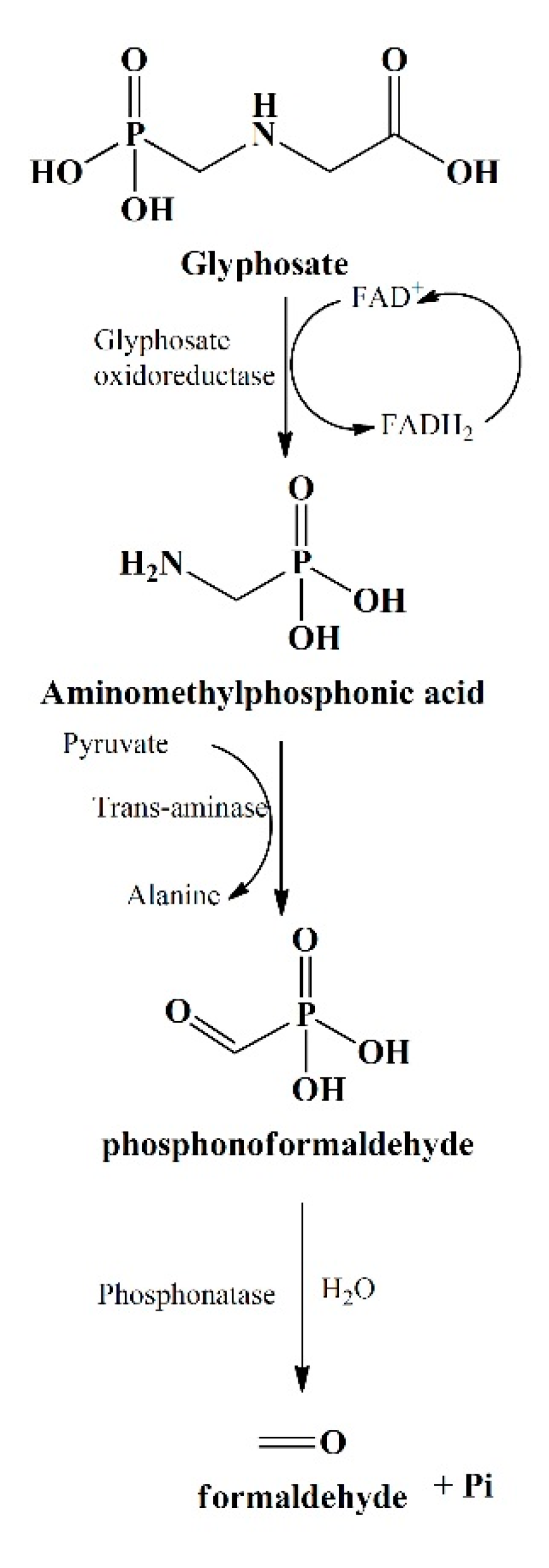
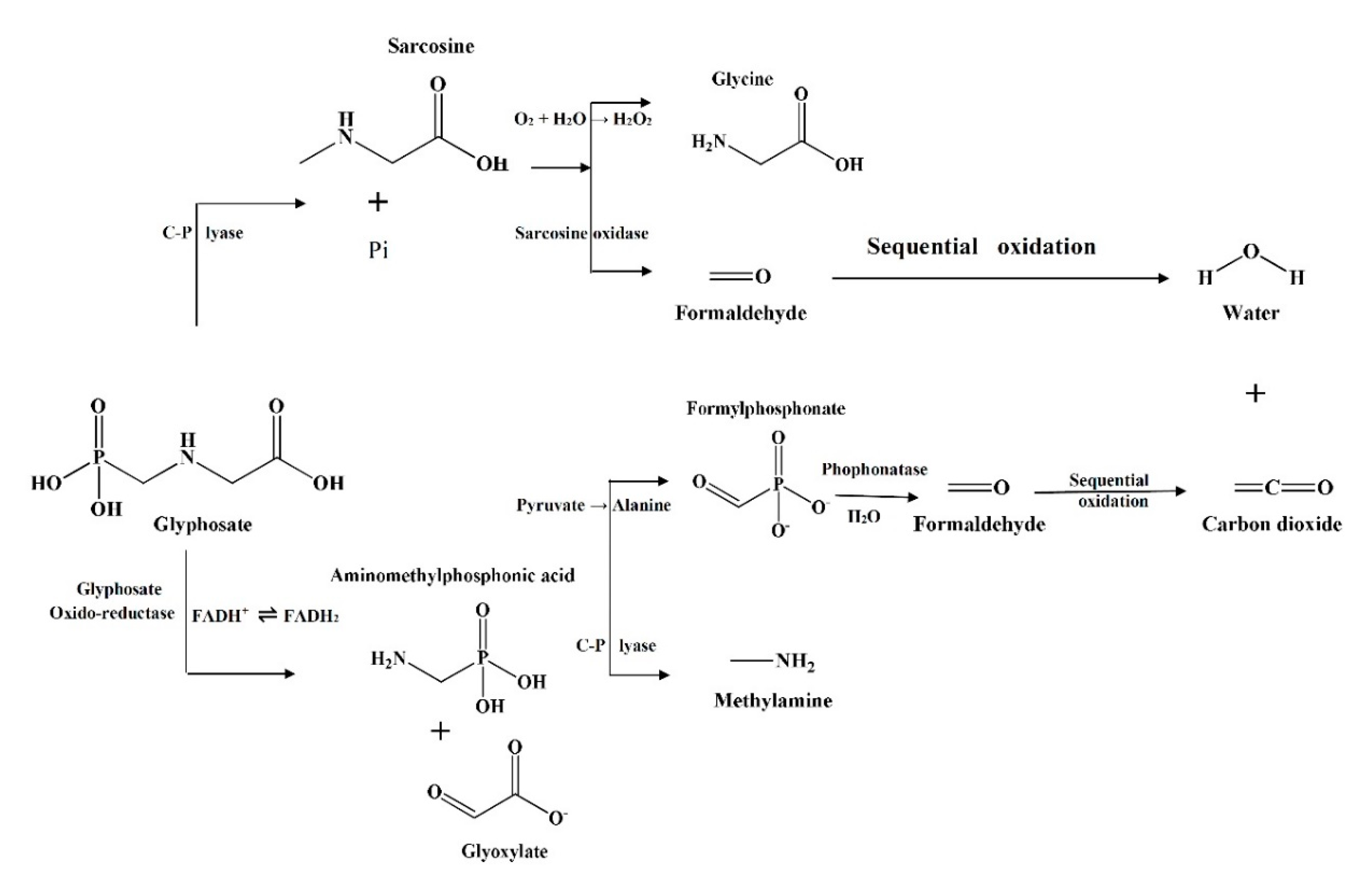
5. Mechanism Underlying Bio-Degradation
6. Enzymatic Mediated Metabolism of Glyphosate
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gill, J.P.K.; Sethi, N.; Mohan, A. Analysis of the glyphosate herbicide in water, soil and food using derivatising agents. Environ. Chem. Lett. 2017, 15, 85–100. [Google Scholar] [CrossRef]
- Zhang, C.; Hu, X.; Luo, J.; Wu, Z.; Wang, L.; Li, B.; Wang, Y.; Sun, G. Degradation dynamics of glyphosate in different types of citrus orchard soils in China. Molecules 2015, 20, 1161–1175. [Google Scholar] [CrossRef]
- Conrad, A.; Schröter-Kermani, C.; Hoppe, H.W.; Rüther, M.; Pieper, S.; Kolossa-Gehring, M. Glyphosate in German adults—Time trend (2001 to 2015) of human exposure to a widely used herbicide. Int. J. Hyg. Environ. Health 2017, 220, 8–16. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Datta, S.; Wani, A.B.; Dhanjal, D.S.; Romero, R.; Singh, J. Glyphosate uptake, translocation, resistance emergence in crops, analytical monitoring, toxicity and degradation: A review. Environ. Chem. Lett. 2020, 18, 663–702. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. In Proceedings of the Pest Management Science; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2008; Volume 64, pp. 319–325. [Google Scholar]
- Dill, G.M. Glyphosate-resistant crops: History, status and future. In Proceedings of the Pest Management Science; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2005; Volume 61, pp. 219–224. [Google Scholar]
- Haslam, E. The Shikimate Pathway: Biosynthesis of Natural Products Series; Butterworths: London, UK, 2014. [Google Scholar]
- Sviridov, A.V.; Shushkova, T.V.; Ermakova, I.T.; Ivanova, E.V.; Epiktetov, D.O.; Leont’evskii, A.A. Microbial degradation of glyphosate herbicides (review). Prikl. Biokhim. Mikrobiol. 2015, 51, 183–190. [Google Scholar] [PubMed]
- Hadi, F.; Mousavi, A.; Noghabi, K.A.; Tabar, H.G.; Salmanian, A.H. New bacterial strain of the genus Ochrobactrum with glyphosate-degrading activity. J. Environ. Sci. Health—Part B Pestic. Food Contam. Agric. Wastes 2013, 48, 208–213. [Google Scholar] [CrossRef]
- Wang, S.; Seiwert, B.; Kästner, M.; Miltner, A.; Schäffer, A.; Reemtsma, T.; Yang, Q.; Nowak, K.M. (Bio)degradation of glyphosate in water-sediment microcosms—A stable isotope co-labeling approach. Water Res. 2016, 99, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Annett, R.; Habibi, H.R.; Hontela, A. Impact of glyphosate and glyphosate-based herbicides on the freshwater environment. J. Appl. Toxicol. 2014, 34, 458–479. [Google Scholar] [CrossRef]
- Hanke, I.; Wittmer, I.; Bischofberger, S.; Stamm, C.; Singer, H. Relevance of urban glyphosate use for surface water quality. Chemosphere 2010, 81, 422–429. [Google Scholar] [CrossRef]
- Shushkova, T.; Ermakova, I.; Leontievsky, A. Glyphosate bioavailability in soil. Biodegradation 2010, 21, 403–410. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. Some Organophosphate Insecticides and Herbicides. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 2017; Volume 112, pp. 1–452. [Google Scholar]
- Niemann, L.; Sieke, C.; Pfeil, R.; Solecki, R. A critical review of glyphosate findings in human urine samples and comparison with the exposure of operators and consumers. J. Verbrauch. Leb. 2015, 10, 3–12. [Google Scholar] [CrossRef]
- Mesnage, R.; Defarge, N.; Spiroux de Vendômois, J.; Séralini, G.E. Potential toxic effects of glyphosate and its commercial formulations below regulatory limits. Food Chem. Toxicol. 2015, 84, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Zhan, H.; Feng, Y.; Fan, X.; Chen, S. Recent advances in glyphosate biodegradation. Appl. Microbiol. Biotechnol. 2018, 102, 5033–5043. [Google Scholar] [CrossRef]
- Grandcoin, A.; Piel, S.; Baurès, E. AminoMethylPhosphonic acid (AMPA) in natural waters: Its sources, behavior and environmental fate. Water Res. 2017, 117, 187–197. [Google Scholar] [CrossRef]
- Xu, X.; Ji, F.; Fan, Z.; He, L. Degradation of glyphosate in soil photocatalyzed by Fe3O4/SiO2/TiO2 under solar light. Int. J. Environ. Res. Public Health 2011, 8, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.C.; Rodrigues, E.G.; Christiani, D.C. Environmental Health Hazards in the Tropics. In Hunter's Tropical Medicine and Emerging Infectious Diseases; Ryan, E.T., Hill, D.R., Solomon, T., Aronson, N.E., Endy, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 200–208. [Google Scholar]
- Chandrasekera, W.U.; Weeratunga, N.P. The lethal impacts of Roundup® (glyphosate) on the fingerlings of guppy, Poecilia reticulata Peters, 1859. Asian Fish. Sci. 2011, 24, 367–378. [Google Scholar]
- Zablotowicz, R.M.; Reddy, K.N. Nitrogenase activity, nitrogen content, and yield responses to glyphosate in glyphosate-resistant soybean. Crop Prot. 2007, 26, 370–376. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, D.G.; Lee, K.W.; Alam, I.; Lee, S.H.; Bahk, J.D.; Lee, B.H. Glyphosate-induced oxidative stress in rice leaves revealed by proteomic approach. Plant Physiol. Biochem. 2008, 46, 1062–1070. [Google Scholar] [CrossRef]
- Miteva, L.P.E.; Ivanov, S.V.; Alexieva, V.S. Alterations in glutathione pool and some related enzymes in leaves and roots of pea plants treated with the herbicide glyphosate. Russ. J. Plant Physiol. 2010, 57, 131–136. [Google Scholar] [CrossRef]
- Orcaray, L.; Zulet, A.; Zabalza, A.; Royuela, M. Impairment of carbon metabolism induced by the herbicide glyphosate. J. Plant Physiol. 2012, 169, 27–33. [Google Scholar] [CrossRef]
- Zabalza, A.; Orcaray, L.; Fernández-Escalada, M.; Zulet-González, A.; Royuela, M. The pattern of shikimate pathway and phenylpropanoids after inhibition by glyphosate or quinate feeding in pea roots. Pestic. Biochem. Physiol. 2017, 141, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.N.; Rimando, A.M.; Duke, S.O. Aminomethylphosphonic acid, a metabolite of glyphosate, causes injury in glyphosate-treated, glyphosate-resistant soybean. J. Agric. Food Chem. 2004, 52, 5139–5143. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.-A.; Nuttens, A.; Larvor, V.; Renault, D.; Couée, I.; Sulmon, C.; Gouesbet, G. Low environmentally relevant levels of bioactive xenobiotics and associated degradation products cause cryptic perturbations of metabolism and molecular stress responses in Arabidopsis thaliana. J. Exp. Bot. 2013, 64, 2753–2766. [Google Scholar] [CrossRef]
- Zobiole, L.H.S.; Kremer, R.J.; Oliveira, R.S.; Constantin, J. Glyphosate affects microorganisms in rhizospheres of glyphosate-resistant soybeans. J. Appl. Microbiol. 2011, 110, 118–127. [Google Scholar] [CrossRef]
- Zobiole, L.H.S.; Oliveira, R.S.; Kremer, R.J.; Constantin, J.; Yamada, T.; Castro, C.; Oliveira, F.A.; Oliveira, A. Effect of glyphosate on symbiotic N2 fixation and nickel concentration in glyphosate-resistant soybeans. Appl. Soil Ecol. 2010, 44, 176–180. [Google Scholar] [CrossRef]
- Yanniccari, M.; Tambussi, E.; Istilart, C.; Castro, A.M. Glyphosate effects on gas exchange and chlorophyll fluorescence responses of two Lolium perenne L. biotypes with differential herbicide sensitivity. Plant Physiol. Biochem. 2012, 57, 210–217. [Google Scholar] [CrossRef]
- Gomes, M.P.; Maccario, S.; Le Manac’h, S.G.; Lucotte, M.; Moingt, M.; Paquet, S.; Labrecque, M.; Juneau, P. Comments on the “Glyphosate herbicide residue determination in samples of environmental importance using spectrophotometric method”. J. Hazard. Mater. 2017, 340, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Marsh, H.V.; Evans, H.J.; Matrone, G. Investigations of the Role of Iron in Chlorophyll Metabolism. II. Effect of Iron Deficiency on Chlorophyll Synthesis. Plant Physiol. 1963, 38, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Mkandawire, M.; Teixeira Da Silva, J.A.; Dudel, E.G. The lemna bioassay: Contemporary issues as the most standardized plant bioassay for aquatic ecotoxicology. Crit. Rev. Environ. Sci. Technol. 2014, 44, 154–197. [Google Scholar] [CrossRef]
- Vivancos, P.D.; Driscoll, S.P.; Bulman, C.A.; Ying, L.; Emami, K.; Treumann, A.; Mauve, C.; Noctor, G.; Foyer, C.H. Perturbations of amino acidmetabolism associated with glyphosate-dependent inhibition of shikimic acid metabolism affect cellular redox homeostasis and alter the abundance of proteins involved in photosynthesis and photorespiration. Plant Physiol. 2011, 157, 256–268. [Google Scholar] [CrossRef]
- Hussain, S.; Siddique, T.; Arshad, M.; Saleem, M. Bioremediation and phytoremediation of pesticides: Recent advances. Crit. Rev. Environ. Sci. Technol. 2009, 39, 843–907. [Google Scholar] [CrossRef]
- Cakmak, I.; Yazici, A.; Tutus, Y.; Ozturk, L. Glyphosate reduced seed and leaf concentrations of calcium, manganese, magnesium, and iron in non-glyphosate resistant soybean. Eur. J. Agron. 2009, 31, 114–119. [Google Scholar] [CrossRef]
- Ding, W.; Reddy, K.N.; Zablotowicz, R.M.; Bellaloui, N.; Arnold Bruns, H. Physiological responses of glyphosate-resistant and glyphosate-sensitive soybean to aminomethylphosphonic acid, a metabolite of glyphosate. Chemosphere 2011, 83, 593–598. [Google Scholar] [CrossRef]
- De María, N.; Becerril, J.M.; García-Plazaola, J.I.; Hernández, A.; De Felipe, M.R.; Fernández-Pascual, M. New insights on glyphosate mode of action in nodular metabolism: Role of shikimate accumulation. J. Agric. Food Chem. 2006, 54, 2621–2628. [Google Scholar] [CrossRef] [PubMed]
- Bonga, J.M. Clonal Propagation of Mature Trees: Problems and Possible Solutions. In Cell and Tissue Culture in Forestry; Springer: Dordrecht, The Netherlands, 1987; pp. 249–271. [Google Scholar]
- Marchiosi, R.; Ferrarese, M.D.L.L.; Bonini, E.A.; Fernandes, N.G.; Ferro, A.P.; Ferrarese-Filho, O. Glyphosate-induced metabolic changes in susceptible and glyphosate-resistant soybean (Glycine max L.) roots. Pest. Biochem. Physiol. 2009, 93, 28–33. [Google Scholar] [CrossRef]
- Sugano, S.; Sugimoto, T.; Takatsuji, H.; Jiang, C.J. Induction of resistance to Phytophthora sojae in soyabean (Glycine max) by salicylic acid and ethylene. Plant Pathol. 2013, 62, 1048–1056. [Google Scholar] [CrossRef]
- Guo, J.; Boxall, A.; Selby, K. Do pharmaceuticals pose a threat to primary producers? Crit. Rev. Environ. Sci. Technol. 2015, 45, 2565–2610. [Google Scholar] [CrossRef]
- Helander, M.; Saloniemi, I.; Omacini, M.; Druille, M.; Salminen, J.P.; Saikkonen, K. Glyphosate decreases mycorrhizal colonization and affects plant-soil feedback. Sci. Total Environ. 2018, 642, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Muangphra, P.; Kwankua, W.; Gooneratne, R. Genotoxic effects of glyphosate or paraquat on earthworm coelomocytes. Environ. Toxicol. 2014, 29, 612–620. [Google Scholar] [CrossRef]
- Pochron, S.; Simon, L.; Mirza, A.; Littleton, A.; Sahebzada, F.; Yudell, M. Glyphosate but not Roundup® harms earthworms (Eisenia fetida). Chemosphere 2020, 241, 125017. [Google Scholar] [CrossRef]
- Székács, I.; Fejes, Á.; Klátyik, S.; Takács, E.; Patkó, D.; Pomóthy, J.; Mörtl, M.; Horváth, R.; Madarász, E.; Darvas, B.; et al. Environmental and toxicological impacts of glyphosate with its formulating adjuvant. Int. J. Biol. Vet. Agric. Food Eng. 2014, 8, 212–218. [Google Scholar]
- Gaur, H.; Bhargava, A. Investigation of Toxicity Induced by Chemicals and Their Mechanisms Using Zebrafish Vertebrate Animal Model. Ph.D. Dissertation, Indian Institute of Technology Hyderabad, Telangana, India, 2019. [Google Scholar]
- Hoopman, A.; North, H.; Rajamohan, A.; Bowsher, J. Toxicity assessment of glyphosate on honey bee (Apis mellifera) spermatozoa [abstract]. In Proceedings of the Society for Integrative & Comparative Biology (SCIB) Annual Meeting, San Francisco, CA, USA, 3–7 January 2018; pp. 3–7. [Google Scholar]
- Bernal, J.; Bernal, J.L.; Martin, M.T.; Nozal, M.J.; Anadón, A.; Martínez-Larrañaga, M.R.; Martínez, M.A. Development and validation of a liquid chromatography–fluorescence–mass spectrometry method to measure glyphosate and aminomethylphosphonic acid in rat plasma. J. Chromatogr. B 2010, 878, 3290–3296. [Google Scholar] [CrossRef]
- Milić, M.; Žunec, S.; Micek, V.; Kašuba, V.; Mikolić, A.; Tariba Lovaković, B.; Živković Semren, T.; Pavičić, I.; Marjanović Čermak, A.M.; Pizent, A.; et al. Oxidative stress, cholinesterase activity, and DNA damage in the liver, whole blood, and plasma of Wistar rats following a 28-day exposure to glyphosate. Arhiv za Higijenu Rada i Toksikologiju 2018, 69, 154–168. [Google Scholar] [CrossRef]
- Gill, J.P.K.; Sethi, N.; Mohan, A.; Datta, S.; Girdhar, M. Glyphosate toxicity for animals. Environ. Chem. Lett. 2018, 16, 401–426. [Google Scholar] [CrossRef]
- Connolly, A.; Leahy, M.; Jones, K.; Kenny, L.; Coggins, M.A. Glyphosate in Irish adults—A pilot study in 2017. Environ. Res. 2018, 165, 235–236. [Google Scholar] [CrossRef]
- Soukup, S.T.; Merz, B.; Bub, A.; Hoffmann, I.; Watzl, B.; Steinberg, P.; Kulling, S.E. Glyphosate and AMPA levels in human urine samples and their correlation with food consumption: Results of the cross-sectional KarMeN study in Germany. Arch. Toxicol. 2020, 94, 1575. [Google Scholar] [CrossRef] [PubMed]
- Australian Drinking Water Guidelines 6. 2011. Available online: http://www.clearwater.asn.au/user-data/.../Aust_drinking_water_guidelines (accessed on 23 February 2015).
- Horth, H.; Blackmore, K. Survey of Glyphosate and AMPA in Groundwaters and Surface Waters in Europe; No: UC8073.02; WRc plc: Swindon, UK, 2009. [Google Scholar]
- Gasnier, C.; Dumont, C.; Benachour, N.; Clair, E.; Chagnon, M.C.; Séralini, G.E. Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology 2009, 262, 184–191. [Google Scholar] [CrossRef]
- Richard, S.; Moslemi, S.; Sipahutar, H.; Benachour, N.; Seralini, G.E. Differential effects of glyphosate and Roundup on human placental cells and aromatase. Environ. Health Perspect. 2005, 113, 716–720. [Google Scholar] [CrossRef]
- Pieniążek, D.; Bukowska, B.; Duda, W. Comparison of the effect of Roundup Ultra 360 SL pesticide and its active compound glyphosate on human erythrocytes. Pestic. Biochem. Physiol. 2004, 79, 58–63. [Google Scholar] [CrossRef]
- Seok, S.J.; Park, J.S.; Hong, J.R.; Gil, H.W.; Yang, J.O.; Lee, E.Y.; Song, H.Y.; Hong, S.Y. Surfactant volume is an essential element in human toxicity in acute glyphosate herbicide intoxication. Clin. Toxicol. 2011, 49, 892–899. [Google Scholar] [CrossRef]
- Koller, V.J.; Fürhacker, M.; Nersesyan, A.; Mišík, M.; Eisenbauer, M.; Knasmueller, S. Cytotoxic and DNA-damaging properties of glyphosate and Roundup in human-derived buccal epithelial cells. Arch. Toxicol. 2012, 86, 805–813. [Google Scholar] [CrossRef]
- Zouaoui, K.; Dulaurent, S.; Gaulier, J.M.; Moesch, C.; Lachâtre, G. Determination of glyphosate and AMPA in blood and urine from humans: About 13 cases of acute intoxication. Forensic Sci. Int. 2013, 226, e20–e25. [Google Scholar] [CrossRef]
- Thongprakaisang, S.; Thiantanawat, A.; Rangkadilok, N.; Suriyo, T.; Satayavivad, J. Glyphosate induces human breast cancer cells growth via estrogen receptors. Food Chem. Toxicol. 2013, 59, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Anifandis, G.; Katsanaki, K.; Lagodonti, G.; Messini, C.; Simopoulou, M.; Dafopoulos, K.; Daponte, A. The effect of glyphosate on human sperm motility and sperm DNA fragmentation. Inter. J. Environ. Res. Public Health 2018, 15, 1117. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, M.; Huras, B.; Bukowska, B. The effect of metabolites and impurities of glyphosate on human erythrocytes (in vitro). Pestic. Biochem. Physiol. 2014, 109, 34–43. [Google Scholar] [CrossRef]
- Coalova, I.; de Molina, M.D.C.R.; Chaufan, G. Influence of the spray adjuvant on the toxicity effects of a glyphosate formulation. Toxicol. Vitro 2014, 28, 1306–1311. [Google Scholar] [CrossRef]
- Martinez, A.; Al-Ahmad, A.J. Effects of glyphosate and aminomethylphosphonic acid on an isogeneic model of the human blood-brain barrier. Toxicol. Lett. 2019, 304, 39–49. [Google Scholar] [CrossRef]
- Li, Q.; Lambrechts, M.J.; Zhang, Q.; Liu, S.; Ge, D.; Yin, R.; Xi, M.; You, Z. Glyphosate and AMPA inhibit cancer cell growth through inhibiting intracellular glycine synthesis. Drug Des. Dev. Ther. 2013, 7, 635. [Google Scholar]
- Hoppin, J.A.; Valcin, M.; Henneberger, P.K.; Kullman, G.J.; Umbach, D.M.; London, S.J.; Alavanja, M.C.; Sandler, D.P. Pesticide use and chronic bronchitis among farmers in the Agricultural Health Study. Am. J. Ind. Med. 2007, 50, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Henneberger, P.K.; Liang, X.; London, S.J.; Umbach, D.M.; Sandler, D.P.; Hoppin, J.A. Exacerbation of Symptoms in Agricultural Pesticide Applicators with Asthma. Int. Arch. Occup. Environ. Health 2014, 87, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Dick, R.E.; Quinn, J.P. Control of glyphosate uptake and metabolism in Pseudomonas sp. 4ASW. FEMS Microbiol. Lett. 1995, 134, 177–182. [Google Scholar] [CrossRef]
- Ermakova, I.T.; Shushkova, T.V.; Sviridov, A.V.; Zelenkova, N.F.; Vinokurova, N.G.; Baskunov, B.P.; Leontievsky, A.A. Organophosphonates utilization by soil strains of Ochrobactrum anthropi and Achromobacter sp. Arch. Microbiol. 2017, 199, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Firdous, S.; Iqbal, S.; Anwar, S. Optimization and Modeling of Glyphosate Biodegradation by a Novel Comamonas odontotermitis P2 through Response Surface Methodology. Pedosphere 2017. [Google Scholar] [CrossRef]
- Firdous, S.; Iqbal, S.; Anwar, S.; Jabeen, H. Identification and analysis of 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene from glyphosate-resistant Ochrobactrum intermedium Sq20. Pest Manag. Sci. 2018, 74, 1184–1196. [Google Scholar] [CrossRef]
- Lerbs, W.; Stock, M.; Parthier, B. Physiological aspects of glyphosate degradation in Alcaligenes spec. strain GL. Arch. Microbiol. 1990, 153, 146–150. [Google Scholar] [CrossRef]
- Moore, J.K.; Braymer, H.D.; Larson, A.D. Isolation of a Pseudomonas sp. which utilizes the phosphonate herbicide glyphosate. Appl. Environ. Microbiol. 1983, 46, 316–320. [Google Scholar] [CrossRef]
- Obojska, A.; Ternan, N.G.; Lejczak, B.; Kafarski, P.; McMullan, G. Organophosphonate utilization by the thermophile GeoBacillus caldoxylosilyticus T20. Appl. Environ. Microbiol. 2002, 68, 2081–2084. [Google Scholar] [CrossRef]
- Pipke, R.; Amrhein, N. Degradation of the phosphonate herbicide glyphosate by Arthrobacter atrocyaneus ATCC 13752. Appl. Environ. Microbiol. 1988, 54, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Pipke, R.; Amrhein, N.; Jacob, G.S.; Schaefer, J.; Kishore, G.M. Metabolism of glyphosate in an Arthrobacter sp. GLP-1. Eur. J. Biochem. 1987, 165, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Bujacz, E.; Wieczorek, P.; Krzys, T.; Goła, Z.; Lejczak, B.; Kavfarski, P. Organophosphonate Utilization by the Wild-Type Strain of Penicillium notatum. Appl. Environ. Microbiol. 1995, 61, 2905–2910. [Google Scholar] [CrossRef]
- Obojska, A.; Lejczak, B.; Kubrak, M. Degradation of phosphonates by Streptomycete isolates. Appl. Microbiol. Biotechnol. 1999, 51, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yang, G.; Zhao, H.; Shi, G.; Geng, Y.; Hou, T.; Tao, K. Isolation, identification and characterization of a glyphosate-degrading bacterium, Bacillus cereus CB4, from soil. J. Gen. Appl. Microbiol. 2012, 58, 263–271. [Google Scholar] [CrossRef]
- Balthazor, T.M.; Hallas, L.E. Glyphosate-degrading microorganisms from industrial activated sludge. Appl. Environ. Microbiol. 1986, 51, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Penaloza-Vazquez, A.; Mena, G.L.; Herrera-Estrella, L.; Bailey, A.M. Cloning and sequencing of the genes involved in glyphosate utilization by Pseudomonas pseudomallei. Appl. Environ. Microbiol. 1995, 61, 538–543. [Google Scholar] [CrossRef]
- McAuliffe, K.S.; Hallas, L.E.; Kulpa, C.F. Glyphosate degradation by Agrobacterium radiobacter isolated from activated sludge. J. Ind. Microbiol. 1990, 6, 219–221. [Google Scholar] [CrossRef]
- Fu, G.-M.; Chen, Y.; Li, R.-Y.; Yuan, X.-Q.; Liu, C.M.; Li, B.; Wan, Y. Pathway and rate-limiting step of glyphosate degradation by Aspergillus oryzae A-F02. Prep. Biochem. Biotechnol. 2017, 47, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Klimek, M.; Lejczak, B.; Kafarski, P.; Forlani, G. Metabolism of the phosphonate herbicide glyphosate by a non-nitrate-utilizing strain of Penicilliumchrysogenum. Pest Manag. Sci. 2001, 57, 815–821. [Google Scholar] [CrossRef]
- Krzyśko-Łupicka, T.; Orlik, A. Use of glyphosate as the sole source of phosphorus or carbon for the selection of soil-borne fungal strains capable to degrade this herbicide. Chemosphere 1997, 34, 2601–2605. [Google Scholar] [CrossRef]
- Krzyśko-Lupicka, T.; Strof, W.; Kubś, K.; Skorupa, M.; Wieczorek, P.; Lejczak, B.; Kafarski, P. The ability of soil-borne fungi to degrade organophosphonate carbon-to-phosphorus bonds. Appl. Microbiol. Biotechnol. 1997, 48, 549–552. [Google Scholar] [CrossRef]
- Pipke, R.; Amrhein, N. Isolation and Characterization of a Mutant of Arthrobacter sp. Strain GLP-1 Which Utilizes the Herbicide Glyphosate as Its Sole Source of Phosphorus and Nitrogen. Appl. Environ. Microbiol. 1988, 54, 2868–2870. [Google Scholar] [CrossRef]
- Kishore, G.M.; Jacob, G.S. Degradation of glyphosate by Pseudomonas sp. PG2982 via a sarcosine intermediate. J. Biol. Chem. 1987, 262, 12164–12168. [Google Scholar] [PubMed]
- Selvapandiyan, A.; Bhatnagar, R.K. Isolation of a glyphosate-metabolising Pseudomonas: Detection, partial purification and localisation of carbon-phosphorus lyase. Appl. Microbiol. Biotechnol. 1994, 40, 876–882. [Google Scholar] [CrossRef]
- Shinabarger, D.L.; Braymer, H.D. Glyphosate catabolism by Pseudomonas sp. strain PG2982. J. Bacteriol. 1986, 168, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Jacob, G.S.; Garbow, J.R.; Hallas, L.E.; Kimack, N.M.; Kishore, G.M.; Schaeffer, J. Metabolism of glyophosate in Pseudomonas sp. strain LBr. Appl. Environ. Microbiol. 1988, 54, 2953–2958. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.P.; Peden, J.M.M.; Dick, R.E. Carbon-phosphorus bond cleavage by Gram-positive and Gram-negative soil bacteria. Appl. Microbiol. Biotechnol. 1989, 31, 283–287. [Google Scholar] [CrossRef]
- Sviridov, A.V.; Shushkova, T.V.; Zelenkova, N.F.; Vinokurova, N.G.; Morgunov, I.G.; Ermakova, I.T.; Leontievsky, A.A. Distribution of glyphosate and methylphosphonate catabolism systems in soil bacteria Ochrobactrum anthropi and Achromobacter sp. Appl. Microbiol. Biotechnol. 2012, 93, 787–796. [Google Scholar] [CrossRef]
- Duke, S.O. Glyphosate degradation in glyphosate-resistant and -susceptible crops and weeds. J. Agric. Food Chem. 2011, 59, 5835–5841. [Google Scholar] [CrossRef]
- Tazdaït, D.; Salah, R.; Grib, H.; Abdi, N.; Mameri, N. Kinetic study on biodegradation of glyphosate with unacclimated activated sludge. Int. J. Environ. Health Res. 2018, 28, 448–459. [Google Scholar] [CrossRef]
- Edwards, V.H. The influence of high substrate concentrations on microbial kinetics. Biotechnol. Bioeng. 1970, 12, 679–712. [Google Scholar] [CrossRef]
- Aiba, S.; Shoda, M.; Nagalani, M. Kinetics of product inhibition in alcohol fermentation. Biotechnol. Bioeng. 1968, 10, 845–864. [Google Scholar] [CrossRef]
- Han, K.; Levenspiel, O. Extended Monod kinetics for substrate, product, and cell inhibition. Biotechnol. Bioeng. 1988, 32, 430–437. [Google Scholar] [CrossRef]
- Luong, J.H.J. Generalization of Monod kinetics for analysis of growth data with substrate inhibition. Biotechnol. Bioeng. 1987, 29, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Tseng, M.M.C.; Wayman, M. Kinetics of yeast growth: Inhibition-threshold substrate concentrations. Can. J. Microbiol. 1975, 21, 994–1003. [Google Scholar] [CrossRef]
- Yano, T.; Koga, S. Dynamic behavior of the chemostat subject to substrate inhibition. Biotechnol. Bioeng. 1969, 11, 139–153. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Singh, J. Kinetic study of the biodegradation of glyphosate by indigenous soil bacterial isolates in presence of humic acid, Fe(III) and Cu(II) ions. J. Environ. Chem. Eng. 2019, 7, 103098. [Google Scholar] [CrossRef]
- Pipke, R.; Schulz, A.; Amrhein, N. Uptake of glycophosphate by an Arthrobacter sp. Appl. Environ. Microbiol. 1987, 53, 974–978. [Google Scholar] [CrossRef] [PubMed]
- Hadi, F.; Mousavi, A.; Salmanian, A.H.; Akbari Noghabi, K. Glyphosate Tolerance in Transgenic Canola by a Modified Glyphosate Oxidoreductase (gox) Gene. Prog. Biol. Sci. 2012, 2, 50–58. [Google Scholar] [CrossRef]
- Hove-Jensen, B.; Zechel, D.L.; Jochimsen, B. Utilization of Glyphosate as Phosphate Source: Biochemistry and Genetics of Bacterial Carbon-Phosphorus Lyase. Microbiol. Mol. Biol. Rev. 2014, 78, 176–197. [Google Scholar] [CrossRef]
- Sviridov, A.V.; Shushkova, T.V.; Ermakova, I.T.; Ivanova, E.V.; Leontievsky, A.A. Glyphosate: Safety risks, biodegradation, and bioremediation. In Current Environmental Issues and Challenges; Springer: Dordrecht, The Netherlands, 2014; pp. 183–195. ISBN 9789401787772. [Google Scholar]
- Villarreal-Chiu, J.F.; Quinn, J.P.; McGrath, J.W. The genes and enzymes of phosphonate metabolism by bacteria, and their distribution in the marine environment. Front. Microbiol. 2012, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.M.; Ye, Q.Z.; Zhu, Z.; Wanner, B.L.; Walsh, C.T. Molecular biology of carbon-phosphorus bond cleavage. Cloning and sequencing of the phn (psiD) genes involved in alkylphosphonate uptake and C-P lyase activity in Escherichia coli B. J. Biol. Chem. 1990, 265, 4461–4471. [Google Scholar]
- Hove-Jensen, B.; McSorley, F.R.; Zechel, D.L. Physiological role of phnP-specified phosphoribosyl cyclic phosphodiesterase in catabolism of organophosphonic acids by the carbon-phosphorus lyase pathway. J. Am. Chem. Soc. 2011, 133, 3617–3624. [Google Scholar] [CrossRef]
- Metcalf, W.W.; Wanner, B.L. Evidence for a fourteen-gene, phnC to phnP locus for phosphonate metabolism in Escherichia coli. Gene 1993, 129, 27–32. [Google Scholar] [CrossRef]
- Hove-Jensen, B.; Rosenkrantz, T.J.; Zechel, D.L.; Willemoës, M. Accumulation of intermediates of the carbon-phosphorus lyase pathway for phosphonate degradation in phn mutants of Escherichia coli. J. Bacteriol. 2010, 192, 370–374. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Santos-Beneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Adelowo, F.E.; Olu-Arotiowa, O.A.; Amuda, O.S. Biodegradation of glyphosate by fungi species. Adv. Biosci. Bioeng. 2014, 2, 104–118. [Google Scholar]
- Arfarita, N.; Djuhari, D.; Prasetya, B.; Imai, T. The application of trichoderma viride strain frp 3 for biodegradation of glyphosate herbicide in contaminated land. J. Agric. Sci. 2016, 38, 275–281. [Google Scholar] [CrossRef]
- Carranza, C.S.; Barberis, C.L.; Chiacchiera, S.M.; Magnoli, C.E. Assessment of growth of Aspergillus spp. from agricultural soils in the presence of glyphosate. Rev. Argent. Microbiol. 2017, 49, 384–393. [Google Scholar] [CrossRef]
- Gard, J.K.; Feng, P.C.C.; Hutton, W.C. Nuclear magnetic resonance tim ecourse studies of glyphosate metabolism by microbial soil isolates. Xenobiotica 1997, 27, 633–644. [Google Scholar] [CrossRef]
- Liu, C.M.; McLean, P.A.; Sookdeo, C.C.; Cannon, F.C. Degradation of the herbicide glyphosate by members of the family Rhizobiaceae. Appl. Environ. Microbiol. 1991, 57, 1799–1804. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, Y.; Pourbabaei, A.A.; Javadi, A.; Abdolmohammad, M.H.; Saffari, M.; Morovvati, A. Biodegradation of glyphosate herbicide by Salinicoccus spp. isolated from Qom Hoze-soltan lake, Iran. Environ. Health Eng. Manag. J. 2015, 2, 31–36. [Google Scholar]
- Shushkova, T.V.; Ermakova, I.T.; Sviridov, A.V.; Leontievsky, A.A. Biodegradation of glyphosate by soil bacteria: Optimization of cultivation and the method for active biomass storage. Microbiology 2012, 81, 44–50. [Google Scholar] [CrossRef]
- Wackett, L.P.; Shames, S.L.; Venditti, C.P.; Walsh, C.T. Bacterial carbon-phosphorus lyase: Products, rates, and regulation of phosphonic and phosphinic acid metabolism. J. Bacteriol. 1987, 169, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Ajish, A.; Sukirtha, T.H. Biodegradation and Bioremediation of Glyphosate herbicide by Nocardia Mediterranie THS 1. Int. J. Sci. Res. 2020, 8, 34–36. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Species | Source | Enzymes/Gene | Location | Metabolites after Degradation | References |
|---|---|---|---|---|---|
| Ochrobactrum anthropi GDOS | Soil | - | Iran | AMPA | [16] |
| Pseudomonas sp. 4ASW | Soil | - | UK | Sarcosine | [71] |
| Comamonas odontotermitis P2 | Soil | Glyphosate oxidoreductase | Pakistan | - | [73] |
| Alcaligenes sp. GL | Selective medium | - | Germany | AMPA (5%) and sarcosine (95%) | [75] |
| Pseudomonas sp. PG298231 | Mixed culture | C-P lyase, sarcosine oxidase | Louisiana | Sarcosine | [76] |
| Geobacillus caldoxylosilyticus T20, Ochrobactrum anthropi LBAA | Soil | UK | AMPA | [77] | |
| Arthrobacter atrocyaneus ATCC 13752 | Microbial collection | C-P lyase | Germany | AMPA | [78] |
| Arthrobacter sp. GLP-1 | Selective medium | - | USA | Sarcosine | [79] |
| Penicillium notatum | Mutation of the wild type | - | Poland | AMPA | [80] |
| Streptomyces sp. StC | Sludge | - | Poland | Sarcosine | [81] |
| Bacillus cereus CB4 | Soil | - | China | AMPA, glyoxylate, sarcosine, glycine and formaldehyde | [82] |
| Flavobacterium sp. GD1 | Sludge | - | Missouri | AMPA | [83] |
| Pseudomonas pseudomallei | Soil | glpA and glpB | USA | AMPA | [84] |
| Achromobacter sp., Rhizobium radiobacter | Sludge | - | USA | AMPA | [85] |
| Aspergillus oryzae A-F02 | Soil | - | China | AMPA and methylamine | [86] |
| Aspergillus niger, Scopulariopsis sp., Trichoderma harzianum | Soil | - | Poland | AMPA | [88] |
| Pseudomonas sp. LBr | Sludge | Glyphosate oxidoreductase and C-P lyase | Missouri | AMPA (95%), sarcosine (5%) | [94] |
| Achromobacter sp. MPS 12 A, Ochrobactrum anthropi GPK 3 | Soil | sarcosine oxidase | Russia | Sacrosine AMPA | [96] |
| Bacillus subtilis, Rhizobium leguminosarum, Streptomyces sp. | Soil | C-P lyase and glyphosate oxidoreductase | India | AMPA and methylamine | [105] |
| Fusarium oxysporum, Trichoderma viridae, Aspergillus niger | Soil | - | Nigeria | AMPA and sarcosine | [116] |
| Trichoderma viride Strain FRP 3 | Soil | - | Indonesia | - | [117] |
| Aspergillus section Flavi and Aspergillus niger | - | - | Argentina | - | [118] |
| Ochrobactrum anthropi S5 | Soil | - | USA | AMPA | [119] |
| Rhizobium meliloti 1021 | Mutation of the wild strain | sarcosine oxidase | Massachusetts | Sarcosine | [120] |
| Salinicoccus sp. | Soil | - | Iran | AMPA | [121] |
| Ochrobactrum anthropi GPK 3, Achromobacter sp. 16 kg | Soil | Glyphosate oxidoreductase transaminase enzyme and phosphonatase | Russia | - | [122] |
| Agrobacterium radiobacter | Wastewater | - | US | Putatively sarcosine | [123] |
| Nocardia mediterranie THS 1 | - | - | India | AMPA | [124] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Kumar, V.; Gill, J.P.K.; Datta, S.; Singh, S.; Dhaka, V.; Kapoor, D.; Wani, A.B.; Dhanjal, D.S.; Kumar, M.; et al. Herbicide Glyphosate: Toxicity and Microbial Degradation. Int. J. Environ. Res. Public Health 2020, 17, 7519. https://doi.org/10.3390/ijerph17207519
Singh S, Kumar V, Gill JPK, Datta S, Singh S, Dhaka V, Kapoor D, Wani AB, Dhanjal DS, Kumar M, et al. Herbicide Glyphosate: Toxicity and Microbial Degradation. International Journal of Environmental Research and Public Health. 2020; 17(20):7519. https://doi.org/10.3390/ijerph17207519
Chicago/Turabian StyleSingh, Simranjeet, Vijay Kumar, Jatinder Pal Kaur Gill, Shivika Datta, Satyender Singh, Vaishali Dhaka, Dhriti Kapoor, Abdul Basit Wani, Daljeet Singh Dhanjal, Manoj Kumar, and et al. 2020. "Herbicide Glyphosate: Toxicity and Microbial Degradation" International Journal of Environmental Research and Public Health 17, no. 20: 7519. https://doi.org/10.3390/ijerph17207519
APA StyleSingh, S., Kumar, V., Gill, J. P. K., Datta, S., Singh, S., Dhaka, V., Kapoor, D., Wani, A. B., Dhanjal, D. S., Kumar, M., Harikumar, S. L., & Singh, J. (2020). Herbicide Glyphosate: Toxicity and Microbial Degradation. International Journal of Environmental Research and Public Health, 17(20), 7519. https://doi.org/10.3390/ijerph17207519