Retrospective Assessment of the Antigenic Similarity of Egg-Propagated and Cell Culture-Propagated Reference Influenza Viruses as Compared with Circulating Viruses across Influenza Seasons 2002–2003 to 2017–2018

 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
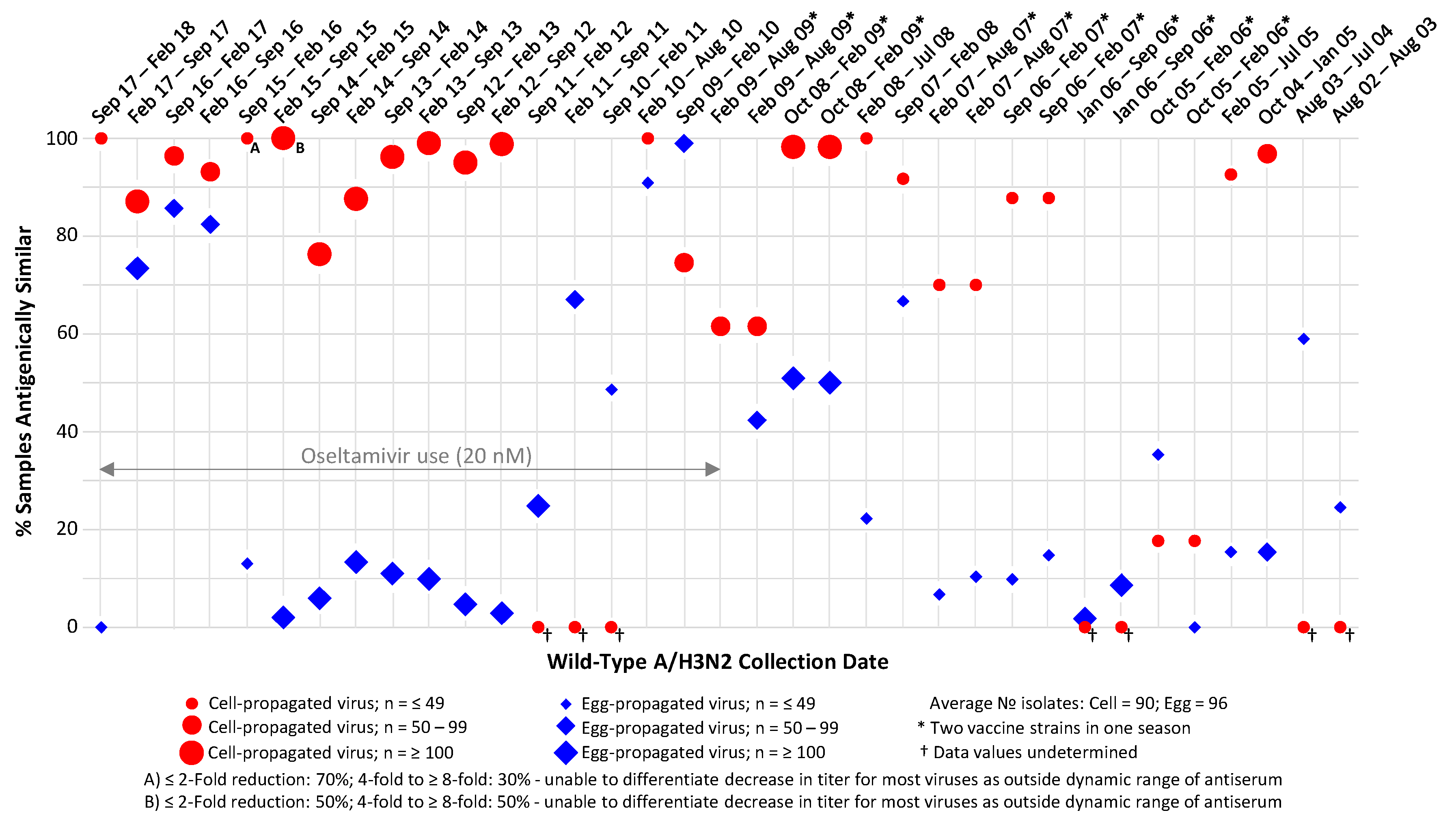
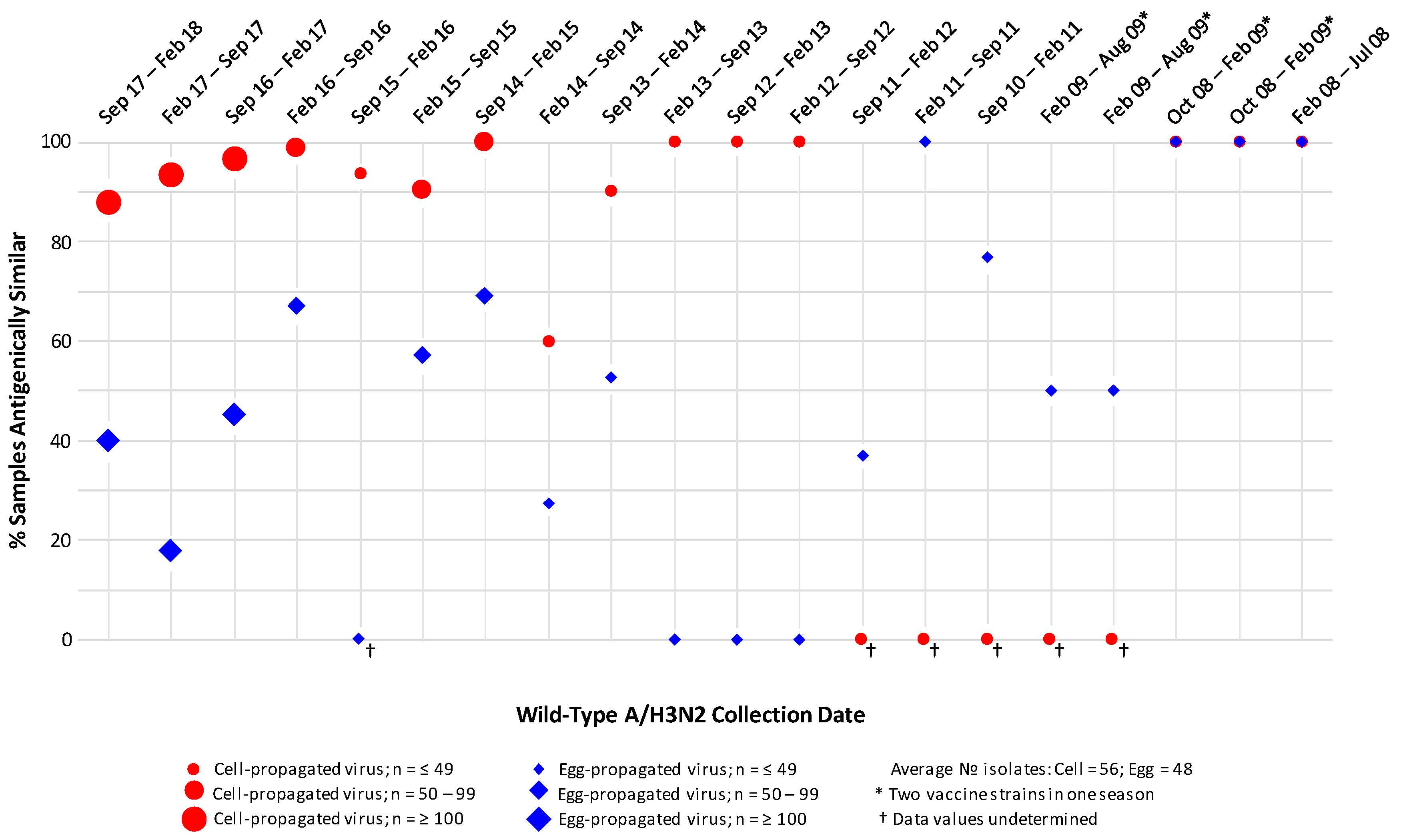
3.1. A/H3N2 Analysis
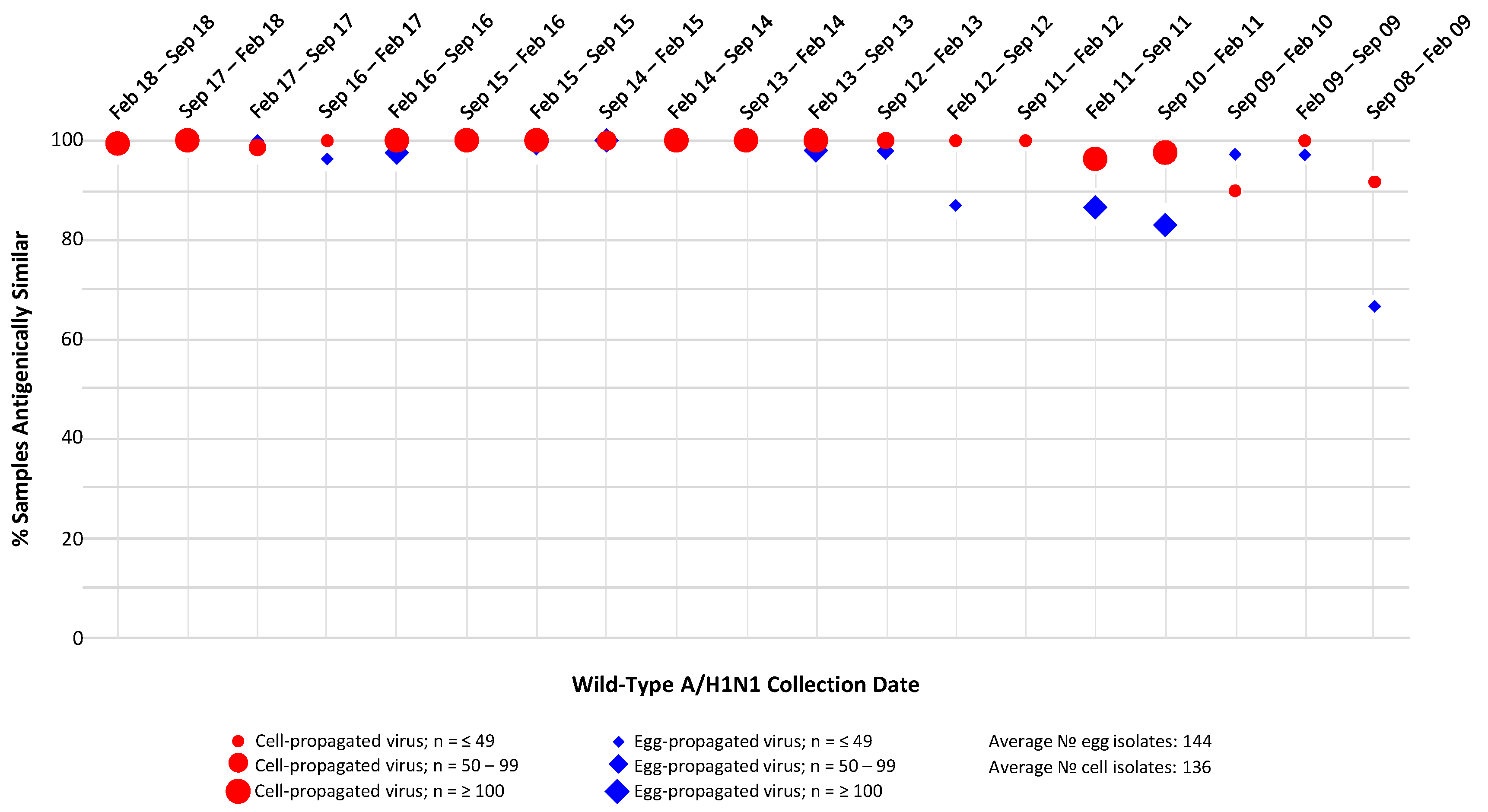
3.2. A/H1N1 Analysis
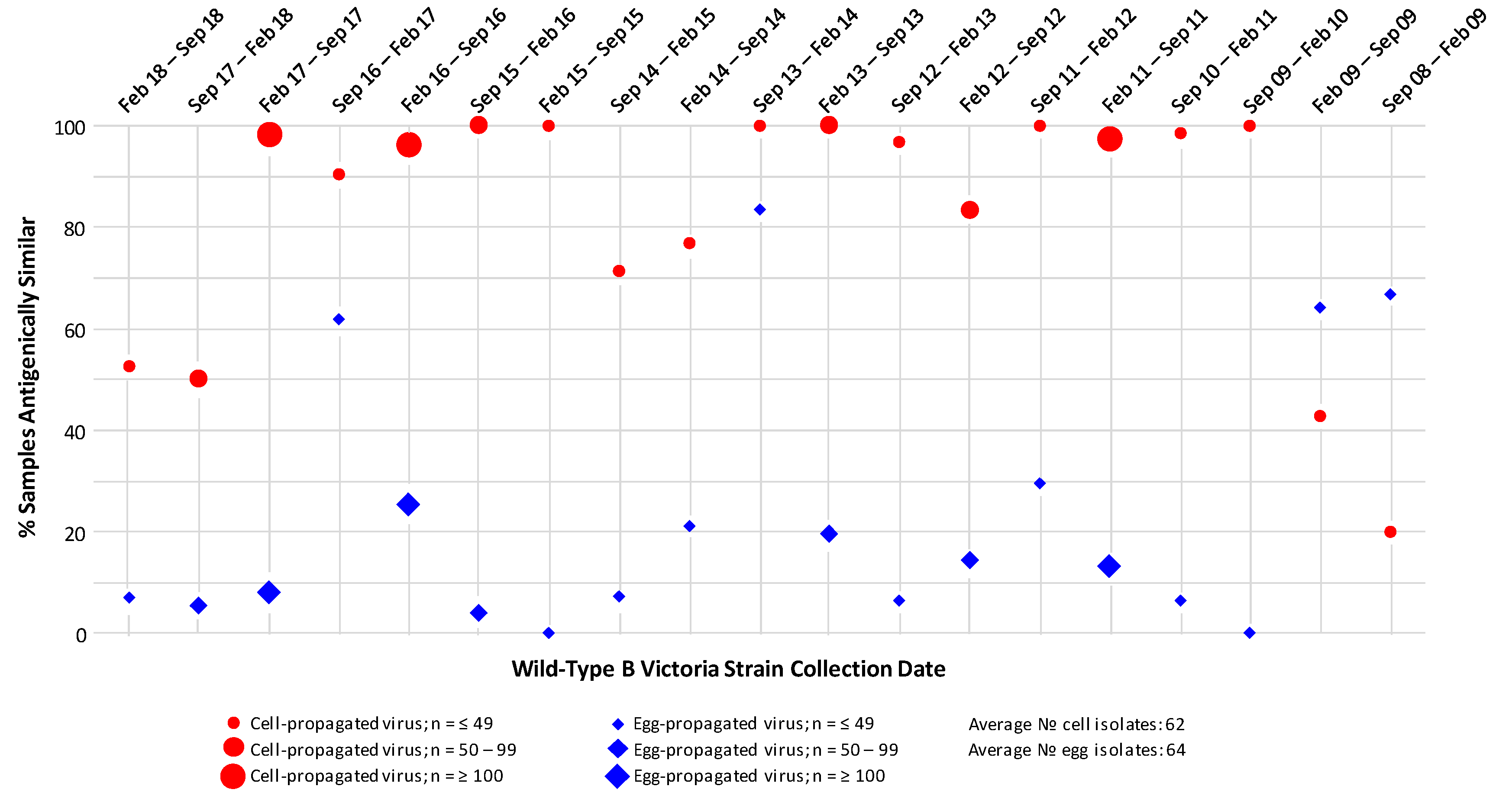
3.3. B/Victoria Strain Analysis
3.4. B/Yamagata Strain Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Fact Sheets. Influenza (Seasonal). Available online: Http://www.Who.Int/en/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 22 May 2020).
- US Government. Centers for Disease Control and Prevention. Influenza. Vaccine Effectiveness—How Well Does the Flu Vaccine Work? Available online: Https://www.Cdc.Gov/flu/vaccines-work/vaccineeffect.Htm (accessed on 22 May 2020).
- Nichols, M.K.; Andrew, M.K.; Hatchette, T.F.; Ambrose, A.; Boivin, G.; Bowie, W.; Chit, A.; Santos, G.D.; ElSherif, M.; Green, K.; et al. Influenza vaccine effectiveness to prevent influenza-related hospitalizations and serious outcomes in canadian adults over the 2011/12 through 2013/14 influenza seasons: A pooled analysis from the canadian immunization research network (cirn) serious outcomes surveillance (sos network). Vaccine 2018, 36, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Restivo, V.; Costantino, C.; Bono, S.; Maniglia, M.; Marchese, V.; Ventura, G.; Casuccio, A.; Tramuto, F.; Vitale, F. Influenza vaccine effectiveness among high-risk groups: A systematic literature review and meta-analysis of case-control and cohort studies. Hum. Vaccines Immunother. 2018, 14, 724–735. [Google Scholar] [CrossRef]
- World Health Organization. Influenza. Who Recommendations on the Composition of Influenza Virus Vaccines. Available online: Http://www.Who.Int/influenza/vaccines/virus/recommendations/en/ (accessed on 22 May 2020).
- Belongia, E.A.; McLean, H.Q. Influenza vaccine effectiveness: Defining the h3n2 problem. Clin. Infect. Dis. 2019, 69, 1817–1823. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C.; Ghedin, E.; Miller, N.; Taylor, J.; Bao, Y.; St George, K.; Grenfell, B.T.; Salzberg, S.L.; Fraser, C.M.; Lipman, D.J.; et al. Whole-genome analysis of human influenza a virus reveals multiple persistent lineages and reassortment among recent h3n2 viruses. PLoS Biol. 2005, 3, e300. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.Y.; Serohijos, A.W.; Choi, J.M.; Shakhnovich, E.I.; Pekosz, A. Influenza a h1n1 pandemic strain evolution—Divergence and the potential for antigenic drift variants. PLoS ONE 2014, 9, e93632. [Google Scholar] [CrossRef]
- Maljkovic Berry, I.; Melendrez, M.C.; Li, T.; Hawksworth, A.W.; Brice, G.T.; Blair, P.J.; Halsey, E.S.; Williams, M.; Fernandez, S.; Yoon, I.K.; et al. Frequency of influenza h3n2 intra-subtype reassortment: Attributes and implications of reassortant spread. BMC Biol. 2016, 14, 117. [Google Scholar] [CrossRef]
- Rejmanek, D.; Hosseini, P.R.; Mazet, J.A.K.; Daszak, P.; Goldstein, T. Evolutionary dynamics and global diversity of influenza a virus. J. Virol. 2015, 89, 10993–11001. [Google Scholar] [CrossRef]
- Schweiger, B.; Bruns, L.; Meixenberger, K. Reassortment between human a(h3n2) viruses is an important evolutionary mechanism. Vaccine 2006, 24, 6683–6690. [Google Scholar] [CrossRef]
- Tewawong, N.; Prachayangprecha, S.; Vichiwattana, P.; Korkong, S.; Klinfueng, S.; Vongpunsawad, S.; Thongmee, T.; Theamboonlers, A.; Poovorawan, Y. Assessing antigenic drift of seasonal influenza a(h3n2) and a(h1n1)pdm09 viruses. PLoS ONE 2015, 10, e0139958. [Google Scholar] [CrossRef]
- Monto, A.S.; Malosh, R.E.; Petrie, J.G.; Martin, E.T. The doctrine of original antigenic sin: Separating good from evil. J. Infect. Dis. 2017, 215, 1782–1788. [Google Scholar] [CrossRef]
- Tan, Y.C.; Blum, L.K.; Kongpachith, S.; Ju, C.H.; Cai, X.; Lindstrom, T.M.; Sokolove, J.; Robinson, W.H. High-throughput sequencing of natively paired antibody chains provides evidence for original antigenic sin shaping the antibody response to influenza vaccination. Clin. Immunol. 2014, 151, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Belongia, E.A.; Simpson, M.D.; King, J.P.; Sundaram, M.E.; Kelley, N.S.; Osterholm, M.T.; McLean, H.Q. Variable influenza vaccine effectiveness by subtype: A systematic review and meta-analysis of test-negative design studies. Lancet Infect. Dis. 2016, 16, 942–951. [Google Scholar] [CrossRef]
- Garten, R.; Blanton, L.; Elal, A.I.A.; Alabi, N.; Barnes, J.; Biggerstaff, M.; Brammer, L.; Budd, A.P.; Burns, E.; Cummings, C.N.; et al. Update: Influenza activity in the united states during the 2017–18 season and composition of the 2018–19 influenza vaccine. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Zhou, H.; Ye, D.; Kemble, G.; Jin, H. Improvement of influenza a/fujian/411/02 (h3n2) virus growth in embryonated chicken eggs by balancing the hemagglutinin and neuraminidase activities, using reverse genetics. J. Virol. 2005, 79, 6763–6771. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.; Chen, L.M.; Carney, P.J.; Garten, R.; Foust, A.; Le, J.; Pokorny, B.A.; Manojkumar, R.; Silverman, J.; Devis, R.; et al. Receptor specificity of influenza a h3n2 viruses isolated in mammalian cells and embryonated chicken eggs. J. Virol. 2010, 84, 8287–8299. [Google Scholar] [CrossRef]
- Kishida, N.; Fujisaki, S.; Yokoyama, M.; Sato, H.; Saito, R.; Ikematsu, H.; Xu, H.; Takashita, E.; Tashiro, M.; Takao, S.; et al. Evaluation of influenza virus a/h3n2 and b vaccines on the basis of cross-reactivity of postvaccination human serum antibodies against influenza viruses a/h3n2 and b isolated in mdck cells and embryonated hen eggs. Clin. Vaccine Immunol. 2012, 19, 897–908. [Google Scholar] [CrossRef]
- Skowronski, D.M.; De Serres, G. Role of egg-adaptation mutations in low influenza a(h3n2) vaccine effectiveness during the 2012–13 season. Clin. Infect. Dis. 2018, 67, 1474–1476. [Google Scholar] [CrossRef]
- Skowronski, D.M.; Janjua, N.Z.; De Serres, G.; Sabaiduc, S.; Eshaghi, A.; Dickinson, J.A.; Fonseca, K.; Winter, A.L.; Gubbay, J.B.; Krajden, M.; et al. Low 2012–13 influenza vaccine effectiveness associated with mutation in the egg-adapted h3n2 vaccine strain not antigenic drift in circulating viruses. PLoS ONE 2014, 9, e92153. [Google Scholar] [CrossRef]
- Wu, N.C.; Zost, S.J.; Thompson, A.J.; Oyen, D.; Nycholat, C.M.; McBride, R.; Paulson, J.C.; Hensley, S.E.; Wilson, I.A. A structural explanation for the low effectiveness of the seasonal influenza h3n2 vaccine. PLoS Pathog. 2017, 13, e1006682. [Google Scholar] [CrossRef]
- Zost, S.J.; Parkhouse, K.; Gumina, M.E.; Kim, K.; Perez, S.D.; Wilson, P.C.; Treanor, J.J.; Sant, A.J.; Cobey, S.; Hensley, S.E. Contemporary h3n2 influenza viruses have a glycosylation site that alters binding of antibodies elicited by egg-adapted vaccine strains. Proc. Natl. Acad. Sci. USA 2017, 114, 12578–12583. [Google Scholar] [CrossRef]
- Dunkle, L.M.; Izikson, R.; Patriarca, P.; Goldenthal, K.L.; Muse, D.; Callahan, J.; Cox, M.M.J.; Team, P.S.C.S. Efficacy of recombinant influenza vaccine in adults 50 years of age or older. N. Engl. J. Med. 2017, 376, 2427–2436. [Google Scholar] [CrossRef]
- Boikos, C.; Sylvester, G.; Sampalis, J.; Mansi, J. Effectiveness of the Cell Culture- and Egg-Derived, Seasonal Influenza Vaccine during the 2017–2018 Northern Hemisphere Influenza Season. Poster Presentation. National Foundation for Infectious Disease, 2018 Clinical Vaccinology Course, Bethesda Md, USA. Available online: Http://www.Nfid.Org/professional-education/conferences. (accessed on 22 May 2020).
- Izurieta, H.S.; Chillarige, Y.; Kelman, J.; Wei, Y.; Lu, Y.; Xu, W.; Lu, M.; Pratt, D.; Chu, S.; Wernecke, M.; et al. Relative effectiveness of cell-cultured and egg-based influenza vaccines among the u.S. Elderly, 2017–2018. J. Infect. Dis. 2018, 220, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Francis Crick Institute. Worldwide Influenza Centre Annual & Interim Reports. Available online: Https://www.Crick.Ac.Uk/partnerships/worldwide-influenza-centre/annual-and-interim-reports (accessed on 22 May 2020).
- van Baalen, C.A.; Els, C.; Sprong, L.; van Beek, R.; van der Vries, E.; Osterhaus, A.D.; Rimmelzwaan, G.F. Detection of nonhemagglutinating influenza a(h3) viruses by enzyme-linked immunosorbent assay in quantitative influenza virus culture. J. Clin. Microbiol. 2014, 52, 1672–1677. [Google Scholar] [CrossRef]
- US Government. Centers for Disease Control and Prevention. Seasonal Influenza. Understanding Influenza Viruses. Antigenic Characterization. Available online: Https://www.Cdc.Gov/flu/professionals/laboratory/antigenic.Htm (accessed on 22 May 2020).
- Vijaykrishna, D.; Holmes, E.C.; Joseph, U.; Fourment, M.; Su, Y.C.; Halpin, R.; Lee, R.T.; Deng, Y.M.; Gunalan, V.; Lin, X.; et al. The contrasting phylodynamics of human influenza b viruses. Elife 2015, 4, e05055. [Google Scholar] [CrossRef] [PubMed]
- The Lancet Respiratory, M. Elderly at risk as severe influenza season looms large. Lancet Respir. Med. 2018, 6, 1. [Google Scholar] [CrossRef]
- Thompson, W.W.; Shay, D.K.; Weintraub, E.; Brammer, L.; Bridges, C.B.; Cox, N.J.; Fukuda, K. Influenza-associated hospitalizations in the united states. Jama 2004, 292, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Blanton, L.; Alabi, N.; Mustaquim, D.; Taylor, C.; Kniss, K.; Kramer, N.; Budd, A.; Garg, S.; Cummings, C.N.; Chung, J.; et al. Update: Influenza activity in the united states during the 2016–17 season and composition of the 2017–18 influenza vaccine. MMWR Morb. Mortal. Wkly. Rep. 2017, 66, 668–676. [Google Scholar] [CrossRef]
- Bonomo, M.E.; Deem, M.W. Predicting influenza h3n2 vaccine efficacy from evolution of the dominant epitope. Clin. Infect. Dis. 2018, 67, 1129–1131. [Google Scholar] [CrossRef]
- Gupta, V.; Earl, D.J.; Deem, M.W. Quantifying influenza vaccine efficacy and antigenic distance. Vaccine 2006, 24, 3881–3888. [Google Scholar] [CrossRef]
- Paules, C.I.; Sullivan, S.G.; Subbarao, K.; Fauci, A.S. Chasing seasonal influenza—The need for a universal influenza vaccine. N. Engl. J. Med. 2018, 378, 7–9. [Google Scholar] [CrossRef]
- Sullivan, S.G.; Chilver, M.B.; Carville, K.S.; Deng, Y.M.; Grant, K.A.; Higgins, G.; Komadina, N.; Leung, V.K.; Minney-Smith, C.A.; Teng, D.; et al. Low interim influenza vaccine effectiveness, australia, 1 may to 24 september 2017. Euro Surveill. 2017, 22, 17–00707. [Google Scholar] [CrossRef] [PubMed]
- Seqirus USA Inc. News. Seqirus Announces Next Major Advancement in Cell-Based Influenza Vaccine Technology. Available online: Https://www.Seqirus.Com/news/seqirus-announces-next-major-advancement-in-cell-based-influenza-vaccine-technology (accessed on 22 May 2020).
- Bruxvoort, K.J.; Luo, Y.; Ackerson, B.; Tanenbaum, H.C.; Sy, L.S.; Gandhi, A.; Tseng, H.F. Comparison of vaccine effectiveness against influenza hospitalization of cell-based and egg-based influenza vaccines, 2017–2018. Vaccine 2019, 37, 5807–5811. [Google Scholar] [CrossRef] [PubMed]
- Klein, N.P.; Fireman, B.; Goddard, K.; Zerbo, O.; Asher, J.; Zhou, J.; King, J.; Lewis, N. Vaccine effectiveness of cell-culture relative to egg-based inactivated influenza vaccine during the 2017–18 influenza season. PLoS ONE 2020, 15, e0229279. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Influenza Virus Characterisation. Summary, Europe. Report November 2014. Available online: Https://ecdc.Europa.Eu/en/seasonal-influenza/surveillance-and-disease-data/influenza-virus-characterisation (accessed on 22 May 2020).
- Mogling, R.; Richard, M.J.; Vliet, S.V.; Beek, R.V.; Schrauwen, E.J.A.; Spronken, M.I.; Rimmelzwaan, G.F.; Fouchier, R.A.M. Neuraminidase-mediated haemagglutination of recent human influenza a(h3n2) viruses is determined by arginine 150 flanking the neuraminidase catalytic site. J. Gen. Virol. 2017, 98, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.P.; Gregory, V.; Collins, P.; Kloess, J.; Wharton, S.; Cattle, N.; Lackenby, A.; Daniels, R.; Hay, A. Neuraminidase receptor binding variants of human influenza a(h3n2) viruses resulting from substitution of aspartic acid 151 in the catalytic site: A role in virus attachment? J. Virol. 2010, 84, 6769–6781. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S.; Gouma, S.; Parkhouse, K.; Chambers, B.S.; Ertl, H.C.; Schmader, K.E.; Halpin, R.A.; Lin, X.; Stockwell, T.B.; Das, S.R.; et al. Poor immunogenicity, not vaccine strain egg adaptation, may explain the low h3n2 influenza vaccine effectiveness in 2012–2013. Clin. Infect. Dis. 2018, 67, 327–333. [Google Scholar] [CrossRef]
- Liu, F.; Tzeng, W.P.; Horner, L.; Kamal, R.P.; Tatum, H.R.; Blanchard, E.G.; Xu, X.; York, I.; Tumpey, T.M.; Katz, J.M.; et al. Both immune priming and egg-adaptation in the vaccine influence antibody responses to circulating a(h1n1)pdm09 viruses following influenza vaccination in adults. J. Infect. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Alvarez, J.J.S.; Ng, S.H.; Nielsen, R.; Zhai, W. Passage adaptation correlates with the reduced efficacy of the influenza vaccine. Clin. Infect. Dis. 2018, 69, 1198–1204. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Influenza Season | Reference Virus | ≤2-Fold | 4-Fold |
|---|---|---|---|
| Feburary 2016–September 2016 | Cell-based | 69% | 24% |
| Feburary 2016–September 2016 | Egg-based | 35% | 47% |
| September 2016–Feburary 2017 | Cell-based | 52% | 45% |
| September 2016–Feburary 2017 | Egg-based | 38% | 48% |
| Clade | Reference Virus | ≤2-Fold | 4-Fold |
|---|---|---|---|
| 3C.2a | Cell-based | 60% | 31% |
| 3C.2a | Egg-based | 19% | 50% |
| 3C.3a | Cell-based | 76% | 20% |
| 3C.3a | Egg-based | 64% | 36% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajaram, S.; Suphaphiphat, P.; van Boxmeer, J.; Haag, M.; Leav, B.; Iheanacho, I.; Kistler, K.; Ortiz de Lejarazu, R. Retrospective Assessment of the Antigenic Similarity of Egg-Propagated and Cell Culture-Propagated Reference Influenza Viruses as Compared with Circulating Viruses across Influenza Seasons 2002–2003 to 2017–2018. Int. J. Environ. Res. Public Health 2020, 17, 5423. https://doi.org/10.3390/ijerph17155423
Rajaram S, Suphaphiphat P, van Boxmeer J, Haag M, Leav B, Iheanacho I, Kistler K, Ortiz de Lejarazu R. Retrospective Assessment of the Antigenic Similarity of Egg-Propagated and Cell Culture-Propagated Reference Influenza Viruses as Compared with Circulating Viruses across Influenza Seasons 2002–2003 to 2017–2018. International Journal of Environmental Research and Public Health. 2020; 17(15):5423. https://doi.org/10.3390/ijerph17155423
Chicago/Turabian StyleRajaram, Sankarasubramanian, Pirada Suphaphiphat, Josephine van Boxmeer, Mendel Haag, Brett Leav, Ike Iheanacho, Kristin Kistler, and Raúl Ortiz de Lejarazu. 2020. "Retrospective Assessment of the Antigenic Similarity of Egg-Propagated and Cell Culture-Propagated Reference Influenza Viruses as Compared with Circulating Viruses across Influenza Seasons 2002–2003 to 2017–2018" International Journal of Environmental Research and Public Health 17, no. 15: 5423. https://doi.org/10.3390/ijerph17155423
APA StyleRajaram, S., Suphaphiphat, P., van Boxmeer, J., Haag, M., Leav, B., Iheanacho, I., Kistler, K., & Ortiz de Lejarazu, R. (2020). Retrospective Assessment of the Antigenic Similarity of Egg-Propagated and Cell Culture-Propagated Reference Influenza Viruses as Compared with Circulating Viruses across Influenza Seasons 2002–2003 to 2017–2018. International Journal of Environmental Research and Public Health, 17(15), 5423. https://doi.org/10.3390/ijerph17155423