The Role of miRNAs in Zearalenone-Promotion of TM3 Cell Proliferation


Abstract
:1. Introduction
2. Material and Methods
2.1. Reagents and Antibodies
2.2. Cell Culture
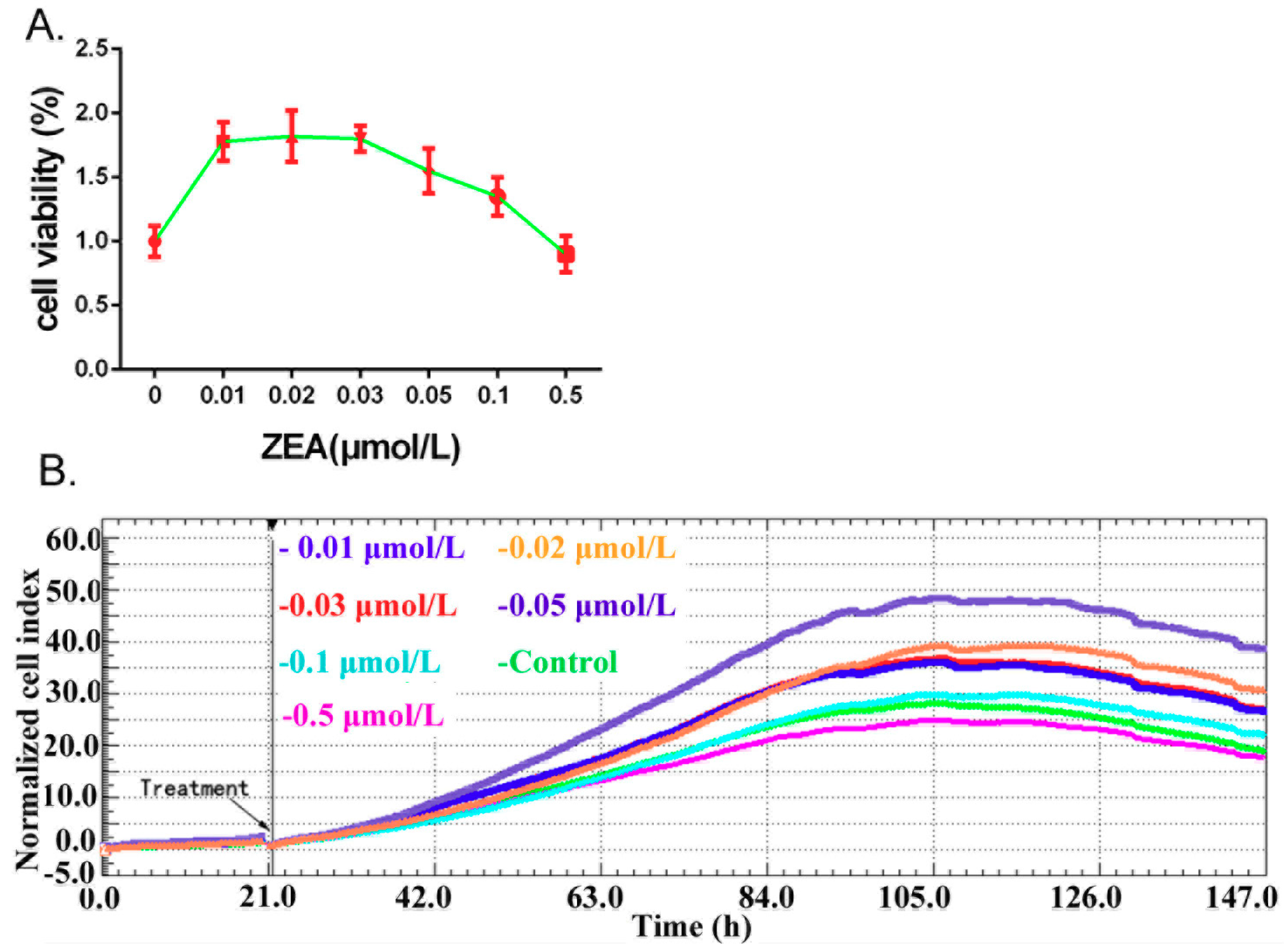
2.3. Cell Proliferation Analysis
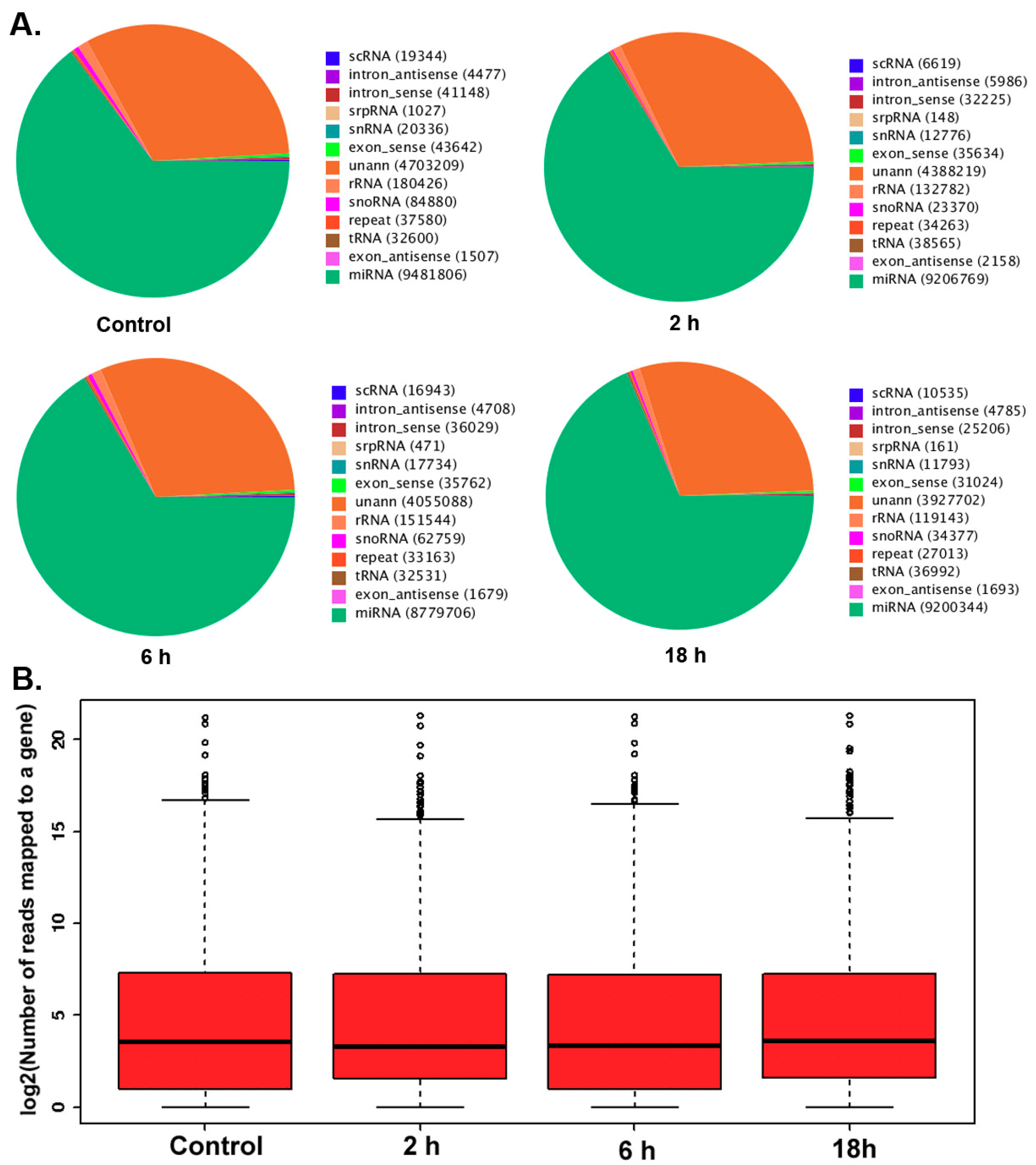
2.4. Analyzing the Differential Expression MicroRNA
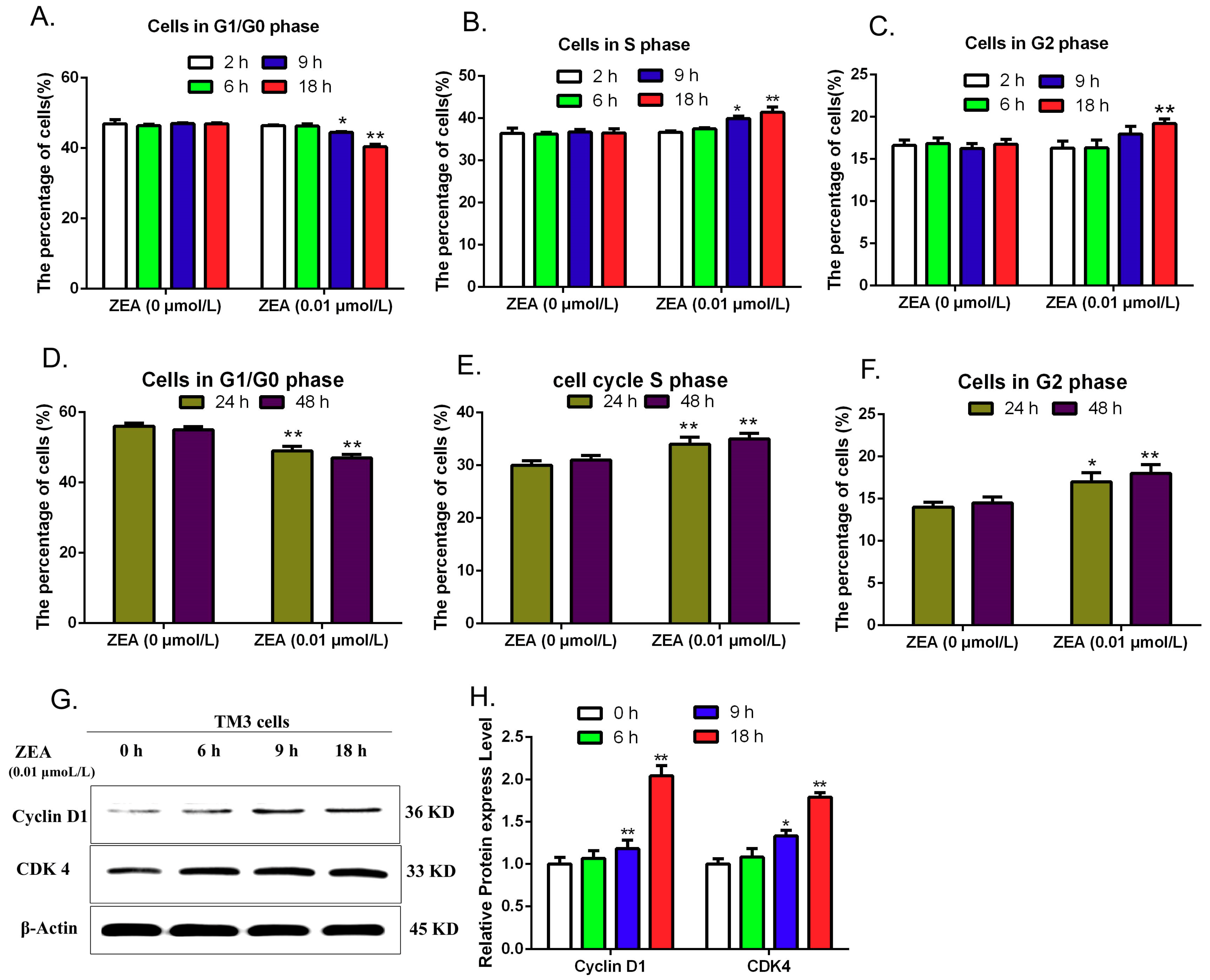
2.5. Analyzing Cell Cycle Distribution
2.6. Confirmation of miRNA Expression by Quantitative RT-PCR
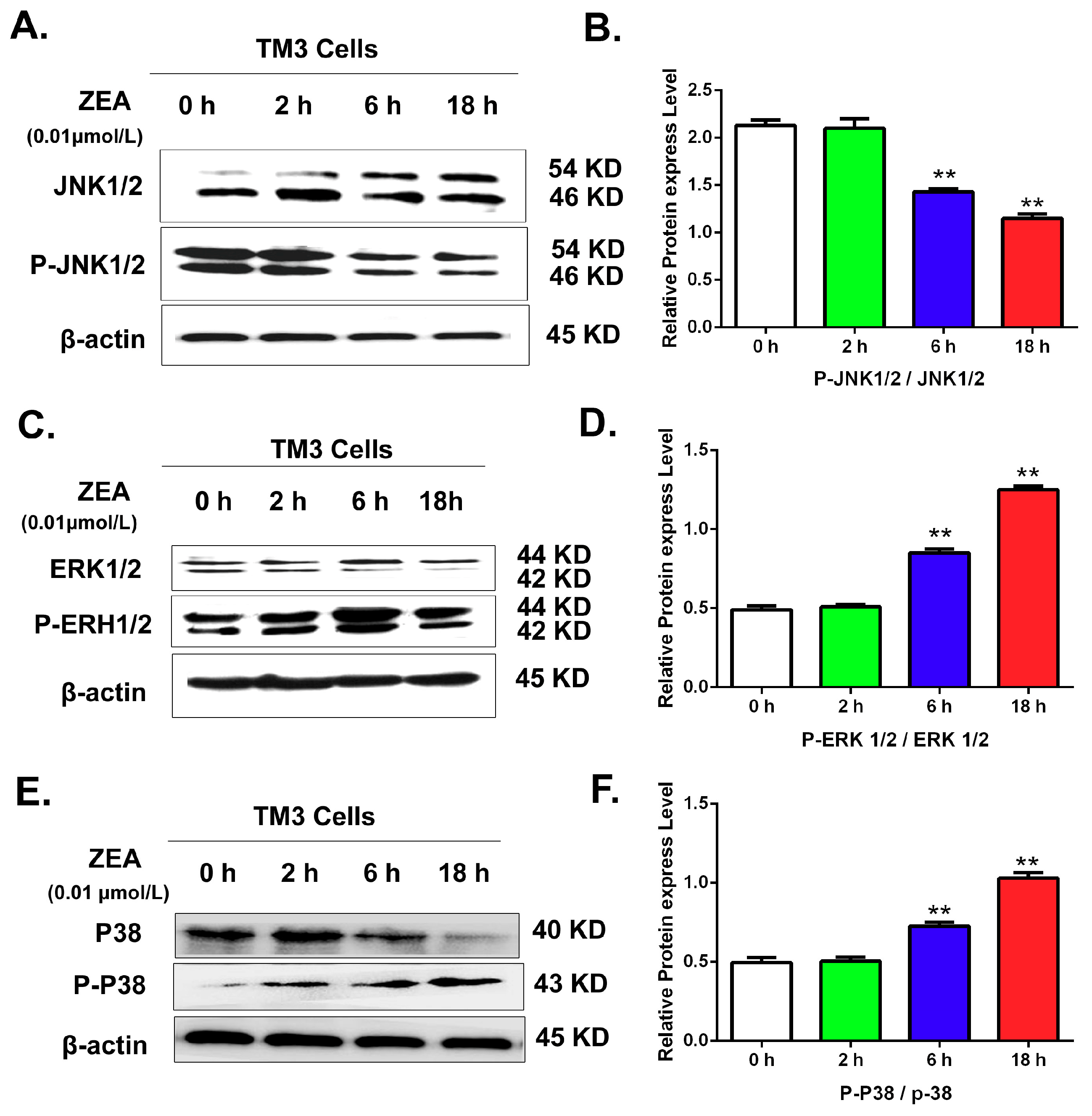
2.7. Western Blotting Analysis
2.8. Statistical Analysis
3. Results
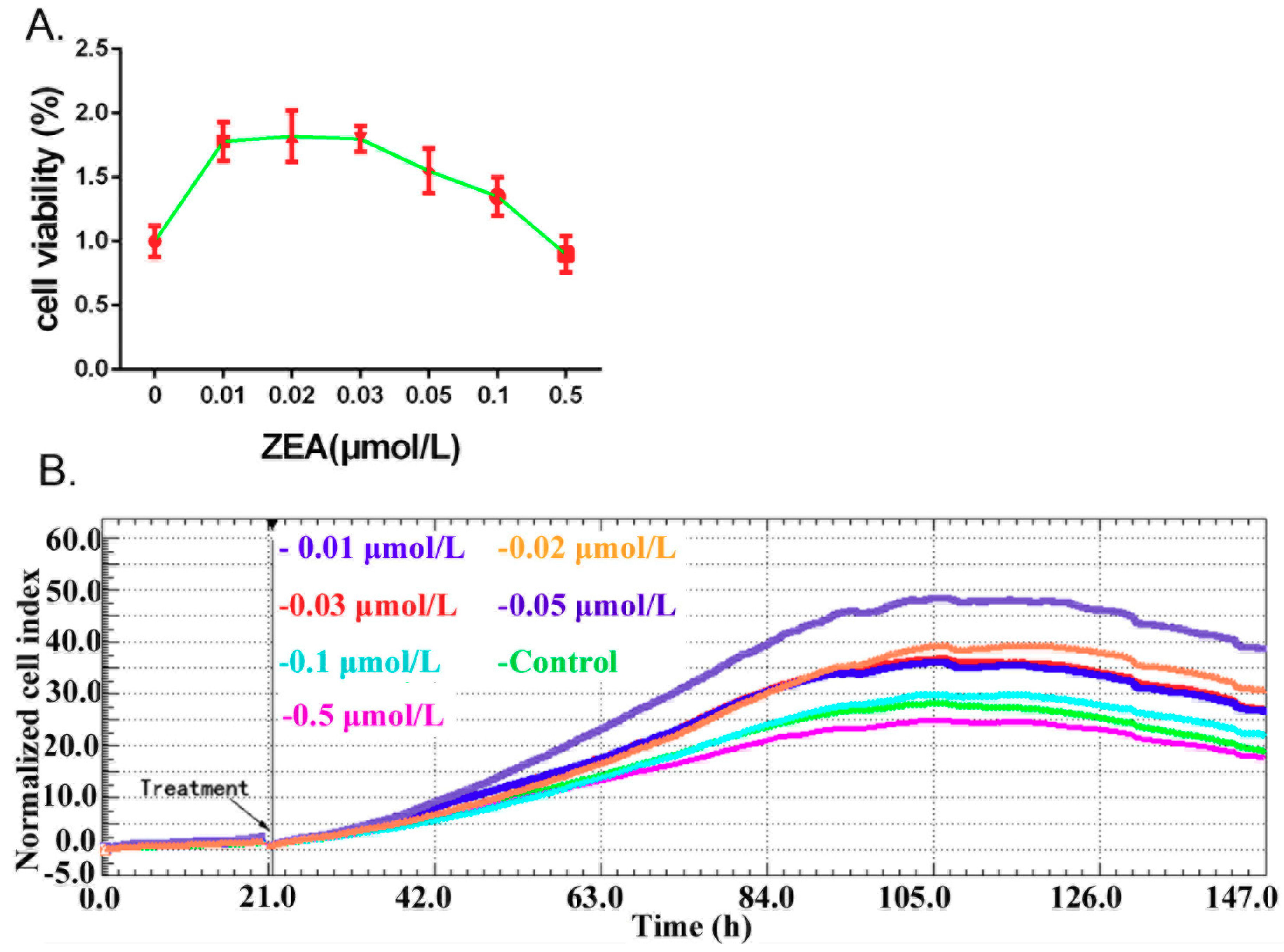
3.1. ZEA Can Stimulate the TM3 Cells Proliferation
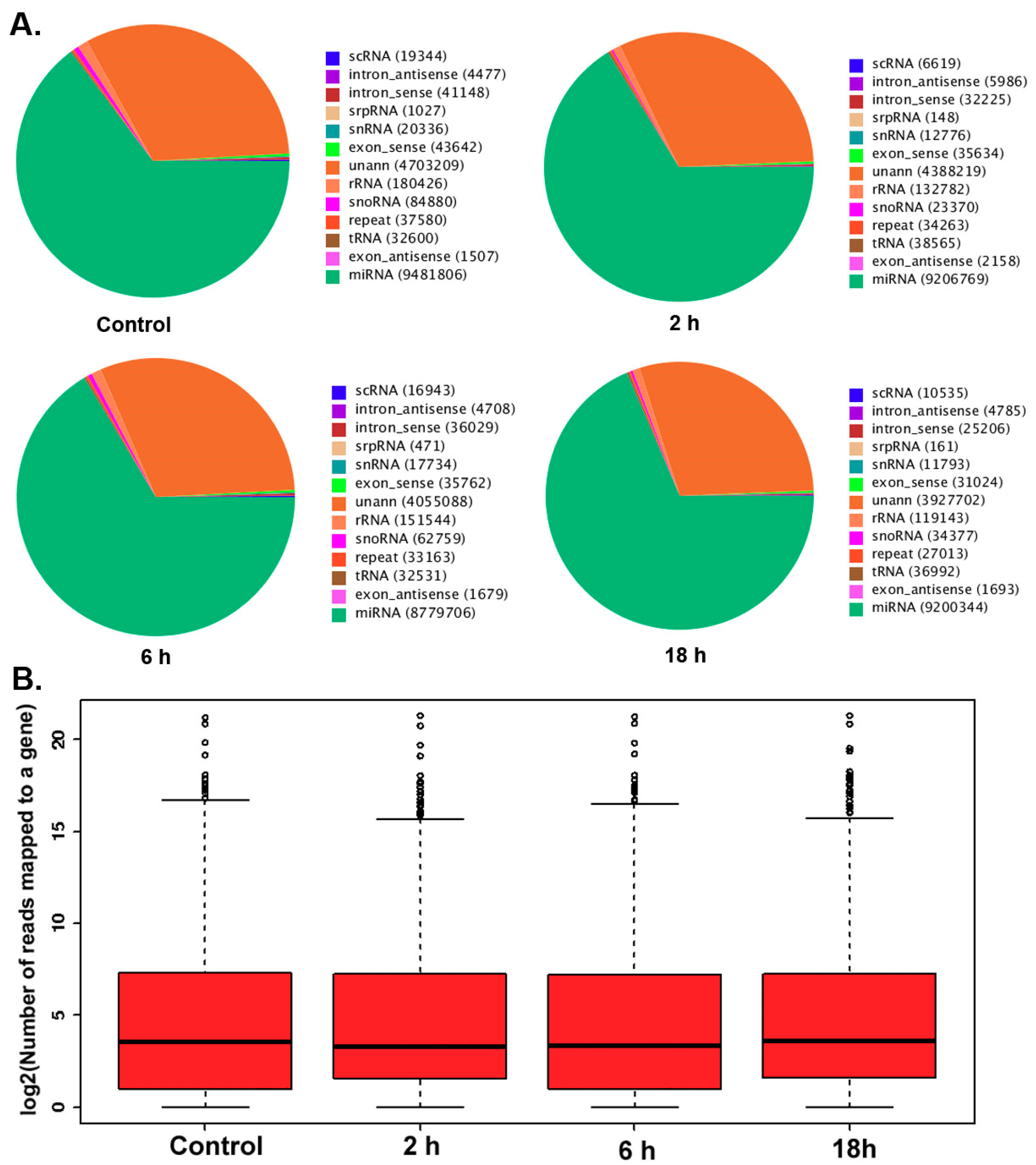
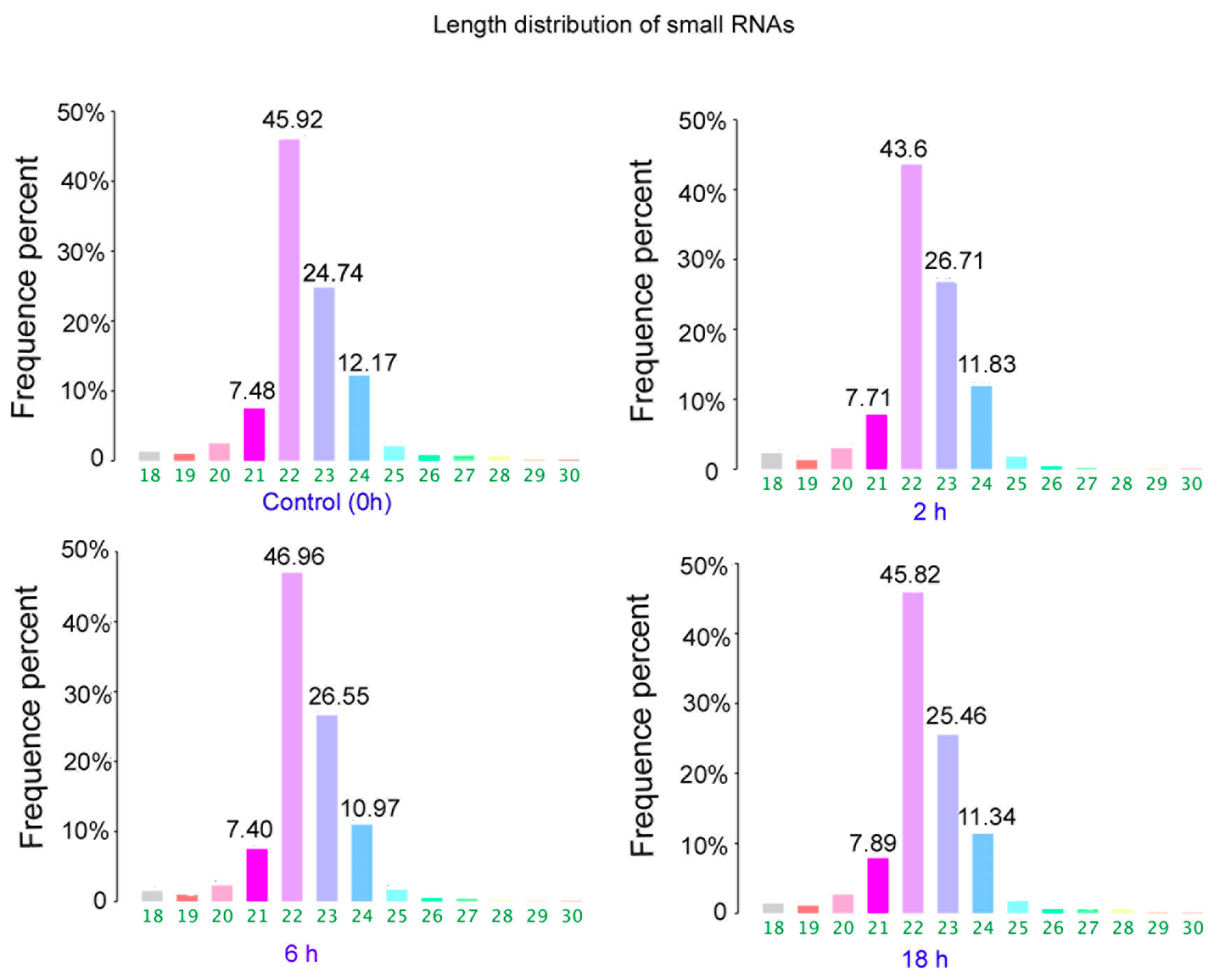
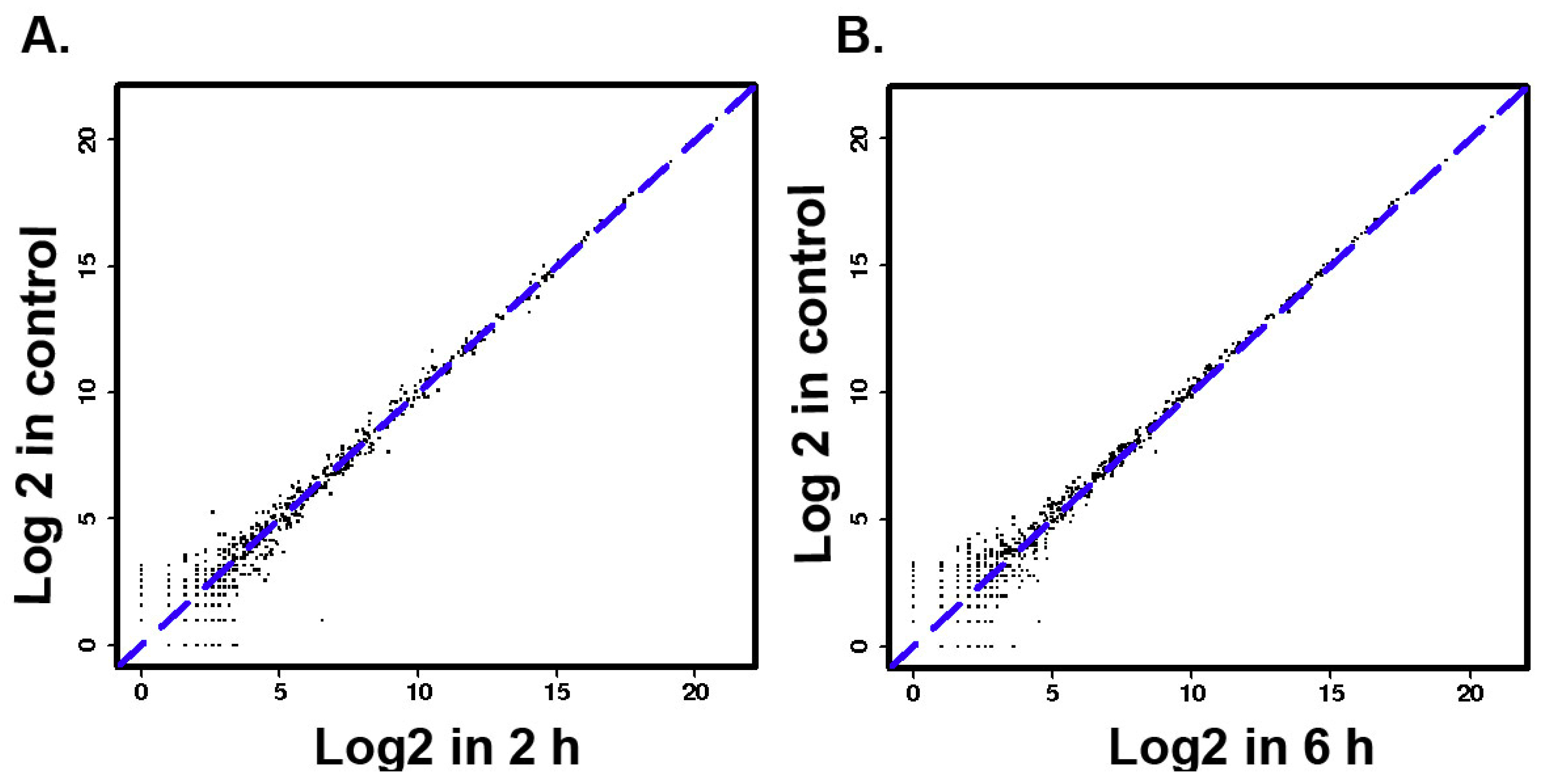
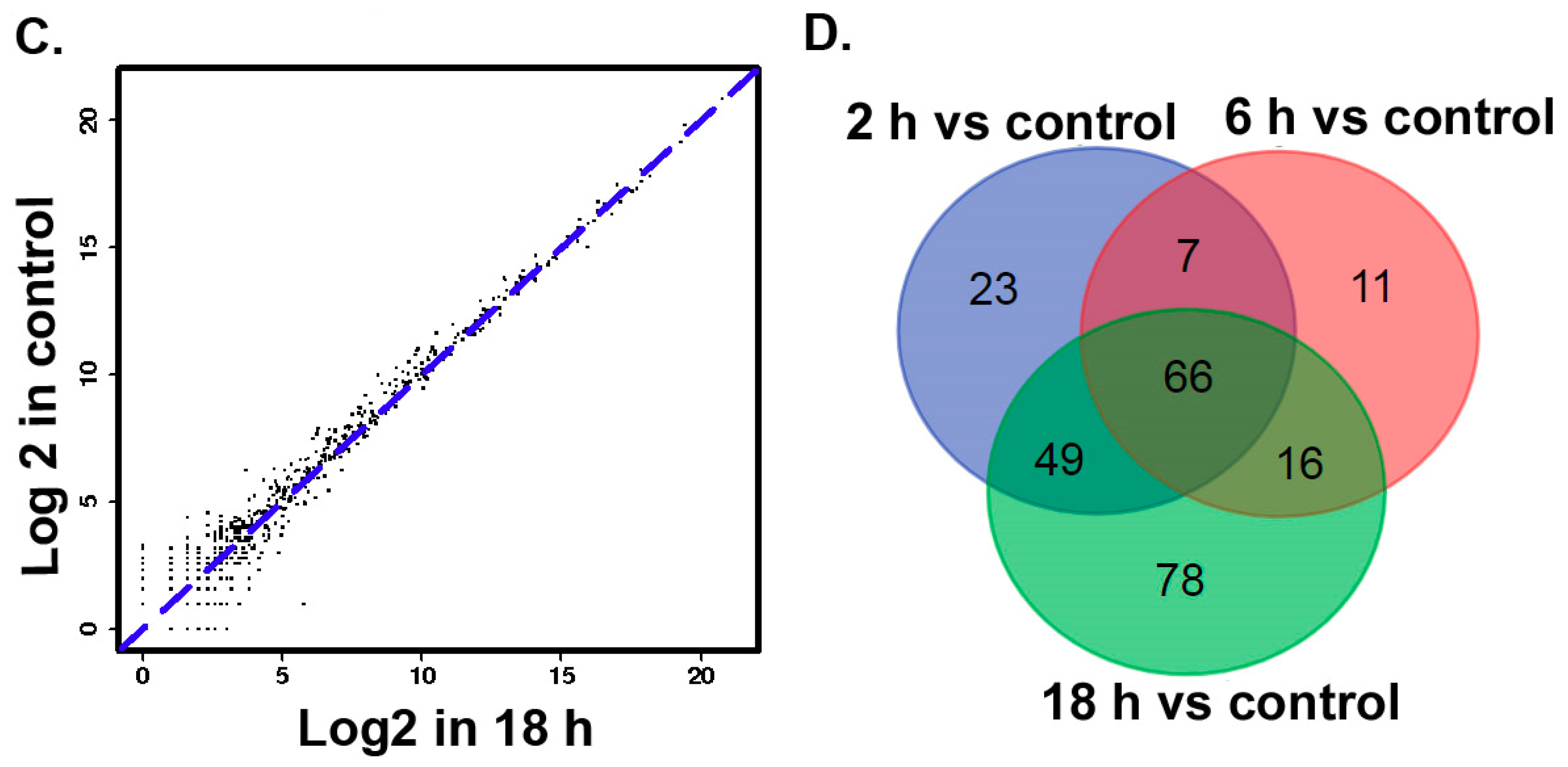
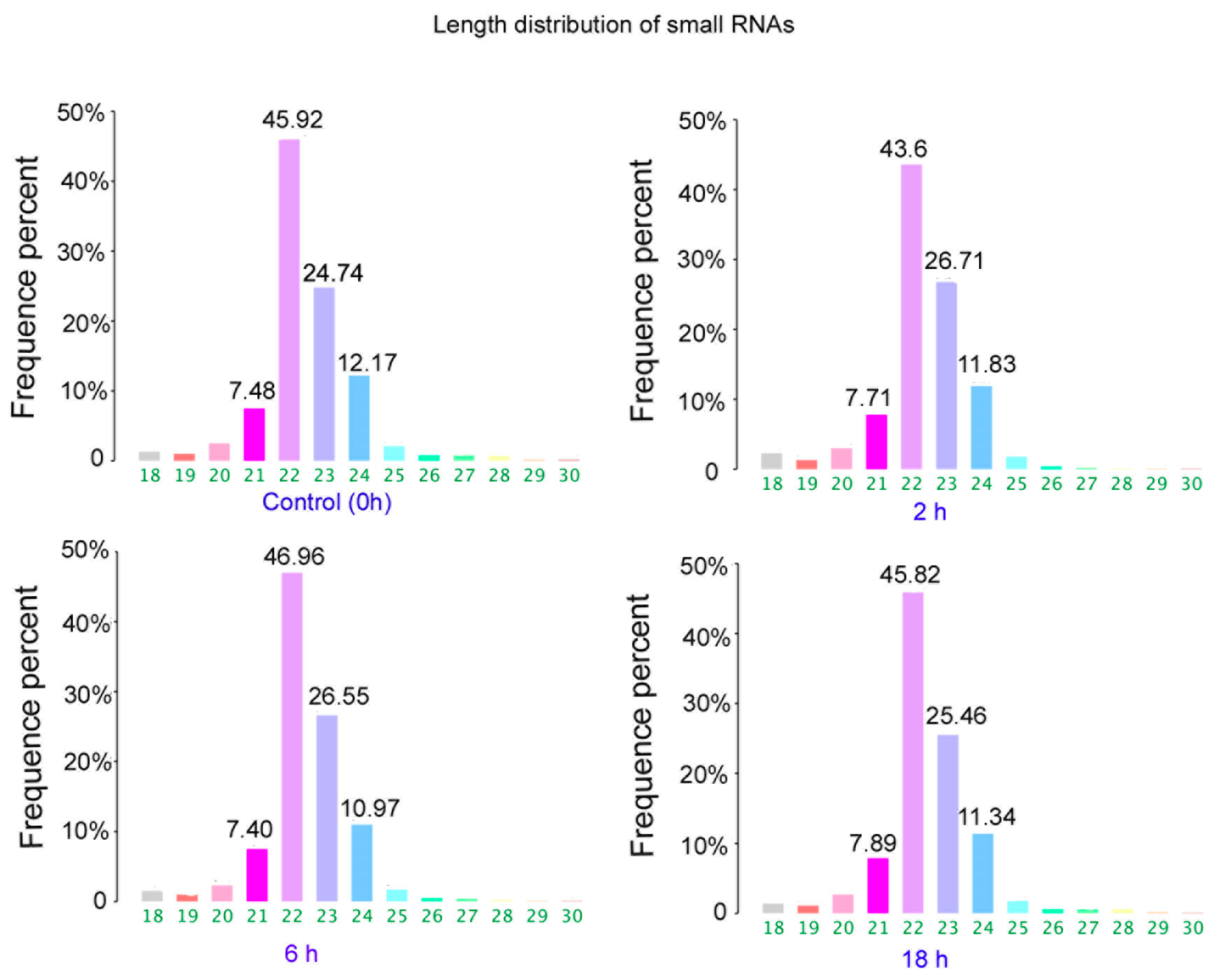
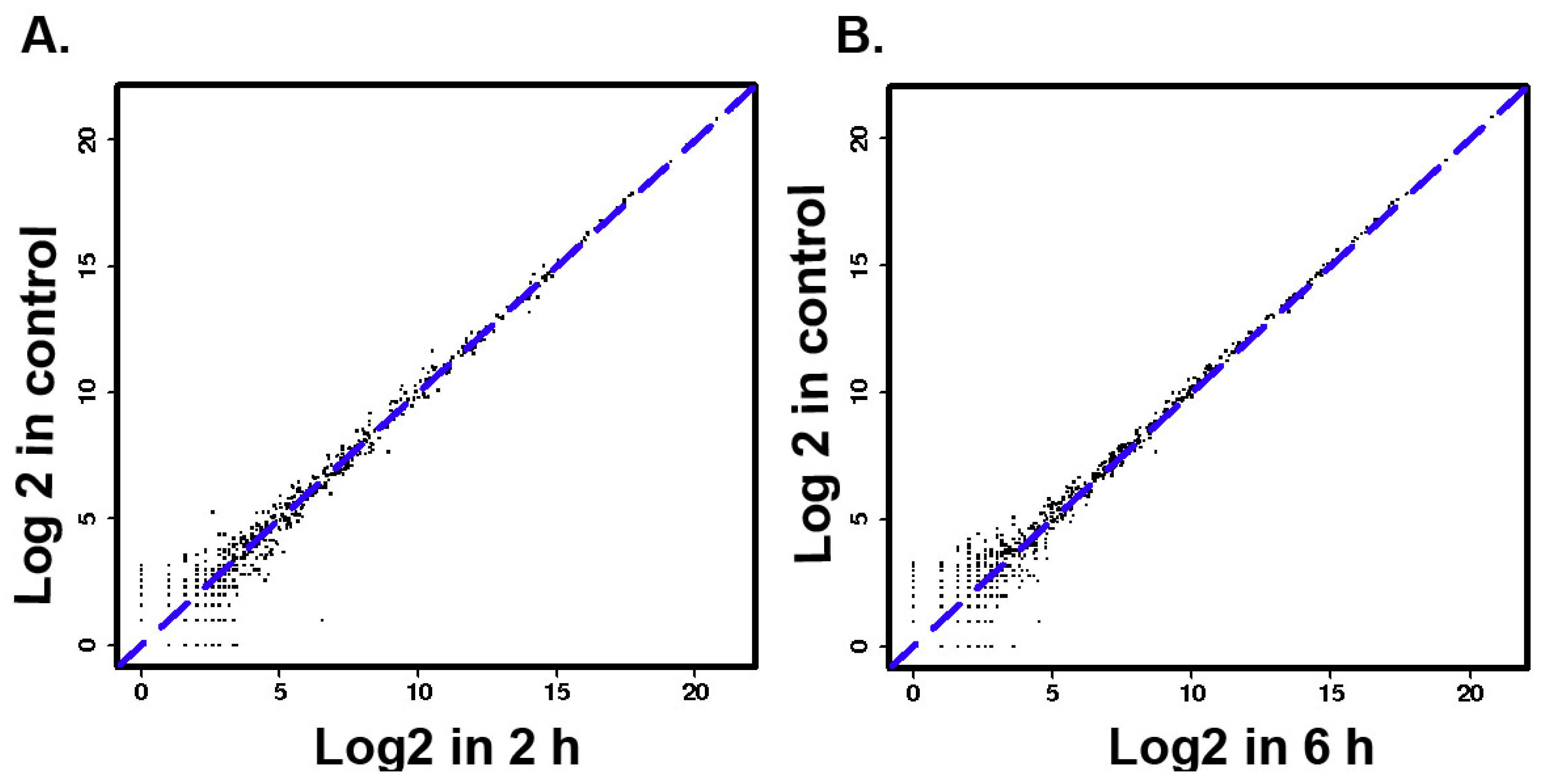
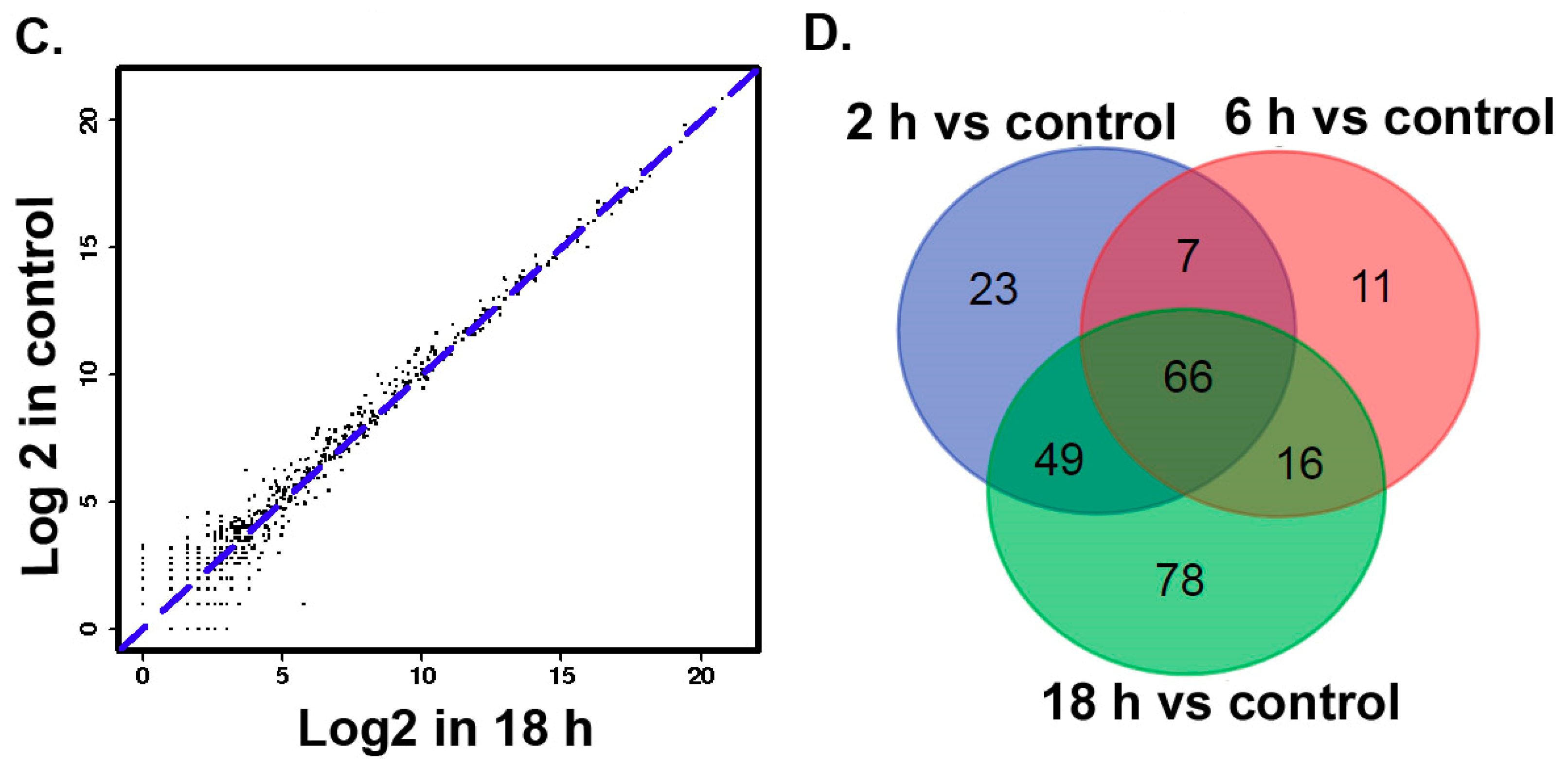
3.2. The Differentially Expressed microRNAs after Treatment with ZEA
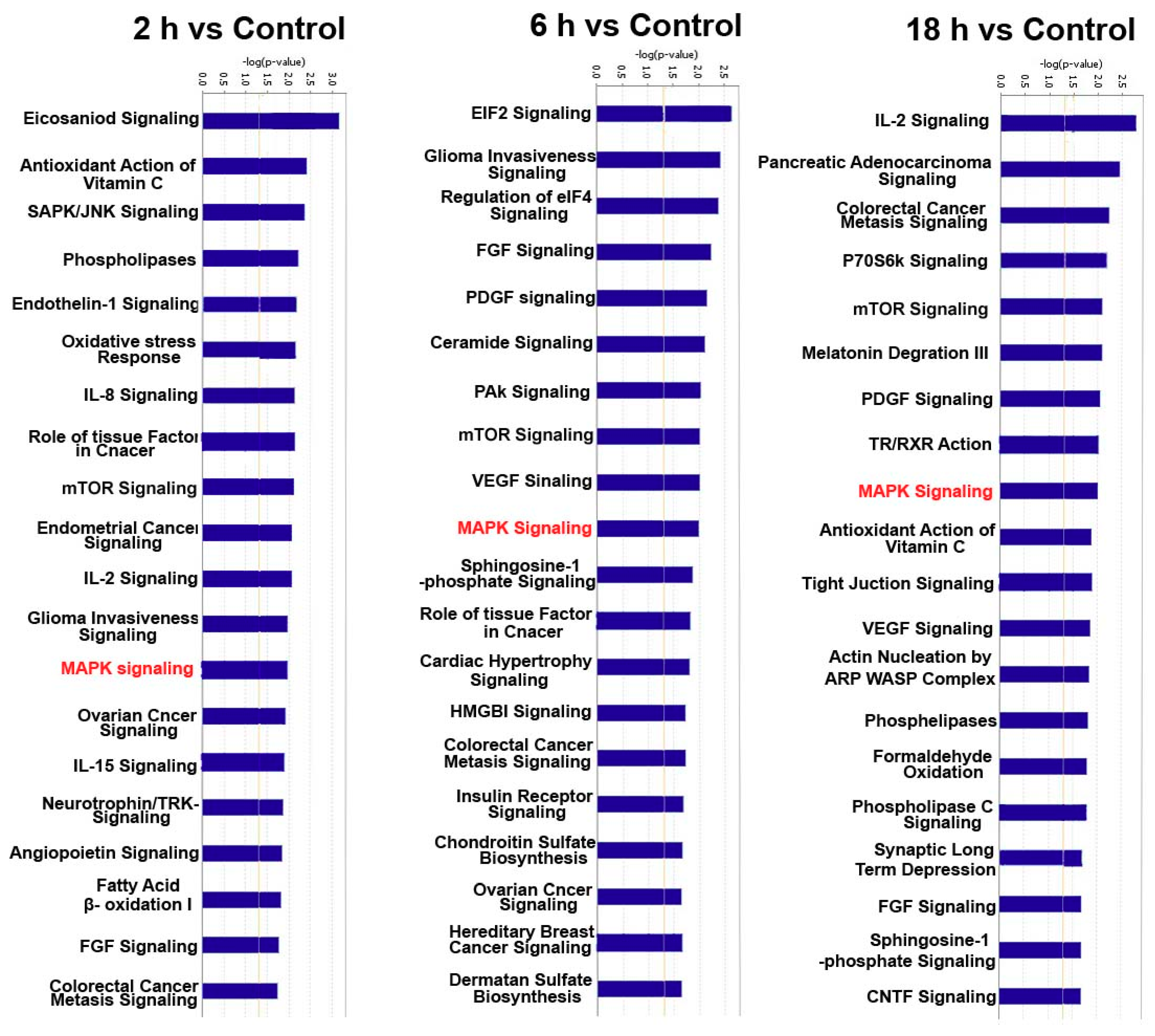
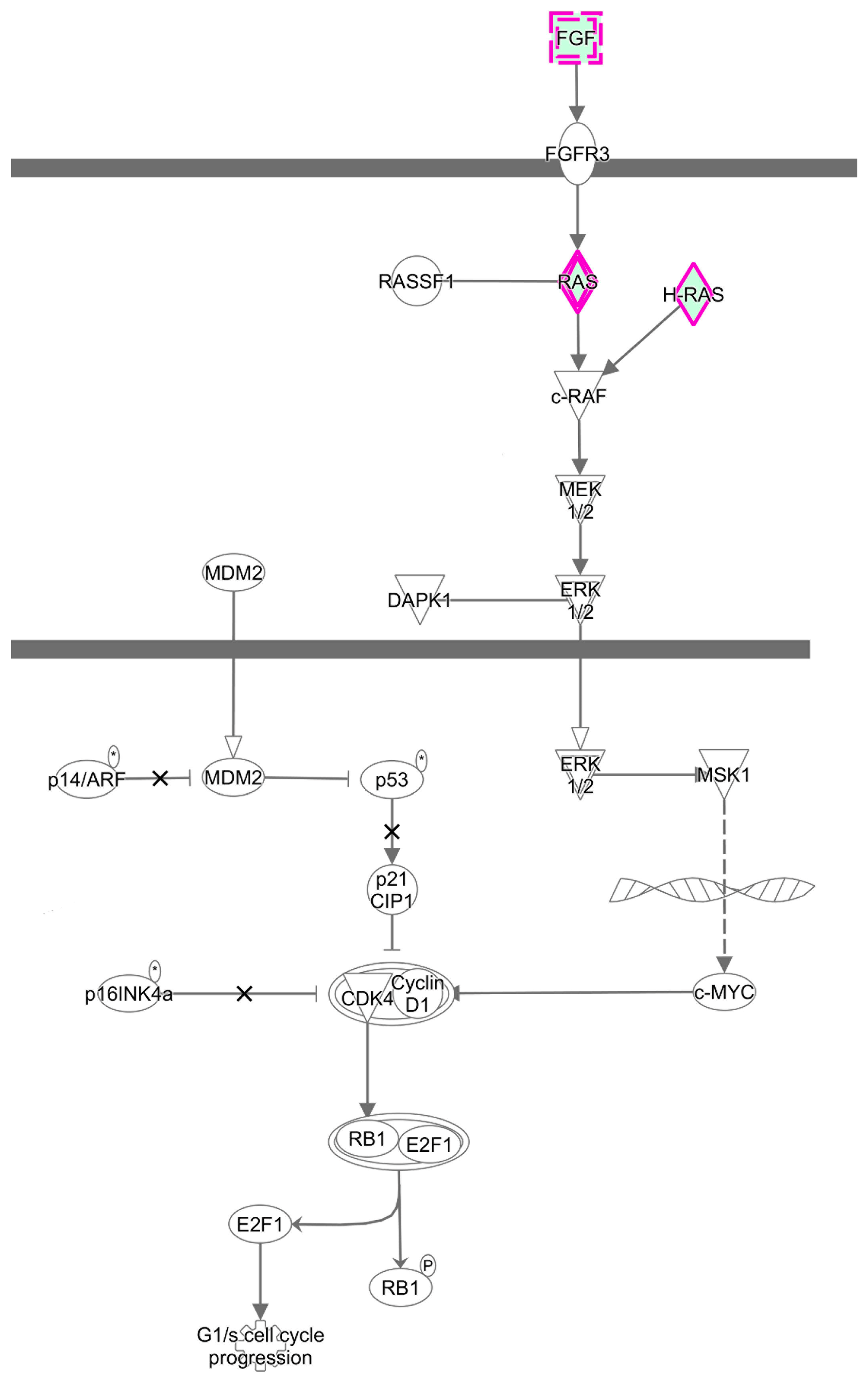
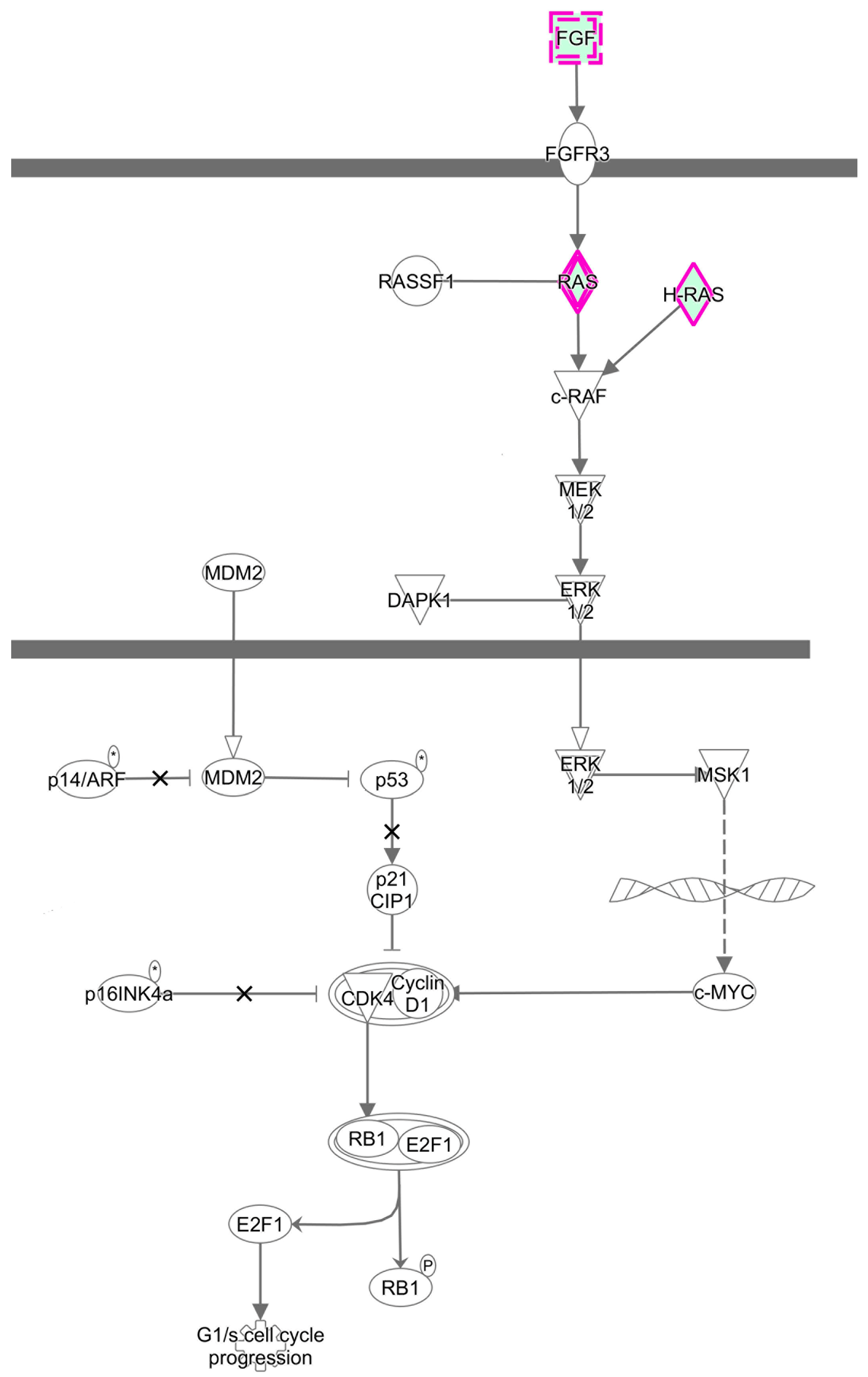
3.3. Predicted the Cell Functions and Pathways of miRNAs Target Genes
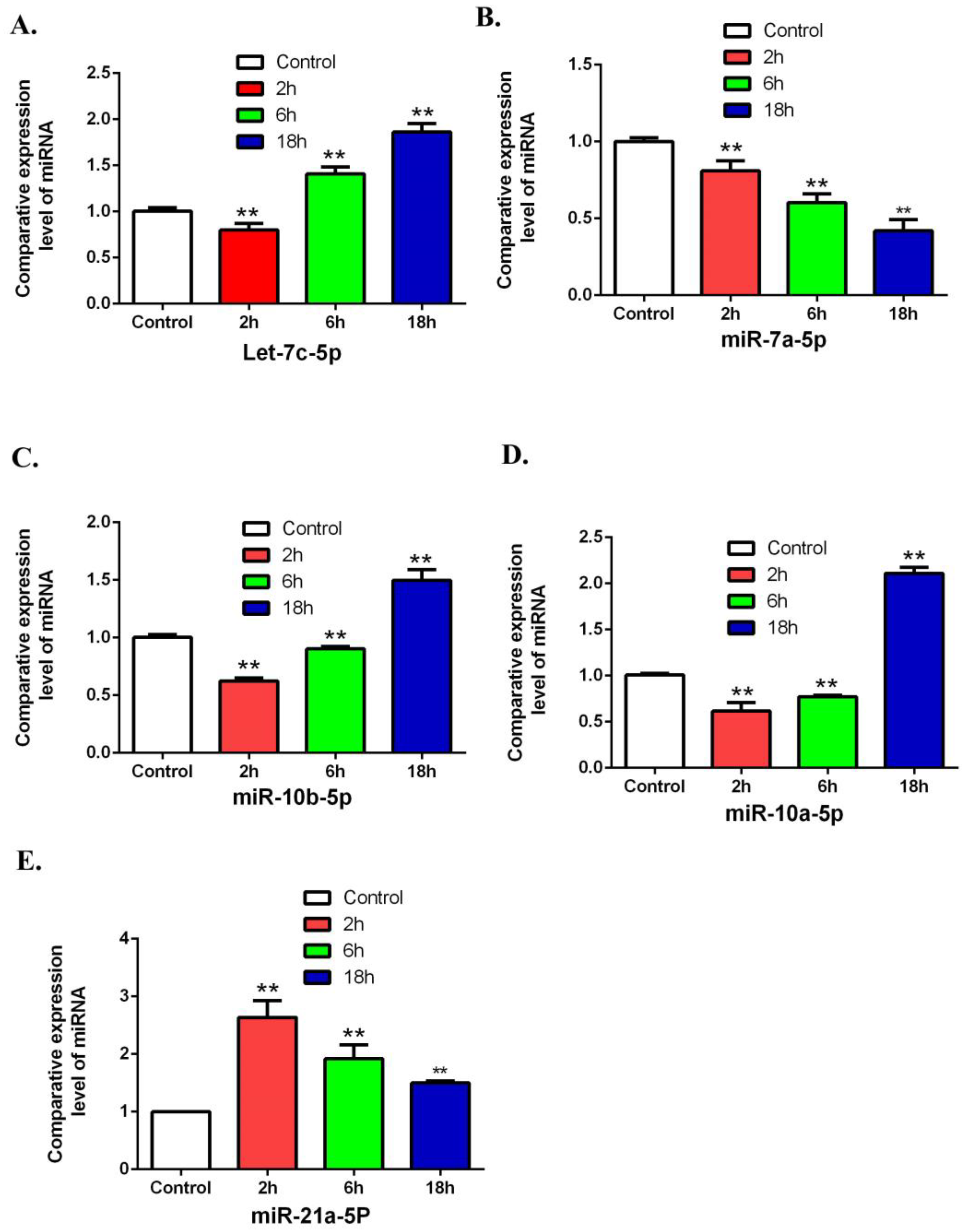
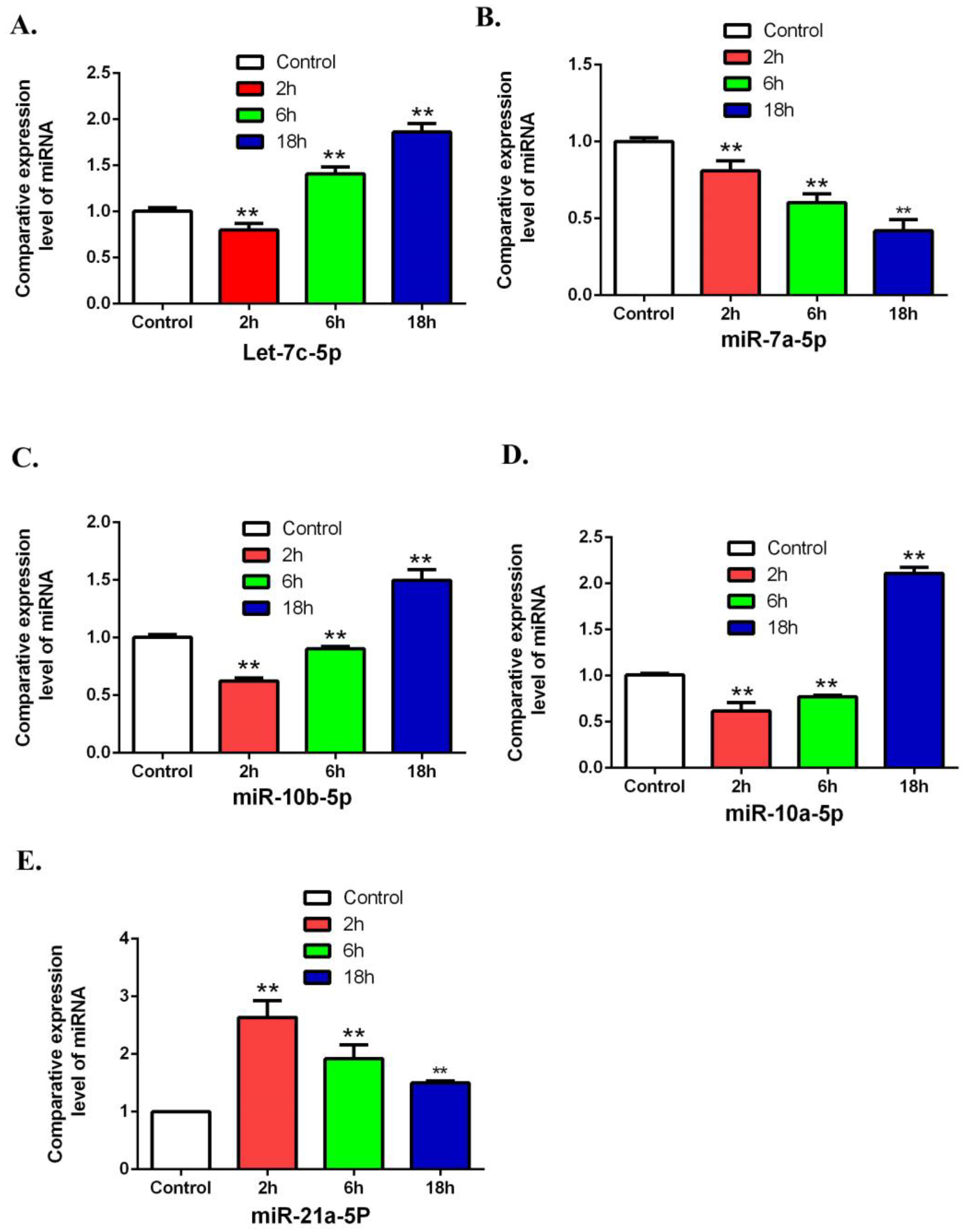
3.4. Validation of Differentially Expressed miRNAs by Using QRT-PCR
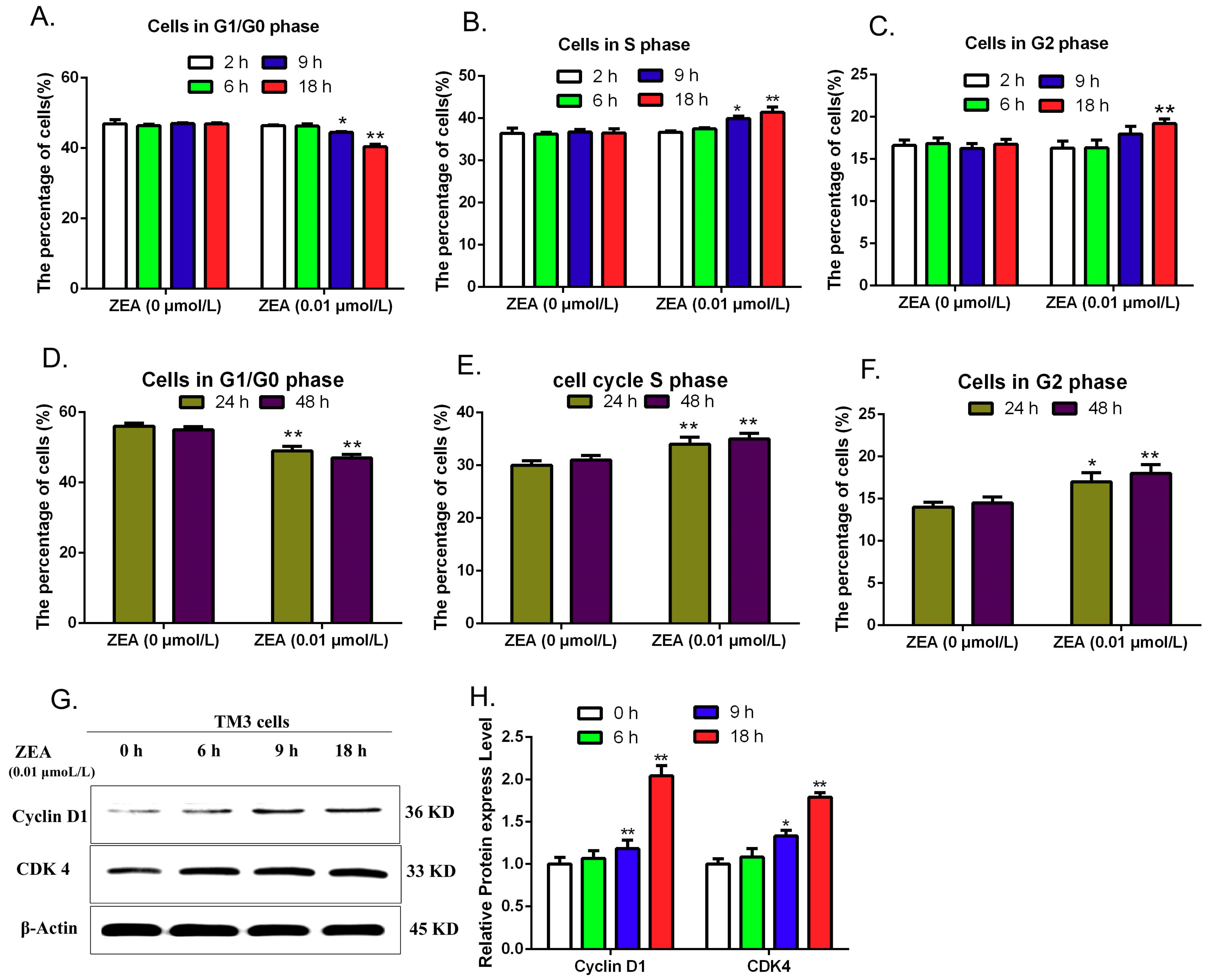
3.5. Validating the Effects of ZEA on Cell Cycle
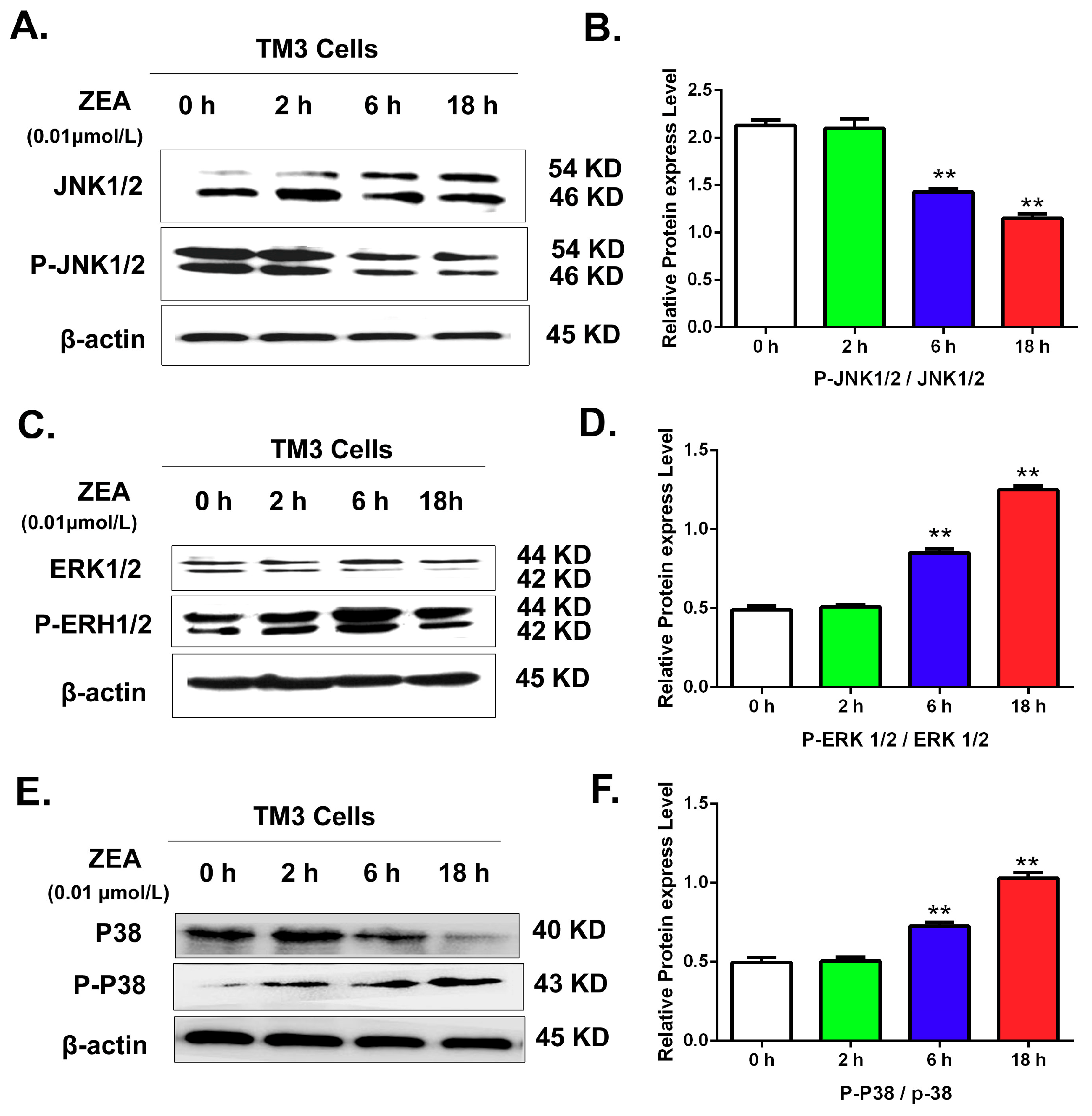
3.6. Validating the Effects of the MAPK Signaling Pathway by Uusing Western Blot
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Full name |
| 3α-HSD | 3α-Hydroxysteroid dehydrogenase |
| 3β -HSD | 3β–Hydroxysteroid dehydrogenase |
| α-ZEA | α-Zearalenol |
| α-ZAL | α-Zearalanol |
| β-ZAL | β-Zearalanol |
| β-ZEA | β-Zearalenol |
| CCK8 | Cell counting kit-8 assay |
| Cdk4 | Cyclin-dependent kinase 4 |
| Cyclin D1 | Cyclin-delta-1 |
| ERK | Extracellular signal-regulated kinase |
| JNK | c-Jun N-terminal kinases |
| MAPK | Mitogen-activated protein kinase |
| MEK | Dual specificity mitogen-activated protein kinase |
| miRNA | MicroRNA |
| RAF | Serine/threonine-protein kinase |
| RTCA | Real-time cell analysis |
| SOAP | Short oLigonucLeotide aLignment program |
| ZAN | Zearalanone |
| ZEA | Zearalenone |
References
- Zheng, W.; Wang, B.; Li, X.; Wang, T.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Bai, J.; Bian, J.; et al. Zearalenone promotes cell proliferation or causes cell death? Toxins (Basel) 2018, 10, 184. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef]
- Skladanka, J.; Adam, V.; Zitka, O.; Mlejnkova, V.; Kalhotka, L.; Horky, P.; Konecna, K.; Hodulikova, L.; Knotova, D.; Balabanova, M.; et al. Comparison of biogenic amines and mycotoxins in alfalfa and red clover fodder depending on additives. Int. J. Environ. Res. Public Health 2017, 14, 418. [Google Scholar] [CrossRef] [PubMed]
- Skladanka, J.; Nedelnik, J.; Adam, V.; Dolezal, P.; Moravcova, H.; Dohnal, V. Forage as a primary source of mycotoxins in animal diets. Int. J. Environ. Res. Public Health 2011, 8, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of zearalenone and its derivatives on the synthesis and secretion of mammalian sex steroid hormones: A review. Food Chem. Toxicol. 2019, 126, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Filannino, A.; Stout, T.A.; Gadella, B.M.; Sostaric, E.; Pizzi, F.; Colenbrander, B.; Dell’Aquila, M.E.; Minervini, F. Dose-response effects of estrogenic mycotoxins (zearalenone, alpha- and beta-zearalenol) on motility, hyperactivation and the acrosome reaction of stallion sperm. Reprod. Biol. Endocrinol. 2011, 9, 134. [Google Scholar] [CrossRef]
- Adibnia, E.; Razi, M.; Malekinejad, H. Zearalenone and 17 beta-estradiol induced damages in male rats reproduction potential; evidence for eralpha and erbeta receptors expression and steroidogenesis. Toxicon 2016, 120, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Mukai, S.; Kuriyagawa, T.; Takagaki, K.; Uno, S.; Kokushi, E.; Otoi, T.; Budiyanto, A.; Shirasuna, K.; Miyamoto, A.; et al. Detection of zearalenone and its metabolites in naturally contaminated follicular fluids by using lc/ms/ms and in vitro effects of zearalenone on oocyte maturation in cattle. Reprod. Toxicol. 2008, 26, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.S.; Takagi, M.; Talukder, A.K.; Marey, M.A.; Kowsar, R.; Abdel-Razek, A.R.K.; Shimizu, T.; Fink-Gremmels, J.; Miyamoto, A. Zearalenone (zen) disrupts the anti-inflammatory response of bovine oviductal epithelial cells to sperm in vitro. Reprod. Toxicol. 2017, 74, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, S.; Lelong, M.; Bourgine, G.; Efstathiou, T.; Saligaut, C.; Pakdel, F. Assessment of the potential activity of major dietary compounds as selective estrogen receptor modulators in two distinct cell models for proliferation and differentiation. Toxicol. Appl. Pharm. 2017, 325, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Khosrokhavar, R.; Rahimifard, N.; Shoeibi, S.; Hamedani, M.P.; Hosseini, M.J. Effects of zearalenone and alpha-zearalenol in comparison with raloxifene on t47d cells. Toxicol. Mech. Methods 2009, 19, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Abassi, H.; Ayed-Boussema, I.; Shirley, S.; Abid, S.; Bacha, H.; Micheau, O. The mycotoxin zearalenone enhances cell proliferation, colony formation and promotes cell migration in the human colon carcinoma cell line hct116. Toxicol. Lett. 2016, 254, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zong, S.; Zeng, G.; Fang, Y.; Peng, J.; Zou, B.; Gao, T.; Zhao, J. The effects of alpha-zearalanol on the proliferation of bone-marrow-derived mesenchymal stem cells and their differentiation into osteoblasts. J. Bone Miner. Metab. 2016, 34, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Cortinovis, C.; Caloni, F.; Schreiber, N.B.; Spicer, L.J. Effects of fumonisin b1 alone and combined with deoxynivalenol or zearalenone on porcine granulosa cell proliferation and steroid production. Theriogenology 2014, 81, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Micrornas: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Wu, Z.; Sun, H.; Zeng, W.; He, J.; Mao, X. Upregulation of mircorna-370 induces proliferation in human prostate cancer cells by downregulating the transcription factor foxo1. PLoS ONE 2012, 7, e45825. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.B.; Xue, L.; Yang, J.; Ma, A.H.; Zhao, J.; Xu, M.; Tepper, C.G.; Evans, C.P.; Kung, H.J.; deVere White, R.W. An androgen-regulated mirna suppresses bak1 expression and induces androgen-independent growth of prostate cancer cells. Proc. Natl. Acad. Sci. USA 2007, 104, 19983–19988. [Google Scholar] [CrossRef]
- Tao, H.; Xiong, Q.; Ji, Z.; Zhang, F.; Liu, Y.; Chen, M. Nfat5 is regulated by p53/mir-27a signal axis and promotes mouse ovarian granulosa cells proliferation. Int. J. Biol. Sci. 2019, 15, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Duan, F.F.; Wang, Y. Micrornas and rna binding protein regulators of micrornas in the control of pluripotency and reprogramming. Curr. Opin. Genet. Dev. 2017, 46, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, X.; Hu, G.; Li, L.; Su, H.; Wang, Y.; Chen, D.; Zhu, Q.; Li, C.; Li, J.; et al. Effects of in utero exposure to dicyclohexyl phthalate on rat fetal leydig cells. Int. J. Environ. Res. Public Health 2016, 13, 246. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Wu, T.; Liu, J.Y.; Gao, P.; Li, K.C.; Guo, Q.Y.; Yuan, M.; Lang, H.Y.; Zeng, L.H.; Guo, G.Z. 1950mhz radio frequency electromagnetic radiation inhibits testosterone secretion of mouse leydig cells. Int. J. Environ. Res. Public Health 2017, 15, 17. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, B.; Huang, K.; He, X.; Luo, Y.; Liang, R.; Luo, H.; Shen, X.L.; Xu, W. Mitochondrial proteomic analysis reveals the molecular mechanisms underlying reproductive toxicity of zearalenone in mltc-1 cells. Toxicology 2014, 324, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Blajet-Kosicka, A.; Kosicki, R.; Twaruzek, M.; Grajewski, J. Determination of moulds and mycotoxins in dry dog and cat food using liquid chromatography with mass spectrometry and fluorescence detection. Food Addit. Contam. B 2014, 7, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Dinleyici, M.; Aydemir, O.; Yildirim, G.K.; Kaya, T.B.; Carman, K.B. Human mature milk zearalenone and deoxynivalenol levels in turkey. Neuro. Endocrinol. Lett. 2018, 39, 325–330. [Google Scholar]
- Chen, F.; Wen, X.; Lin, P.; Chen, H.; Wang, A.; Jin, Y. Herp depletion inhibits zearalenone-induced apoptosis through autophagy activation in mouse ovarian granulosa cells. Toxicol. Lett. 2019, 301, 1–10. [Google Scholar] [CrossRef]
- Li, P.; Yang, S.; Zhang, X.; Huang, S.; Wang, N.; Wang, M.; Long, M.; He, J. Zearalenone changes the diversity and composition of caecum microbiota in weaned rabbit. Biomed. Res. Int. 2018, 2018, 3623274. [Google Scholar] [CrossRef]
- Xiao, Y.X.; Xu, S.W.; Zhao, S.C.; Liu, K.X.; Lu, Z.J.; Hou, Z.Z. Protective effects of selenium against zearalenone-induced apoptosis in chicken spleen lymphocyte via an endoplasmic reticulum stress signaling pathway. Cell Stress Chaperones 2019, 24, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Schumann, B.; Winkler, J.; Mickenautsch, N.; Warnken, T.; Danicke, S. Effects of deoxynivalenol (don), zearalenone (zen), and related metabolites on equine peripheral blood mononuclear cells (pbmc) in vitro and background occurrence of these toxins in horses. Mycotoxin Res. 2016, 32, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, W.; Bian, X.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis and cytoprotective autophagy in primary leydig cells. Toxicol. Lett. 2014, 226, 182–191. [Google Scholar] [CrossRef]
- Zheng, W.; Pan, S.; Wang, G.; Wang, Y.J.; Liu, Q.; Gu, J.; Yuan, Y.; Liu, X.Z.; Liu, Z.P.; Bian, J.C. Zearalenone impairs the male reproductive system functions via inducing structural and functional alterations of sertoli cells. Environ. Toxicol. Pharmacol. 2016, 42, 146–155. [Google Scholar] [CrossRef]
- Yang, J.Y.; Zhang, Y.F.; Wang, Y.Q.; Cui, S. Toxic effects of zearalenone and alpha-zearalenol on the regulation of steroidogenesis and testosterone production in mouse leydig cells. Toxicol. In Vitro 2007, 21, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Wang, Y.; Ma, L.; Chen, X.; Lu, Y.; Ge, F.; Chen, Y.; Chen, X.; Lian, Q.; Jin, X.D.; et al. Zearalenone delays rat leydig cell regeneration. Toxicol. Sci. 2018, 164, 60–71. [Google Scholar] [CrossRef]
- Nasmyth, K. A prize for proliferation. Cell 2001, 107, 689–701. [Google Scholar] [CrossRef]
- Kaufmann, T.; Tai, L.; Ekert, P.G.; Huang, D.C.; Norris, F.; Lindemann, R.K.; Johnstone, R.W.; Dixit, V.M.; Strasser, A. The bh3-only protein bid is dispensable for DNA damage- and replicative stress-induced apoptosis or cell-cycle arrest. Cell 2007, 129, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Huang, Q.; Pan, S.; Fan, W.; Wang, G.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Regulation of oncogenes and gap junction intercellular communication during the proliferative response of zearalenone in tm3 cells. Hum. Exp. Toxicol. 2017, 36, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, B.; He, X.; Cheng, W.H.; Xu, W.; Luo, Y.; Liang, R.; Luo, H.; Huang, K. Analysis of individual and combined effects of ochratoxin A and zearalenone on hepg2 and kk-1 cells with mathematical models. Toxins (Basel) 2014, 6, 1177–1192. [Google Scholar] [CrossRef] [PubMed]
- Yip, K.Y.; Wan, M.L.Y.; Wong, A.S.T.; Korach, K.S.; El-Nezami, H. Combined low-dose zearalenone and aflatoxin b1 on cell growth and cell-cycle progression in breast cancer mcf-7 cells. Toxicol. Lett. 2017, 281, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.H.; Zhao, Z.X.; Dai, J.; Geng, D.C.; Xu, Y.Z. Microrna-221 regulates osteosarcoma cell proliferation, apoptosis, migration, and invasion by targeting cdkn1b/p27. J. Cell. Biochem. 2019, 120, 4665–4674. [Google Scholar] [CrossRef]
- Eilam-Frenkel, B.; Naaman, H.; Brkic, G.; Veksler-Lublinsky, I.; Rall, G.; Shemer-Avni, Y.; Gopas, J. Microrna 146-5p, mir-let-7c-5p, mir-221 and mir-345-5p are differentially expressed in respiratory syncytial virus (rsv) persistently infected hep-2 cells. Virus Res. 2018, 251, 34–39. [Google Scholar] [CrossRef]
- Wang, Y.; Blelloch, R. Cell cycle regulation by micrornas in stem cells. Results Probl. Cell Differ. 2011, 53, 459–472. [Google Scholar]
- Tatay, E.; Espin, S.; Garcia-Fernandez, A.J.; Ruiz, M.J. Estrogenic activity of zearalenone, alpha-zearalenol and beta-zearalenol assessed using the e-screen assay in mcf-7 cells. Toxicol. Mech. Methods 2018, 28, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Li, C.; Xu, Y.; Chen, S.; Zhao, Y.; Chen, L.; Jiang, Y.; Liu, Z.; Fan, R.; Sun, L.; et al. Differential expression of micrornas in tm3 leydig cells of mice treated with brain-derived neurotrophic factor. Cell Bioch. Funct. 2017, 35, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, J.; Ochiya, T. Extracellular micrornas and oxidative stress in liver injury: A systematic mini review. J. Clin. Biochem. Nutr. 2018, 63, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewski, J.; Miturski, R.; Semczuk, A.; Kotarski, J.; Jakowicki, J. Tissue zearalenone concentration in normal, hyperplastic and neoplastic human endometrium. Ginekologia Polska 1998, 69, 363–366. [Google Scholar] [PubMed]
- Yu, Z.; Zhang, L.; Wu, D.; Liu, F. Anti-apoptotic action of zearalenone in mcf-7 cells. Ecotoxicol. Environ. Saf. 2005, 62, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Belli, P.; Bellaton, C.; Durand, J.; Balleydier, S.; Milhau, N.; Mure, M.; Mornex, J.F.; Benahmed, M.; Le Jan, C. Fetal and neonatal exposure to the mycotoxin zearalenone induces phenotypic alterations in adult rat mammary gland. Food Chem. Toxicol. 2010, 48, 2818–2826. [Google Scholar] [CrossRef] [PubMed]
- Becci, P.J.; Johnson, W.D.; Hess, F.G.; Gallo, M.A.; Parent, R.A.; Taylor, J.M. Combined two-generation reproduction-teratogenesis study of zearalenone in the rat. J. Appl. Toxicol. 1982, 2, 201–206. [Google Scholar] [CrossRef]
- Zheng, S.; Shen, H.; Jia, Q.; Jing, C.; Lin, J.; Zhang, M.; Zhang, X.; Zhang, B.; Liu, Y. S100a6 promotes proliferation of intrahepatic cholangiocarcinoma cells via the activation of the p38/mapk pathway. Future Oncol. 2017, 13, 2053–2063. [Google Scholar] [CrossRef]
- Gerthoffer, W.T.; Singer, C.A. Mapk regulation of gene expression in airway smooth muscle. Respir. Physiol. Neurobiol. 2003, 137, 237–250. [Google Scholar] [CrossRef]
- Martin, E.D.; Bassi, R.; Marber, M.S. P38 mapk in cardioprotection—Are we there yet? Br. J. Pharmacol. 2015, 172, 2101–2113. [Google Scholar] [CrossRef]
- Gorog, D.A.; Jabr, R.I.; Tanno, M.; Sarafraz, N.; Clark, J.E.; Fisher, S.G.; Cao, X.B.; Bellahcene, M.; Dighe, K.; Kabir, A.M.; et al. Mapkapk-2 modulates p38-mapk localization and small heat shock protein phosphorylation but does not mediate the injury associated with p38-mapk activation during myocardial ischemia. Cell Stress Chaperones 2009, 14, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Guldal, C.G.; Ahmad, A.; Korshunov, A.; Squatrito, M.; Awan, A.; Mainwaring, L.A.; Bhatia, B.; Parathath, S.R.; Nahle, Z.; Pfister, S.; et al. An essential role for p38 mapk in cerebellar granule neuron precursor proliferation. Acta Neuropathol. 2012, 123, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Rousseau, S. P38 map-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [PubMed]
- Zarubin, T.; Han, J. Activation and signaling of the p38 map kinase pathway. Cell Res. 2005, 15, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Edington, H.D.; Jukic, D.M.; Rao, U.N.; Land, S.R.; Kirkwood, J.M. Impact of ifnalpha2b upon pstat3 and the mek/erk mapk pathway in melanoma. Cancer Immunol. Immunother. 2008, 57, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Liu, G.W.; Ding, L.; Jiang, T.; Shao, S.H.; Gao, Y.; Lu, Y. Hoxa3 promotes tumor growth of human colon cancer through activating egfr/ras/raf/mek/erk signaling pathway. J. Cell. Biochem. 2018, 119, 2864–2874. [Google Scholar] [CrossRef] [PubMed]
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Wong, E.W.; Chang, F.; Lehmann, B.; Terrian, D.M.; Milella, M.; Tafuri, A.; et al. Roles of the raf/mek/erk pathway in cell growth, malignant transformation and drug resistance. Biochim. Ebiophys. Acta 2007, 1773, 1263–1284. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer | Primer Sequence |
|---|---|---|
| miR-21a-5p | RT primer | UAGCUUAUCAGACUGAUGUUGA |
| Forward primer | GCAGTAGCTTATCAGACTGATA | |
| Reverse primer | GGTCCAGTTTTTTTTTTTTTTTCAAC | |
| miR-10a-5p | RT primer | UACCCUGUAGAUCCGAAUUUGUG |
| Forward primer | GCAGTACCCTGTAGATCCGA | |
| Reverse primer | GGTCCAGTTTTTTTTTTTTTTTCAC | |
| miR-10b-5p | RT primer | UACCCUGUAGAACCGAAUUUGUG |
| Forward primer | CAGTACCCTGTAGAACCGA | |
| Reverse primer | GGTCCAGTTTTTTTTTTTTTTTCAC | |
| miR-7a-5p | RT primer | UGGAAGACUAGUGAUUUUGUUGU |
| Forward primer | CGCAGTGGAAGACTAGTGA | |
| Reverse primer | GTCCAGTTTTTTTTTTTTTTTACAACA | |
| let-7c-5p | RT primer | UGAGGUAGUAGGUUGUAUGGUU |
| Forward primer | GCAGTGAGGTAGTAGGTTGT | |
| Reverse primer | GGTCCAGTTTTTTTTTTTTTTTAACCA | |
| U6 | Forward primer | GGAACGATACAGAGAAGATTAGC |
| Reverse primer | TGGAACGCTTCACGAATTTGCG |
| miRNA Title | 2 h VS. Control | 6 h VS. Control | 18 h VS. Control | |||
|---|---|---|---|---|---|---|
| Fold Changes log2 | p-Value | Fold Changes log2 | p-Value | Fold Changes log2 | p-Value | |
| mmu-let-7a-5p | −0.12 | 1.04 × 10−184 | −0.04 | 2.06 × 10−21 | 0.14 | 1.94 × 10−251 |
| mmu-let-7b-3p | 0.45 | 0.00 | 0.42 | 9.89 × 10−4 | −0.91 | 1.07 × 10−8 |
| mmu-let-7b-5p | −0.10 | 0.00 | −0.03 | 2.13 × 10−58 | −0.39 | 0.00 |
| mmu-let-7c-5p | −0.01 | 0.00 | 0.06 | 9.80 × 10−134 | 0.14 | 0.00 |
| mmu-let-7e-5p | −0.07 | 1.01 ×10−29 | −0.03 | 6.96 × 10−7 | −0.14 | 1.55 × 10−109 |
| mmu-let-7f-5p | −0.17 | 0.00 | −0.12 | 5.31 × 10−152 | 0.28 | 0.00 |
| mmu-let-7i-3p | 5.56 | 9.77 × 10−24 | 3.63 | 3.48 × 10−6 | 4.80 | 3.40 × 10−14 |
| mmu-let-7i-5p | −0.05 | 0.00 | 0.02 | 1.73 × 10−57 | −0.07 | 0.00 |
| mmu-miR-100-5p | −0.13 | 1.19 × 10−132 | −0.14 | 1.82 × 10−166 | 0.40 | 0.00 |
| mmu-miR-10a-5p | −0.17 | 0.00 | −0.02 | 5.98 × 10−9 | 0.38 | 0.00 |
| mmu-miR-10b-5p | −0.21 | 6.45 × 10−254 | −0.07 | 1.36 × 10−26 | 0.31 | 0.00 |
| mmu-miR-1198-5p | −0.19 | 0.00 | −0.08 | 2.27 × 10−4 | −0.27 | 1.12 × 10−30 |
| mmu-miR-125a-5p | −0.03 | 3.88 × 10−4 | −0.07 | 6.84 × 10−19 | 0.03 | 2.28 × 10−5 |
| mmu-miR-125b-5p | 0.06 | 0.00 | −0.05 | 5.03 × 10−5 | 0.19 | 9.56 × 10−73 |
| mmu-miR-128-3p | 0.06 | 2.44 × 10−3 | 0.13 | 3.37 × 10−10 | −0.25 | 7.47 × 10−31 |
| mmu-miR-139-3p | 3.36 | 0.00 | 3.70 | 7.45 × 10−4 | 3.04 | 1.21 × 10−2 |
| mmu-miR-140-3p | 0.30 | 5.09 × 10−24 | 0.13 | 3.49 × 10−5 | 0.36 | 9.26 × 10−36 |
| mmu-miR-143-3p | 0.10 | 0.00 | 0.07 | 7.51 × 10−12 | 0.36 | 5.60 × 10−299 |
| mmu-miR-148a-3p | −0.06 | 2.76 × 10−33 | 0.15 | 8.57 × 10−191 | 0.18 | 4.63 × 10−275 |
| mmu-miR-149-3p | 1.28 | 0.00 | 1.14 | 1.15 × 10−21 | −1.40 | 2.31 × 10−14 |
| mmu-miR-151-3p | −0.07 | 3.47 × 10−17 | 0.08 | 1.75 × 10−19 | −0.17 | 1.94 × 10−85 |
| mmu-miR-16-1-3p | −0.20 | 0.00 | −0.52 | 1.05 × 10−18 | −0.94 | 1.75 × 10−50 |
| mmu-miR-182-3p | −0.59 | 6.70 × 10−5 | −0.39 | 6.57 × 10−3 | −1.57 | 3.28 × 10−19 |
| mmu-miR-182-5p | −0.11 | 0.00 | −0.06 | 7.50 × 10−14 | 0.12 | 6.94 × 10−67 |
| mmu-miR-183-5p | −0.05 | 5.08 × 10−20 | 0.02 | 2.03 × 10−4 | 0.10 | 4.05 × 10−74 |
| mmu-miR-196b-5p | −0.15 | 0.00 | −0.07 | 5.29 × 10−4 | 0.09 | 1.02 × 10−5 |
| mmu-miR-199a-3p | 0.29 | 1.22 × 10−69 | 0.07 | 3.34 × 10−5 | 0.31 | 5.12 × 10−80 |
| mmu-miR-199a-5p | −0.48 | 0.00 | −0.22 | 4.46 × 10−75 | 0.93 | 0.00 |
| mmu-miR-199b-3p | 0.29 | 1.92 × 10−69 | 0.07 | 5.92 × 10−5 | 0.31 | 1.40 × 10−79 |
| mmu-miR-206-3p | −0.22 | 0.00 | −0.20 | 6.43 × 10−9 | −0.83 | 8.43 × 10−110 |
| mmu-miR-21a-5p | 0.15 | 0.00 | 0.04 | 5.92 × 10−210 | 0.03 | 2.08 × 10−114 |
| mmu-miR-221-5p | −0.75 | 0.00 | −0.32 | 5.85 × 10−11 | −1.10 | 2.09 × 10−85 |
| mmu-miR-222-3p | −0.02 | 4.47 × 10−4 | −0.10 | 5.31 × 10−63 | −0.46 | 0.00 |
| mmu-miR-23a-3p | 0.55 | 0.00 | 0.26 | 5.69 × 10−5 | 0.38 | 1.56 × 10−9 |
| mmu-miR-24-3p | 0.17 | 4.78 × 10−33 | 0.08 | 3.27 × 10−7 | −0.05 | 4.25 × 10−4 |
| mmu-miR-25-5p | −0.12 | 0.01 | −0.38 | 6.66 × 10−16 | −0.69 | 1.64 × 10−44 |
| mmu-miR-27a-3p | 0.83 | 0.00 | 0.40 | 4.16 × 10−88 | 0.28 | 6.97 × 10−44 |
| mmu-miR-27a-5p | 0.13 | 0.00 | −0.13 | 5.94 × 10−8 | −0.80 | 2.57 × 10−196 |
| mmu-miR-27b-3p | 0.57 | 5.61 × 10−287 | 0.27 | 7.54 × 10−59 | 0.31 | 2.35 × 10−80 |
| mmu-miR-28a-3p | −0.13 | 0.00 | −0.13 | 1.31 × 10−5 | 0.14 | 7.11 × 10−7 |
| mmu-miR-296-3p | 0.21 | 8.10 × 10−38 | −0.05 | 2.18 × 10−3 | −0.34 | 1.19 × 10−82 |
| mmu-miR-30c-2-3p | −0.35 | 0.00 | −0.35 | 2.65 × 10−13 | −0.60 | 6.46 × 10−33 |
| mmu-miR-30d-5p | −0.09 | 4.16 × 10−36 | −0.12 | 3.95 × 10−67 | 0.10 | 6.42 × 10−51 |
| mmu-miR-30e-3p | −0.16 | 0.00 | −0.13 | 1.28 × 10−5 | −0.18 | 4.95 × 10−10 |
| mmu-miR-342-5p | −1.12 | 1.39 × 10−143 | −0.32 | 7.78 × 10−17 | −1.09 | 4.07 × 10−138 |
| mmu-miR-351-3p | −0.41 | 0.00 | −0.44 | 2.25 × 10−8 | −0.77 | 2.97 × 10−20 |
| mmu-miR-365-1-5p | −0.57 | 7.98 × 10−13 | −0.40 | 3.68 × 10−7 | −1.04 | 2.44 × 10−33 |
| mmu-miR-365-2-5p | −0.51 | 0.00 | −0.24 | 2.82 × 10−7 | −1.13 | 3.12 × 10−95 |
| mmu-miR-374b-5p | 0.36 | 5.55 × 10−9 | 0.20 | 1.95 × 10−3 | 0.27 | 1.76 × 10−5 |
| mmu-miR-378a-3p | 0.17 | 0.00 | −0.07 | 1.08 × 10−6 | 0.11 | 2.11 × 10−17 |
| mmu-miR-423-5p | −0.10 | 3.14 × 10−38 | −0.16 | 5.19 × 10−92 | −0.44 | 0.00 |
| mmu-miR-486a-3p | −0.29 | 0.00 | −0.18 | 3.14 × 10−4 | −0.55 | 6.25 × 10−26 |
| mmu-miR-501-3p | −0.43 | 3.95 × 10−45 | −0.20 | 2.82 × 10−11 | 0.46 | 4.11 × 10−71 |
| mmu-miR-503-5p | 0.45 | 0.00 | 0.21 | 1.04 × 10−8 | 0.27 | 3.56 × 10−14 |
| mmu-miR-532-5p | −0.16 | 7.28 × 10−52 | −0.05 | 3.51 × 10−7 | 0.39 | 0.00 |
| mmu-miR-574-5p | −0.47 | 0.00 | −0.07 | 3.05 × 10−7 | −0.59 | 0.00 |
| mmu-miR-615-3p | 0.26 | 9.94 × 10−97 | 0.16 | 2.72 × 10−34 | 0.11 | 2.35 × 10−17 |
| mmu-miR-615-5p | −0.73 | 0.00 | −0.30 | 3.96 × 10−6 | −0.75 | 1.29 × 10−26 |
| mmu-miR-669c-5p | −0.28 | 9.79 × 10−78 | −0.05 | 1.32 × 10−3 | −0.22 | 4.30 × 10−50 |
| mmu-miR-671-3p | −0.42 | 0.00 | −0.23 | 1.00 × 10−5 | −1.00 | 1.11 × 10−66 |
| mmu-miR-7a-5p | −0.03 | 1.80 × 10−8 | −0.19 | 0.00 | −0.47 | 0.00 |
| mmu-miR-877-5p | −0.44 | 0.00 | −0.34 | 8.23 × 10−7 | −1.55 | 5.50 × 10−76 |
| mmu-miR-92b-5p | −0.92 | 2.48 × 10−20 | −0.40 | 1.58 × 10−5 | −1.19 | 4.37 × 10−30 |
| mmu-miR-93-5p | −0.12 | 0.00 | −0.25 | 4.43 × 10−9 | 0.09 | 1.52 × 10−2 |
| mmu-miR-98-5p | −0.08 | 3.33 × 10−6 | −0.07 | 4.45 × 10−5 | −0.16 | 5.41 × 10−19 |
| mmu-miR-99b-3p | −0.10 | 0.00 | −0.11 | 1.33 × 10−9 | −0.70 | 1.32 × 10−274 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, W.; Fan, W.; Feng, N.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Bai, J.; Bian, J.; et al. The Role of miRNAs in Zearalenone-Promotion of TM3 Cell Proliferation. Int. J. Environ. Res. Public Health 2019, 16, 1517. https://doi.org/10.3390/ijerph16091517
Zheng W, Fan W, Feng N, Lu N, Zou H, Gu J, Yuan Y, Liu X, Bai J, Bian J, et al. The Role of miRNAs in Zearalenone-Promotion of TM3 Cell Proliferation. International Journal of Environmental Research and Public Health. 2019; 16(9):1517. https://doi.org/10.3390/ijerph16091517
Chicago/Turabian StyleZheng, Wanglong, Wentong Fan, Nannan Feng, Nanyan Lu, Hui Zou, Jianhong Gu, Yan Yuan, Xuezhong Liu, Jianfa Bai, Jianchun Bian, and et al. 2019. "The Role of miRNAs in Zearalenone-Promotion of TM3 Cell Proliferation" International Journal of Environmental Research and Public Health 16, no. 9: 1517. https://doi.org/10.3390/ijerph16091517
APA StyleZheng, W., Fan, W., Feng, N., Lu, N., Zou, H., Gu, J., Yuan, Y., Liu, X., Bai, J., Bian, J., & Liu, Z. (2019). The Role of miRNAs in Zearalenone-Promotion of TM3 Cell Proliferation. International Journal of Environmental Research and Public Health, 16(9), 1517. https://doi.org/10.3390/ijerph16091517