Integrated Effects of Co-Inoculation with Phosphate-Solubilizing Bacteria and N2-Fixing Bacteria on Microbial Population and Soil Amendment Under C Deficiency


Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Properties and Pretreatment
2.2. Microorganisms
2.3. Experimental Design
2.4. Sampling and Analytical Methods
2.5. Statistical Analysis
3. Results
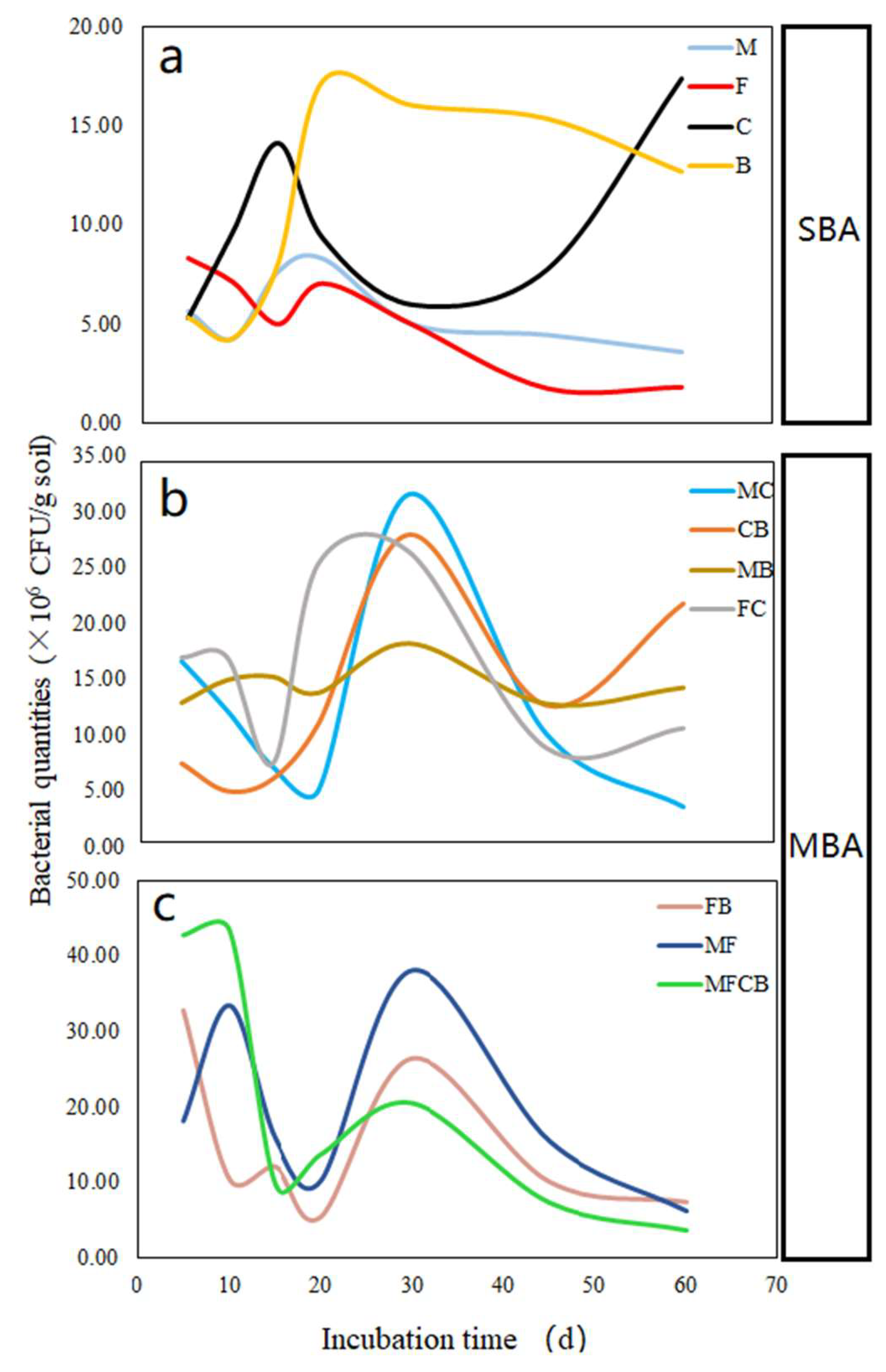
3.1. Dynamic Growth of Bacteria in Incubation Soil
3.2. Inoculants, Incubation Time, and Their Interactions on Soil Characteristics
3.3. C and N Contents of the Incubation Soil
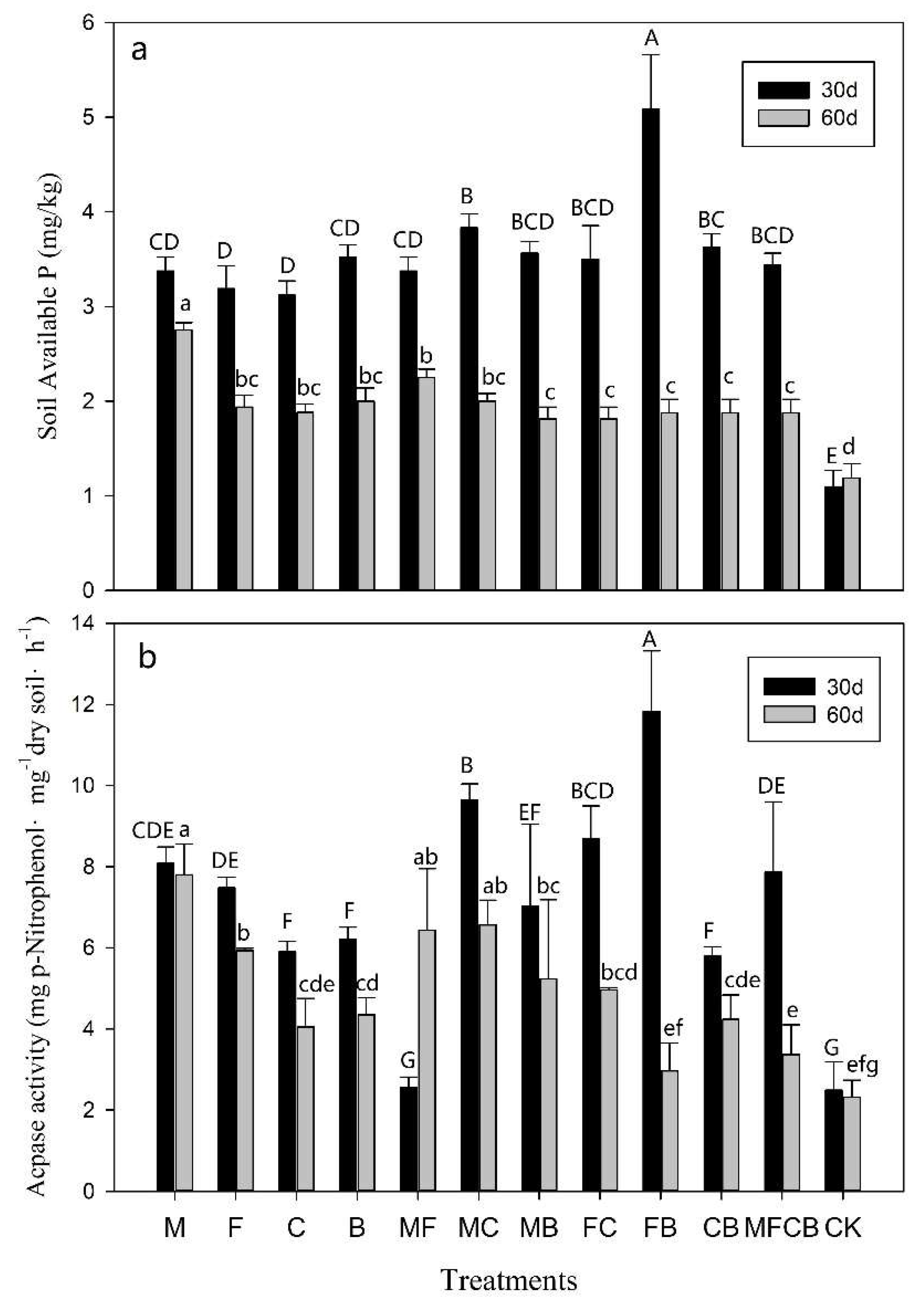
3.4. AcPase Activity and SAP Concentrations
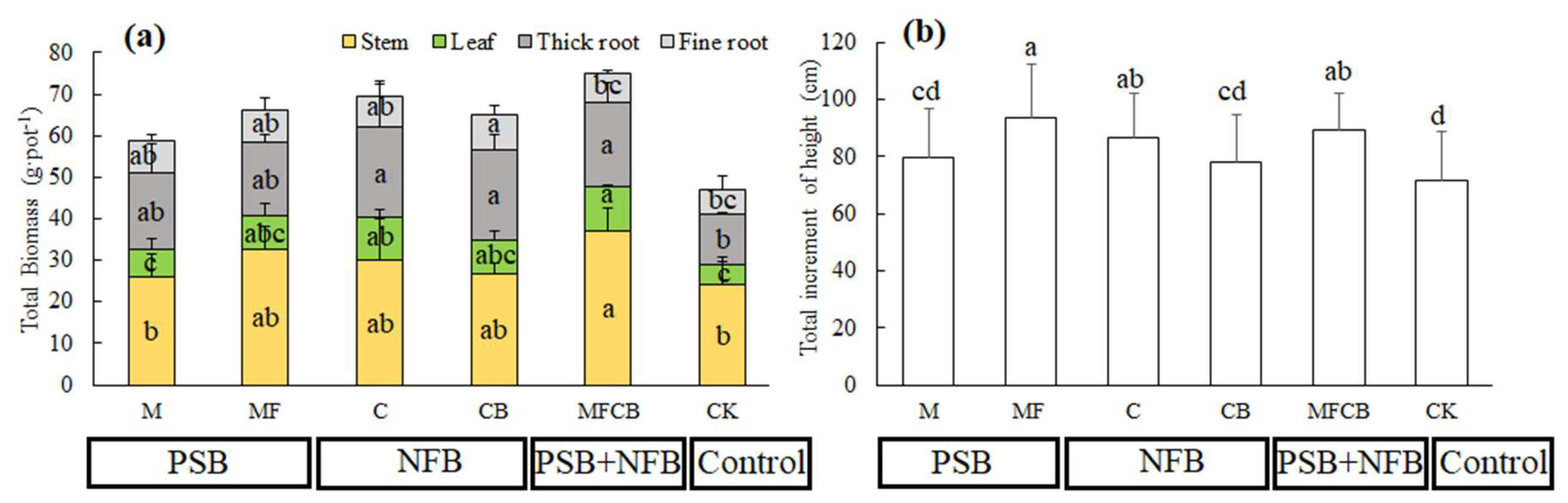
3.5. Seedling Height and Biomass Accumulation
4. Discussion
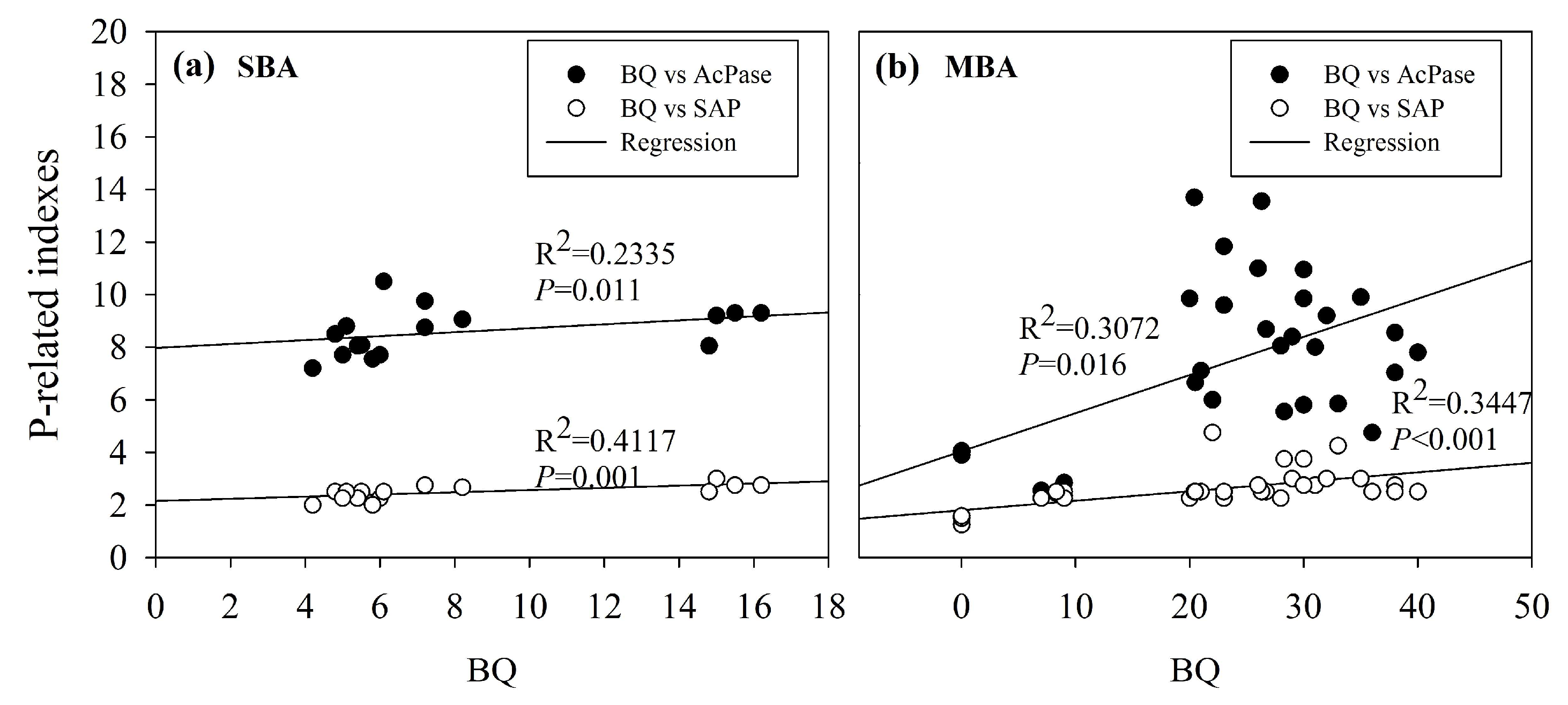
4.1. Changes of BQ Influenced by C Resources and Interactions of Bacteria
4.2. Additions of Bacteria Improved Soil Nutrients with Different Patterns
4.3. Co-Inoculation with NFB and PSB Resulted in Higher Plant Biomass Accumulation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fazhu, Z.; Jiao, S.; Chengjie, R.; Di, K.; Jian, D.; Xinhui, H.; Gaihe, Y.; Yongzhong, F.; Guangxin, R. Land use change influences soil C, N, and P stoichiometry under ‘Grain-to-Green Program’ in China. Sci. Rep. 2015, 5, 10195. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.Q.; Chang, S.; Clinton, P.W.; Johnson, C.E.; Wang, H.L.; Xu, Z.H. Preface to the special issue for the 8th International Symposium on Forest Soils: Linking Soil Processes to Forest Productivity and Water Protection under Global Change. J. Soil Sediments 2017, 17, 2215–2217. [Google Scholar] [CrossRef][Green Version]
- Tang, Y.C.; Zhang, X.Y.; Li, D.D.; Wang, H.M.; Chen, F.S.; Fu, X.L.; Fang, X.M.; Sun, X.M.; Yu, G.R. Impacts of nitrogen and phosphorus additions on the abundance and community structure of ammonia oxidizers and denitrifying bacteria in Chinese fir plantations. Soil Biol. Biochem. 2016, 103, 284–293. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Smil, V. Phosphorus in the Environment: Natural Flows and Human Interferences. Annu. Rev. Energy Environ. 2000, 25, 53–88. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Kloepper, J.W. Plant-microbes interactions in enhanced fertilizer-use efficiency. Appl. Microbiol. Biotechnol. 2009, 85, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gyaneshwar, P.; Kumar, G.N.; Parekh, L.J.; Poole, P.S. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 2002, 245, 83–93. [Google Scholar] [CrossRef]
- Kaur, G.; Reddy, M.S. Effects of Phosphate-Solubilizing Bacteria, Rock Phosphate and Chemical Fertilizers on Maize-Wheat Cropping Cycle and Economics. Pedosphere 2015, 25, 428–437. [Google Scholar] [CrossRef]
- Megali, L.; Glauser, G.; Rasmann, S. Fertilization with beneficial microorganisms decreases tomato defenses against insect pests. Agron. Sustain. Dev. 2014, 34, 649–656. [Google Scholar] [CrossRef]
- Yin, H.J.; Zhao, W.Q.; Li, T.; Cheng, X.Y.; Liu, Q. Balancing straw returning and chemical fertilizers in China: Role of straw nutrient resources. Renew. Sustain. Energy Rev. 2018, 81, 2695–2702. [Google Scholar] [CrossRef]
- Liu, D.; Fang, S.; Tian, Y.; Dun, X. Seasonal and clonal variations of microbial biomass and processes in the rhizosphere of poplar plantations. Appl. Soil Ecol. 2014, 78, 65–72. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Yevdokimov, I.; Gattinger, A.; Buegger, F.; Munch, J.C.; Schloter, M. Changes in microbial community structure in soil as a result of different amounts of nitrogen fertilization. Biol. Fert. Soils 2008, 44, 1103–1106. [Google Scholar] [CrossRef]
- Behera, B.C.; Yadav, H.; Singh, S.K.; Mishra, R.R.; Sethi, B.K.; Dutta, S.K.; Thatoi, H.N. Phosphate solubilization and acid phosphatase activity of Serratia sp. isolated from mangrove soil of Mahanadi river delta, Odisha, India. J. Genet. Eng. Biotechnol. 2017, 15, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Debska, B.; Dlugosz, J.; Piotrowska-Dlugosz, A.; Banach-Szott, M. The impact of a bio-fertilizer on the soil organic matter status and carbon sequestration-results from a field-scale study. J. Soil Sediments 2016, 16, 2335–2343. [Google Scholar] [CrossRef]
- Yilmaz, E.; Sonmez, M. The role of organic/bio-fertilizer amendment on aggregate stability and organic carbon content in different aggregate scales. Soil Tillage Res. 2017, 168, 118–124. [Google Scholar] [CrossRef]
- Huang, Y.M.; Wu, Q.S.; Li, Y. Arbuscular Mycorrhizal Fungi Alter Plant Growth, Soil Aggregate Stability, and Rhizospheric Organic Carbon Pools of Citrus. Adv. Mater. Res. 2013, 610–613, 3063–3066. [Google Scholar] [CrossRef]
- Gunasekara, A.S.; Xing, B.S. Sorption and desorption of naphthalene by soil organic matter: Importance of aromatic and aliphatic components. J. Environ. Qual. 2003, 32, 240–246. [Google Scholar] [CrossRef]
- Yu, X.; Liu, X.; Zhu, T.H.; Liu, G.H.; Mao, C. Co-inoculation with phosphate-solubilzing and nitrogen-fixing bacteria on solubilization of rock phosphate and their effect on growth promotion and nutrient uptake by walnut. Eur. J. Soil Biol. 2012, 50, 112–117. [Google Scholar] [CrossRef]
- Hameeda, B.; Harini, G.; Rupela, O.P.; Wani, S.P.; Reddy, G. Growth promotion of maize by phosphate-solubilizing bacteria isolated from composts and macrofauna. Microbiol. Res. 2008, 163, 234–242. [Google Scholar] [CrossRef]
- Valetti, L.; Iriarte, L.; Fabra, A. Growth promotion of rapeseed (Brassica napus) associated with the inoculation of phosphate solubilizing bacteria. Appl. Soil Ecol. 2018, 132, 1–10. [Google Scholar] [CrossRef]
- Hu, J.; Wei, Z.; Weidner, S.; Friman, V.P.; Xu, Y.C.; Shen, Q.R.; Jousset, A. Probiotic Pseudomonas communities enhance plant growth and nutrient assimilation via diversity-mediated ecosystem functioning. Soil Biol. Biochem. 2017, 113, 122–129. [Google Scholar] [CrossRef]
- Venieraki, A.; Dimou, M.; Vezyri, E.; Kefalogianni, I.; Argyris, N.; Liara, G.; Pergalis, P.; Chatzipavlidis, I.; Katinakis, P. Characterization of Nitrogen-Fixing Bacteria Isolated from Field-Grown Barley, Oat, and Wheat. J. Microbiol. 2011, 49, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sarma, M.V.R.K.; Saharan, K.; Srivastava, R.; Kumar, L.; Sahai, V.; Bisaria, V.S.; Sharma, A.K. Effect of formulated root endophytic fungus Piriformospora indica and plant growth promoting rhizobacteria fluorescent pseudomonads R62 and R81 on Vigna mungo. World J. Microbiol. Biotechnol. 2012, 28, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Mengual, C.; Schoebitz, M.; Azcon, R.; Roldan, A. Microbial inoculants and organic amendment improves plant establishment and soil rehabilitation under semiarid conditions. J. Environ. Manag. 2014, 134, 1–7. [Google Scholar] [CrossRef]
- Ortiz, N.; Armada, E.; Duque, E.; Roldán, A.; Azcón, R. Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought tolerance under natural soil conditions: Effectiveness of autochthonous or allochthonous strains. J. Plant Physiol. 2015, 174, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Wyciszkiewicz, M.; Saeid, A.; Dobrowolska-Iwanek, J.; Chojnacka, K. Utilization of microorganisms in the solubilization of low-quality phosphorus raw materiaL. Ecol. Eng. 2016, 89, 109–113. [Google Scholar] [CrossRef]
- Mittal, V.; Singh, O.; Nayyar, H.; Kaur, J.; Tewari, R. Stimulatory effect of phosphate-solubilizing fungal strains (Aspergillus awamori and Penicillium citrinum) on the yield of chickpea (Cicer arietinum L. cv. GPF2). Soil Biol. Biochem. 2008, 40, 718–727. [Google Scholar] [CrossRef]
- Dadrasan, M.; Chaichi, M.R.; Pourbabaee, A.A.; Yazdani, D.; Keshavarz-Afshar, R. Deficit irrigation and biological fertilizer influence on yield and trigonelline production of fenugreek. Ind. Crops Prod. 2015, 77, 156–162. [Google Scholar] [CrossRef]
- Sanders, E.R. Aseptic Laboratory Techniques: Plating Methods. JoVE 2012, 63, e3064. [Google Scholar] [CrossRef]
- SR, O. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Misc. Pap. Inst. Agric. Res. 1954, 939, 1–19. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Igual, J.M.; Valverde, A.; Cervantes, E.; Velazquez, E. Phosphate-solubilizing bacteria as inoculants for agriculture: Use of updated molecular techniques in their study. Agronomie 2001, 21, 561–568. [Google Scholar] [CrossRef]
- Tejada, M.; Gonzalez, J.L.; Hernandez, M.T.; Garcia, C. Application of different organic amendments in a gasoline contaminated soil: Effect on soil microbial properties. Bioresour. Technol. 2008, 99, 2872–2880. [Google Scholar] [CrossRef] [PubMed]
- Jing, Z.W.; Chen, R.R.; Wei, S.P.; Feng, Y.Z.; Zhang, J.B.; Lin, X.G. Response and feedback of C mineralization to P availability driven by soil microorganisms. Soil Biol. Biochem. 2017, 105, 111–120. [Google Scholar] [CrossRef]
- Tahir, M.; Khalid, U.; Ijaz, M.; Shah, G.M.; Naeem, M.A.; Shahid, M.; Mahmood, K.; Ahmad, N.; Kareem, F. Combined application of bio-organic phosphate and phosphorus solubilizing bacteria (Bacillus strain MWT 14) improve the performance of bread wheat with low fertilizer input under an arid climate. Braz. J. Microbiol. 2018, 49 (Suppl. S1), 15–24. [Google Scholar] [CrossRef] [PubMed]
- Wani, P.; Khan, M.; Zaidi, A. Co-inoculation of nitrogen-fixing and phosphate-solubilizing bacteria to promote growth, yield and nutrient uptake in chickpea. Acta Agron. Hung. 2007, 55, 315–323. [Google Scholar] [CrossRef]
- Vassilev, N.; Vassileva, M.; Fenice, M.; Federici, F. Immobilized cell technology applied in solubilization of insoluble inorganic (rock) phosphates and P plant acquisition. Bioresour. Technol. 2001, 79, 263–271. [Google Scholar] [CrossRef]
- Shanmugam, S.R.; Chaganti, S.R.; Lalman, J.A.; Heath, D.D. Effect of inhibitors on hydrogen consumption and microbial population dynamics in mixed anaerobic cultures. Int. J. Hydrog. Energy 2014, 39, 249–257. [Google Scholar] [CrossRef]
- Paerl, H.W.; Pinckney, J.L. A mini-review of microbial consortia: Their roles in aquatic production and biogeochemical cycling. Microb. Ecol. 1996, 31, 225–247. [Google Scholar] [CrossRef]
- Wei, Y.; Zhao, Y.; Shi, M.; Cao, Z.; Lu, Q.; Yang, T.; Fan, Y.; Wei, Z. Effect of organic acids production and bacterial community on the possible mechanism of phosphorus solubilization during composting with enriched phosphate-solubilizing bacteria inoculation. Bioresour. Technol. 2018, 247, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Saxena, J.; Rana, G.; Pandey, M. Impact of addition of biochar along with Bacillus sp on growth and yield of French beans. Sci. Horticult. 2013, 162, 351–356. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Hadas, A.; Sofer, M.; Molina, J.A.E.; Barak, P.; Clapp, C.E. Assimilation of nitrogen by soil microbial population: NH4 versus organic N. Soil Biol. Biochem. 1992, 24, 137–143. [Google Scholar] [CrossRef]
- Yang, L.J.; Zhang, L.L.; Geisseler, D.; Wu, Z.J.; Gong, P.; Xue, Y.; Yu, C.X.; Juan, Y.H.; Horwath, W.R. Available C and N affect the utilization of glycine by soil microorganisms. Geoderma 2016, 283, 32–38. [Google Scholar] [CrossRef]
- Wang, H.Y.; Nie, Y.; Butterly, C.R.; Wang, L.; Chen, Q.H.; Tian, W.; Song, B.B.; Xi, Y.G.; Wang, Y. Fertilization alters microbial community composition and functional patterns by changing the chemical nature of soil organic carbon: A field study in a Halosol. Geoderma 2017, 292, 17–24. [Google Scholar] [CrossRef]
- Cai, F.; Chen, W.; Wei, Z.; Pang, G.; Li, R.X.; Ran, W.; Shen, Q.R. Colonization of Trichoderma harzianum strain SQR-T037 on tomato roots and its relationship to plant growth, nutrient availability and soil microflora. Plant Soil 2015, 388, 337–350. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, H.; Guo, Y.; Tian, T. Influence of Chicken Manure Fertilization on Antibiotic-Resistant Bacteria in Soil and the Endophytic Bacteria of Pakchoi. Int. J. Environ. Res. Public Health 2016, 13, 662. [Google Scholar] [CrossRef]
- Marschner, P.; Kandeler, E.; Marschner, B. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 2003, 35, 453–461. [Google Scholar] [CrossRef]
- Frey, S.D.; Knorr, M.; Parrent, J.L.; Simpson, R.T. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests. Ecol. Manag. 2004, 196, 159–171. [Google Scholar] [CrossRef]
- Liu, J.X.; Huang, W.J.; Zhou, G.Y.; Zhang, D.Q.; Liu, S.Z.; Li, Y.Y. Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests. Glob. Chang. Biol. 2013, 19, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Li, Y.Y.; Xu, Y.; Liu, S.G.; Huang, W.J.; Fang, X.; Yin, G.C. Phosphorus uptake in four tree species under nitrogen addition in subtropical China. Environ. Sci. Pollut. R 2017, 24, 20005–20014. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Bolan, N. Lead immobilization and bioavailability in microbial and root interface. J. Hazard. Mater. 2013, 261, 777–783. [Google Scholar] [CrossRef] [PubMed]
- De Bolle, S.; Gebremikael, M.T.; Maervoet, V.; De Neve, S. Performance of phosphate-solubilizing bacteria in soil under high phosphorus conditions. Biol. Fert. Soils 2013, 49, 705–714. [Google Scholar] [CrossRef]
- Sanz-Saez, A.; Heath, K.D.; Burke, P.V.; Ainsworth, E.A. Inoculation with an enhanced N-2-fixing Bradyrhizobium japonicum strain (USDA110) does not alter soybean (Glycine maxMerr.) response to elevated [CO2]. Plant Cell Environ. 2015, 38, 2589–2602. [Google Scholar] [CrossRef] [PubMed]
- Porto, D.S.; Farias, E.D.C.; Chaves, J.D.; Souza, B.F.; de Medeiros, R.D.; Zilli, J.E.; da Silva, K. Symbiotic effectiveness of Bradyrhizobium ingae in promoting growth of Inga edulis Mart. seedlings. Rev. Bras. Cienc. Solo 2017, 41, e0160222. [Google Scholar] [CrossRef]
- Hacquard, S.; Garrido-Oter, R.; Gonzalez, A.; Spaepen, S.; Ackermann, G.; Lebeis, S.; McHardy, A.C.; Dangl, J.L.; Knight, R.; Ley, R.; et al. Microbiota and Host Nutrition across Plant and Animal Kingdoms. Cell Host Microbe 2015, 17, 603–616. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inoculants Type | Treatment Code | M: Bacillus megaterium | F: Pseudomonas fluorescens | C: Azotobacter chroococcum | B: Azospirillum brasilence |
|---|---|---|---|---|---|
| SBA | M | 5 | 0 | 0 | 0 |
| F | 0 | 5 | 0 | 0 | |
| C | 0 | 0 | 5 | 0 | |
| B | 0 | 0 | 0 | 5 | |
| MBA | MF | 2.5 | 2.5 | 0 | 0 |
| MC | 2.5 | 0 | 2.5 | 0 | |
| MB | 2.5 | 0 | 0 | 2.5 | |
| FC | 0 | 2.5 | 2.5 | 0 | |
| FB | 0 | 2.5 | 0 | 2.5 | |
| CB | 0 | 0 | 2.5 | 2.5 | |
| MFCB | 1.25 | 1.25 | 1.25 | 1.25 | |
| Control | Control | 0 | 0 | 0 | 0 |
| Treatment Code | Incubation Duration (d) | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 5 | 10 | 15 | 20 | 30 | 45 | 60 | |
| MFCB | 0.167 ± 0 | 42.7 ± 5.3Aa | 43.3 ± 4.7Aa | 9.7 ± 1.5Dc | 13.6 ± 3Cbc | 20 ± 0.6Bd | 7.3 ± 1DEbcde | 3.7 ± 1Fefg |
| MF | 0.167 ± 0 | 18C ± 0.3c | 33.3 ± 1.9Bb | 15.67 ± 1.3Da | 10 ± 1.2Ecd | 38 ± 1.2Aa | 15.33 ± D1a | 6.1 ± 1Fef |
| FC | 0.167 ± 0 | 17 ± 6Bc | 17 ± 5.5Bc | 7.2 ± 1DEde | 25.6 ± 3.8Aa | 26.6 ± 3.2Ac | 8.7 ± 2.8CDbc | 10.5 ± 2Ccd |
| MB | 0.167 ± 0 | 12.9 ± 2BCc | 15 ± 1.6ABc | 15.3 ± 2ABa | 13.8 ± 1Bc | 18.3 ± 2Af | 12.8 ± 2.7BCab | 14.3 ± 0.3Bc |
| FB | 0.167 ± 0 | 33 ± 4.3Ab | 10.3 ± 4.1Ccde | 12 ± 2Cb | 5.3 ± 1DEefg | 26.3 ± 3.5Bc | 10 ± 3.2Cabc | 7.3 ± 2Dde |
| CB | 0.167 ± 0 | 7.3 ± 2Dd | 4.8 ± 2.3DEFde | 5.9 ± 1.8DEde | 11 ± 1Ccd | 28.3 ± 5.7Ac | 12.7 ± 5Cab | 22 ± 5.3Ba |
| MC | 0.167 ± 0 | 16.7 ± 5Bc | 12.1 ± 0.2Ccd | 7 ± 1DEde | 4.8 ± 1.8EFfg | 32 ± 2.6Ab | 10 ± 3.5CDabc | 3.3 ± 1FGefg |
| B | 0.167 ± 0 | 5.3 ± 0.6Ed | 4.3 ± 2.2EFe | 8.0 ± 1Dde | 17.2 ± 3.5Ab | 16 ± 0.8ABe | 15.3 ± 4ABCa | 12.7 ± 3Cc |
| C | 0.167 ± 0 | 5.3 ± 3.2DEFd | 9.7 ± 1.5Ccde | 14.0 ± 0.5Bc | 9.3 ± 3Cde | 6.0 ± 1DEf | 7.7 ± 1CDbcd | 17.3 ± 2Ab |
| F | 0.167 ± 0 | 8.3 ± 1.5Ad | 7.1 ± 2.5ABde | 5.0 ± 0.4BCe | 7.1 ± 3ABdefg | 5.0 ± 0.8BCf | 1.8 ± 0.8De | 1.9 ± 0.7Dg |
| M | 0.167 ± 0 | 5.6 ± 1.5BCd | 4.2 ± 1.6CDEe | 7.7 ± 1.5ABcd | 8.3 ± 3Adef | 5.0 ± 0.6CDf | 4.5 ± 0CDEcde | 3.6 ± 1DEefg |
| Variables | Inoculants | Incubation Duration | Inoculants × Incubation Duration | |||
|---|---|---|---|---|---|---|
| F-test | Sig. | F-test | Sig. | F-test | Sig. | |
| TC | 1.82 | nd | 54.48 | ** | 2.03 | * |
| TN | 1.86 | nd | 43.11 | ** | 2.93 | ** |
| NH4+-N | 10.90 | ** | 24.57 | ** | 3.11 | ** |
| NO3--N | 20.27 | ** | 48.50 | ** | 5.87 | ** |
| SAP | 26.58 | ** | 2162.43 | ** | 21.40 | ** |
| AcPase | 24.42 | ** | 67.18 | ** | 21.34 | ** |
| IN | 19.18 | ** | 75.34 | ** | 7.65 | ** |
| Inoculants | TC | TN | NH4+-N | NO3--N | SAP | AcPase | IN | |
|---|---|---|---|---|---|---|---|---|
| 30d-60d | 30d-60d | 30d-60d | 30d-60d | 30d-60d | 30d-60d | 30d-60d | ||
| SBA | M | ** | ** | ** | ** | nd | ** | |
| F | nd | * | * | * | ** | * | ** | |
| C | * | nd | nd | nd | ** | nd | nd | |
| B | nd | nd | ** | nd | ** | nd | ** | |
| MBA | MF | ** | ** | ** | * | ** | ** | ** |
| MC | ** | * | nd | ** | ** | ** | ** | |
| MB | * | * | nd | ** | ** | * | nd | |
| FC | * | ** | nd | ** | ** | ** | nd | |
| FB | * | ** | * | ** | ** | ** | ** | |
| CB | nd | nd | * | nd | ** | * | ** | |
| MFCB | nd | nd | * | nd | ** | ** | ** | |
| Control | CK | nd | nd | nd | nd | nd | nd | nd |
| Inoculants | TC (g·kg–1) | TN (g·kg–1) | IN (mg·kg–1) | ||||
|---|---|---|---|---|---|---|---|
| 30 d | 60 d | 30 d | 60 d | 30 d | 60 d | ||
| SBA | M | 4.6 ± 0.2a | 4.0 ± 0.3a * | 0.9 ± 0.1a | 0.7 ± 0.1a * | 34.8 ± 1.6b | 19.6 ± 2.4cd * |
| F | 4.1 ± 0.2a-d | 3.9 ± 0.1a | 0.9 ± 0.1ab | 0.7 ± 0.1a * | 34.3 ± 2.5bc | 23.9 ± 6.6abc * | |
| C | 4.3 ± 0.2a-d | 4.0 ± 0.2a * | 0.7 ± 0.1c | 0.7 ± 0.1a | 10.7 ± 2.8f | 9.7 ± 3.5f | |
| B | 4.1 ± 0.1cd | 3.9 ± 0.1a | 0.8 ± 0.1bc | 0.7 ± 0.1a | 30.4 ± 8.5cd | 8.7 ± 1.2f * | |
| MBA | MF | 4.5 ± 0.3ab | 4.0 ± 0.6a * | 0.9 ± 0.0a | 0.6 ± 0.1a * | 43.1 ± 4.8a | 28.6 ± 3.6ab * |
| MC | 4.5 ± 0.4a | 3.8 ± 0.1a * | 0.8 ± 0.1abc | 0.7 ± 0.1a * | 23.7 ± 7.4de | 17.5 ± 4.2cde * | |
| MB | 4.4 ± 0.1abc | 3.9 ± 0.1a * | 0.8 ± 0.1abc | 0.7 ± 0.1a * | 25.9 ± 5.5d | 22.3 ± 2.2bc | |
| FC | 4.2 ± 0.2a-d | 3.8 ± 0.1a * | 0.9 ± 0.1ab | 0.7 ± 0.1a * | 22.4 ± 3.4de | 18.4 ± 4.9cde | |
| FB | 4.2 ± 0.1bcd | 3.8 ± 0.1a * | 0.9 ± 0.1ab | 0.7 ± 0.1a * | 27.7 ± 1.3cd | 18.9 ± 2.5cd * | |
| CB | 4.2 ± 0.1cd | 4.2 ± 0.1a | 0.7 ± 0.1c | 0.7 ± 0.1a | 21.2 ± 4.7bc | 12.2 ± 0.9def * | |
| MFCB | 4.3 ± 0.1a-d | 4.1 ± 0.1a | 0.8 ± 0.1abc | 0.7 ± 0.1a | 20.2 ± 0.2e | 30.8 ± 12.8a * | |
| CK | CK | 4.0 ± 0.1d | 4.0 ± 0.2a | 0.7 ± 0.01c | 0.7 ± 0.0a | 10.9 ± 1.4f | 10.8 ± 0.1ef |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Chen, Z.; Fu, X. Integrated Effects of Co-Inoculation with Phosphate-Solubilizing Bacteria and N2-Fixing Bacteria on Microbial Population and Soil Amendment Under C Deficiency. Int. J. Environ. Res. Public Health 2019, 16, 2442. https://doi.org/10.3390/ijerph16132442
Wang Z, Chen Z, Fu X. Integrated Effects of Co-Inoculation with Phosphate-Solubilizing Bacteria and N2-Fixing Bacteria on Microbial Population and Soil Amendment Under C Deficiency. International Journal of Environmental Research and Public Health. 2019; 16(13):2442. https://doi.org/10.3390/ijerph16132442
Chicago/Turabian StyleWang, Zhikang, Ziyun Chen, and Xiangxiang Fu. 2019. "Integrated Effects of Co-Inoculation with Phosphate-Solubilizing Bacteria and N2-Fixing Bacteria on Microbial Population and Soil Amendment Under C Deficiency" International Journal of Environmental Research and Public Health 16, no. 13: 2442. https://doi.org/10.3390/ijerph16132442
APA StyleWang, Z., Chen, Z., & Fu, X. (2019). Integrated Effects of Co-Inoculation with Phosphate-Solubilizing Bacteria and N2-Fixing Bacteria on Microbial Population and Soil Amendment Under C Deficiency. International Journal of Environmental Research and Public Health, 16(13), 2442. https://doi.org/10.3390/ijerph16132442