Exogenous Glycinebetaine Promotes Soil Cadmium Uptake by Edible Amaranth Grown during Subtropical Hot Season

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Crops and Soil
2.2. Sample Determination
2.3. Data Analysis
3. Results and Discussion
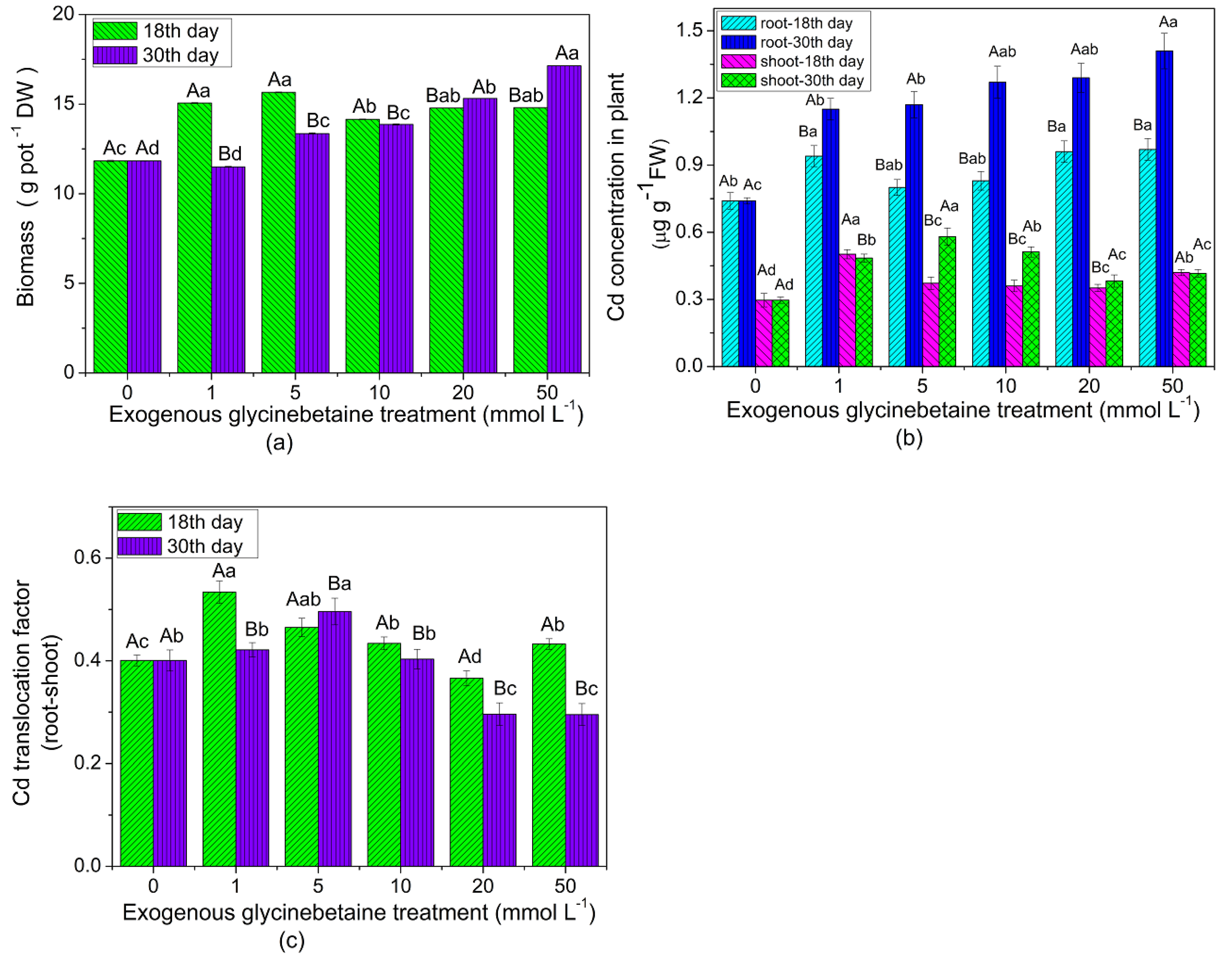
3.1. Biomass and Cd Concentration in Edible Amaranth after Exogenous Glycinebetaine Treatment at Two Growth Stages
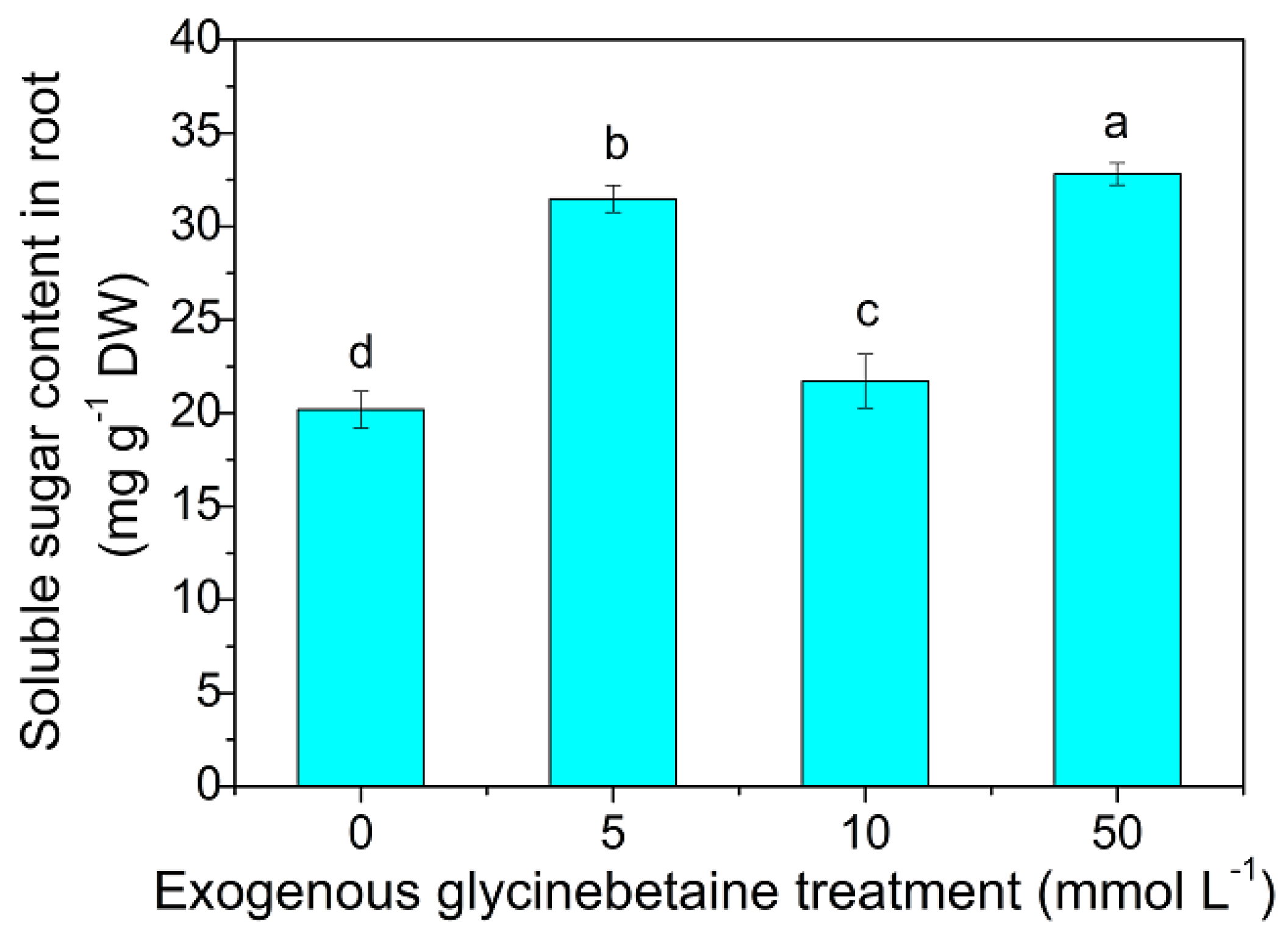
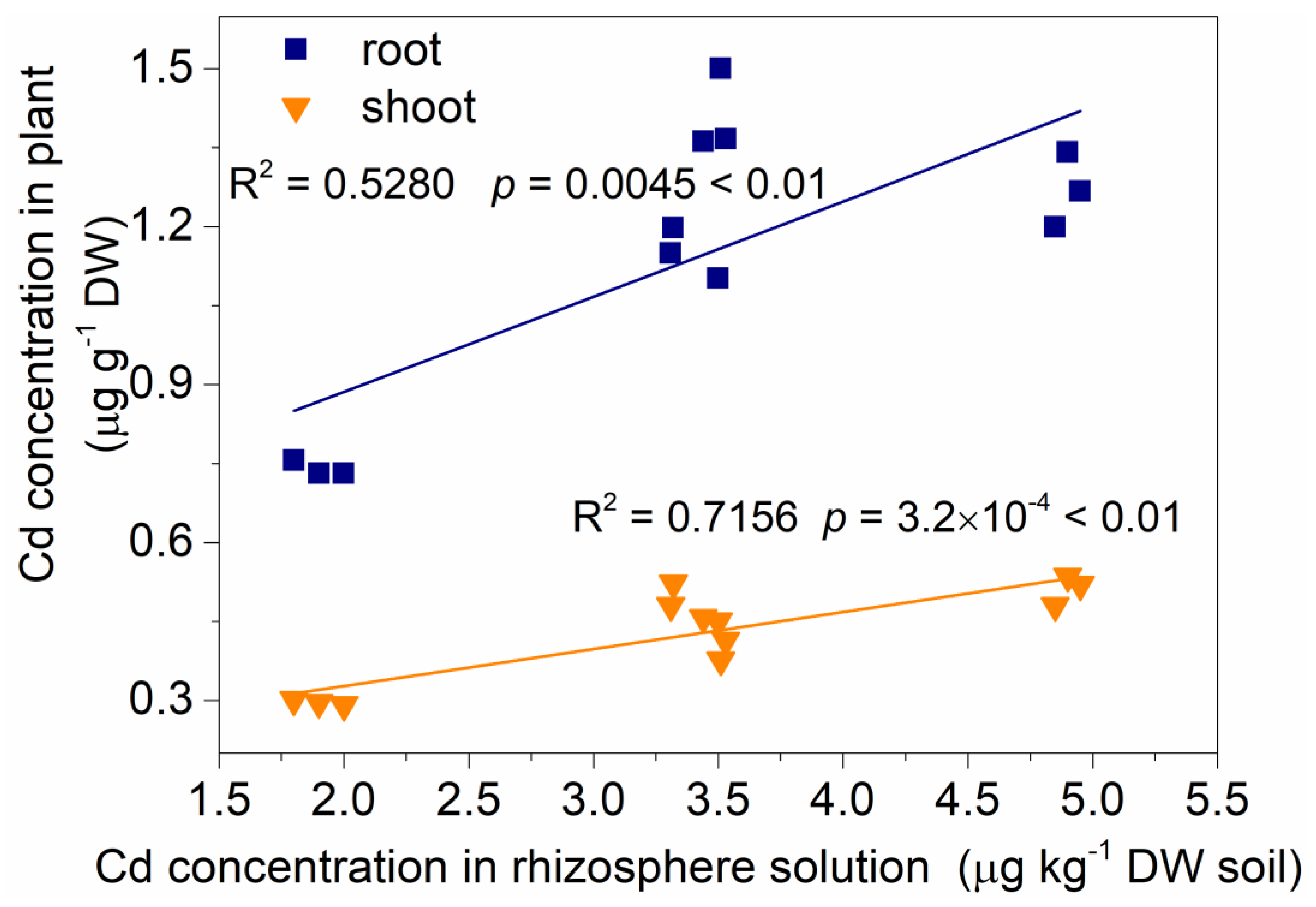
3.2. Soluble Sugar Content in the Amaranth Roots, Contents of Soluble Sugar, DOC, and LMWOAs in the Rhizosphere Solution and their Relationship with Cd Mobilization in Rhizosphere Soil
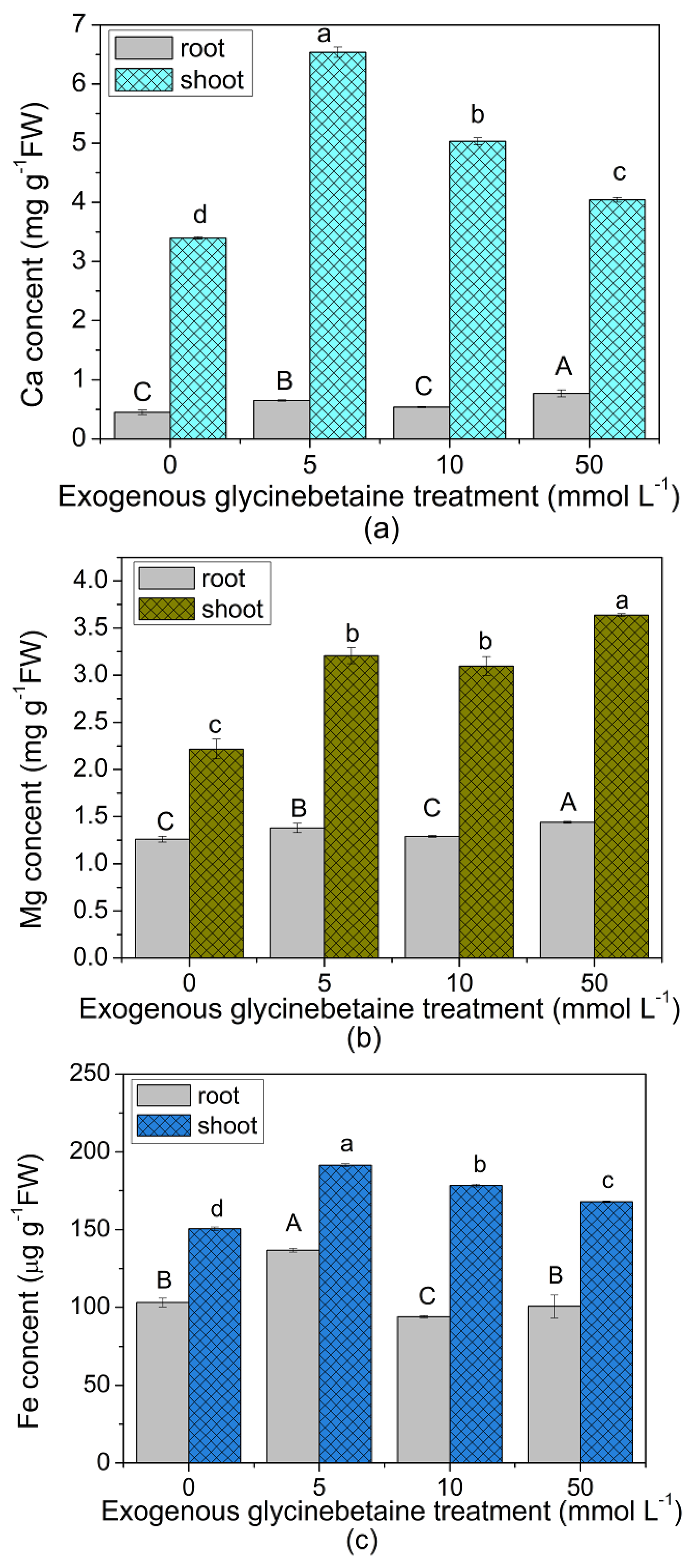
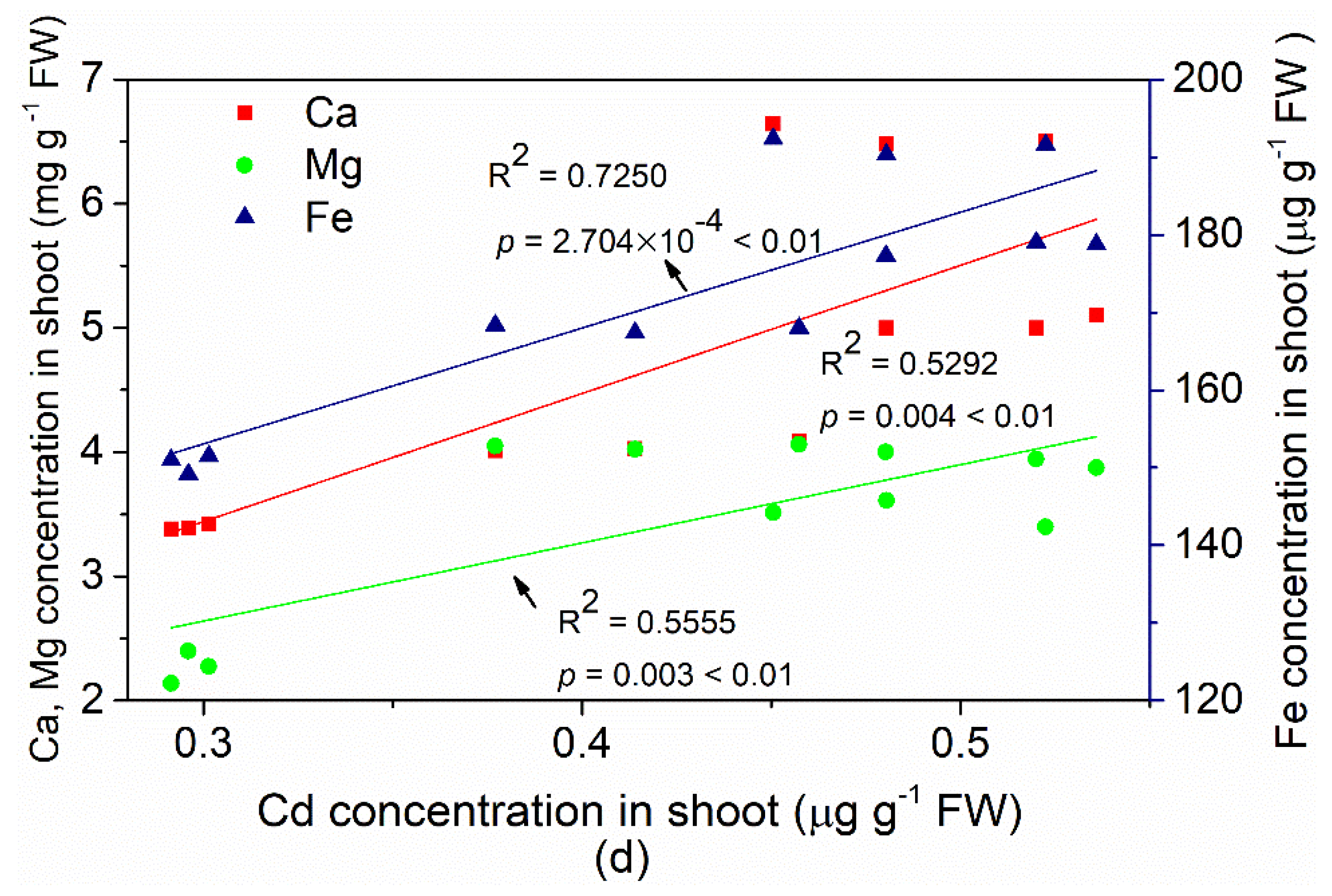
3.3. Changes in Ca, Mg and Fe Contents in the Leaves and their Relationship with Cd Uptake by Edible Amaranth
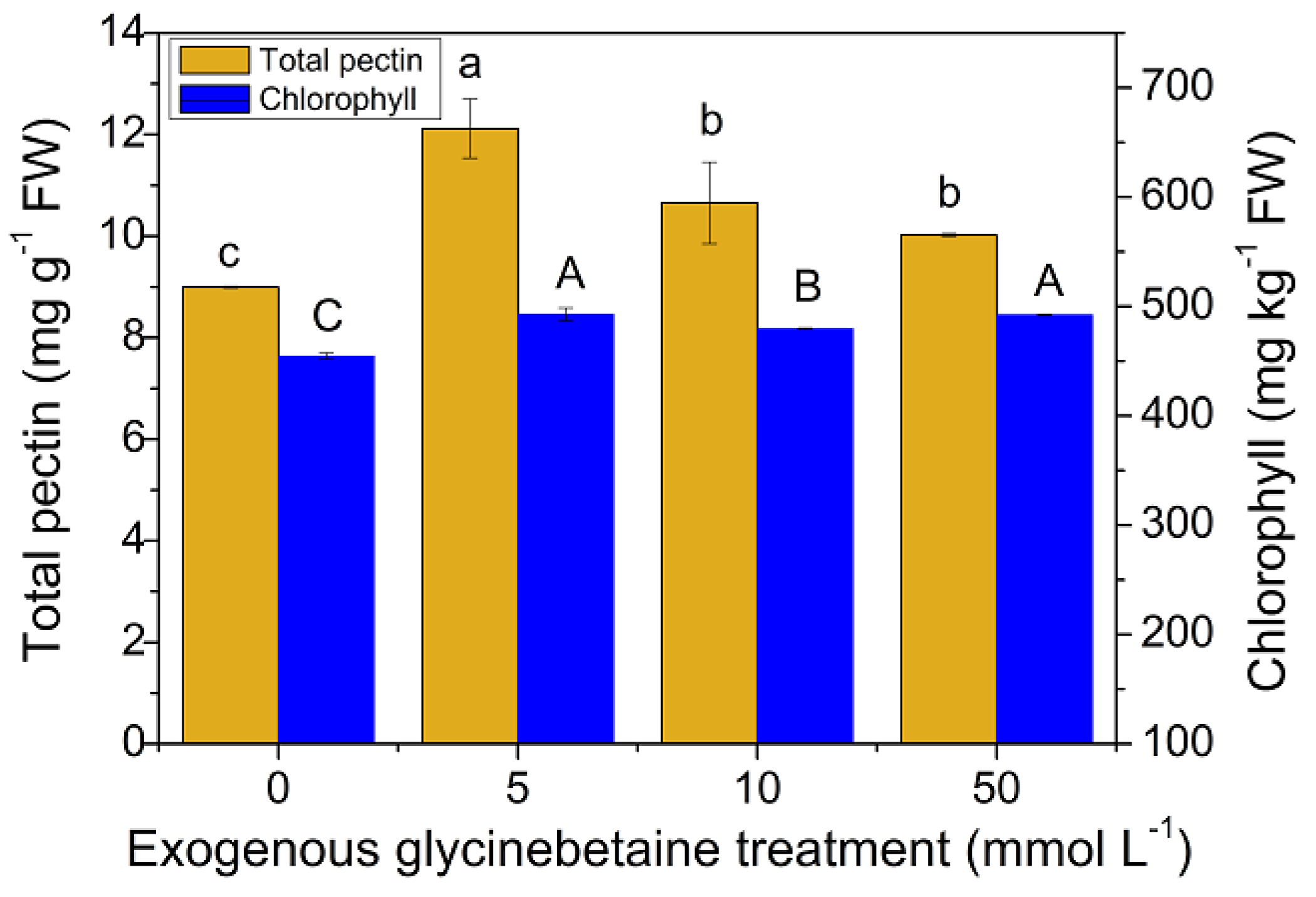
3.4. Changes in the Pectin and Chlorophyll Contents and Their Relationship with the Cd Uptake and Accumulation in Amaranth Leaves
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Bio/Technol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Nusrat, N.; Shahbaz, M.; Perveen, S. Modulation in growth, photosynthetic efficiency, activity of antioxidants and mineral ions by foliar application of glycinebetaine on pea (Pisum sativum L.) under salt stress. Acta Physiol. Plant. 2014, 36, 2985–2998. [Google Scholar] [CrossRef]
- Wei, D.; Zhang, W.; Wang, C.; Meng, Q.; Li, G.; Chen, T.H.H.; Yang, X. Genetic engineering of the biosynthesis of glycinebetaine leads to alleviate salt-induced potassium efflux and enhances salt tolerance in tomato plants. Plant Sci. 2017, 257, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Salama, K.H.A.; Mansour, M.M.; Al-Malawi, H.A. Glycinebetaine priming improves salt tolerance of wheat. Biologia 2015, 70, 1334–1339. [Google Scholar] [CrossRef]
- Korkmaz, A.; Şirikçi, R.; Kocaçınar, F.; Değer, Ö.; Demirkırıan, A.R. Alleviation of salt-induced adverse effects in pepper seedlings by seed application of glycinebetaine. Sci. Hortic. 2012, 148, 197–205. [Google Scholar] [CrossRef]
- Anjum, S.A.; Farooq, M.; Wang, L.C.; Xue, L.L.; Wang, S.G.; Wang, L.; Zhang, S.; Chen, M. Gas exchange and chlorophyll synthesis of maize cultivars are enhanced by exogenously-applied glycinebetaine under drought conditions. Plant Soil Environ. 2011, 57, 326–331. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Saleem, M.F.; Jamil, M.; Khan, I.H. Impact of foliar applied glycinebetaine on growth and physiology of wheat (Triticum aestivum L.) under drought conditions. Pak. J. Agric. Sci. 2014, 51, 327–334. [Google Scholar]
- Park, E.-J.; Jeknic, Z.; Chen, T.H.H. Exogenous application of glycinebetaine increases chilling tolerance in tomato plants. Plant Cell Physiol. 2006, 47, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Kanechi, M.; Hikosaka, Y.; Uno, Y. Application of sugarbeet pure and crude extracts containing glycinebetaine affects root growth, yield, and photosynthesis of tomato grown during summer. Sci. Hortic. 2013, 152, 9–15. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Wang, J.; Zhang, W.; Meng, Q.; Chen, T.H.H.; Murata, N.; Yang, X. Glycinebetaine enhances the tolerance of tomato plants to high temperature during germination of seeds and growth of seedlings. Plant Cell Environ. 2011, 34, 1931–1943. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, P.; Jokinen, K.; Kontturi, M.; Peltonen-Sainio, P.; Pehu, E.; Somersalo, S. Foliar application of glycinebetaine—A novel product from sugar beet—As an approach to increase tomato yield. Ind. Crop. Prod. 1998, 7, 139–148. [Google Scholar] [CrossRef]
- Mäkelä, P.; Kontturi, M.; Pehu, E.; Somersalo, S. Photosynthetic response of drought- and salt-stressed tomato and turnip rape plants to foliar-applied glycinebetaine. Physiol. Plant. 1999, 105, 45–50. [Google Scholar] [CrossRef]
- Alia; Kondo, Y.; Sakamoto, A.; Nonaka, H.; Hayashi, H.; Saradhi, P.P.; Chen, T.H.H.; Murata, N. Enhanced tolerance to light stress of transgenic arabidopsis plants that express the coda gene for a bacterial choline oxidase. Plant Mol. Biol. 1999, 40, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-P.; Tian, F.-X.; Zhang, M.; Wang, W. The overaccumulation of glycinebetaine alleviated damages to psii of wheat flag leaves under drought and high temperature stress combination. Acta Physiol. Plant. 2014, 36, 2743–2753. [Google Scholar] [CrossRef]
- Alia; Hayashi, H.; Chen, T.H.H.; Murata, N. Transformation with a gene for choline oxidase enhances the cold tolerance of arabidopsis during germination and early growth. Plant Cell Environ. 1998, 21, 232–239. [Google Scholar] [CrossRef]
- Wang, G.P.; Zhang, X.Y.; Li, F.; Luo, Y.; Wang, W. Overaccumulation of glycine betaine enhances tolerance to drought and heat stress in wheat leaves in the protection of photosynthesis. Photosynthetica 2010, 48, 117–126. [Google Scholar] [CrossRef]
- Lin, X.; Mou, R.; Cao, Z.; Xu, P.; Wu, X.; Zhu, Z.; Chen, M. Characterization of cadmium-resistant bacteria and their potential for reducing accumulation of cadmium in rice grains. Sci. Total Environ. 2016, 569–570, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Bian, R.; Joseph, S.; Cui, L.; Pan, G.; Li, L.; Liu, X.; Zhang, A.; Rutlidge, H.; Wong, S.; Chia, C.; et al. A three-year experiment confirms continuous immobilization of cadmium and lead in contaminated paddy field with biochar amendment. J. Hazard. Mater. 2014, 272, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Hoque, M.A.; Okuma, E.; Banu, M.N.A.; Shimoishi, Y.; Nakamura, Y.; Murata, Y. Exogenous proline and glycinebetaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J. Plant Physiol. 2009, 166, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Ashraf, M. Exogenously applied glycinebetaine enhances seed and seed oil quality of maize (Zea mays L.) under water deficit conditions. Environ. Exp. Bot. 2011, 71, 249–259. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hasanuzzaman, M.; Fujita, M. Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants 2010, 16, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Liu, L.; Ibrahim, W.; Cai, Y.; Wu, F. Alleviating effects of exogenous glutathione, glycinebetaine, brassinosteroids and salicylic acid on cadmium toxicity in rice seedlings (Oryza sativa). Agrotechnology 2013, 2, 107. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Bharwana, S.A.; Rizwan, M.; Ishaque, W.; Farid, M.; Mahmood, K.; Iqbal, Z. Cadmium stress in cotton seedlings: Physiological, photosynthesis and oxidative damages alleviated by glycinebetaine. S. Afr. J. Bot. 2016, 104, 61–68. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, J.; Zhu, L.; Yang, G.; Li, D. Transfer of Cadmium from Soil to Vegetable in the Pearl River Delta area, South China. PLoS ONE 2014, 9, e108572. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, Y.; Fu, H.; Cui, Z.; Shi, L.; Wang, L.; Liu, Z. Health risk of heavy metals in food crops grown on reclaimed tidal flat soil in the Pearl River Estuary, China. J. Hazard. Mater. 2012, 227–228, 148–154. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, Y.; Fang, B.; Li, C.; Yang, J.; Teng, Z.; He, X.; Zhang, Y. Effect of temperature on cadmium content of brown rice in different cultrivars. China Rice (Chin.) 2016, 22, 31–35. [Google Scholar]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Xu, W.; Wang, C.-W.; Wang, W.-G.; Zhang, J.-F. Quick determination of pectin in pectin extract from premna microphylla. Food Drug (Chin.) 2006, 8, 53–54. [Google Scholar]
- Li, Q.; Cai, S.; Mo, C.; Chu, B.; Peng, L.; Yang, F. Toxic effects of heavy metals and their accumulation in vegetables grown in a saline soil. Ecotoxicol. Environ. Saf. 2010, 73, 84–88. [Google Scholar] [CrossRef] [PubMed]
- He, B.-Y.; Yu, D.-P.; Chen, Y.; Shi, J.-L.; Xia, Y.; Li, Q.-S.; Wang, L.-L.; Ling, L.; Zeng, E.Y. Use of low-calcium cultivars to reduce cadmium uptake and accumulation in edible amaranth (Amaranthus mangostanus L.). Chemosphere 2017, 171, 588–594. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Ling, L.; Zhang, L.; Li, M.; Li, Q.; Mei, X.; Li, H.; Tan, L. Cultivar-specific differences in heavy metal (Cd, Cr, Cu, Pb, and Zn) concentrations in water spinach (Ipomoea aquatic ‘Forsk’) grown on metal-contaminated soil. Plant Soil 2015, 386, 251–262. [Google Scholar] [CrossRef]
- Gao, Y.; Li, C.; Lou, K. Effect of spraying glycine betaine on physiological responses of processing tomato under drought stress. J. Plant Nutr. Fertil. (Chin.) 2012, 18, 426–432. [Google Scholar]
- Dessureault-Rompré, J.; Luster, J.; Schulin, R.; Tercier-Waeber, M.-L.; Nowack, B. Decrease of labile Zn and Cd in the rhizosphere of hyperaccumulating Thlaspi caerulescens with time. Environ. Pollut. 2010, 158, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Yanai, J.; Zhao, F.-J.; McGrath, S.P.; Kosaki, T. Effect of soil characteristics on Cd uptake by the hyperaccumulator Thlaspi caerulescens. Environ. Pollut. 2006, 139, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-M.; Li, Q.-S.; Yang, P.; Ye, H.-J.; Chen, Z.-S.; Guo, S.-H.; Wang, L.-L.; He, B.-Y.; Zeng, E.Y. Impact of osmoregulation on the differences in cd accumulation between two contrasting edible amaranth cultivars grown on cd-polluted saline soils. Environ. Pollut. 2017, 224, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Vaculík, M.; Konlechner, C.; Langer, I.; Adlassnig, W.; Puschenreiter, M.; Lux, A.; Hauser, M.-T. Root anatomy and element distribution vary between two Salix caprea isolates with different Cd accumulation capacities. Environ. Pollut. 2012, 163, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Carrier, P.; Baryla, A.; Havaux, M. Cadmium distribution and microlocalization in oilseed rape (Brassica napus) after long-term growth on cadmium-contaminated soil. Planta 2003, 216, 939–950. [Google Scholar] [PubMed]
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, N.K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Lu, L.; Tian, S.; Zhang, M.; Zhang, J.; Yang, X.; Jiang, H. The role of Ca pathway in Cd uptake and translocation by the hyperaccumulator Sedum alfredii. J. Hazard. Mater. 2010, 183, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Goel, D.; Singh, A.K.; Yadav, V.; Babbar, S.B.; Murata, N.; Bansal, K.C. Transformation of tomato with a bacterial codA gene enhances tolerance to salt and water stresses. J. Plant Physiol. 2011, 168, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, P.; Kärkkäinen, J.; Somersalo, S. Effect of glycinebetaine on chloroplast ultrastructure, chlorophyll and protein content, and RuBPCO activities in tomato grown under drought or salinity. Biol. Plant. 2000, 43, 471–475. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Zou, Y.-N.; He, X.-H. Contributions of arbuscular mycorrhizal fungi to growth, photosynthesis, root morphology and ionic balance of citrus seedlings under salt stress. Acta Physiol. Plant. 2010, 32, 297–304. [Google Scholar] [CrossRef]
- Brunner, I.; Luster, J.; Günthardt-Goerg, M.S.; Frey, B. Heavy metal accumulation and phytostabilisation potential of tree fine roots in a contaminated soil. Environ. Pollut. 2008, 152, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GB/mM | Cdtotal (µg pot−1) | Rhizosphere Soil Solution | ||
|---|---|---|---|---|
| DOC (µg g−1 DW) | LMWOAs (µg g−1 DW) | Soluble Sugar (µg kg−1 DW) | ||
| 0 | 1.02 ± 0.02d | 64.5 ± 1.01d | 1.24 ± 0.14d | 6.14 ± 0.21d |
| 5 | 1.63 ± 0.01c | 82.0 ± 1.23c | 3.65 ± 0.11c | 12.0 ± 0.10c |
| 10 | 1.77 ± 0.04a | 90.1 ± 0.80b | 5.75 ± 0.29a | 15.3 ± 0.45b |
| 50 | 1.69 ± 0.02b | 93.1 ± 0.20a | 4.51 ± 0.21b | 16.2 ± 0.19a |
| GB average | 1.70 | 88.4 | 4.64 | 14.5 |
| GB average/0 (fold) | 1.66 | 1.37 | 3.73 | 2.36 |
| Statistical Indicators | DOC | LMWOAs | Soluble Sugar |
|---|---|---|---|
| R2-value | 0.874 | 0.867 | 0.855 |
| p-value | 0.043 < 0.05 | 0.045 < 0.05 | 0.049 < 0.05 |
| Ca | Mg | Fe | Pectin | Chlorophyll | |
|---|---|---|---|---|---|
| Ca | 1 | ||||
| Mg | 0.095 | 1 | |||
| Fe | 0.924 * | 0.344 | 1 | ||
| Total pectin | 0.856 * | 0.171 | 0.448 | 1 | |
| Chlorophyll | 0.370 | 0.679 * | 0.589 * | 0.479 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, W.-Q.; Lei, Y.-K.; Yang, P.; Li, Q.-S.; Wang, L.-L.; He, B.-Y.; Xu, Z.-M.; Zhou, C.; Ye, H.-J. Exogenous Glycinebetaine Promotes Soil Cadmium Uptake by Edible Amaranth Grown during Subtropical Hot Season. Int. J. Environ. Res. Public Health 2018, 15, 1794. https://doi.org/10.3390/ijerph15091794
Yao W-Q, Lei Y-K, Yang P, Li Q-S, Wang L-L, He B-Y, Xu Z-M, Zhou C, Ye H-J. Exogenous Glycinebetaine Promotes Soil Cadmium Uptake by Edible Amaranth Grown during Subtropical Hot Season. International Journal of Environmental Research and Public Health. 2018; 15(9):1794. https://doi.org/10.3390/ijerph15091794
Chicago/Turabian StyleYao, Wei-Qing, Yong-Kang Lei, Ping Yang, Qu-Sheng Li, Li-Li Wang, Bao-Yan He, Zhi-Min Xu, Chu Zhou, and Han-Jie Ye. 2018. "Exogenous Glycinebetaine Promotes Soil Cadmium Uptake by Edible Amaranth Grown during Subtropical Hot Season" International Journal of Environmental Research and Public Health 15, no. 9: 1794. https://doi.org/10.3390/ijerph15091794
APA StyleYao, W.-Q., Lei, Y.-K., Yang, P., Li, Q.-S., Wang, L.-L., He, B.-Y., Xu, Z.-M., Zhou, C., & Ye, H.-J. (2018). Exogenous Glycinebetaine Promotes Soil Cadmium Uptake by Edible Amaranth Grown during Subtropical Hot Season. International Journal of Environmental Research and Public Health, 15(9), 1794. https://doi.org/10.3390/ijerph15091794