Susceptibility of Multidrug-Resistant Bacteria, Isolated from Water and Plants in Nigeria, to Ceragenins

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Maintenance of Bacterial Isolates
2.2. Identification of Isolates
2.3. Susceptibility Testing
2.4. Scanning Electron Microscopy (SEM)
2.5. Biofilm Study Using XTT Assay
2.6. Confocal Laser Scanning Microscopy
3. Results and Discussion
3.1. Susceptibility of Isolated Bacteria
3.2. Scanning Electron Microscopy (SEM)
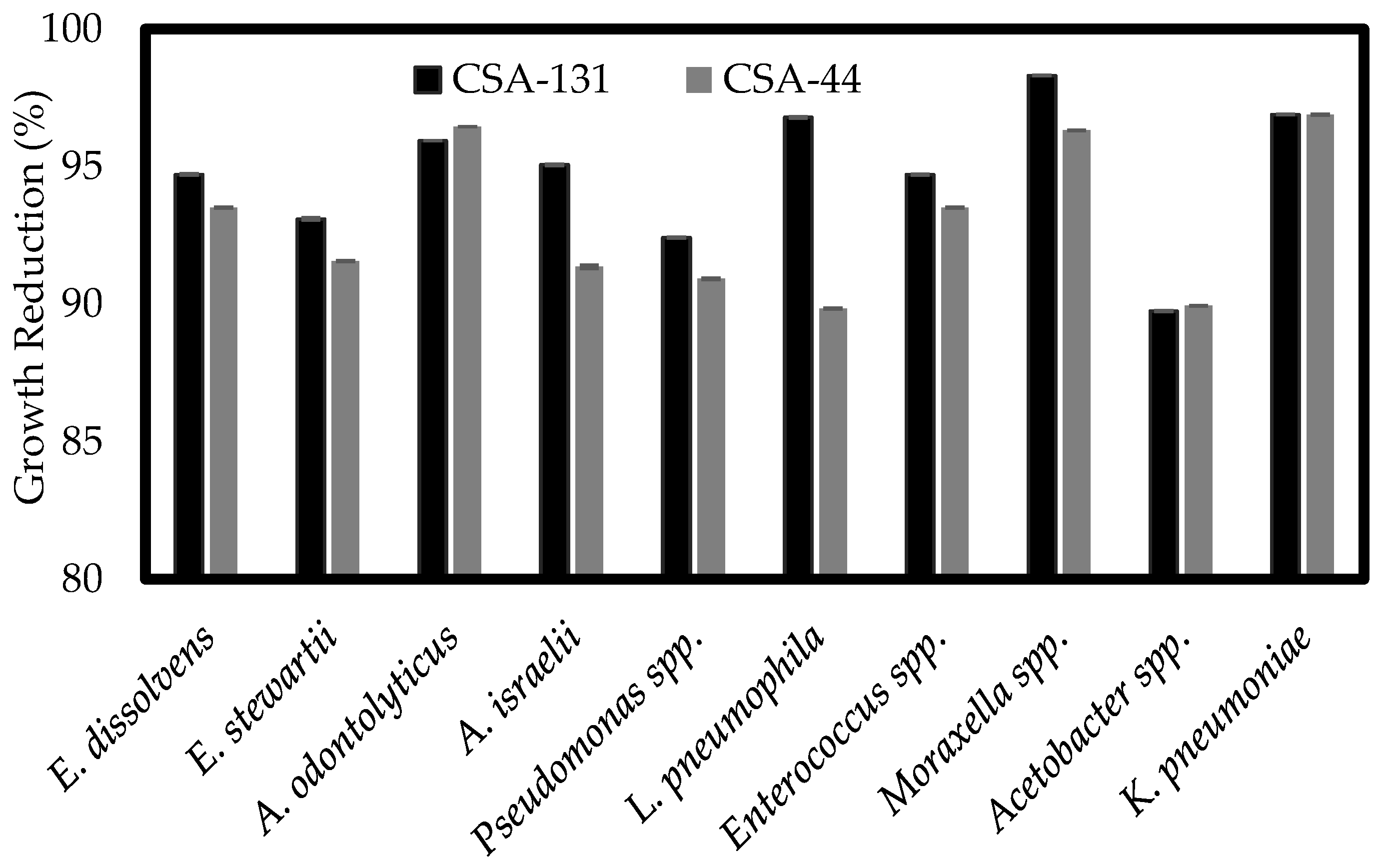
3.3. Determination of Susceptibility Profiles of Bacterial Biofilms
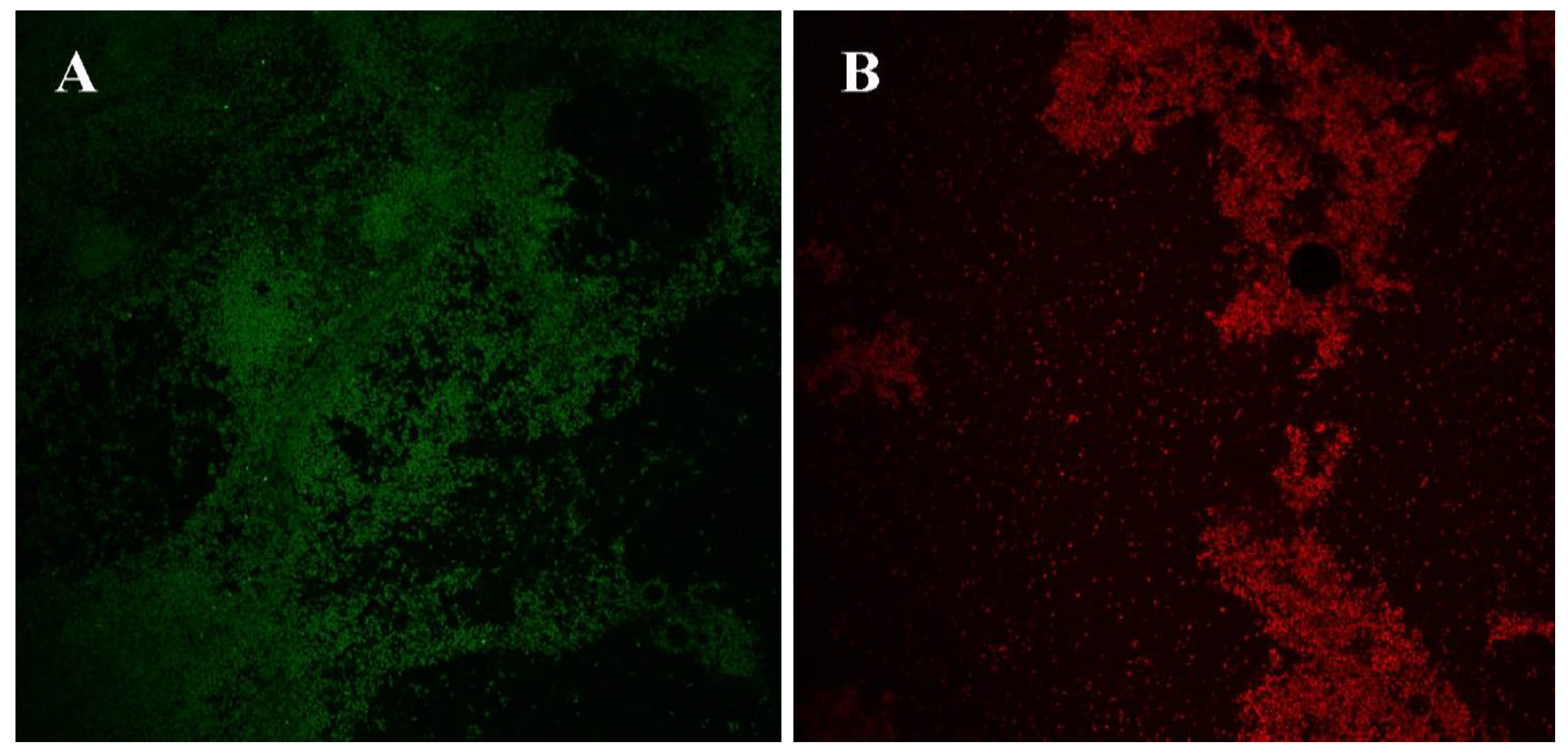
3.4. Confocal Laser Scanning of Biofilms
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Padiyara, P.; Inoue, H.; Sprenger, M. Global governance mechanisms to address antimicrobial resistance. Infect. Dis. 2018, 11, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Aoki, W.; Kuroda, K.; Ueda, M. Next generation of antimicrobial peptides as molecular targeted medicines. J. Biosci. Bioeng. 2012, 114, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Simpkin, V.L.; Renwick, M.J.; Kelly, R.; Mossialos, E. Incentivising innovation in antibiotic drug discovery and development: Progress, challenges and next steps. J. Antibiot. 2017, 70, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.M.; Levy, S.B. Food animals and antimicrobials: Impacts on human health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef]
- Zhang, L.-J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Ganz, T. The role of antimicrobial peptides in innate immunity. Integr. Comp. Biol. 2003, 43, 300–304. [Google Scholar] [CrossRef]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: Biological and therapeutic considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef]
- Nagant, C.; Pitts, B.; Stewart, P.S.; Feng, Y.; Savage, P.B.; Dehaye, J.-P. Study of the effect of antimicrobial peptide mimic, CSA-13, on an established biofilm formed by Pseudomonas aeruginosa. Microbiologyopen 2013, 2, 318–325. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Rovig, J.; Weber, S.; Hilton, B.; Forouzan, M.M.; Savage, P.B. Susceptibility of colistin-resistant, Gram-negative bacteria to antimicrobial peptides and ceragenins. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef]
- Durnaś, B.; Wnorowska, U.; Pogoda, K.; Deptuła, P.; Wątek, M.; Piktel, E.; Głuszek, S.; Gu, X.; Savage, P.B.; Niemirowicz, K.; et al. Candidacidal activity of selected ceragenins and human cathelicidin LL-37 in experimental settings mimicking infection sites. PLoS ONE 2016, 11, e0157242. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Rovig, J.; Holden, B.S.; Taylor, M.F.; Weber, S.; Wilson, J.; Hilton, B.; Zaugg, A.L.; Ellis, S.W.; Yost, C.D.; et al. Ceragenins are active against drug-resistant Candida auris clinical isolates in planktonic and biofilm forms. J. Antimicrob. Chemother. 2018, 73, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.E.; Snarr, J.; Chaudhary, V.; Jennings, J.D.; Shaw, H.; Christiansen, B.; Wright, J.; Jia, W.; Bishop, R.E.; Savage, P.B. In vitro evaluation of the potential for resistance development to ceragenin CSA-13. J. Antimicrob. Chemother. 2012, 67, 2665–2672. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.M.; Holden, B.S.; Savage, P.B. Ceragenins as non-peptide mimics of endogenous antimicrobial peptides. In Fighting Antimicrobial Resistance, 1st ed.; Budimir, A., Ed.; IAPC Publishing: Zagreb, Croatia, 2018; pp. 139–169. ISBN 978-953-56942-6-7. [Google Scholar]
- Sinclair, K.; Pham, T.; Farnsworth, R.; Williams, D.L.; Loc-Carrillo, C.; Horne, L.A.; Ingebretsen, S.H.; Bloebaum, R.D. Development of a broad spectrum polymer-released antimicrobial coating for the prevention of resistant strain bacterial infections. J. Biomed. Mater. Res. A 2012, 100, 2732–2738. [Google Scholar] [CrossRef] [PubMed]
- Bucki, R.; Niemirowicz, K.; Wnorowska, U.; Byfield, F.J.; Piktel, E.; Wątek, M.; Janmey, P.A.; Savage, P.B. Bactericidal activity of ceragenin CSA-13 in cell culture and in an animal model of peritoneal infection. Antimicrob. Agents Chemother. 2015, 59, 6274–6282. [Google Scholar] [CrossRef]
- Olekson, M.; You, T.; Savage, P.B.; Leung, K.P. Ceragenin peptide-mimics inhibit biofilms and affect mammalian cell viability and migration in vitro. FEBS Open Bio 2017, 7, 953–967. [Google Scholar] [CrossRef]
- Li, C.; Peters, A.S.; Meredith, E.L.; Allman, G.W.; Savage, P.B. Design and synthesis of potent sensitizers of Gram-negative bacteria based on a cholic acid scaffolding. J. Am. Chem. Soc. 1998, 120, 2961–2962. [Google Scholar] [CrossRef]
- Cheesbrough, M. District Laboratory Practice in Tropical Countries; Cambridge University Press: Cambridge, UK, 2006; pp. 143–157. ISBN 978-0-521-67631-1. [Google Scholar]
- Cruickshank, R.; Duguid, J.O.; Marmon, B.P.; Swain, R.H.A. Medical Microbiology: The Practice of Medical Microbiology, 12th ed.; Churchill Livingstone: Edinburg, NY, USA, 1980; p. 585. ISBN 978-0443011115. [Google Scholar]
- Holt, J.G. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams & Wilkins: Baltimore, MD, USA, 2000; pp. 532–551. ISBN 978-0683006032. [Google Scholar]
- Wikler, M.A. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically: Approved Standard; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2006; ISBN 1-56238-783-9. [Google Scholar]
- Hashemi, M.M.; Holden, B.S.; Taylor, M.F.; Wilson, J.; Coburn, J.; Hilton, B.; Nance, T.; Gubler, S.; Genberg, C.; Deng, S.; et al. Antibacterial and antifungal activities of poloxamer micelles containing ceragenin CSA-131 on ciliated tissues. Molecules 2018, 23, 596. [Google Scholar] [CrossRef]
- Bucki, R.; Sostarecz, A.G.; Byfield, F.J.; Savage, P.B.; Janmey, P.A. Resistance of the antibacterial agent ceragenin CSA-13 to inactivation by DNA or F-actin and its activity in cystic fibrosis sputum. J. Antimicrob. Chemother. 2007, 60, 535–545. [Google Scholar] [CrossRef]
- Ding, B.; Guan, Q.; Walsh, J.P.; Boswell, J.S.; Winter, T.W.; Winter, E.S.; Boyd, S.S.; Li, C.; Savage, P.B. Correlation of the antibacterial activities of cationic peptide antibiotics and cationic ateroid antibiotics. J. Med. Chem. 2002, 45, 663–669. [Google Scholar] [CrossRef]
- Stewart, P.S. Antimicrobial tolerance in biofilms. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Sandasi, M.; Leonard, C.; Viljoen, A. The in vitro antibiofilm activity of selected culinary herbs and medicinal plants against Listeria monocytogenes. Lett. Appl. Microbiol. 2010, 50, 30–35. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Holden, B.S.; Durnaś, B.; Bucki, R.; Savage, P.B. Ceragenins as mimics of endogenous antimicrobial peptides. J. Antimicrob. Agents 2017, 3, 141. [Google Scholar] [CrossRef]
- Pollard, J.; Wright, J.; Feng, Y.; Geng, D.; Genberg, C.; Savage, P.B. Activities of ceragenin CSA-13 against established biofilms in an in vitro model of catheter decolonization. Antiinfect. Agents Med. Chem. 2009, 8, 290–294. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Rovig, J.; Bateman, J.; Holden, B.S.; Modelzelewski, T.; Gueorguieva, I.; von Dyck, M.; Bracken, R.; Genberg, C.; Deng, S.; et al. Preclinical testing of a broad-spectrum antimicrobial endotracheal tube coated with an innate immune synthetic mimic. J. Antimicrob. Chemother. 2018, 73, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Leszczyńska, K.; Namiot, D.; Byfield, F.J.; Cruz, K.; Zendzian-Piotrowska, M.; Fein, D.E.; Savage, P.B.; Diamond, S.; McCulloch, C.A.; Janmey, P.A. Antibacterial activity of the human host defence peptide LL-37 and selected synthetic cationic lipids against bacteria associated with oral and upper respiratory tract infections. J. Antimicrob. Chemother. 2013, 68, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.-Z.; Feng, Y.; Pollard, J.; Chin, J.N.; Rybak, M.J.; Bucki, R.; Epand, R.F.; Epand, R.M.; Savage, P.B. Ceragenins: Cholic acid-based mimics of antimicrobial peptides. Acc. Chem. Res. 2008, 41, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Oloso, N.O.; Fagbo, S.; Garbati, M.; Olonitola, S.O.; Awosanya, E.J.; Aworh, M.K.; Adamu, H.; Odetokun, I.A.; Fasina, F.O. Antimicrobial resistance in food animals and the environment in Nigeria: A review. Int. J. Environ. Res. Public health 2018, 15, 1284. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Isolation Source | |
|---|---|---|
| 1 | Enterococcus spp. | Rubber plant |
| 2 | Actinomyces odontolyticus | Rubber plant |
| 3 | Actinomyces israelii | Rubber plant |
| 4 | Acetobacter spp. | Rubber plant |
| 5 | Moraxella spp. | Rubber plant |
| 6 | Enterobacter dissolvens | Water |
| 7 | Pseudomonas spp. | Rubber plant |
| 8 | Klebsiella pneumoniae | Water |
| 9 | Legionella pneumophila | Water |
| 10 | Erwinia Stewartii | Water |
| Strains | Chl | Kan | Col | Pol B | Ery | Tet | Van | Amp |
|---|---|---|---|---|---|---|---|---|
| Enterobacter dissolvens | 8 | 64 | >100 | 100 | 2 | 2 | nm | nm |
| Erwinia stewartii | 8 | 2 | 100 | 100 | 1 | 2 | nm | nm |
| Enterococcus spp. | 64 | 100 | >100 | 100 | 8 | 32 | nm | nm |
| Pseudomonas spp. | 32 | 16 | >100 | >100 | 16 | 8 | nm | nm |
| Klebsiella pneumoniae | 32 | 32 | >100 | 100 | 16 | 1 | nm | nm |
| Acetobacter spp. | 32 | 32 | 16 | 8 | 32 | 16 | nm | nm |
| Moraxella spp. | 64 | 64 | >100 | 100 | 32 | 32 | nm | nm |
| Legionella pneumophila | 64 | 64 | 32 | 16 | 8 | 4 | nm | nm |
| Actinomyces odontolyticus | 4 | nm | nm | nm | nm | nm | 1 | 1 |
| Actinomyces israelii | 4 | nm | nm | nm | nm | nm | 2 | 2 |
| Strains | CSA-13 | CSA-44 | CSA-131 | CSA-144 |
|---|---|---|---|---|
| Enterobacter dissolvens | 1(8) | 2(10) | 2(8) | 2(10) |
| Erwinia stewartii | 2(8) | 4(10) | 2(8) | 4(10) |
| Actinomyces odontolyticus | 1(1–2) | 1(1) | 1(1) | 2(2) |
| Actinomyces israelii | 2(4) | 1(2) | 2(4) | 2(4) |
| Pseudomonas spp. | 1(1) | 4(4) | 1(1) | 4(4) |
| Legionella pneumophila | 8(8) | 4(8) | 4(4) | 16(32) |
| Enterococcus spp. | 16(32) | 8(8) | 4(32) | 32(100) |
| Moraxella spp. | 10(32) | 4(16) | 4(16) | 24(100) |
| Acetobacter spp. | 2(32) | 4(64) | 2(32) | 4(64) |
| Klebsiella pneumoniae | 2(4) | 4(8) | 2(4) | 4(4) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashemi, M.M.; Mmuoegbulam, A.O.; Holden, B.S.; Coburn, J.; Wilson, J.; Taylor, M.F.; Reiley, J.; Baradaran, D.; Stenquist, T.; Deng, S.; et al. Susceptibility of Multidrug-Resistant Bacteria, Isolated from Water and Plants in Nigeria, to Ceragenins. Int. J. Environ. Res. Public Health 2018, 15, 2758. https://doi.org/10.3390/ijerph15122758
Hashemi MM, Mmuoegbulam AO, Holden BS, Coburn J, Wilson J, Taylor MF, Reiley J, Baradaran D, Stenquist T, Deng S, et al. Susceptibility of Multidrug-Resistant Bacteria, Isolated from Water and Plants in Nigeria, to Ceragenins. International Journal of Environmental Research and Public Health. 2018; 15(12):2758. https://doi.org/10.3390/ijerph15122758
Chicago/Turabian StyleHashemi, Marjan M., Augusta O. Mmuoegbulam, Brett S. Holden, Jordan Coburn, John Wilson, Maddison F. Taylor, Joseph Reiley, Darius Baradaran, Tania Stenquist, Shenglou Deng, and et al. 2018. "Susceptibility of Multidrug-Resistant Bacteria, Isolated from Water and Plants in Nigeria, to Ceragenins" International Journal of Environmental Research and Public Health 15, no. 12: 2758. https://doi.org/10.3390/ijerph15122758
APA StyleHashemi, M. M., Mmuoegbulam, A. O., Holden, B. S., Coburn, J., Wilson, J., Taylor, M. F., Reiley, J., Baradaran, D., Stenquist, T., Deng, S., & Savage, P. B. (2018). Susceptibility of Multidrug-Resistant Bacteria, Isolated from Water and Plants in Nigeria, to Ceragenins. International Journal of Environmental Research and Public Health, 15(12), 2758. https://doi.org/10.3390/ijerph15122758