Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals

 ,
, 

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Moisture and Ash
2.2. Proteins
2.3. Carbohydrates
2.4. Lipids
2.5. Total Dietary Fiber
2.6. Carotenoids
3. Results and Discussion
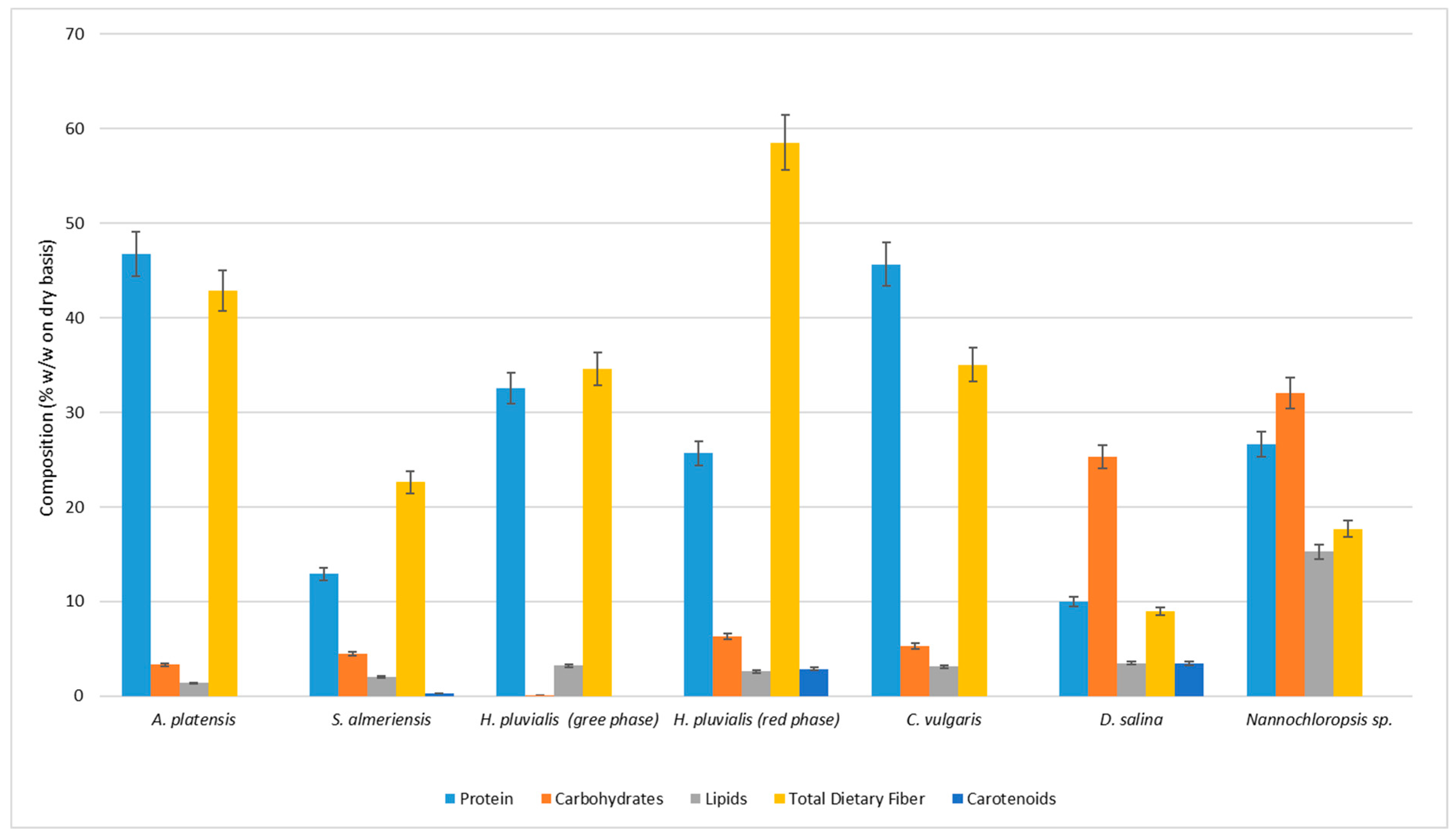
3.1. Microalgae Composition
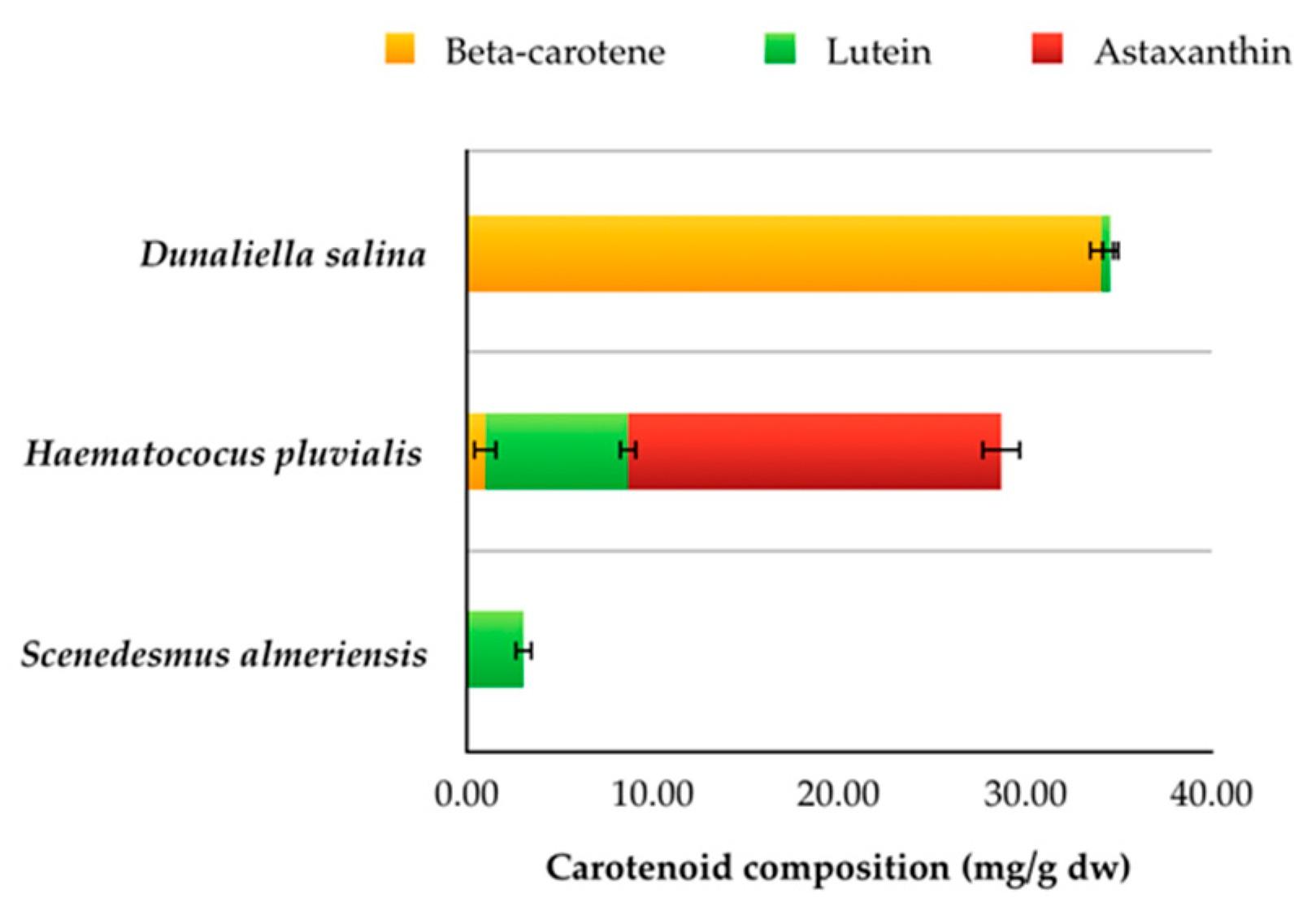
3.2. Carotenoids
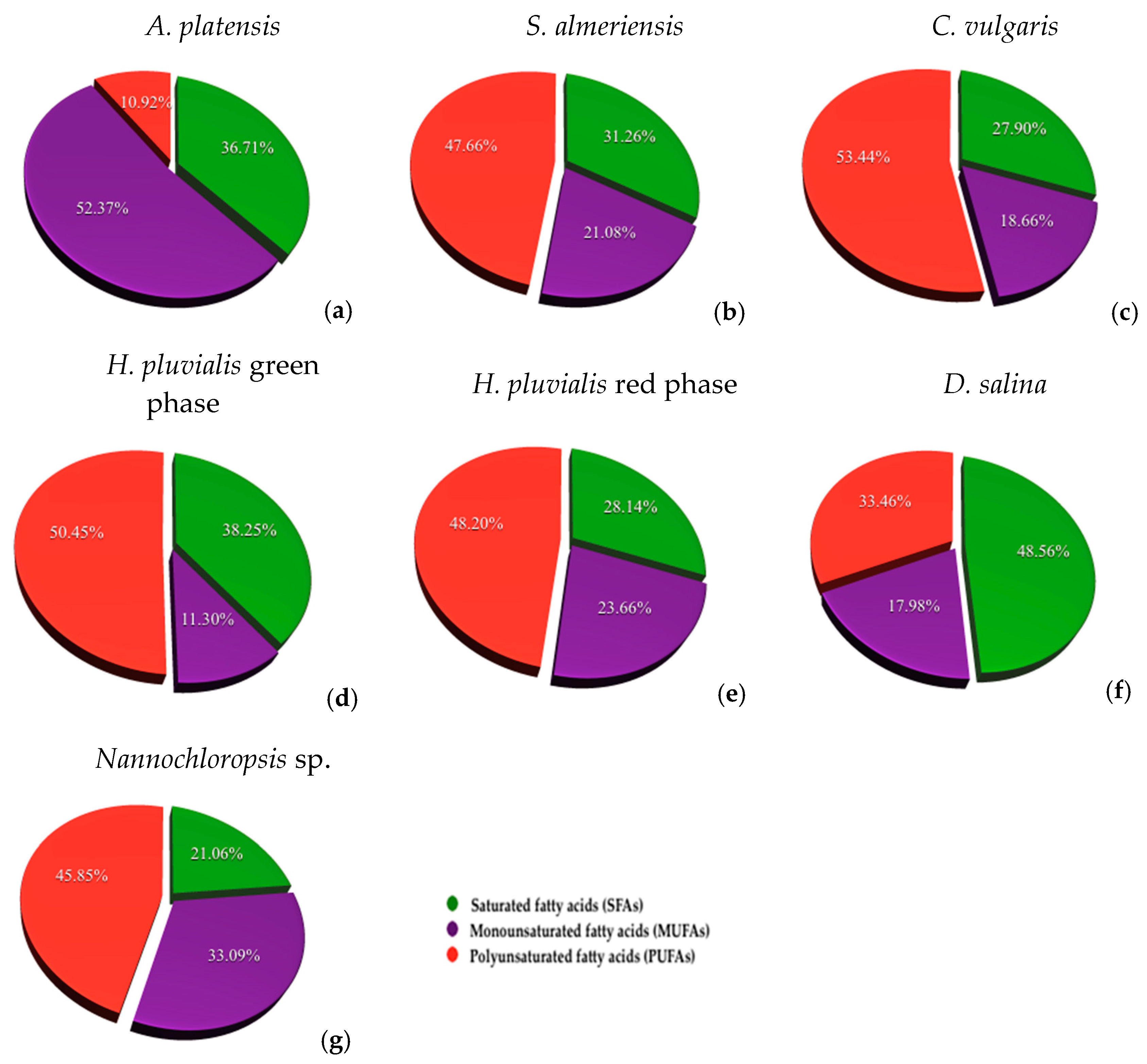
3.3. Fatty Acid Composition
3.4. Comparison with Literature About Microalgae Composition
3.5. Microalgae Market and New Challenges
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- United States Department of State, Economics and Social Affairs. The Impact of Population Momentum on Future Population Growth. Available online: https://esa.un.org/unpd/wpp/publications/Files/PopFacts_2017-4_Population-Momentum.pdf (accessed on 11 August 2018).
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Hamed, I. The Evolution and Versatility of Microalgal Biotechnology: A Review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1104–1123. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef]
- Matos, Â.P.; Feller, R.; Moecke, E.H.S.; de Oliveira, J.V.; Junior, A.F.; Derner, R.B.; Sant’Anna, E.S. Chemical Characterization of Six Microalgae with Potential Utility for Food Application. J AOCS J. Am. Oil Chem. Soc. 2016, 93, 963–972. [Google Scholar] [CrossRef]
- Muhaemin, M.; Kaswadji, R.F. Biomass Nutrient Profiles of Marine Microalgae Dunaliella salina. J. Penelit. Sains 2009, 13, 64–67. [Google Scholar]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Affan, M.A.; Jang, J.; Kang, M.H.; Ko, A.R.; Jeon, S.M.; Oh, C.; Heo, S.J.; Lee, Y.H.; Ju, S.J.; et al. Morphological, molecular, and biochemical characterization of astaxanthin-producing green microalga Haematococcus sp. KORDI03 (haematococcaceae, chlorophyta) isolated from Korea. J. Microbiol. Biotechnol. 2015, 25, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.F.; Fernández, J.M.; Acién, F.G.; Rueda, A.; Pérez-Parra, J.; Molina, E. Influence of culture conditions on the productivity and lutein content of the new strain Scenedesmus almeriensis. Process Biochem. 2008, 43, 398–405. [Google Scholar] [CrossRef]
- Food and Drug Administration, USA. Substances Generally Recognized as Safe. Available online: https://www.federalregister.gov/d/2016-19164/p-1 (accessed on 11 August 2018).
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef] [PubMed]
- García, J.L.; de Vicente, M.; Galán, B. Microalgae, old sustainable food and fashion nutraceuticals. Microb. Biotechnol. 2017, 10, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Caporgno, M.P.; Mathys, A. Trends in Microalgae Incorporation into Innovative Food Products with Potential Health Benefits. Front. Nutr. 2018, 5, 58. [Google Scholar] [CrossRef] [PubMed]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef]
- Stanic-Vucinic, D.; Minic, S.; Nikolic, M.R.; Velickovic, T.C. Spirulina Phycobiliproteins as Food Components and Complements. In Microalgal Biotechnology, 1st ed.; Jacob-Lopes, E., Queiroz Zepka, L., Queiroz, M.I., Eds.; IntechOpen: London, UK, 2018; pp. 129–149. [Google Scholar]
- Harrison, E.H. Mechanisms involved in the intestinal absorption of dietary vitamin A and provitamin A carotenoids. Biochim. Biophys. Acta 2012, 1821, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Yaakob, Z.; Ali, E.; Zainal, A.; Mohamad, M.; Takriff, M.S. An overview: Biomolecules from microalgae for animal feed and aquaculture. J. Biol. Res. 2014, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Hemaiswarya, S.; Raja, R.; Kumar, R.R.; Ganesan, V.; Anbazhagan, C. Microalgae: A sustainable feed source for aquaculture. World J. Microbiol. Biotechnol. 2011, 27, 1737–1746. [Google Scholar] [CrossRef]
- Sørensen, M.; Gong, Y.; Bjarnason, F.; Vasanth, G.K.; Dahle, D.; Huntley, M.; Kiron, V. Nannochloropsis oceania-derived defatted meal as an alternative to fishmeal in Atlantic salmon feeds. PLoS ONE 2017, 12, e0179907. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.Y.; Deng, D.F.; Dominy, W. A defatted microalgae (Haematococcus pluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931). Aquaculture 2012, 354–355, 50–55. [Google Scholar] [CrossRef]
- Kiron, V.; Phromkunthong, W.; Huntley, M.; Archibald, I.; De Scheemaker, G. Marine microalgae from biorefinery as a potential feed protein source for Atlantic salmon, common carp and whiteleg shrimp. Aquac. Nutr. 2012, 18, 521–531. [Google Scholar] [CrossRef]
- Nicoletti, M. Microalgae Nutraceuticals. Foods 2016, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Bishop, M.W.; Zubeck, H.M. Evaluation of Microalgae for use as Nutraceuticals and Nutritional Supplements. J. Nutr. Food Sci. 2012, 2, 147. [Google Scholar] [CrossRef]
- Guillerme, J.-B.; Couteau, C.; Coiffard, L. Applications for Marine Resources in Cosmetics. Cosmetics 2017, 4, 35. [Google Scholar] [CrossRef]
- Enzing, C.; Ploeg, M.; Barbosa, M.; Sijtsma, L. Microalgae-based products for the food and feed sector: An outlook for Europe. JRC Sci. Policy Rep. 2014, 82, 19–37. [Google Scholar]
- Meticulous Market Research Algae Products Market Worth 3318.1 Million USD by 2022. 2018. Available online: https://www.meticulousresearch.com/algae-products-market-size-2017-2022/ (accessed on 11 August 2018).
- Carotenoids Market Analysis by Source (Natural, Synthetic), By Product (Beta-Carotene, Lutein, Lycopene, Astaxanthin, Zeaxanthin, Canthaxanthin), by Application (Food, Supplements, Feed, Pharmaceuticals, Cosmetics), and Segment Forecasts, 2014–2025. 2016. Available online: https://www.grandviewresearch.com/industry-analysis/carotenoids-market (accessed on 16 August 2018).
- Grand View Research Omega 3 Market Size, Share & Trend Analysis Report by Application (Supplements & Functional Foods, Pharmaceuticals, Infant formulas, Pet & Animal Feed), by Region, and Segment Forecasts, 2012–2022. 2018. Available online: https://www.grandviewresearch.com/industry-analysis/omega-3-market (accessed on 17 August 2018).
- Business Wire Global Algae Protein Market 2017–2023—$800 + Million Industry Trends, Opportunities and Forecasts-ResearchAndMarkets.com. 2018. Available online: https://www.businesswire.com/news/home/20180703005135/en/Global-Algae-Products-Market-Analysis-Trends-Industry (accessed on 17 August 2018).
- Brenner Pharma Food Algal Plant Proteins (MicroAlgae) a Disruptive Opportunity Market Trends, Technologies, Key Success Factors. 2017. Available online: https://www.globalfoodforums.com/wp-content/uploads/2017/05/2017-PTT_Bus-G.Brenner-Brenner.pdf (accessed on 17 August 2018).
- De Jesus Raposo, M.F.; de Morais, R.M.S.C.; de Morais, A.M.M.B. Health applications of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Patias, L.D.; Maroneze, M.M.; Siqueira, S.F.; de Menezes, C.R.; Zepka, L.Q.; Jacob-Lopes, E. Single-Cell Protein as a Source of Biologically Active Ingredients for the Formulation of Antiobesity Foods. Altern. Replace. Foods 2018, 317–353. [Google Scholar]
- UNI EN ISO 712:2011. Cereals and Cereals Products—Determination of Moisture Content—Reference Method. Available online: https://www.iso.org/standard/44807.html (accessed on 12 August 2018).
- UNI EN ISO 2171:2011. Cereals, Pulse and by-Products—Determination of Ash Yield by Incineration. Available online: https://www.iso.org/standard/37264.html (accessed on 12 August 2018).
- UNI EN ISO 20483:2014. Cereals and Pulse—Determination of Nitrogen Content and Calculation of the Crude Protein—Kjeldhal Method. Available online: https://www.iso.org/standard/59162.html (accessed on 12 August 2018).
- Myklestad, S.; Haug, A. Production of carbohydrates by the marine diatom Chaetoceros affinis var. willei (Gran) Hustedt. I. Effect of the concentration of nutrients in the culture medium. J. Exp. Mar. Biol. Ecol. 1972, 9, 125–136. [Google Scholar] [CrossRef]
- UNI EN 15086:2006. Foodstuffs—Determination of Isomalt, Lactitol, Mannitol, Sorbitol, and Xylitol in Foodstuffs. Available online: http://store.uni.com/catalogo/index.php/uni-en-iso-5943-2007.html?_store=en&_from_store=it (accessed on 13 August 2018).
- Tang, Y.; Zhang, Y.; Rosenberg, J.; Betenbaugh, M.; Wang, F. Optimization of One-Step in Situ Transesterification Method for Accurate Quantification of EPA in Nannochloropsis gaditana. Appl. Sci. 2016, 6, 343. [Google Scholar] [CrossRef]
- UNI ISO 12966-2:2011. Animal and Vegetables Fat and Oils—Gas Chromatography of Fatty Acid Methyl Esters—Part 2: Preparation of Methyl Esters of Fatty Acids. Available online: https://www.iso.org/standard/43172.html (accessed on 13 August 2018).
- UNI ISO 12966-4:2011. Animal and Vegetables Fat and Oils—Gas Chromatography of Fatty Acid Methyl Esters—Part 4: Determination by Capillary Chromatography. Available online: https://www.iso.org/standard/63503.html (accessed on 13 August 2018).
- Association Official Agricultural Chemists (AOAC). AOAC Official Methods of Analysis; Association Official Agricultural Chemists: Gaithersburg, MO, USA, 1998; pp. 136–138. [Google Scholar]
- Li, Y.; Miao, F.; Geng, Y.; Lu, D.; Zhang, C.; Zeng, M. Accurate quantification of astaxanthin from Haematococcus crude extract spectrophotometrically. Chin. J. Oceanol. Limnol. 2012, 30, 627–637. [Google Scholar] [CrossRef]
- Sanzo, G.; Mehariya, S.; Martino, M.; Larocca, V.; Casella, P.; Chianese, S.; Musmarra, D.; Balducchi, R.; Molino, A.; Sanzo, G.D.; et al. Supercritical Carbon Dioxide Extraction of Astaxanthin, Lutein, and Fatty Acids from Haematococcus pluvialis Microalgae. Mar. Drugs 2018, 16, E334. [Google Scholar] [CrossRef] [PubMed]
- UNI EN 12823-2:2000. Foodstuffs Determination of Vitamin a by High Performance Liquid Chromatography Measurement of β-carotene. Available online: http://store.uni.com/catalogo/index.php/uni-en-12823-2-2000.html?_store=it&_from_store=en (accessed on 13 August 2018).
- Ruen-ngam, D.; Shotipruk, A.; Pavasant, P. Comparison of extraction methods for recovery of astaxanthin from Haematococcus pluvialis. Sep. Sci. Technol. 2011, 46, 64–70. [Google Scholar] [CrossRef]
- López-González, D.; Fernandez-Lopez, M.; Valverde, J.L.L.; Sanchez-Silva, L. Kinetic analysis and thermal characterization of the microalgae combustion process by thermal analysis coupled to mass spectrometry. Appl. Energy 2014, 114, 227–237. [Google Scholar] [CrossRef]
- Carrillo-Reyes, J.; Barragán-Trinidad, M.; Buitrón, G. Biological pretreatments of microalgal biomass for gaseous biofuel production and the potential use of rumen microorganisms: A review. Algal Res. 2016, 18, 341–351. [Google Scholar] [CrossRef]
- García-González, M.; Moreno, J.; Manzano, J.C.; Florencio, F.J.; Guerrero, M.G. Production of Dunaliella salina biomass rich in 9-cis-β-carotene and lutein in a closed tubular photobioreactor. J. Biotechnol. 2005, 115, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tang, X.; Kapoore, R.V.; Xu, C.; Vaidyanathan, S. Influence of nutrient status on the accumulation of biomass and lipid in Nannochloropsis salina and Dunaliella salina. Energy Convers. Manag. 2015, 106, 62–72. [Google Scholar] [CrossRef]
- Chidambara Murthy, K.N.; Vanitha, A.; Rajesha, J.; Mahadeva Swamy, M.; Sowmya, P.R.; Ravishankar, G. In vivo antioxidant activity of carotenoids from Dunaliella salina—A green microalga. Life Sci. 2005, 76, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Lamers, P.P.; Van De Laak, C.C.W.; Kaasenbrood, P.S.; Lorier, J.; Janssen, M.; De Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Carotenoid and fatty acid metabolism in light-stressed Dunaliella salina. Biotechnol. Bioeng. 2010, 106, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Hata, N.; Ogbonna, J.C.; Hasegawa, Y.; Taroda, H.; Tanaka, H. Production of astaxanthin by Haematococcus pluvialis in a sequential heterotrophic-photoautotrophic culture. J. Appl. Phycol. 2001, 13, 395–402. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sevilla, J.M.; Acién Fernández, F.G.; Molina Grima, E. Biotechnological production of lutein and its applications. Appl. Microbiol. Biotechnol. 2010, 86, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.A.C.L.D.; Monteiro, M.P.C.; Robbs, P.G.; Leite, S.G.F. Growth and Chemical Composition of Spirulina Maxima and Spirulina Platensis Biomass at Different Temperatures. Aquac. Int. 1999, 7, 261–275. [Google Scholar] [CrossRef]
- Choi, G.-G.; Bae, M.-S.; Ahn, C.-Y.; Oh, H.-M. Enhanced biomass and gamma-linolenic acid production of mutant strain Arthrospira platensis. J. Microbiol. Biotechnol. 2008, 18, 539–544. [Google Scholar] [PubMed]
- Golmakani, M.-T.; Rezaei, K.; Mazidi, S.; Razavi, S.H. γ-Linolenic acid production by Arthrospira platensis using different carbon sources. Eur. J. Lipid Sci. Technol. 2012, 114, 306–314. [Google Scholar] [CrossRef]
- Sánchez, J.F.; Fernández-Sevilla, J.M.; Acién, F.G.; Cerón, M.C.; Pérez-Parra, J.; Molina-Grima, E. Biomass and lutein productivity of Scenedesmus almeriensis: Influence of irradiance, dilution rate and temperature. Appl. Microbiol. Biotechnol. 2008, 79, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Davidi, L.; Katz, A.; Pick, U. Characterization of major lipid droplet proteins from Dunaliella. Planta 2012, 236, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, H.; Martel, A.; Jiménez Del Río, M.; García Reina, G. Oleic acid is the main fatty acid related with carotenogenesis in Dunaliella salina. J. Appl. Phycol. 1999, 11, 15–19. [Google Scholar] [CrossRef]
- Van Wagenen, J.; Miller, T.W.; Hobbs, S.; Hook, P.; Crowe, B.; Huesemann, M. Effects of Light and Temperature on Fatty Acid Production in Nannochloropsis Salina. Energies 2012, 5, 731–740. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, L.; Wang, J.; Chen, Y.; Gao, X.; Zhang, Z.; Liu, T. Attached cultivation for improving the biomass productivity of Spirulina platensis. Bioresour. Technol. 2015, 181, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Gershwin, M.E.; Belay, A.; Belay, M.E.G.A. Spirulina in Human Nutrition and Health; CRC Press: Boca Raton, FL, USA, 2007; p. 328. [Google Scholar]
- Depraetere, O.; Pierre, G.; Noppe, W.; Vandamme, D.; Foubert, I.; Michaud, P.; Muylaert, K. Influence of culture medium recycling on the performance of Arthrospira platensis cultures. Algal Res. 2015, 10, 48–54. [Google Scholar] [CrossRef]
- D’Alessandro, E.B.; Antoniosi Filho, N.R. Concepts and studies on lipid and pigments of microalgae: A review. Renew. Sustain. Energy Rev. 2016, 58, 832–841. [Google Scholar] [CrossRef]
- Ho, S.H.; Huang, S.W.; Chen, C.Y.; Hasunuma, T.; Kondo, A.; Chang, J.S. Characterization and optimization of carbohydrate production from an indigenous microalga Chlorella vulgaris FSP-E. Bioresour. Technol. 2013, 135, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Batista, A.P.; Gouveia, L.; Bandarra, N.M.; Franco, J.M.; Raymundo, A. Comparison of microalgal biomass profiles as novel functional ingredient for food products. Algal Res. 2013, 2, 164–173. [Google Scholar] [CrossRef]
- Yusof, Y.A.M. Fatty acids composition of microalgae Chlorella vulgaris can be modulated by varying carbon dioxide concentration in outdoor culture. Afr. J. Biotechnol. 2011, 10, 13536–13542. [Google Scholar]
- Mitra, D.; Hans van Leeuwen, J.; Lamsal, B. Heterotrophic/mixotrophic cultivation of oleaginous Chlorella vulgaris on industrial co-products. Algal Res. 2012, 1, 40–48. [Google Scholar] [CrossRef]
- Bonnefond, H.; Moelants, N.; Talec, A.; Bernard, O.; Sciandra, A. Concomitant effects of light and temperature diel variations on the growth rate and lipid production of Dunaliella salina. Algal Res. 2016, 14, 72–78. [Google Scholar] [CrossRef]
- Rebolloso-Fuentes, M.M.; Navarro-Pérez, A.; García-Camacho, F.; Ramos-Miras, J.J.; Guil-Guerrero, J.L. Biomass Nutrient Profiles of the Microalga Nannochloropsis. J. Agric. Food Chem. 2001, 49, 2966–2972. [Google Scholar] [CrossRef] [PubMed]
- Wahidin, S.; Idris, A.; Shaleh, S.R.M. The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Alboresi, A.; Perin, G.; Vitulo, N.; Diretto, G.; Block, M.A.; Jouhet, J.; Meneghesso, A.; Valle, G.; Giuliano, G.; Maréchal, E.; et al. Light Remodels Lipid Biosynthesis in Nannochloropsis gaditana by Modulating Carbon Partitioning between Organelles. Plant Physiol. 2016, 171, 2468–2482. [Google Scholar] [CrossRef] [PubMed]
- Perin, G.; Simionato, D.; Bellan, A.; Carone, M.; Occhipinti, A.; Maffei, M.E.; Morosinotto, T. Cultivation in industrially relevant conditions has a strong influence on biological properties and performances of Nannochloropsis gaditana genetically modified strains. Algal Res. 2017, 28, 88–99. [Google Scholar] [CrossRef]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Aussant, J.; Guihéneuf, F.; Stengel, D.B. Impact of temperature on fatty acid composition and nutritional value in eight species of microalgae. Appl. Microbiol. Biotechnol. 2018, 12, 5279–5297. [Google Scholar] [CrossRef] [PubMed]
- Bienvenido, O.J. Nutrient composition and protein quality of rice relative to other cereals. In Human Nutrition; The Collaboration of the International Rice Research Institute Food and Agriculture Organization of the United Nations: Rome, Italy, 1993; Available online: http://www.fao.org/docrep/x2184e/x2184e04.htm (accessed on 16 October 2018).
- Norman, F.H.; Odunfa, S.A.; Cherl-Ho, L.; Quintero-Ramírez, R.; Lorence-Quiñones, A.; Wacher-Radarte, C. Nutrient Composition and Protein Quality of Rice Relative to Other Cereals. In Fermented Cereals; A Global Perspective. FAO Agricultural Services Bulletin; No. 138.1999; ISBN 92-5-104296-9. Available online: http://www.fao.org/docrep/t0567e/T0567E0d.htm (accessed on 17 October 2018).
- Frolich, W.; Asp, N.G. Dietary fiber content in cereals in Norway. Cereal Chem. 1981, 58, 524–527. [Google Scholar]
- AgroCycle. Characterisation of Agricultural Waste Co- and by-Products. 2016. Available online: http://www.agrocycle.eu/files/2017/10/D1.2_AgroCycle.pdf (accessed on 17 October 2018).
- Piccolo, A. Spirulina—A Livelihood and a Business Venture. REPORT/RAPPORT: SF/2011/16. 2012. Available online: http://www.fao.org/3/a-az386e.pdf (accessed on 17 October 2018).
- Voort, M.P.J.; van der Vulsteke, E.; de Visser, C.L.M. Marco-Economics of Algae Products. Output Report WP2A7.02 of the EnAlgae Project, Swansea. June 2015. Available online: http://www.enalgae.eu/public-deliverables.htm (accessed on 17 October 2018).
- Li, J.; Daling, Z.; Jianfeng, N.; Shen, S.; Wang, G. An economic assessment of astaxanthin production by large scale cultivation of Haematococcus pluvialis. Biotechnol. Adv. 2011, 29, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Microalgae for aquaculture: Opportunities and constraints. J. Appl. Psychol. 1997, 9, 393–401. [Google Scholar]
- Draaisma, R.B.; Wijffels, R.H.; Ellen Slegers, P.M.; Brentner, L.B.; Roy, A.; Barbosa, M.J. Food commodities from microalgae. Curr. Opin. Biotechnol. 2013, 24, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialization. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.M. Chlorella and Spirulina Microalgae as Sources of Functional Foods, Nutraceuticals, and Food Supplements; an Overview. MOJ Food Process. Technol. 2018, 6, 45–58. [Google Scholar] [CrossRef]
- Habib, M.A.B.; Parvin, M.; Huntington, T.C.; Hasan, M.R. A Review of Culture, Production and Use of Spirulina as Food for Humans and Feeds for Domestic Animals and Fish; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008; p. 33. ISBN 978-92-5-106106-0. [Google Scholar]
- Molino, A.; Rimauro, J.; Casella, P.; Cerbone, A.; Larocca, V.; Karatza, D.; Hristoforou, E.; Chianese, S.; Musmarra, D. Microalgae valorisation via accelerated solvent extraction: Optimization of the operative conditions. Chem. Eng. Trans. 2018, 65, 835–840. [Google Scholar]
- Molino, A.; Rimauro, J.; Casella, P.; Cerbone, A.; Larocca, V.; Chianese, S.; Karatza, D.; Mehariya, S.; Ferraro, A.; Hristoforou, E.; et al. Extraction of astaxanthin from microalga Haematococcus pluvialis in red phase by using generally recognized as safe solvents and accelerated extraction. J. Biotechnol. 2018, 283, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Reyes, F.A.; Mendiola, J.A.; Ibañez, E.; Del Valle, J.M. Astaxanthin extraction from Haematococcus pluvialis using CO2-expanded ethanol. J. Supercrit. Fluids 2014, 92, 75–83. [Google Scholar] [CrossRef]
- Lin, J.H.; Lee, D.J.; Chang, J.S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Landrum, J.T.; Bone, R. Lutein, zeaxanthin, and the macular pigment. Arch. Biochem. Biophys. 2001, 385, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.N.; Chen, T.P.; Yang, B.; Liu, J.; Chen, F. Lipid production from Nannochloropsis. Mar. Drugs 2016, 14, E61. [Google Scholar] [CrossRef] [PubMed]
- Babuskin, S.; Krishnan, K.R.; Babu, P.A.S.; Sivarajan, M.; Sukumar, M. Functional foods enriched with marine microalga Nannochloropsis oculata as a source of ω-3 fatty acids. Food Technol. Biotechnol. 2014, 52, 292–299. [Google Scholar]
- Rodríguez De Marco, E.; Steffolani, M.E.; Martínez, M.; León, A.E. The use of Nannochloropsis sp. as a source of omega-3 fatty acids in dry pasta: Chemical, technological and sensory evaluation. Int. J. Food Sci. Technol. 2018, 53, 499–507. [Google Scholar] [CrossRef]
- Gbadamosi, O.K.; Lupatsch, I. Effects of dietary Nannochloropsis salina on the nutritional performance and fatty acid profile of Nile tilapia, Oreochromis niloticus. Algal Res. 2018, 33, 48–54. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; López, G.; Sáez, M.I.; Jiménez, J.A.; Barros, A.; Hidalgo, L.; Camacho-Rodríguez, J.; Martínez, T.F.; Cerón-García, M.C.; Alarcón, F.J. Effects of the microalga Scenedesmus almeriensis as fishmeal alternative in diets for gilthead sea bream, Sparus aurata, juveniles. Aquaculture 2014, 431, 34–43. [Google Scholar] [CrossRef]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Safi, C.; Ursu, A.V.; Laroche, C.; Zebib, B.; Merah, O.; Pontalier, P.Y.; Vaca-Garcia, C. Aqueous extraction of proteins from microalgae: Effect of different cell disruption methods. Algal Res. 2014, 3, 61–65. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Pignolet, O.; Silvestre, F.; Vaca-Garcia, C.; Pontalier, P.Y. Influence of microalgae cell wall characteristics on protein extractability and determination of nitrogen-to-protein conversion factors. J. Appl. Phycol. 2013, 25, 523–529. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Composition | Arthrospira platensis | Scenedesmus almeriensis | Haematococcus pluvialis (Green Phase) | Haematococcus pluvialis (Red Phase) | Chlorella vulgaris | Dunaliella salina | Nannochloropsis sp. |
|---|---|---|---|---|---|---|---|
| Moisture * | 6.45 ± 0.15 | 8.89 ± 0.32 | 5.03 ± 0.12 | 2.79 ± 0.23 | 1.92 ± 0.09 | 6.63 ± 0.25 | 1.90 ± 0.05 |
| Ash # | 5.71 ± 0.32 | 57.61 ± 2.20 | 29.49 ± 0.22 | 4.02 ± 0.22 | 10.88 ± 0.51 | 48.74 ± 2.50 | 8.31 ± 0.42 |
| Composition (% w/w on a Dry Basis ± SD) | Arthrospira platensis | Scenedesmus almeriensis | Haematococcus pluvialis (Green Phase) | Haematococcus pluvialis (Red Phase) | Chlorellavulgaris | Dunaliella salina | Nannochloropsis sp. |
|---|---|---|---|---|---|---|---|
| Protein | 46.76 ± 0.95 | 12.93 ± 0.69 | 32.59 ± 1.20 | 25.69 ± 1.27 | 45.64 ± 1.20 | 10.03 ± 0.57 | 26.67 ± 1.10 |
| Carbohydrates | 3.32 ± 0.05 | 4.51 ± 0.41 | 0.13 ± 0.01 | 6.30 ± 0.24 | 5.30 ± 0.50 | 25.31 ± 1.55 | 32.05 ± 0.70 |
| Lipids | 1.40 ± 0.12 | 2.05 ± 0.12 | 3.24 ± 0.11 | 2.60 ± 0.10 | 3.13 ± 0.21 | 3.49 ± 0.10 | 15.30 ± 0.24 |
| Total dietary Fiber | 42.82 ± 1.20 | 22.60± 1.50 | 34.56 ± 0.90 | 58.52 ± 2.56 | 35.04 ± 1.60 | 8.97 ± 0.50 | 17.67 ± 0.80 |
| Carotenoids | <Ldl * | 0.30 ± 0.05 | <Ldl * | 2.87 ± 0.15 | <Ldl * | 3.46 ± 0.15 | <Ldl * |
| Fatty Acid Composition (mg/100 g on a dry basis ± SD) | Artrhospira platensis | Scenedesmus almeriensis | Haematococcuspluvialis (Green Phase) | Haematococcus pluvialis (Red Phase) | Chlorella vulgaris | Dunaliella salina | Nannochloropsis sp. |
|---|---|---|---|---|---|---|---|
| SFAs * | |||||||
| Tridecanoic acid | nd # | nd # | 10.90 ± 0.22 | nd # | 11.21 ± 0.39 | nd # | 233.20 ± 0.67 |
| Palmitic acid | 253.32 ± 0.73 | 249.07 ± 0.90 | 521.73 ± 0.49 | 506.68 ± 0.27 | 598.75 ± 2.20 | 965.00 ± 1.15 | 26.43 ± 1.50 |
| Pentadecanoic acid | nd # | nd # | nd # | 3.22 ± 0.18 | nd # | nd # | 294.71 ± 0.38 |
| Heptadecanoic acid | nd # | nd # | 19.28 ± 0.42 | nd # | nd # | nd # | 1253.38 ± 2.35 |
| Stearic acid | 9.94 ± 0.24 | 33.83 ± 1.23 | 130.36 ± 1.98 | 42.07 ± 0.35 | 22.14 ± 1.39 | 567.68 ± 0.56 | nd # |
| Arachidic acid | 124.38 ± 0.95 | 66.22 ± 2.49 | nd # | 63.12 ± 1.37 | nd # | nd # | 382.51 ± 1.94 |
| ∑ other SFAs | - | - | 56.64 ± 1.49 | 31.64 ± 1.19 | 29.28 ± 0.94 | - | 16.71 ± 0.62 |
| ∑ SFAs | 387.64 ± 1.61 | 349.11 ± 4.58 | 738.91 ± 4.40 | 646.73 ± 2.74 | 661.37 ± 4.32 | 1532.68 ± 1.70 | 2316.54 ± 2.02 |
| MUFAs * | |||||||
| Palmitoleic acid | 38.47 ± 0.54 | 60.04 ± 0.90 | 51.23 ± 0.88 | 4.82 ± 0.61 | 15.69 ± 0.98 | nd # | 2588.39 ± 1.79 |
| cis-9-Octadecenoic acid (oleic acid) | 65.06 ± 0.96 | 109.11 ± 1.67 | nd # | 486.17 ± 0.23 | 226.43 ± 0.74 | 567.56 ± 1.29 | nd # |
| Myristoleic acid | 16.11 ± 0.94 | 16.99 ± 1.25 | 33.65 ± 0.68 | nd # | 23.19 ± 0.97 | nd # | 483.93 ± 0.86 |
| Nervonic acid | nd # | nd # | nd # | nd # | nd # | nd # | 522.39 ± 0.23 |
| Erucic acid | 433.46 ± 0.83 | nd # | 51.78 ± 0.70 | nd # | nd # | nd # | nd # |
| ∑ other MUFAs | - | 49.28 ± 1.13 | 81.54 ± 0.70 | 52.60 ± 0.62 | 177.04 ± 1.37 | - | 44.59 ± 0.85 |
| ∑ MUFAs | 553.10 ± 1.75 | 235.42 ± 2.80 | 218.20 ± 0.93 | 543.60 ± 1.13 | 442.35 ± 1.26 | 567.56 ± 1.29 | 3639.30 ± 2.77 |
| PUFAs * | |||||||
| cis-8,11,14-Eicosatrienoic acid | nd # | nd # | nd # | 13.94 ± 0.57 | nd # | nd # | 802.59 ± 0.82 |
| Linoelaidic acid | nd # | nd # | nd # | nd # | nd # | nd # | 590.51 ± 1.35 |
| Linoleic acid | 76.92 ± 0.51 | 161.41 ± 0.23 | 251.61 ± 0.50 | 714.43 ± 1.06 | 691.87 ± 0.77 | 519.75 ± 0.63 | nd # |
| γ-Linolenic acid | 38.40 ± 0.77 | 370.84 ± 0.84 | 723.07 ± 0.21 | 205.85 ± 0.45 | 574.54 ± 0.35 | 536.22 ± 0.12 | nd # |
| Arachidonic acid | nd # | nd # | nd # | 173.34 ± 0.27 | nd # | nd # | nd # |
| cis-5,8,11,14,17-Eicosapentaenoic acid | nd # | nd # | nd # | nd # | nd # | nd # | 3650.82 ± 1.97 |
| ∑ others PUFAs | - | - | - | - | - | - | - |
| ∑ PUFAs | 115.31 ± 0.98 | 532.25 ± 1.00 | 974.68 ± 0.59 | 1107.55 ± 1.75 | 1266.40 ± 0.47 | 1055.97 ± 0.75 | 5043.91 ± 3.28 |
| Microalgae | Protein | Carbohydrates | Lipids | Total Dietary Fiber | Reference |
|---|---|---|---|---|---|
| Arthrospira platensis | 50–63 | 7.7–22.2 | - | - | [4,63,64,65] |
| 42.08 ± 0.10 | 21.5 | - | 8.5 | [5] | |
| - | - | 4–9 | - | [66] | |
| 46.76 ± 0.95 | 3.32 ± 0.05 | 1.40 ± 0.12 | 42.82 ± 1.20 | This Study | |
| Scenedesmus almeriensis | 49.4–55 | - | - | - | [49,59] |
| - | 24.6 | - | - | [9] | |
| - | - | 1.58 | - | [66] | |
| −12.93 ± 0.69 | 4.51 ± 0.41 | 2.05 ± 0.12 | 22.60 ± 1.50 | This Study | |
| Haematococcus pluvialis (green phase) | 32.59 ± 1.20 | 0.13 ± 0.01 | 3.24 ± 0.11 | 34.56 ± 0.90 | This Study |
| Haematococcus pluvialis (red phase) | 10.2–17 | - | - | - | [7,51] |
| 25.69 ± 1.27 | 6.30 ± 0.24 | 2.60 ± 0.10 | 58.52 ± 2.56 | This Study | |
| Chlorella vulgaris | 20–60.38 | - | - | - | [67] |
| 25.50–48.19 | 59.71 | - | 16.37–25.95 | [68,69] | |
| - | - | 12–26 | - | [66] | |
| 45.64 ± 1.20 | 5.30 ± 0.50 | 3.13 ± 0.21 | 35.04 ± 1.60 | This Study | |
| Dunaliella salina | 55 | 25–40 | - | - | [49] |
| - | 17 | - | [70] | ||
| 10.03 ± 0.57 | 25.31 ± 1.55 | 3.49 ± 0.10 | 8.97 ± 0.50 | This Study | |
| Nannochloropsis sp. | 41.6–42.1 | 16.7–18.6 | - | - | [5] |
| 28.8 ± 0.63 | 28.7 ± 0.48–40.4 | - | - | [71] | |
| - | 0.39 | - | - | [49] | |
| - | - | >40 | - | [72] | |
| - | - | 25.6–30 | - | [72] | |
| 26.67 ± 1.10 | 32.05 ± 0.70 | - | 17.67 ± 0.80 | This Study |
| Cereals | Moisture | Ash | Proteins | Carbohydrates | Lipids | Total Dietary Fiber | Carotene |
|---|---|---|---|---|---|---|---|
| Wheat | 14 | 1.4 | 10.6 | 61.6–69.7 | 1.4 | 10.5–14.4 | 0.02 |
| Barley | 14 | 1.9 | 11 | 55.8 | 3.4 | 4.7 | n.a. * |
| Rye | 14 | 1.8 | 8.7 | 60.9–71.8 | 1.5 | 13.1 | 0 |
| Brown Rice | 14 | 1.4 | 7.3 | 64.3–71.1 | 2.2 | 4 | 0 |
| Sorghum | 14 | 2.6 | 8.3 | 57.4–62.9 | 3.9 | 13.8 | 10 |
| Oats | 14 | 2.3 | 9.3 | 62.9–63 | 5.9 | 5.5 | 0 |
| Maize | 14 | 1.4 | 9.8 | 60.9–63.6 | 4.9 | 9 | 0.37 |
| Products Name | Application | Characteristics | Regulations | Price |
|---|---|---|---|---|
| A. platensis extract | Additive in food and drugs, coloring agent, dietary supplements | Filtered aqueous extract from dried biomass, principal colorant phycocyanin | CFR-TITLE 21-FDA | US$29.75/120 mL |
| €70–200 kg [81] | ||||
| Dried algae meal (genus Spongiococcum) | Additive for chicken skins, and eggs | A mixture of dried biomass, molasses, corn step liquor after fermentation | CFR-TITLE 21-FDA | US$50/kg [82] |
| Dried T. chuii | Novel food and food supplement; ingredient for sauce, special salt and condiment | Humidity ≤7.0% | Regulation (EC) 2017/2470 Directive 2002/46/CE | US$50/kg [82] |
| Protein 35–40% | ||||
| Carbohydrates 30–32%, ash 14–16% | ||||
| Fiber 2–3% | ||||
| Fat 5–8% | ||||
| SFAs 29–31% | ||||
| MUFAs 21–24% | ||||
| PUFAs 44–49% | ||||
| Iodine ≤15 mg/kg | ||||
| O. aurita | Novel food, food additive in flavored pasta, fish soups, marines terrines, broth preparation, crackers, frozen breaded fish | Diatom with silicon 3.3% | Regulation (EC) 2017/2470 | |
| H. pluvialis meal | Color additive for fish feed (salmonids) | Dry and comminuted solid biomass containing not less than 1.5% astaxanthin | CFR-TITLE 21-FDA | US$489/kg [83] |
| C. vulgaris | Food source falling into the generally recognized as safe (GRAF) category | FDA (2016) Summary: Substances generally regarded as safe (Final Rule). U.S. Food and Drug Administration (HHS). | 50–600/kg [84] | |
| Protein powder and lipid ingredient derived from C. vulgaris | Food source falling into the generally recognized as safe (GRAF) category | FDA (2016) Summary: Substances generally regarded as safe (Final Rule). U.S. Food and Drug Administration (HHS). | €8.30/kg (Production cost) [85] | |
| Beta-carotene (E 160 IV) (73.95 or 73.1095 beta-carotene) | Additive for food, feed, for all drugs including those for eyes, dietary supplements and cosmetics | E-160 IV: oil essential extract from D. salina; beta-carotene content not less than 20% and other carotenes may be present; 73.95: Physical state, solid, 1% percent solution in chloroform; Beta-carotene content 96–100% | Regulation (EC) 231/2012 | US$50–500/ kg |
| Regulation (EC) 1103/2015 | ||||
| Regulation (EC) 1170/2009 | €150/kg [86] | |||
| CFR-TITLE 21-FDA | ||||
| Astaxanthin | Color additive for fish feed | Physical state, solid, 0.05 percent solution in chloroform Astaxanthin content minimum 96% | CFR-TITLE 21-FDA | €99/kg (astaxanthin content: 0.5%) [83] |
| Astaxanthin-rich oleoresin from H. pluvialis | Novel food and food supplement | Astaxanthin is extracted by CO2-SFE or diluted ethyl acetate using olive oil, sunflower oil or medium chain triglycerides. | Regulation (EC) 2017/2470 Directive 2002/46/CE | €499/kg (astaxanthin content: 5%) [83] |
| Protein 0.3–4.4% | ||||
| Carbohydrates 0–52.8% | ||||
| Ash 0.0–4.2% | ||||
| Fiber <1% | ||||
| Fat 42.2–99% | ||||
| Total astaxanthin 2.9–11.1% | ||||
| All-trans astaxanthin 79.9–91.5% | ||||
| 9-cis astaxanthin 0.3–17.3% | ||||
| 13-cis astaxanthin 0.2–7.0% | ||||
| Beta-carotene 0.01–0.3% | ||||
| Lutein 0–1.8% | ||||
| Canthaxanthin 0–1.30% | ||||
| Algal oil from Ulkenia sp. | Novel food for bakery products, cereal bars, non-alcoholic beverages | Non-saponificable fraction ≤ 4.5% | Regulation (EC) 2017/2470 | US$80–160/kg [86] |
| Trans fatty acids ≤ 1.0% | ||||
| DHA content ≥ 32% | ||||
| Schizochytrium sp. Oil rich in DHA and EPA | Novel food and food supplement for adult, pregnant and lactating women, baby food | Non-saponificable fraction ≤ 3.5% | Regulation (EC) 2017/2470 Directive 2002/46/CE | US$80–160/kg [86] |
| Trans fatty acids ≤ 1.0% | ||||
| DHA content ≥ 22.5% | ||||
| EPA content ≥ 10% | ||||
| Nannochloropsis sp.derived oil (rich in EPA) | Safe for use in dietary supplements | FDA (2015) US Food and Drug Administration New Dietary Ingredient Notification Report #826 | US$80–160/kg [86] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molino, A.; Iovine, A.; Casella, P.; Mehariya, S.; Chianese, S.; Cerbone, A.; Rimauro, J.; Musmarra, D. Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. https://doi.org/10.3390/ijerph15112436
Molino A, Iovine A, Casella P, Mehariya S, Chianese S, Cerbone A, Rimauro J, Musmarra D. Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals. International Journal of Environmental Research and Public Health. 2018; 15(11):2436. https://doi.org/10.3390/ijerph15112436
Chicago/Turabian StyleMolino, Antonio, Angela Iovine, Patrizia Casella, Sanjeet Mehariya, Simeone Chianese, Antonietta Cerbone, Juri Rimauro, and Dino Musmarra. 2018. "Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals" International Journal of Environmental Research and Public Health 15, no. 11: 2436. https://doi.org/10.3390/ijerph15112436
APA StyleMolino, A., Iovine, A., Casella, P., Mehariya, S., Chianese, S., Cerbone, A., Rimauro, J., & Musmarra, D. (2018). Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals. International Journal of Environmental Research and Public Health, 15(11), 2436. https://doi.org/10.3390/ijerph15112436