Influence of Afforestation on the Species Diversity of the Soil Seed Bank and Understory Vegetation in the Hill-Gullied Loess Plateau, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Samples and Standing Vegetation Investigation
2.3. Germination Experiments
2.4. Statistical Analysis
- (1)
- Species number as the richness index (S).
- (2)
- Shannon–Wiener diversity index (H’):
- (3)
- Pielou evenness index (J):
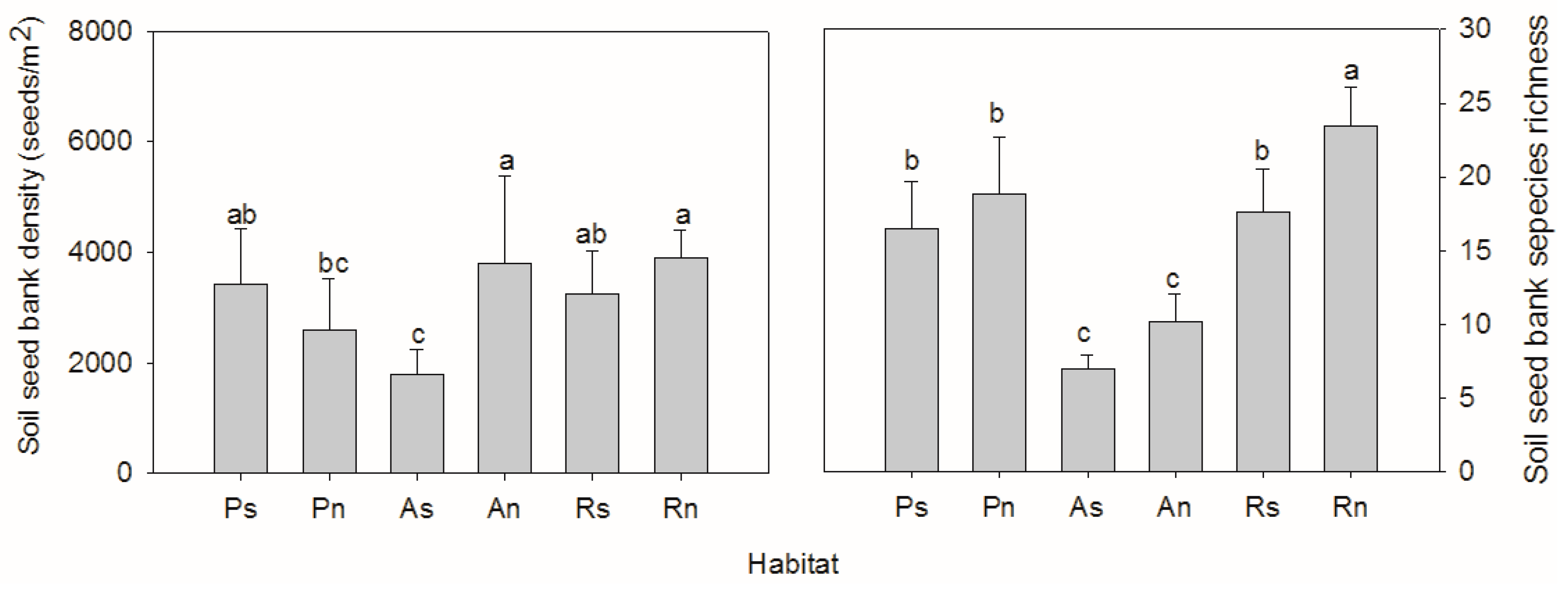
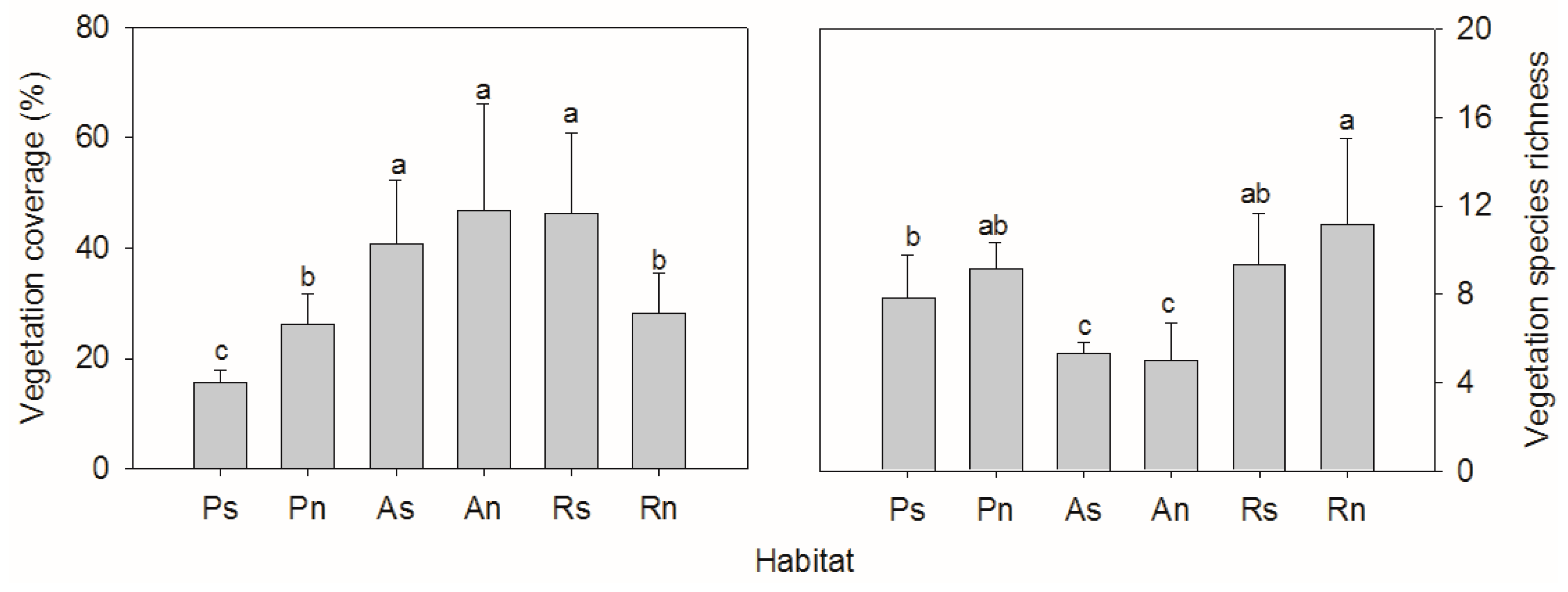
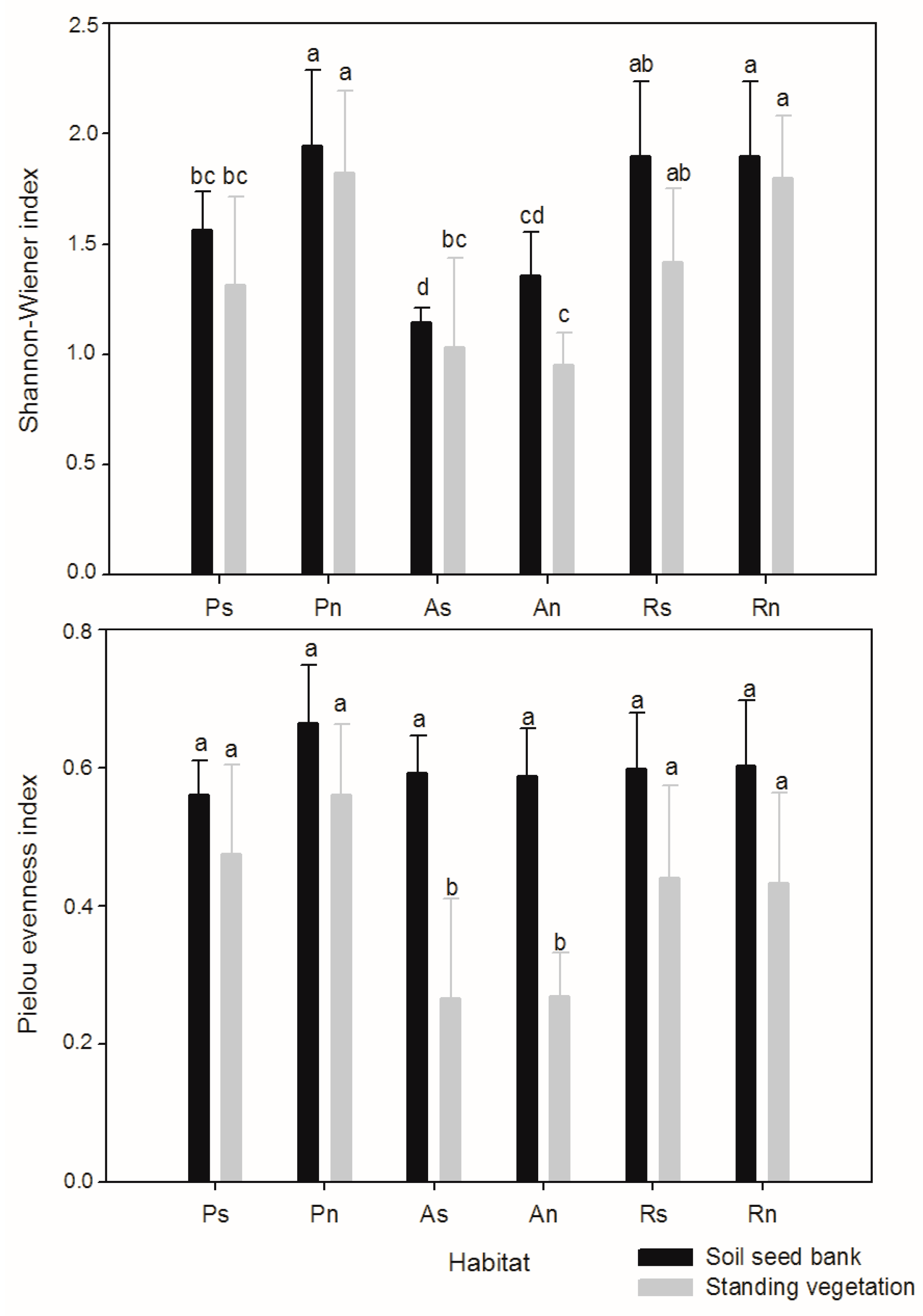
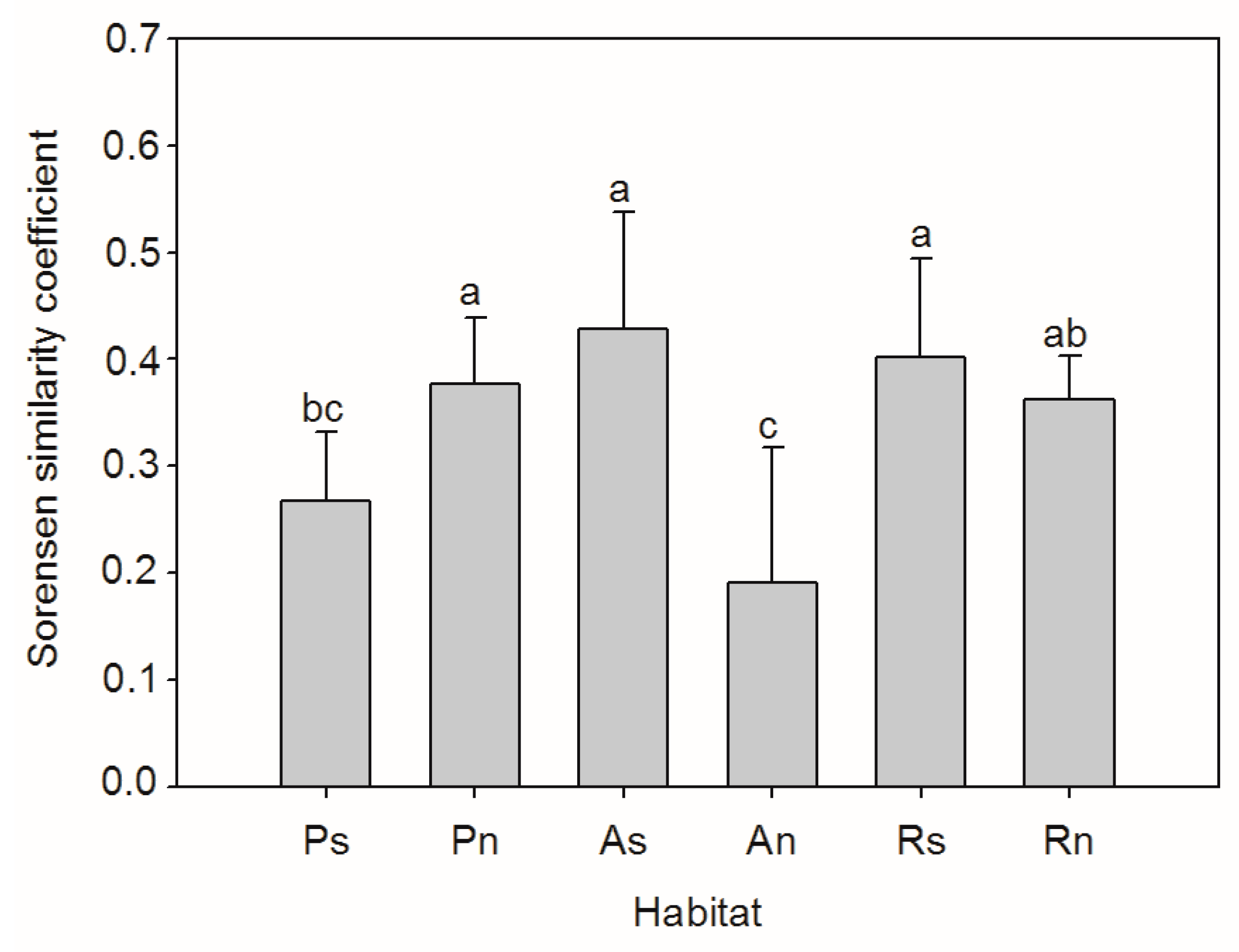
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pimentel, D.; Kounang, N. Ecology of Soil Erosion in Ecosystems. Ecosystems 1998, 1, 416–426. [Google Scholar] [CrossRef]
- Zhang, Q.J.; Fu, B.J.; Chen, L.D.; Zhao, W.W.; Yang, Q.K.; Liu, G.B.; Gulinck, H. Dynamics and driving factors of agricultural landscape in the semiarid hilly area of the Loess Plateau, China. Agric. Ecosyst. Environ. 2004, 103, 535–543. [Google Scholar] [CrossRef]
- Zhao, G.; Mu, X.; Wen, Z.; Wang, F.; Gao, P. Soil erosion, conservation and eco-environment changes in the Loess Plateau of China. Land Degrad. Dev. 2013, 24, 499–510. [Google Scholar] [CrossRef]
- Shi, H.; Shao, M.A. Soil and water loss from the Loess Plateau in China. J. Arid Environ. 2000, 45, 9–20. [Google Scholar] [CrossRef]
- Cerdà, A. Parent material and vegetation affect soil erosion in eastern Spain. Soil Sci. Soc. Am. J. 1999, 63, 362–368. [Google Scholar] [CrossRef]
- Lieskovský, J.; Kenderessy, P. Modeling the effect of vegetation cover and different tillage practices on soil erosion in vineyards: A case study in Vrable (Slovakia) using watem/sedem. Land Degrad. Dev. 2014, 25, 288–296. [Google Scholar] [CrossRef]
- Zhou, Z.C.; Shangguan, Z.P.; Zhao, D. Modeling vegetation coverage and soil erosion in the Loess Plateau Area of China. Ecol. Model. 2006, 198, 263–268. [Google Scholar] [CrossRef]
- Zheng, F.; He, X.; Gao, X.; Zhang, C.; Tang, K. Effects of erosion patterns on nutrient loss following deforestation on the Loess Plateau of China. Agric. Ecosyst. Environ. 2005, 108, 85–97. [Google Scholar] [CrossRef]
- Cerdà, A.; García-Fayos, P. The influence of seed size and shape on their removal by water erosion. Catena 2002, 48, 293–301. [Google Scholar] [CrossRef]
- García-Fayos, P.; Bochet, E.; Cerdà, A. Seed removal susceptibility through soil erosion shapes vegetation composition. Plant Soil 2010, 334, 289–297. [Google Scholar] [CrossRef]
- Jiao, J.; Zou, H.; Jia, Y.; Wang, N. Research progress on the effects of soil erosion on vegetation. Acta Ecol. Sin. 2009, 29, 85–91. [Google Scholar] [CrossRef]
- García-Fayos, P.; García-Ventoso, B.; Cerdà, A. Limitations to plant establishment on eroded slopes in southeastern Spain. J. Veg. Sci. 2000, 11, 77–86. [Google Scholar] [CrossRef]
- De Luís, M.; Raventós, J.; González-Hidalgo, J.C. Fire and torrential rainfall: Effects on seedling establishment in Mediterranean gorse shrublands. J. Int. Assoc. Wildland Fire 2005, 14, 413–422. [Google Scholar] [CrossRef]
- Wang, N.; Jiao, J.Y.; Lei, D.; Chen, Y.; Wang, D.L. Effect of rainfall erosion: Seedling damage and establishment problems. Land Degrad. Dev. 2014, 25, 565–572. [Google Scholar] [CrossRef]
- Becerra, P.I.; Montenegro, G. The widely invasive tree Pinus radiata facilitates regeneration of native woody species in a semi-arid ecosystem. Appl. Veg. Sci. 2013, 16, 173–183. [Google Scholar] [CrossRef]
- Gomez-Aparicio, L.; Zamora, R.; Gomez, J.M.; Hodar, J.A.; Catr, J.; Baraza, E. Applying plant facilitation to forest restoration: A meta-analysis of the useof shrubs as nurse plants. Ecol. Appl. 2004, 14, 1128–1138. [Google Scholar] [CrossRef]
- Ciccarese, L.; Mattsson, A.; Pettenella, D. Ecosystem services from forest restoration: Thinking ahead. New For. 2012, 43, 543–560. [Google Scholar] [CrossRef]
- Li, W. Degradation and restoration of forest ecosystems in China. For. Ecol. Manag. 2004, 201, 33–41. [Google Scholar]
- Chirino, E.; Bonet, A.; Bellot, J.; Sanchez, J.R. Effects of 30-year-old Aleppo pine plantations on runoff, soil erosion, and plant diversity in a semi-arid landscape in south eastern Spain. Catena 2006, 65, 19–29. [Google Scholar] [CrossRef]
- Rey, F.; Berger, F. Management of Austrian black pine on marly lands for sustainable protection against erosion (Southern Alps, france). New For. 2006, 31, 535–543. [Google Scholar] [CrossRef]
- Breshears, D.D.; Nyhan, J.W.; Heil, C.E.; Wilcox, B.P. Effects of woody plants on microclimate in asemiarid woodland: Soil temperature and evaporation in canopy and intercanopy patches. Int. J. Plant Sci. 1998, 159, 1010–1017. [Google Scholar] [CrossRef]
- Wu, J.P.; Liu, Z.F.; Sun, Y.X.; Zhou, L.X.; Lin, Y.B.; Fu, S.L. Introduced Eucalytus urophylla plantations change the composition of the soil microbial community in subtropical China. Land Degrad. Dev. 2013, 24, 400–406. [Google Scholar] [CrossRef]
- Cusack, D.; Montagnini, F. The role of native species plantations in recovery of understory woody diversity in degraded pasturelands of Costa Rica. For. Ecol. Manag. 2004, 188, 1–15. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Kou, M.; Jiao, J.; Yin, Q.; Wang, N.; Wang, Z.; Li, Y.; Yu, W.; Wei, Y.; Yan, F.; Cao, B. Successional trajectory over 10 years of vegetation restoration of abandoned slope croplands in the Hill-gully region of the Loess Plateau. Land Degrad. Dev. 2016, 27, 919–932. [Google Scholar] [CrossRef]
- Gilliam, F.S. The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Bremer, L.L.; Farley, K.A. Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodivers. Conserv. 2010, 19, 3893–3915. [Google Scholar] [CrossRef]
- Chen, L.; He, Z.; Zhu, X.; Du, J.; Yang, J.; Li, J. Impacts of afforestation on plant diversity, soil properties, and soil organic carbon storage in a semi-arid grassland of northwestern China. Catena 2016, 147, 300–307. [Google Scholar] [CrossRef]
- Butler, R.; Montagnini, F.; Arroyo, P. Woody understory plant diversity in pure and mixed native tree plantations at La Selva Biological Station, Costa Rica. For. Ecol. Manag. 2008, 255, 2251–2263. [Google Scholar] [CrossRef]
- Martín-García, J.; Jactel, H.; Oria-de-Rueda, J.A.; Diez, J.J. The Effects of Poplar Plantations on Vascular Plant Diversity in Riparian Landscapes. Forests 2016, 7, 50. [Google Scholar] [CrossRef]
- Norton, D.; Forbes, A. Can exotic pine trees assist in restoration? Appl. Veg. Sci. 2013, 16, 169–170. [Google Scholar] [CrossRef]
- Bakker, J.P.; Poschlod, P.; Strykstra, R.J.; Bekker, R.M.; Thompson, K. Seed banks and seed dispersal: Important topics in restoration ecology. Acta Bot. Neerl. 1996, 45, 461–490. [Google Scholar] [CrossRef]
- Plue, J.; Dupouey, J.L.; Verheyen, K.; Hermy, M. Forest seed banks along an intensity gradient of ancient agriculture. Seed Sci. Res. 2009, 19, 103–114. [Google Scholar] [CrossRef]
- Zamora, C.O.; Montagnini, F. Seed rain and seed dispersal agents in pure and mixed plantations of native trees and abandoned pastures at La Selva Biological Station, Costa Rica. Restor. Ecol. 2007, 15, 453–461. [Google Scholar] [CrossRef]
- Thompson, K. Seeds and seed banks. New Physiol. 1987, 106, 23–34. [Google Scholar] [CrossRef]
- Plue, J.; Gils, B.V.; Peppler-Lisbach, C.; Schrijver, A.D.; Verheyen, K.; Hermy, M. Seed-bank convergence under different tree species during forest development. Perspect. Plant Ecol. Evol. Syst. 2010, 2, 211–218. [Google Scholar] [CrossRef]
- Bossuyt, B.; Hermy, M. Influence of land use history on seed banks in European temperate forest ecosystems: A review. Ecography 2001, 24, 225–238. [Google Scholar] [CrossRef]
- Plue, J.; Verheyen, K.; Calster, H.V.; Marage, D.; Thompson, K.; Kalamees, R.; Jankowska-Blaszczuk, M.; Bossuyt, B.; Hermy, M. Seed banks of temperate deciduous forests during secondary succession. J. Veg. Sci. 2010, 21, 965–978. [Google Scholar] [CrossRef]
- Van Calster, H.; Chevalier, R.; Wyngene, B.; Archaux, F.; Verheyen, K.; Hermy, M. Long-term seed bank dynamics in a temperate forest under conversion from coppice-with-standards to high forest management. Appl. Veg. Sci. 2008, 11, 251–260. [Google Scholar] [CrossRef]
- Bossuyt, B.; Honnay, O. Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J. Veg. Sci. 2008, 19, 875–884. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Physiological ecology of natural regeneration of harvested and disturbed forest stands: Implications for forest management. For. Ecol. Manag. 2002, 158, 195–221. [Google Scholar] [CrossRef]
- Cao, S.; Chen, L.; Shankman, D.; Wang, C.; Wang, X.; Zhang, H. Excessive reliance on afforestation in China’s arid and semi-arid regions: Lessons in ecological restoration. Earth Sci. Rev. 2011, 104, 240–245. [Google Scholar] [CrossRef]
- Jiao, J.; Zhang, Z.; Bai, W.; Jia, Y.; Wang, N. Assessing the ecological success of restoration by afforestation on the Chinese Loess Plateau. Restor. Ecol. 2012, 20, 240–249. [Google Scholar] [CrossRef]
- Jiao, J.; Tzanopoulos, J.; Xofis, P.; Bai, W.; Ma, X.; Mitchley, J. Can the study of natural vegetation succession assist in the control of soil erosion on abandoned croplands on the Loess Plateau, China? Restor. Ecol. 2007, 15, 391–399. [Google Scholar] [CrossRef]
- Tateno, R.; Tokuchi, N.; Yamanaka, N.; Du, S.; Otsuki, K.; Shimamura, T.; Xue, Z.; Wang, S.; Hou, Q. Comparison of litterfall production and leaf litter decomposition between an exotic blacklocustplantation and an indigenous oak forest near Yan’an on the Loess Plateau, China. For. Ecol. Manag. 2007, 241, 84–90. [Google Scholar] [CrossRef]
- Qiu, L.; Zhang, X.; Cheng, J.; Yin, X. Effects of black locust (Robinia pseudoacacia) on soil properties in the loessial gully region of the Loess Plateau, China. Plant Soil 2010, 332, 207–217. [Google Scholar] [CrossRef]
- Chang, R.; Fu, B.; Liu, G.; Liu, S. Soil carbonsequestration potential for “Grain for Green” project in Loess Plateau, China. Environ. Manag. 2011, 48, 1158–1172. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, P.; Feger, K.; Wei, X.; Sun, G.; Bonell, M.; Xiong, W.; Zhang, S.; Xu, L. Annual runoff and evapotranspiration of forestlands and non-forestlands in selected basins of the Loess Plateau of China. Ecohydrology 2011, 4, 277–287. [Google Scholar] [CrossRef]
- He, X.; Tian, J.; Tang, K.; Sun, J.; Matthews, J.A. Bio-climatic imprints on a Holocene loess palaeosol from China. J. Asian Earth Sci. 2004, 22, 455–464. [Google Scholar]
- Lü, H.; Liu, D.; Guo, Z. Natural vegetation of geological and historical periods in Loess Plateau. Chin. Sci. Bull. 2003, 48, 411–416. [Google Scholar] [CrossRef]
- Wang, N.; Jiao, J.; JIa, Y.; Wang, D. Seed persistence in the soil on eroded slopes in the hilly-gullied Loess Plateau region, China. Seed Sci. Res. 2011, 21, 295–304. [Google Scholar] [CrossRef]
- Wang, N.; Jiao, J.; JIa, Y.; Bai, W.; Zhang, Z. Germinable soil seed banks and the restoration potential of abandoned cropland on the Chinese hilly-gullied Loess Plateau. Environ. Manag. 2010, 6, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yang, Y.; Leng, P.S.; Dou, D.Q.; Zhang, B.; Hou, B.F. Characteristics of soil seed bank in plantation forest in the rocky mountain region of Beijing, China. J. For. Res. 2013, 24, 91–97. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, Y. Factors affecting species diversity of plant communities and the restoration process in the loess area of China. Ecol. Eng. 2010, 36, 345–350. [Google Scholar] [CrossRef]
- Jia, G.; Liu, B.; Wang, G.; Zhang, B. The microbial biomass and activity in soil with shrub (Caragana korshinskii K.) plantation in the semi-arid loess plateau in China. Eur. J. Soil Biol. 2010, 46, 6–10. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, J.; Liu, G.; Yamanaka, N. Soil properties in natural grassland, Caragana korshinskii planted shrubland, and Robinia pseudoacacia planted forest in gullies on the hilly Loess Plateau, China. Catena 2014, 119, 116–124. [Google Scholar] [CrossRef]
- Kou, M.; Garcia-Fayos, P.; Hu, S.; Jiao, J. The effect of Robinia pseudoacacia afforestation on soil and vegetation properties in the Loess Plateau (China): A chronosequence approach. For. Ecol. Manag. 2016, 375, 146–158. [Google Scholar] [CrossRef]
- Zahawi, R.A.; Augspurger, C.K. Early plant succession in abandoned pastures in Ecuador. Biotropica 1999, 31, 540–552. [Google Scholar] [CrossRef]
- Ruprecht, E. Successfully recovered grassland: A promising example from Romanian old-fields. Restor. Ecol. 2006, 14, 473–480. [Google Scholar] [CrossRef]
- Jiang, W.; Yang, S.; Yang, X.; Gu, N. Negative impacts of afforestation and economic forestry on the Chinese Loess Plateau and proposed solutions. Quatern. Int. 2016, 399, 165–173. [Google Scholar] [CrossRef]
- Shi, H.; Wen, Z.; Paull, D.; Jiao, F. Distribution of Natural and Planted Forests in the Yanhe River Catchment: Have We Planted Trees on the Right Sites? Forests 2016, 7, 258. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Soil Seed Bank (seeds/m2) | Vegetation Cover (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ps | Pn | As | An | Rs | Rn | Ps | Pn | As | An | Rs | Rn | |
| Annuals/biennials | ||||||||||||
| Amaranthus tricolor | 2 | |||||||||||
| Androsace septentrionalis | 23 | 28 | 4 | |||||||||
| Artemisia hedinii | 94 | 6 | 250 | 553 | 11 | 0.2 | ||||||
| Artemisia scoparia | 1701 | 966 | 983 | 2072 | 1463 | 1448 | 0.1 | 0.1 | ||||
| Bidens pilosa | 3 | 6 | ||||||||||
| Bothriospermum chinense | 5 | 31 | 4 | 4 | ||||||||
| Chenopodium serotinum | 11 | 11 | 4 | 2 | ||||||||
| Digitaria sanguinalis | 2 | |||||||||||
| Dracocephalum moldavica | 20 | 18 | 28 | 11 | 7 | 4 | 0.0 | 0.8 | 0.1 | |||
| Echinochloa crusgali | 2 | |||||||||||
| Euphorbia humifusa | 2 | 15 | 6 | 67 | 6 | 13 | ||||||
| Eragrostis pilosa | 17 | 2 | 6 | 15 | ||||||||
| Galium aparine | 0.3 | |||||||||||
| Incarvillea sinensis | 0.1 | |||||||||||
| Ixeris polycephala | 2 | 11 | 11 | |||||||||
| Kochia scoparia | 3 | 2 | ||||||||||
| Leonurus artemisia | 17 | |||||||||||
| Linum stelleroides | 12 | 15 | 26 | |||||||||
| Plantago asiatica | 2 | 6 | 2 | |||||||||
| Salsola collina | 61 | 57 | 2 | 0.6 | 0.9 | |||||||
| Saussurea japonica | 0.1 | 0.1 | 0.2 | |||||||||
| Setaria viridis | 78 | 21 | 22 | 122 | 35 | 83 | 0.2 | |||||
| Sonchus oleraceus | 2 | |||||||||||
| Swertia bimaculata | 123 | 370 | 133 | |||||||||
| Torilis scabra | 2 | |||||||||||
| Torularia humilis | 11 | 158 | 50 | |||||||||
| Perennial herbs | ||||||||||||
| Artemisia giraldii | 12 | 6 | 2 | 41 | 0.6 | 0.8 | 9.4 | 1.2 | ||||
| Artemisia mongolica | 0.3 | 0.4 | 0.1 | |||||||||
| Astragalus adsurgens | 3 | 39 | 2 | 2 | ||||||||
| Astragalus scaberrimus | 0.1 | |||||||||||
| Bupleurum yinchowense | 11 | 0.4 | ||||||||||
| Dendranthema indicum | 3 | 11 | 1.7 | 4.9 | ||||||||
| Diarthron linifolium | 0.0 | |||||||||||
| Geranium wilfordii | 3 | |||||||||||
| Gerbera anandria | 7 | |||||||||||
| Glycyrrhiza uralensis | 0.6 | 0.8 | 0.8 | 0.2 | ||||||||
| Heteropappus altaicus | 3 | 18 | 6 | 78 | 20 | 20 | 0.5 | 0.7 | 0.2 | 0.1 | ||
| Ixeridium chinense | 57 | 11 | 11 | 11 | 55 | 13 | ||||||
| Ixeris sonchifolia Bge. Hance | 149 | 48 | 22 | 37 | 175 | 0.1 | 0.1 | |||||
| Leontopodium leontopodioides | 2 | 0.3 | ||||||||||
| Linum perenne | 2 | |||||||||||
| Melilotus officinalis | 3 | 17 | 4 | 18 | 0.1 | 1.7 | 0.7 | 0.1 | 0.3 | |||
| Oxytropis discolor | 2 | 0.1 | ||||||||||
| Patrinia scabiosaefolia | 12 | 23 | 6 | 9 | 74 | 2.1 | 0.8 | 3.6 | ||||
| Polygala tenuifolia | 17 | 2 | 11 | 2 | 0.1 | 0.2 | ||||||
| Potentilla bifurca | 0.7 | 0.1 | ||||||||||
| Potentilla tanacetifolia | 66 | 44 | 2 | 4 | 0.2 | 0.8 | 0.0 | |||||
| Pulsatilla chinensis | 0.2 | |||||||||||
| Rubia cordifolia | 2 | |||||||||||
| Serratula centauroides | 0.2 | |||||||||||
| Thalictrum aquilegifolium | 0.7 | |||||||||||
| Vicia amoena Fisch. ex DC. | 2 | 0.3 | 4.4 | |||||||||
| Viola dissecta | 2 | 29 | 9 | 15 | 0.1 | |||||||
| Viola philippica | 0.1 | |||||||||||
| Perennial grasses | ||||||||||||
| Bothriochloa ischcemum | 3 | 11 | 556 | 103 | 4 | 0.2 | 29.0 | 1.3 | ||||
| Carex lanceolata | 20 | |||||||||||
| Cleistogenes caespitosa | 0.2 | |||||||||||
| Cleistogenes chinensis | 6 | 8 | 311 | 17 | 0.3 | 0.4 | 3.6 | 0.1 | ||||
| Cleistogenes hancei | 6 | 0.3 | ||||||||||
| Cleistogenes squarrosa | 17 | 1.0 | 1.3 | |||||||||
| Helictotrichon schellianum | 0.1 | |||||||||||
| Koeleria cristata | 0.5 | |||||||||||
| Leymus secalinus | 2 | |||||||||||
| Melica radula | 98 | |||||||||||
| Melica scabrosa | 54 | 9 | 24 | 0.2 | 0.4 | |||||||
| Pennisetum centrasiaticum | 0.2 | |||||||||||
| Phragmites australis | 0.7 | 0.4 | ||||||||||
| Poa sphondylodes | 63 | 193 | 907 | 134 | 1074 | 0.1 | ||||||
| Stipa bungeana | 743 | 89 | 39 | 17 | 11 | 48 | 9.0 | 4.1 | 0.5 | 2.3 | 0.6 | 0.6 |
| Roegneria kamoji | 2 | 0.1 | 0.0 | |||||||||
| Shrubs/Sub-shrubs | ||||||||||||
| Ampelopsis glandulosa | 0.8 | 1.7 | ||||||||||
| Artemisia gmelinii | 52 | 180 | 11 | 89 | 41 | 326 | 1.3 | 11.8 | 2.2 | 30.0 | 7.7 | 8.5 |
| Buddleja alternifolia | 3 | 114 | 6 | 3.1 | ||||||||
| Caragana korshinskii | 1.2 | 3.3 | ||||||||||
| Clematis fruticosa | 4 | 2 | 0.9 | |||||||||
| Lespedeza cuneata | 2 | 0.1 | ||||||||||
| Lespedeza davurica | 5 | 9 | 44 | 33 | 77 | 13 | 0.7 | 0.8 | 3.3 | 3.4 | 0.3 | |
| Lespedeza floribunda | 3.9 | |||||||||||
| Periploca sepium | 2 | 0.5 | 0.8 | 0.2 | ||||||||
| Prinsepia utilis | 2.3 | |||||||||||
| Sophora davidii | 24 | 7.9 | ||||||||||
| Syringa oblata | 6 | |||||||||||
| Rubus parvifolius | 0.1 | |||||||||||
| Ziziphus jujuba | 0.3 | |||||||||||
| Trees | ||||||||||||
| Ailanthus giraldii | 0.7 | |||||||||||
| Robinia pseudoacacia | 71 | 2 | ||||||||||
| Ulmus pumila | 3.3 | |||||||||||
| Items | Ps | Pn | As | An | Rs | Rn | |
|---|---|---|---|---|---|---|---|
| Soil seed bank density (seeds/m2) | A | 2109 | 1523 | 1061 | 2583 | 2277 | 1803 |
| P | 247 | 246 | 67 | 156 | 151 | 385 | |
| G | 870 | 310 | 594 | 941 | 560 | 1291 | |
| S | 60 | 190 | 56 | 122 | 260 | 354 | |
| T | 71 | 2 | |||||
| Soil seed bank species richness | A | 13 | 12 | 6 | 10 | 17 | 14 |
| P | 9 | 14 | 4 | 5 | 10 | 14 | |
| G | 6 | 5 | 2 | 3 | 4 | 9 | |
| S | 3 | 3 | 2 | 2 | 5 | 6 | |
| T | 1 | 1 | |||||
| Vegetation cover (%) | A | 1.2 | 1.1 | 1.2 | 0.2 | ||
| P | 1.6 | 6.0 | 4.5 | 4.2 | 9.9 | 15.1 | |
| G | 10.3 | 5.3 | 30.7 | 3.8 | 5.5 | 2.1 | |
| S | 2.5 | 13.8 | 5.5 | 34.2 | 30.3 | 10.9 | |
| T | 3.3 | 0.7 | |||||
| Vegetation species richness | A | 6 | 3 | 3 | 2 | ||
| P | 6 | 12 | 6 | 5 | 5 | 12 | |
| G | 5 | 5 | 4 | 3 | 3 | 7 | |
| S | 3 | 3 | 2 | 3 | 9 | 6 | |
| T | 1 | 1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Jiao, J.; Jia, Y.; Wang, D. Influence of Afforestation on the Species Diversity of the Soil Seed Bank and Understory Vegetation in the Hill-Gullied Loess Plateau, China. Int. J. Environ. Res. Public Health 2017, 14, 1285. https://doi.org/10.3390/ijerph14101285
Wang N, Jiao J, Jia Y, Wang D. Influence of Afforestation on the Species Diversity of the Soil Seed Bank and Understory Vegetation in the Hill-Gullied Loess Plateau, China. International Journal of Environmental Research and Public Health. 2017; 14(10):1285. https://doi.org/10.3390/ijerph14101285
Chicago/Turabian StyleWang, Ning, Juying Jiao, Yanfeng Jia, and Dongli Wang. 2017. "Influence of Afforestation on the Species Diversity of the Soil Seed Bank and Understory Vegetation in the Hill-Gullied Loess Plateau, China" International Journal of Environmental Research and Public Health 14, no. 10: 1285. https://doi.org/10.3390/ijerph14101285
APA StyleWang, N., Jiao, J., Jia, Y., & Wang, D. (2017). Influence of Afforestation on the Species Diversity of the Soil Seed Bank and Understory Vegetation in the Hill-Gullied Loess Plateau, China. International Journal of Environmental Research and Public Health, 14(10), 1285. https://doi.org/10.3390/ijerph14101285