
2,4,6-Tribromophenol Interferes with the Thyroid Hormone System by Regulating Thyroid Hormones and the Responsible Genes in Mice

 , ,
, , 
Abstract
:1. Introduction
2. Experimental Section
2.1. Chemicals
2.2. Animals
2.3. Quantitative Real-Time qPCR
2.4. Histology
2.5. Serum Hormone Analysis
2.6. Statistical Analysis
3. Results
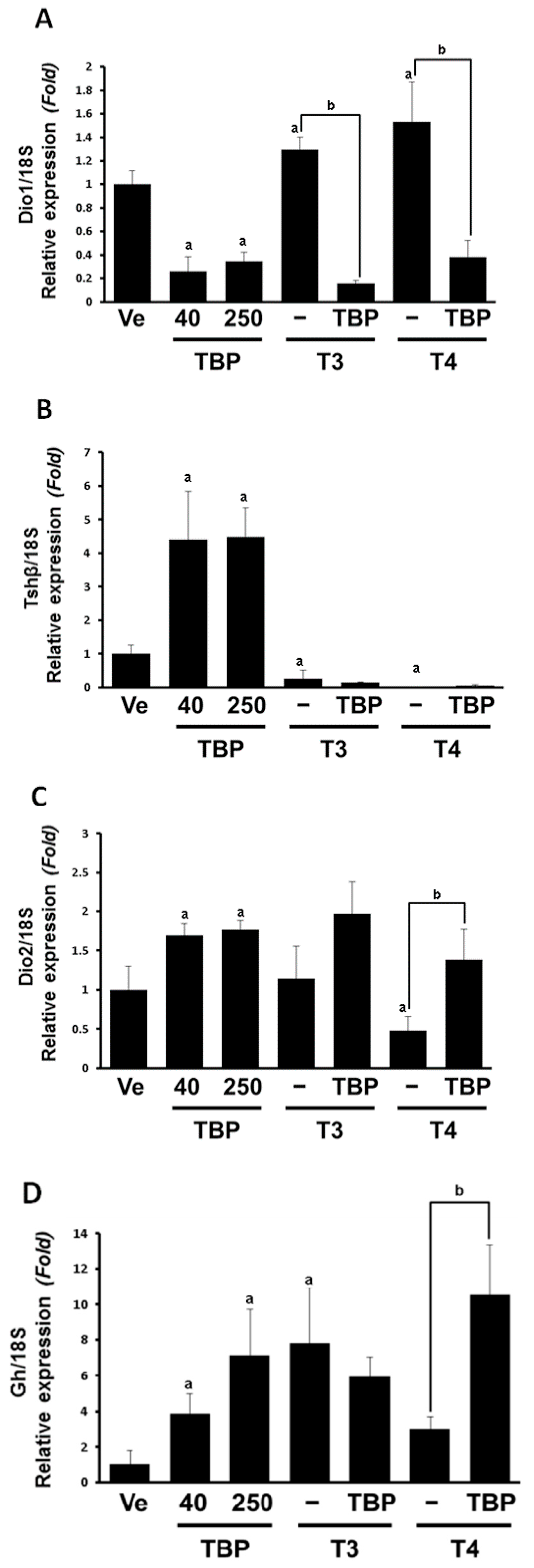
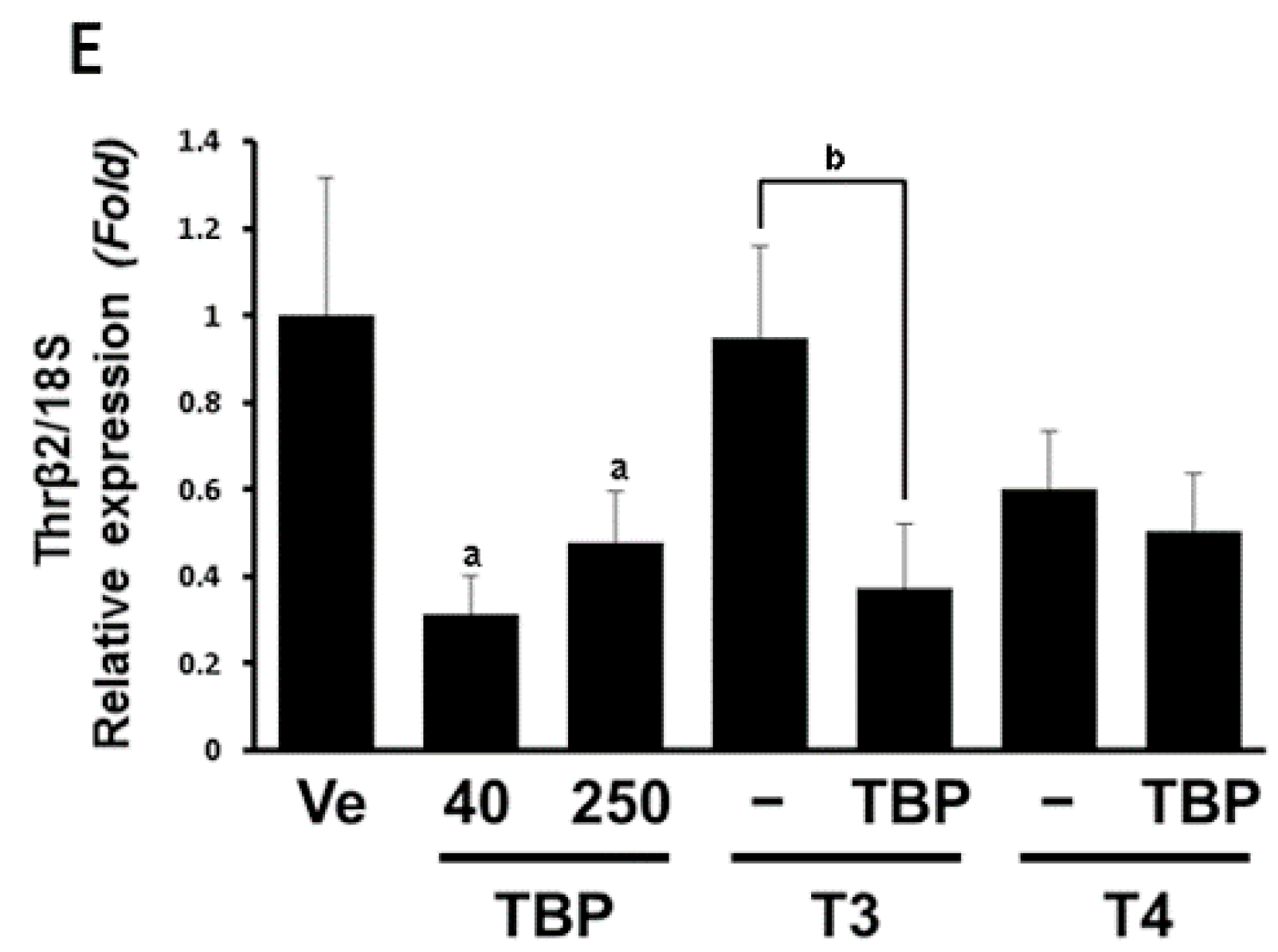
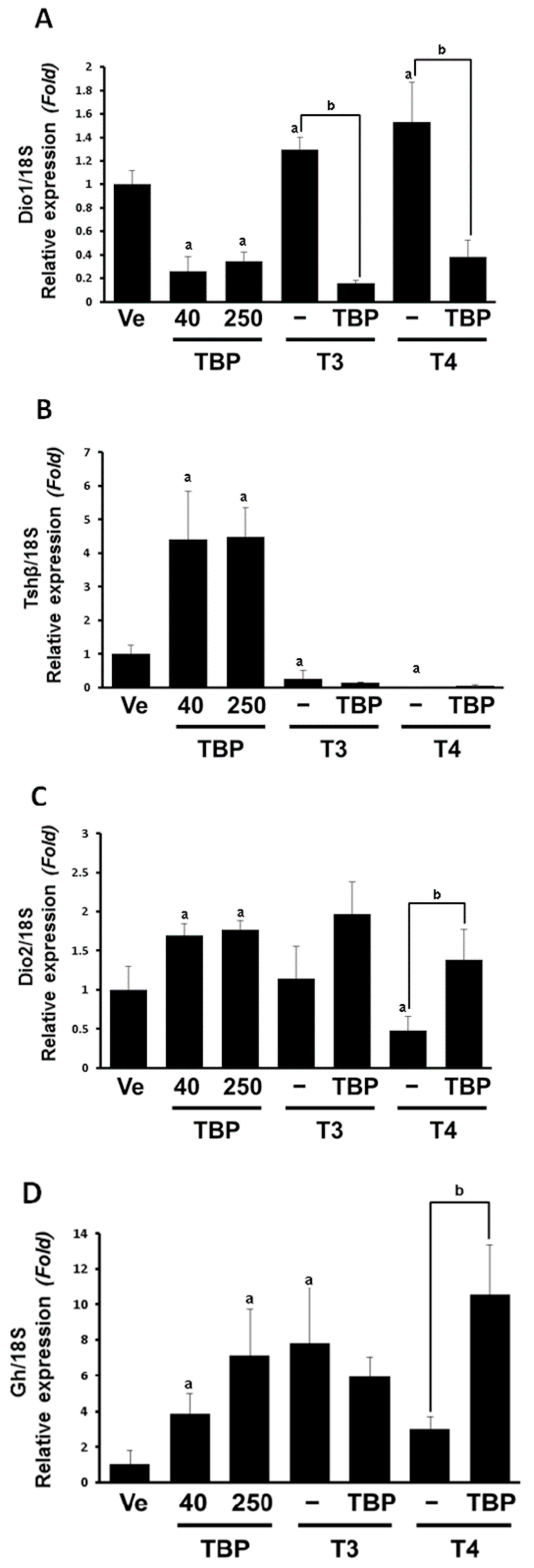
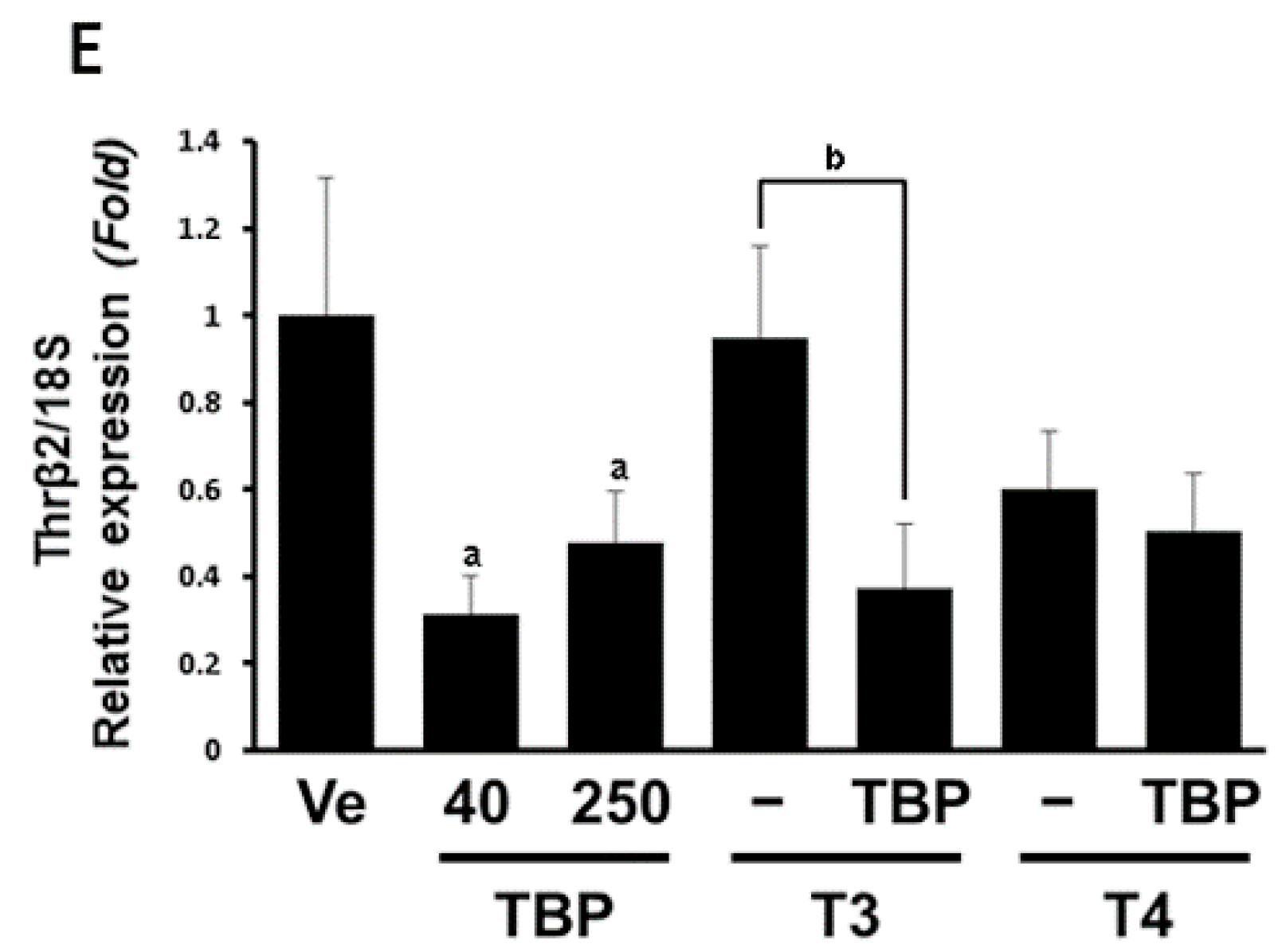
3.1. The mRNA Expression of Thyroid Hormone-Related Genes in the Pituitary Gland
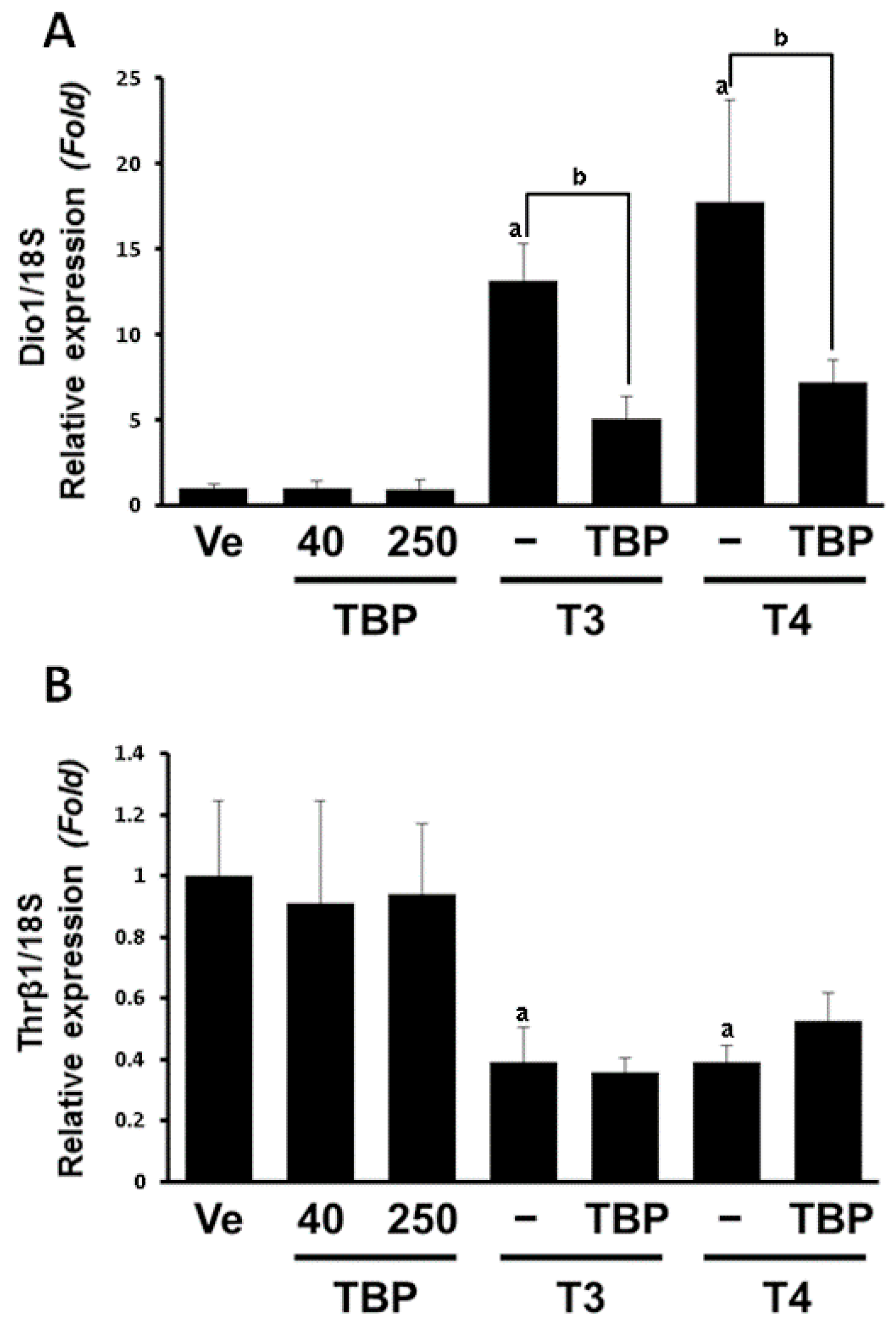
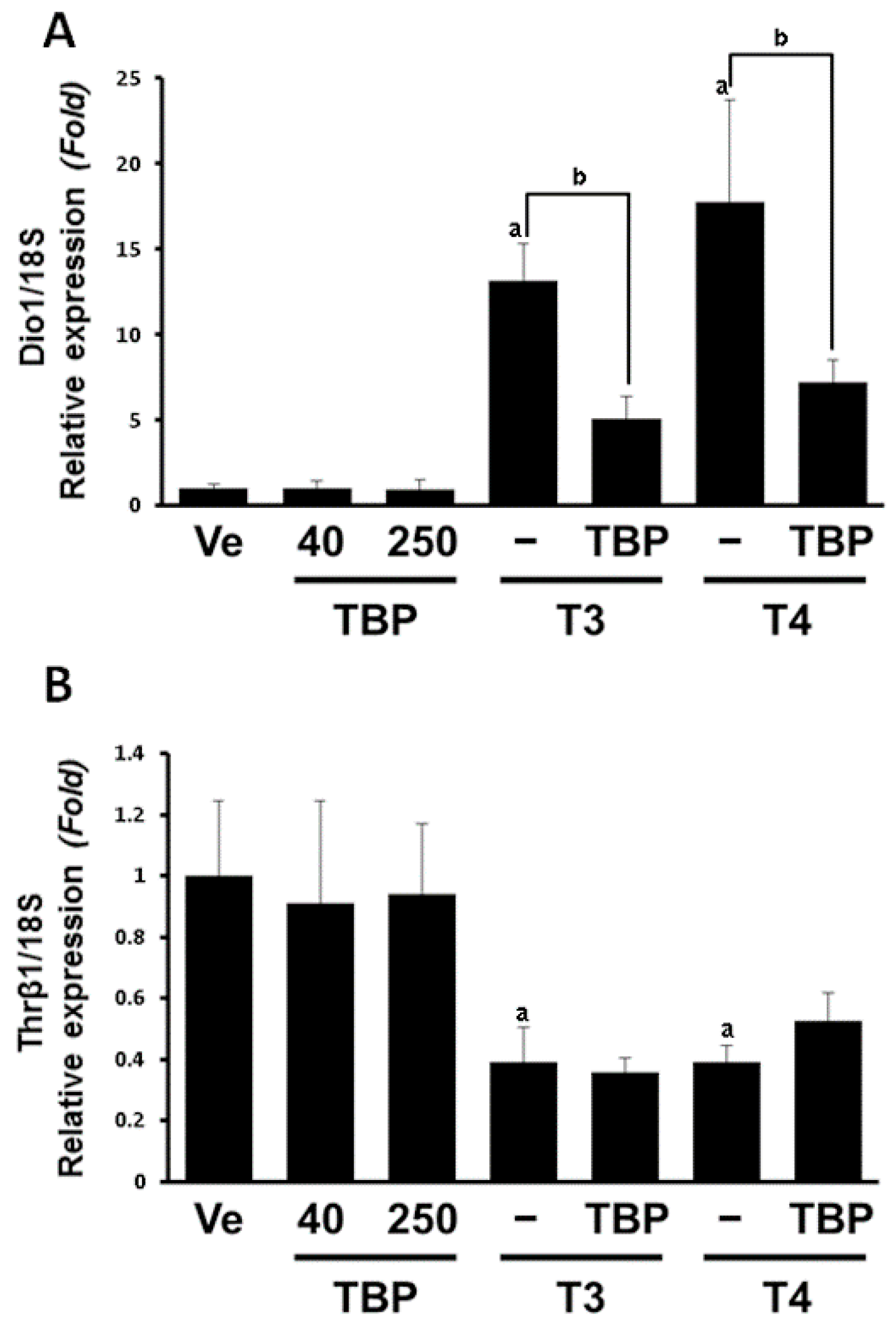
3.2. The mRNA Expression of Dio1 and Thrβ1 in the Liver
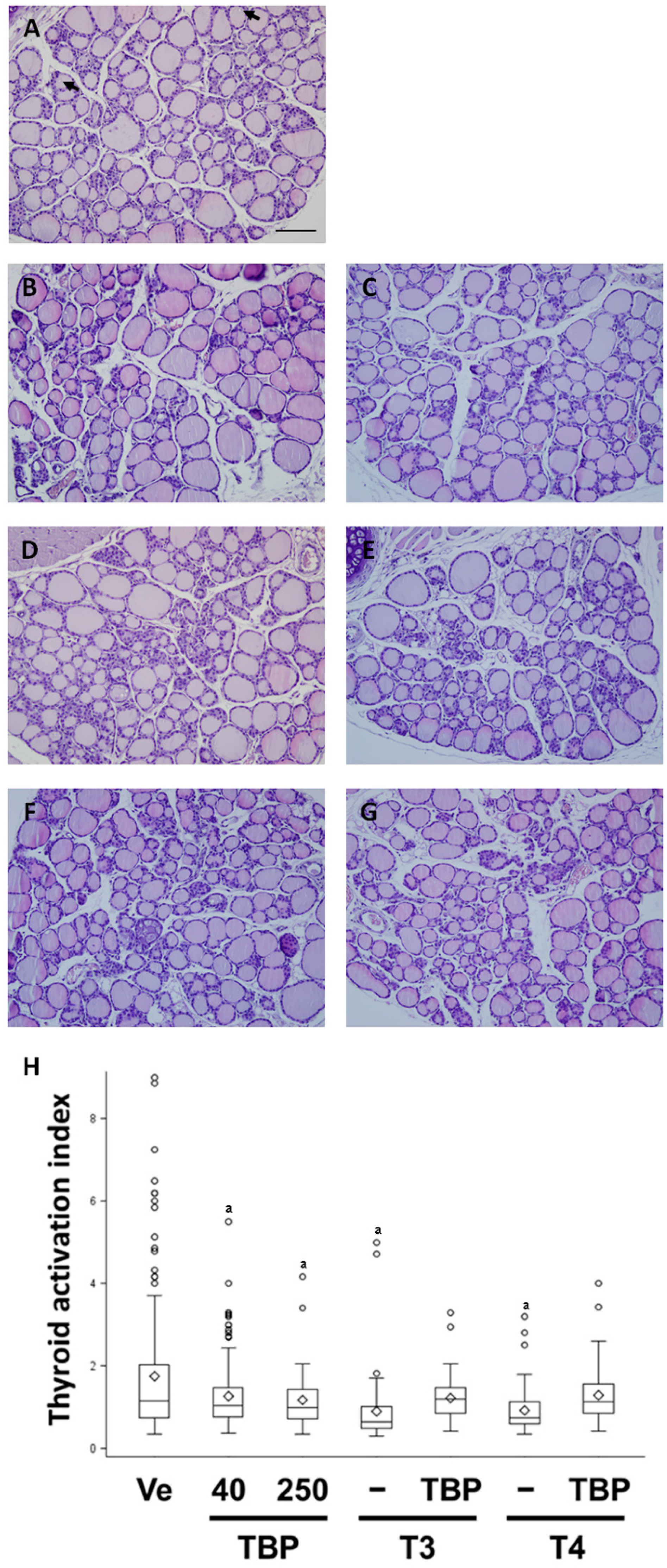
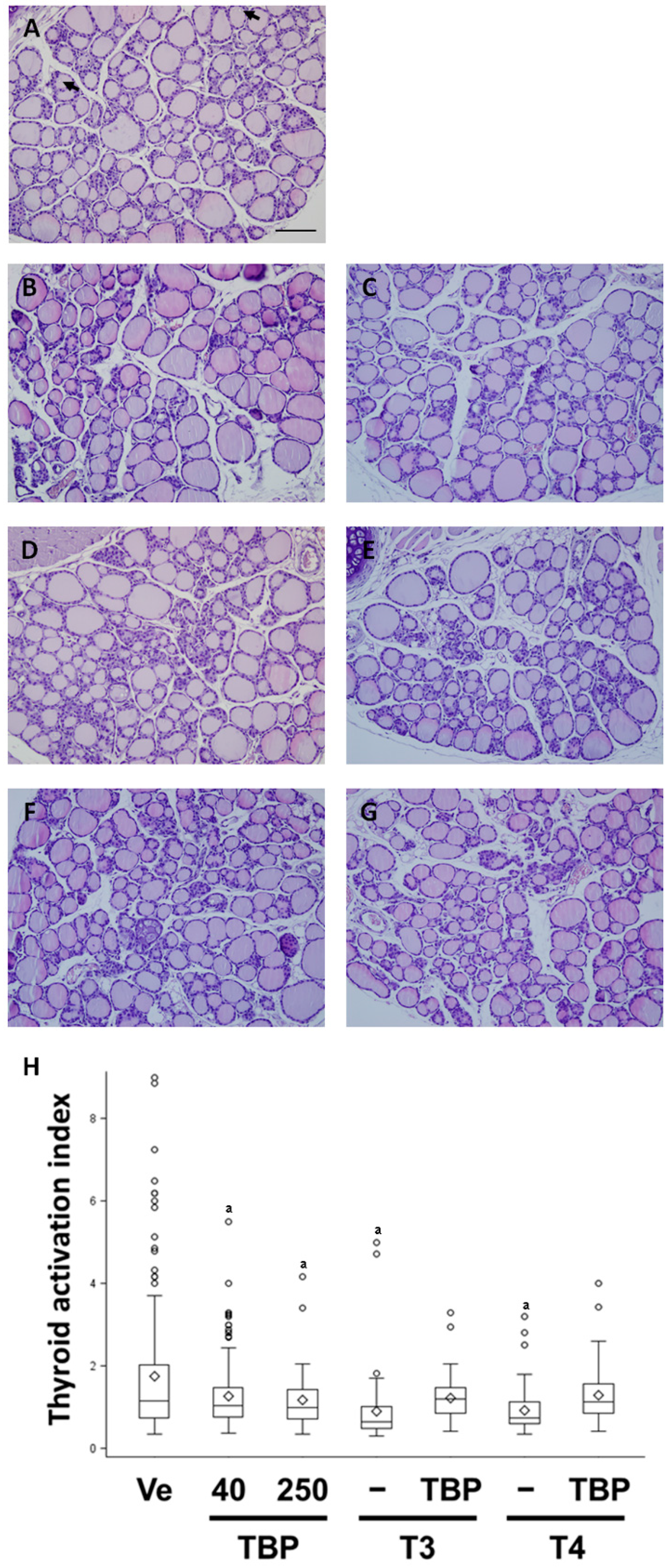
3.3. Morphological and Histological Analysis of Thyroid Gland
3.4. Concentration of Free T3 (fT3) and T4 (fT4) Levels in Serum
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vaitkus, J.A.; Farrar, J.S.; Celi, F.S. Thyroid hormone mediated modulation of energy expenditure. Int. J. Mol. Sci. 2015, 16, 16158–16175. [Google Scholar] [CrossRef] [PubMed]
- Schussler, G.C. The thyroxine-binding proteins. Thyroid 2000, 10, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Faix, J.D. Principles and pitfalls of free hormone measurements. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Grajower, M.M.; Surks, M.I. Effect of decreased hepatic nuclear l-triiodothyronine receptors on the response of hepatic enzymes to l-triiodothyronine in tumor-bearing rats. Endocrinology 1979, 104, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Bassett, J.H.; Harvey, C.B.; Williams, G.R. Mechanisms of thyroid hormone receptor-specific nuclear and extra nuclear actions. Mol. Cell. Endocrinol. 2003, 213, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, T.; Hiroi, H.; Tsutsumi, O.; Ishikawa, T.; Osuga, Y.; Fujiwara, T.; Inoue, S.; Muramatsu, M.; Momoeda, M.; Taketani, Y. The activity of bisphenol a depends on both the estrogen receptor subtype and the cell type. Endocr. J. 2002, 49, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.M.; An, B.S.; Yang, H.; Choi, K.C.; Jeung, E.B. Biomarker genes for detecting estrogenic activity of endocrine disruptors via estrogen receptors. Int. J. Environ. Res. Public Health 2012, 9, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, K.; Tagami, T.; Akamizu, T.; Usui, T.; Saijo, M.; Kanamoto, N.; Hataya, Y.; Shimatsu, A.; Kuzuya, H.; Nakao, K. Thyroid hormone action is disrupted by bisphenol a as an antagonist. J. Clin. Endocrinol. Metab. 2002, 87, 5185–5190. [Google Scholar] [PubMed]
- Guo, Y.; Zhou, B. Thyroid endocrine system disruption by pentachlorophenol: An in vitro and in vivo assay. Aquat. Toxicol. 2013, 142–143, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Sharan, S.; Nikhil, K.; Roy, P. Disruption of thyroid hormone functions by low dose exposure of tributyltin: An in vitro and in vivo approach. Gen. Comp. Endocrinol. 2014, 206, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Zhai, W.; Huang, Z.; Chen, L.; Feng, C.; Li, B.; Li, T. Thyroid endocrine disruption in zebrafish larvae after exposure to mono-(2-ethylhexyl) phthalate (mehp). PLoS ONE 2014, 9, e92465. [Google Scholar] [CrossRef] [PubMed]
- Arrojo E Drigo, R.; Fonseca, T.L.; Werneck-de-Castro, J.P.; Bianco, A.C. Role of the type 2 iodothyronine deiodinase (D2) in the control of thyroid hormone signaling. Biochim. Biophys. Acta 2013, 1830, 3956–3964. [Google Scholar] [CrossRef] [PubMed]
- Covaci, A.; Harrad, S.; Abdallah, M.A.; Ali, N.; Law, R.J.; Herzke, D.; de Wit, C.A. Novel brominated flame retardants: A review of their analysis, environmental fate and behaviour. Environ. Int. 2011, 37, 532–556. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, I.; Sakai, S. Environmental release and behavior of brominated flame retardants. Environ. Int. 2003, 29, 665–682. [Google Scholar] [CrossRef]
- Meerts, I.A.; van Zanden, J.J.; Luijks, E.A.; van Leeuwen-Bol, I.; Marsh, G.; Jakobsson, E.; Bergman, A.; Brouwer, A. Potent competitive interactions of some brominated flame retardants and related compounds with human transthyretin in vitro. Toxicol. Sci. 2000, 56, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Ezechiáš, M.; Svobodová, K.; Cajthaml, T. Hormonal activities of new brominated flame retardants. Chemosphere 2012, 87, 820–824. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Liu, C.; Yu, L.; Zhou, B. Chronic exposure to environmental levels of tribromophenol impairs zebrafish reproduction. Toxicol. Appl. Pharmacol. 2010, 243, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Legler, J.; Brouwer, A. Are brominated flame retardants endocrine disruptors? Environ. Int. 2003, 29, 879–885. [Google Scholar] [CrossRef]
- Olsen, C.M.; Meussen-Elholm, E.T.; Holme, J.A.; Hongslo, J.K. Brominated phenols: Characterization of estrogen-like activity in the human breast cancer cell-line MCF-7. Toxicol. Lett. 2002, 129, 55–63. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Kalisnik, M. A histometric thyroid gland activation index (preliminary report). J. Microsc. 1972, 95, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.E.; Wilson, V.; Noriega, N.; Lambright, C.; Furr, J.; Stoker, T.E.; Laws, S.C.; Goldman, J.; Cooper, R.L.; Foster, P.M. Use of the laboratory rat as a model in endocrine disruptor screening and testing. ILAR J. 2004, 45, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.C.; Leung, P.C.; Jeung, E.B. Biology and physiology of calbindin-D9k in female reproductive tissues: Involvement of steroids and endocrine disruptors. Reprod. Biol. Endocrinol. 2005, 3, 66. [Google Scholar] [CrossRef] [PubMed]
- Baur, A.; Bauer, K.; Jarry, H.; Köhrle, J. 3,5-diiodo-l-thyronine stimulates type 1 5’deiodinase activity in rat Anterior Pituitaries in vivo and in Reaggregate Cultures and GH3 Cells in vitro. Endocrinology 1997, 138, 3242–3248. [Google Scholar] [CrossRef] [PubMed]
- Luongo, C.; Martin, C.; Vella, K.; Marsili, A.; Ambrosio, R.; Dentice, M.; Harney, J.W.; Salvatore, D.; Zavacki, A.M.; Larsen, P.R. The selective loss of the type 2 iodothyronine deiodinase in mouse thyrotrophs increases basal TSH but blunts the thyrotropin response to hypothyroidism. Endocrinology 2015, 156, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Medina-Gomez, G.; Calvo, R.M.; Obregon, M.J. Thermogenic effect of triiodothyroacetic acid at low doses in rat adipose tissue without adverse side effects in the thyroid axis. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E688–E697. [Google Scholar] [CrossRef] [PubMed]
- Maia, A.L.; Goemann, I.M.; Meyer, E.L.; Wajner, S.M. Deiodinases: The balance of thyroid hormone: Type 1 iodothyronine deiodinase in human physiology and disease. J. Endocrinol. 2011, 209, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Mullur, R.; Liu, Y.Y.; Brent, G.A. Thyroid hormone regulation of metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [PubMed]
- Menezes-Ferreira, M.M.; Petrick, P.A.; Weintraub, B.D. Regulation of thyrotropin (TSH) bioactivity by tsh-releasing hormone and thyroid hormone. Endocrinology 1986, 118, 2125–2130. [Google Scholar] [CrossRef] [PubMed]
- De Greef, W.J.; Rondeel, J.M.; van Haasteren, G.A.; Klootwijk, W.; Visser, T.J. Regulation of hypothalamic trh production and release in the rat. Acta Med. Austriaca 1992, 19, 77–79. [Google Scholar] [PubMed]
- Taurog, A.; Dorris, M.L.; Doerge, D.R. Mechanism of simultaneous iodination and coupling catalyzed by thyroid peroxidase. Arch. Biochem. Biophys. 1996, 330, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Lamas, L.; Taurog, A. The importance of thyroglobulin structure in thyroid peroxidase-catalyzed conversion of diiodotyrosine to thyroxine. Endocrinology 1977, 100, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Shivaraj, G.; Prakash, B.D.; Sonal, V.; Shruthi, K.; Vinayak, H.; Avinash, M. Thyroid function tests: A review. Eur. Rev. Med. Pharmacol. Sci. 2009, 13, 341–349. [Google Scholar] [PubMed]
- Salmela, P.I.; Wide, L.; Juustila, H.; Ruokonen, A. Effects of thyroid hormones (T4,T3), bromocriptine and Triac on inappropriate TSH hypersecretion. Clin. Endocrinol. 1988, 28, 497–507. [Google Scholar] [CrossRef]
- Ernest, S.R.; Wade, M.G.; Lalancette, C.; Ma, Y.Q.; Berger, R.G.; Robaire, B.; Hales, B.F. Effects of chronic exposure to an environmentally relevant mixture of brominated flame retardants on the reproductive and thyroid system in adult male rats. Toxicol. Sci. Off. J. Soc. Toxicol. 2012, 127, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Norrgran, J.; Jones, B.; Lindquist, N.G.; Bergman, A. Decabromobiphenyl, polybrominated diphenyl ethers, and brominated phenolic compounds in serum of cats diagnosed with the endocrine disease feline hyperthyroidism. Arch. Environ. Contam. Toxicol. 2012, 63, 161–168. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hormones | Ve | TBP (mg/kg) | T3 (100 μg/kg) | T4 (100 μg/kg) | |||
|---|---|---|---|---|---|---|---|
| - | 40 | 250 | - | TBP (250 mg/kg) | - | TBP (250 mg/kg) | |
| Free T3 (pg/mL) | 1.64 ± 0.16 | 1.33 ± 0.22 a | 0.93 ± 0.06 a | <0.3 a | <0.3 | 1.64 ± 0.24 | 0.73 ± 0.09 b |
| Free T4 (ng/dL) | 1.80 ± 0.24 | 1.21 ± 0.22 a | 1.30 ±0.23 a | 2.33 ± 0.27 a | 1.99 ± 0.11 b | 2.76 ± 0.24 a | 1.62 ± 0.17 b |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Ahn, C.; Hong, E.-J.; An, B.-S.; Hyun, S.-H.; Choi, K.-C.; Jeung, E.-B. 2,4,6-Tribromophenol Interferes with the Thyroid Hormone System by Regulating Thyroid Hormones and the Responsible Genes in Mice. Int. J. Environ. Res. Public Health 2016, 13, 697. https://doi.org/10.3390/ijerph13070697
Lee D, Ahn C, Hong E-J, An B-S, Hyun S-H, Choi K-C, Jeung E-B. 2,4,6-Tribromophenol Interferes with the Thyroid Hormone System by Regulating Thyroid Hormones and the Responsible Genes in Mice. International Journal of Environmental Research and Public Health. 2016; 13(7):697. https://doi.org/10.3390/ijerph13070697
Chicago/Turabian StyleLee, Dongoh, Changhwan Ahn, Eui-Ju Hong, Beum-Soo An, Sang-Hwan Hyun, Kyung-Chul Choi, and Eui-Bae Jeung. 2016. "2,4,6-Tribromophenol Interferes with the Thyroid Hormone System by Regulating Thyroid Hormones and the Responsible Genes in Mice" International Journal of Environmental Research and Public Health 13, no. 7: 697. https://doi.org/10.3390/ijerph13070697
APA StyleLee, D., Ahn, C., Hong, E.-J., An, B.-S., Hyun, S.-H., Choi, K.-C., & Jeung, E.-B. (2016). 2,4,6-Tribromophenol Interferes with the Thyroid Hormone System by Regulating Thyroid Hormones and the Responsible Genes in Mice. International Journal of Environmental Research and Public Health, 13(7), 697. https://doi.org/10.3390/ijerph13070697