
Comparison of Effect of Two-Hour Exposure to Forest and Urban Environments on Cytokine, Anti-Oxidant, and Stress Levels in Young Adults


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Subjects
2.3. Procedure
2.4. Psychological Indices
2.5. Physiological Indices
2.6. Statistical Analyses
3. Results
3.1. Effect of Exposure to Forest Environment on Stress Levels
3.2. Effect of Exposure to Forest Environment on Serum GPx and Pro-Inflammatory Cytokine Levels
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| GPx | glutathione peroxidase |
| IFN | interferon |
| IL | interleukin |
| NK | Natural killer |
| SRI-MF | stress response inventory-modified form |
| TNF | tumor necrosis factor |
References
- Canter, A.; Cluff, L.E.; Imboden, J.B. Hypertensive reactions to immunization inoculation and antecedent psychological vulnerability. J. Psychosom. Res. 1972, 16, 99–101. [Google Scholar] [CrossRef]
- Fischer, C.L.; Daniels, J.C.; Levin, W.C.; Kimzey, S.L.; Cobb, E.K.; Ritzmann, S.E. Effects of the space flight environment on man’s immune system. II—Lymphocyte counts and reactivity. Aerosp. Med. 1972, 43, 1122–1125. [Google Scholar]
- Lovallo, W.R. Stress and Health: Biological and Psychological Interactions, 2nd ed.; Sage Publications: Thousand Oaks, CA, USA, 2005; pp. 133–154. [Google Scholar]
- Reus, G.Z.; Fries, G.R.; Stertz, L.; Badawy, M.; Passos, I.C.; Barichello, T.; Kapczinski, F.; Quevedo, J. The Role of Inflammation and Microgial Activation in the Pathology of Psychiatric Disorders. Neuroscience 2015, 300, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Leboyer, M.; Soreca, I.; Scott, J.; Frye, M.; Henry, C.; Tamouza, R.; Kupfer, D.J. Can bipolar disorder be viewed as a multi-system inflammatory disease? J. Affect. Disord. 2012, 141, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Myint, A.M.; Kim, Y.K. Cytokine-serotonin interaction through IOO-A neurodegeneration hypothesis of depression. Med. Hypothesis 2003, 61, 519–525. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Harrison, N.A.; Brydon, L.; Walker, C.; Gray, M.A.; Steptoe, A.; Critchley, H.D. Inflammation causes mood changes through alterations in subgenual cingulate activity and mesolimbic connectivity. Biol. Psychiatry 2009, 66, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Eisenberger, N.I.; Berkman, E.T.; Inagaki, T.K.; Rameson, L.T.; Mashal, N.M.; Irwin, M.R. Inflammation-induced anhedonia: Endotoxin reduces ventral striatum responses to reward. Biol. Psychiatry 2010, 68, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Boche, D.; Perry, V.H.; Nicoll, J.A. Review: Activation patterns of microglia and their identification in the human brain. Neuropathol. Appl. Neurobiol. 2013, 39, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Barger, S.W.; Goodwin, M.E.; Porter, M.M.; Beggs, M.L. Glutamate release from activated microglia requires the oxidative burst and lipid peroxidation. J. Neurochem. 2007, 101, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Takaki, J.; Fujimori, K.; Miura, M.; Suzuki, T.; Sekino, Y.; Sato, K. l-glutamate released from activated microglia downregulates astrocytic l-glutamate transporter expression in neuroinflammation: The “collusion” hypothesis for increased extracellular l-glutamate concentration in neuroinflammation. J. Neuroinflamm. 2012, 9, 275. [Google Scholar] [CrossRef] [PubMed]
- Maiorini, C.; Khorooshi, R.; Ruffini, F.; Løbner, M.; Bergami, A.; Garzetti, L.; Martino, G.; Owens, T.; Furlan, R. Lentiviral-mediated administration of IL-25 in the CNS induces alternative activation of microglia. Gene Ther. 2013, 20, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Ekdahl, C.T. Microglial activation—Tuning and purging adult neurogenesis. Front Pharmacol. 2012, 3, 41. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Leak, R.K.; Shi, Y.; Suenaga, J.; Gao, Y.; Zheng, P.; Chen, J. Microglial and macrophage polarization—New prospects for brain repair. Nat. Rev. Neurol. 2015, 11, 56–64. [Google Scholar] [CrossRef] [PubMed]
- John, R.F. The functions of cytokines and their uses in toxicology. Int. J. Exp. Pathol. 2001, 82, 171–192. [Google Scholar]
- Na, K.S.; Jung, H.Y.; Kim, Y.K. The role of pro-inflammatory cytokines in the neuroinflammation and neurogenesis of schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 48, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Woodroofe, M.N. Cytokine production in the central nervous system. Neurology 1995, 45, 6–10. [Google Scholar] [CrossRef]
- Mills, G.C. Hemoglobin catabolism I. Glutathione peroxidase, and erythrocyte enzyme which protects hemoglobin from oxidative breakdown. J. Biol. Chem. 1957, 229, 189–197. [Google Scholar] [PubMed]
- Flohé, L.; Günzler, W.A.; Schock, H.H. Glutathione peroxidase: A selenoenzyme. FEBS Lett. 1973, 32, 132–134. [Google Scholar] [CrossRef]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Chu, F.F.; Doroshow, J.H.; Esworthy, E.S. Expression, characterization, and tissue distribution of a new cellular selenium dependent glutathione peroxidase, GSH-Px-GI. J. Biol. Chem. 1993, 268, 2571–2576. [Google Scholar] [PubMed]
- Takahashi, K.; Avissar, N.; Whitin, J.; Cohen, H. Purification and characterization of human plasma glutathione peroxidase: Aselenoglycoprotien distinct from the known cellular enzyme. Arch. Biochem. Biophys. 1987, 256, 677–686. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M.; Valente, M.; Ferri, L.; Gregolin, C. Purification from pig liver of a protein which protects liposomes and biomembranes from peroxidative degradation and exhibits glutathione peroxidase activity on phosphatidylcholine hydroperoxides. Biochim. Biophys. Acta 1982, 710, 197–211. [Google Scholar] [CrossRef]
- Loureiro, G.; Rabaca, M.A.; Blanco, B.; Andrade, S.; Chieira, C.; Pereira, C. Urban versus rural environment-any differences in aeroallergens sensitization in an allergic population of Cova ba Beira, Portugal? Eur. Ann. Allergy Clin. Immunol. 2005, 37, 187–193. [Google Scholar] [PubMed]
- Kawakami, K.; Kawamoto, M.; Nomura, M.; Otani, H.; Nabika, T.; Gonda, T. Effects of phytoncides on blood pressure under restraint stress in SHRSP. Clin. Exp. Parmacol. Physiol. 2004, 31 (Suppl. S2), 27–28. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Calderon, R.; Reed, W.; Palacios-Moreno, J.; Keefe, S.; Herritt, L.; Brooks, D.; Torres-Jardón, R.; Calderón-Garcidueñas, L. Urban air pollution produces up-regulation of myocardial inflammatory genes and dark chocolate provides cardioprotection. Exp. Toxicol. Pathol. 2012, 64, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Calfapietra, C.; Fares, S.; Manes, F.; Morani, A.; Sgrigna, G.; Loreto, F. Role of Biogenic Volatile Organic Compounds (BVOC) emitted by urban trees on ozone concentration in cities: A review. Environ. Pollut. 2013, 183, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.G.; Cho, M.G.; Kim, J.I. Effects of Phytoncide Aromatherapy on Stress, Symptoms of Stress and Heart Rate Variability among Nursing Students. J. Biol. Nurs. Sci. 2012, 14, 249–257. [Google Scholar] [CrossRef]
- Joung, D.W.; Song, C.R.; Ikei, H.; Okuda, T.; Igarashi, M.; Koizumi, H. Physiological and psychological effects of olfactory stimulation with D-Limonene. Adv. Hortic. Sci. 2014, 28, 90–94. [Google Scholar]
- Mao, G.X.; Guang, L.X.; Bao, C.Y.; Mei, C.Z.; Hua, H.Z.; Dong, L.Y.; Zhen, W.Y.; Lian, H.X.; Fu, W.G.; Jing, Y. Effects of Short-Term Forest Bathing on Human Health in a Broad-Leaved Evergreen Forest in Zhenjian Province, China. Biomed. Environ. Sci. 2012, 25, 317–324. [Google Scholar] [PubMed]
- Woo, J.S.; Choi, S.R.; Lee, Y.K.; Min, Y.K.; Jung, B.H. Distribution Characteristics of Phytoncide (Monoterpene) by the Type of Recreational Forests in Gyeonggi-Do; Chungbuk Institute of Health and Environment: Cheongju, Korea, 2014. [Google Scholar]
- Ki, M.R. Theory and Practice of Case-Crossover Study Design. Korean J. Epidemiol. 2008, 30, 1–10. [Google Scholar] [CrossRef]
- Koh, K.B.; Park, J.K.; Kim, C.H. Development of the Stress Response Inventory. J. Korean Neuropsychiatr. Assoc. 2000, 39, 707–719. [Google Scholar]
- Choi, S.M.; Kang, T.Y.; Woo, J.M. Development and Validation of a Modified form of the Stress Response Inventory for Workers. J. Korean Neuropsychiatr. Assoc. 2006, 45, 541–553. [Google Scholar]
- Mao, G.X.; Cao, Y.B.; Lan, X.G.; He, Z.H.; Chen, Z.M.; Wang, Y.Z.; Hu, X.L.; Lv, Y.D.; Wang, G.F.; Yan, J. Therapeutic effect of forest bathing on human hypertension in the elderly. J. Cardiol. 2012, 60, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Sugama, S.; Fujita, M.; Conti, B. Stress induced morphological microglial activation in the rodent brain: Involvement of interleukin-18. Neuroscience 2007, 146, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Najjar, S.; Pearlman, D.M.; Alper, K.; Najjar, A.; Devinsky, O. Neuroinflammation and psychiatric illness. J. Neuroinflamm. 2013, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Schiepers, O.J.; Wichers, M.C.; Maes, M. Cytokines and major depression. Progr. Neuropsychopharmacol. Biol. Psychiatry 2005, 53, 23–34. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.M.; Scully, P.; Fitzgerald, P.; Scott, L.V.; Dinan, T.G. Plasma cytokine profiles in depressed patients who fail to respond to selective serotonin reuptake inhibitor therapy. J. Psychiatr. Res. 2007, 41, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Couch, Y.; Anthony, D.C.; Dolgov, O.; Revischin, A.; Festoff, B.; Santos, A.I.; Steinbusch, H.W.; Strekalova, T. Microglial activation, increased TNF and SERT expression in the prefrontal cortex define stress-altered behavior in mice susceptible to anhedonia. Brain Behav. Immun. 2013, 29, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.J.; Buckley, P.; Seabolt, W.; Mellor, A.; Kirkpatrick, B. Metaanalysis of cytokine alterations in schizophrenia: Clinical status and antipsychotic effects. Biol. Psychiatry 2011, 70, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Potvin, S.; Stip, E.; Sepehry, A.A.; Gendron, A.; Bah, R.; Kouassi, E. Inflammatory cytokine alteration in schizophrenia: A systematic quantitative review. Biol. Psychiatry 2008, 63, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Van Berkel, B.N.; Bossong, M.G.; Boelaard, R.; Kloet, R.; Schuitemaker, A.; Caspers, E.; Luurtsema, G.; Windhorst, A.D.; Cahn, W.; Lammertsma, A.A.; et al. Microglia activation in recent-onset schizophrenia: A quantitative (R)-[11C]PK11195 positron emission tomography study. Biol. Psychiatry 2008, 64, 820–822. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.H.; Andersson, P.B.; Gordon, S. Macrophages and inflammation in the central nervous system. Trends Neurosci. 1993, 16, 268–273. [Google Scholar] [CrossRef]
- Dheen, S.T.; Kaur, C.; Ling, E.A. Microglial activation and its implications in the brain diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.H.; Cunningham, C.; Holmes, C. Systemic infections and inflammation affect chronic neurodegeneration. Nat. Rev. Immunol. 2007, 7, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Ceretta, L.B.; Reus, G.Z.; Abelaira, H.M.; Jomada, L.K.; Schwalm, M.T.; Hoepers, N.J.; Tomazzi, C.D.; Gulbis, K.G.; Ceretta, R.A.; Quevedo, J. Increased prevalence of mood disorders and suicidal ideation in type 2 diabetic patients. Acta Diabetol. 2012, 49, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Abelaira, H.M.; Reus, G.Z.; Neotti, M.V.; Quevedo, J. The role of mTOR in depression and antidepressant responses. Life Sci. 2014, 101, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Abelaira, H.M.; Reus, G.Z.; Quevedo, J. Animal models as tools to study the pathophysiology of depression. Rev. Bras. Psiquiatr. 2014, 35, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Hoyo-Becerra, C.; Schlaak, J.F.; Hermann, D.M. Insights from interferon-alpha-related depression for the pathogenesis of depression associated with inflammation. Brain Behav. Immun. 2014, 42, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Iseme, R.A.; McEvoy, M.; Kelly, B.; Agnew, L.; Attia, J.; Walker, F.R. Autoantibodies and depression: Evidence for a causal link? Neurosci. Biobehav. Rev. 2014, 40, 62–79. [Google Scholar] [CrossRef] [PubMed]
- McInnis, O.A.; Matheson, K.; Anisman, H. Living with the unexplained: Coping, distress, and depression among women with chronic fatigue syndrome and/or fibromyalgia compared to an autoimmune disorder. Anx. Stress Cop. 2014, 27, 601–618. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Parameter | Mean (Standard Deviation) | p | ||
|---|---|---|---|---|
| Group A | Group B | |||
| Total sample number | 19 | 22 | ||
| Sex | Male | 7 | 7 | 0.735 |
| Female | 12 | 15 | ||
| Age (years) | 22.37(1.54) | 23.09(3.28) | 0.384 | |
| Height (cm) | 167.88(9.17) | 165.67(7.42) | 0.399 | |
| Weight (kg) | 67.74(15.72) | 60.51(11.85) | 0.102 | |
| Smoking | Yes/No | 3/16 | 5/17 | 0.576 |
| Drinking | Yes/No | 11/8 | 21/1 | 0.004 ** |
| Student loan | Yes/No | 6/13 | 5/16 | 0.583 |
| Part-time job | Yes/No | 11/8 | 10/11 | 0.516 |
| Economic Status | Good | 3 | 4 | 0.760 |
| Fair | 14 | 17 | ||
| Poor | 2 | 1 | ||
| Schedule | Group A | Group B | ||
|---|---|---|---|---|
| Item | Location | Item | Location | |
| 09:30–10:30 | Description of study procedure and subject allocation (Seoul Paik Hospital) | |||
| 10:30–12:30 | Pre-test | Seoul Paik Hospital (urban environment) | Pre-test | Pine Fragrant Forest (forest environment) |
| 12:30–14:30 | Wash-out period | Hwado-eup (rural environment) | Wash-out period | Hwado-eup (rural environment) |
| 14:30–15:30 | Transportation | Transportation | ||
| 15:30–17:30 | Post-test | Gapyeong Pine Fragrant Forest (forest environment) | Post-test | Seoul Paik Hospital (urban environment) |
| Variable | Subscales | Mean (Standard Deviation) | t | p | |
|---|---|---|---|---|---|
| Urban | Forest | ||||
| SRI-MF | Somatic symptoms | 7.61 (6.02) | 5.02 (5.16) | 2.347 | 0.024 * |
| Depressive symptoms | 8.76 (5.73) | 6.02 (5.18) | 2.339 | 0.024 * | |
| Anger symptoms | 5.34 (4.57) | 4 (4.15) | 1.630 | 0.111 | |
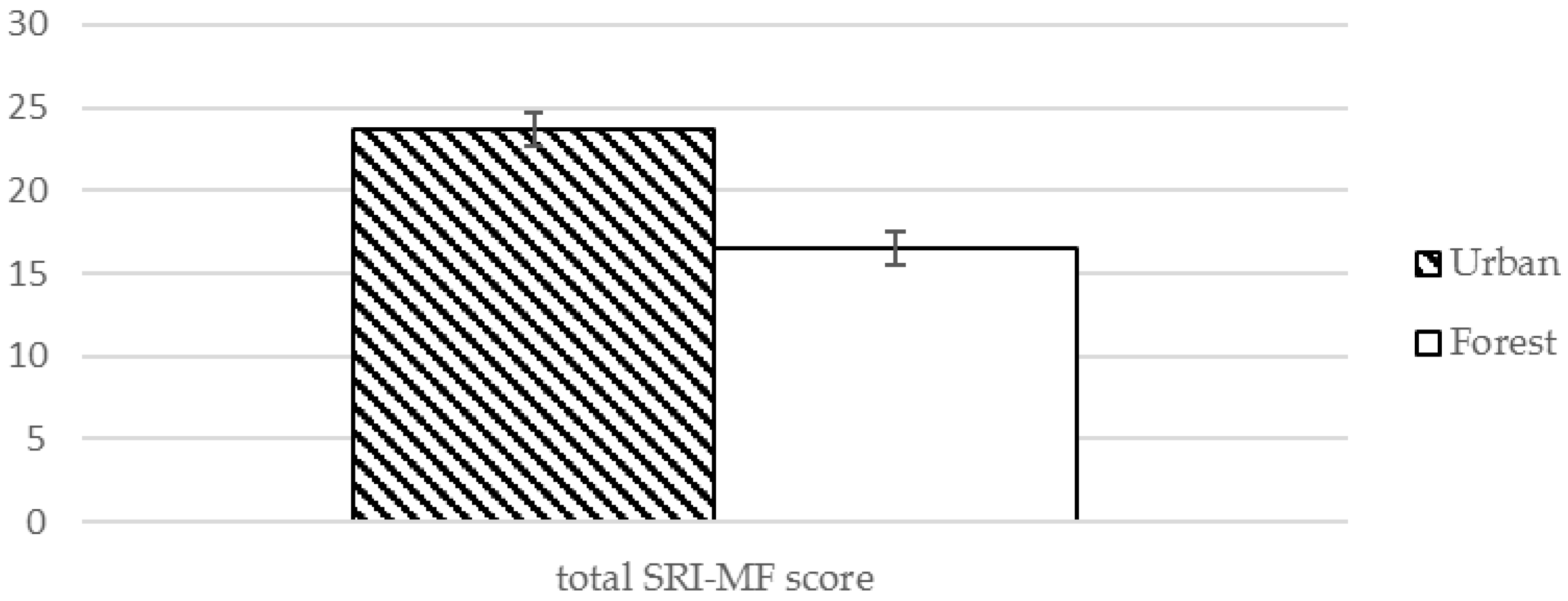
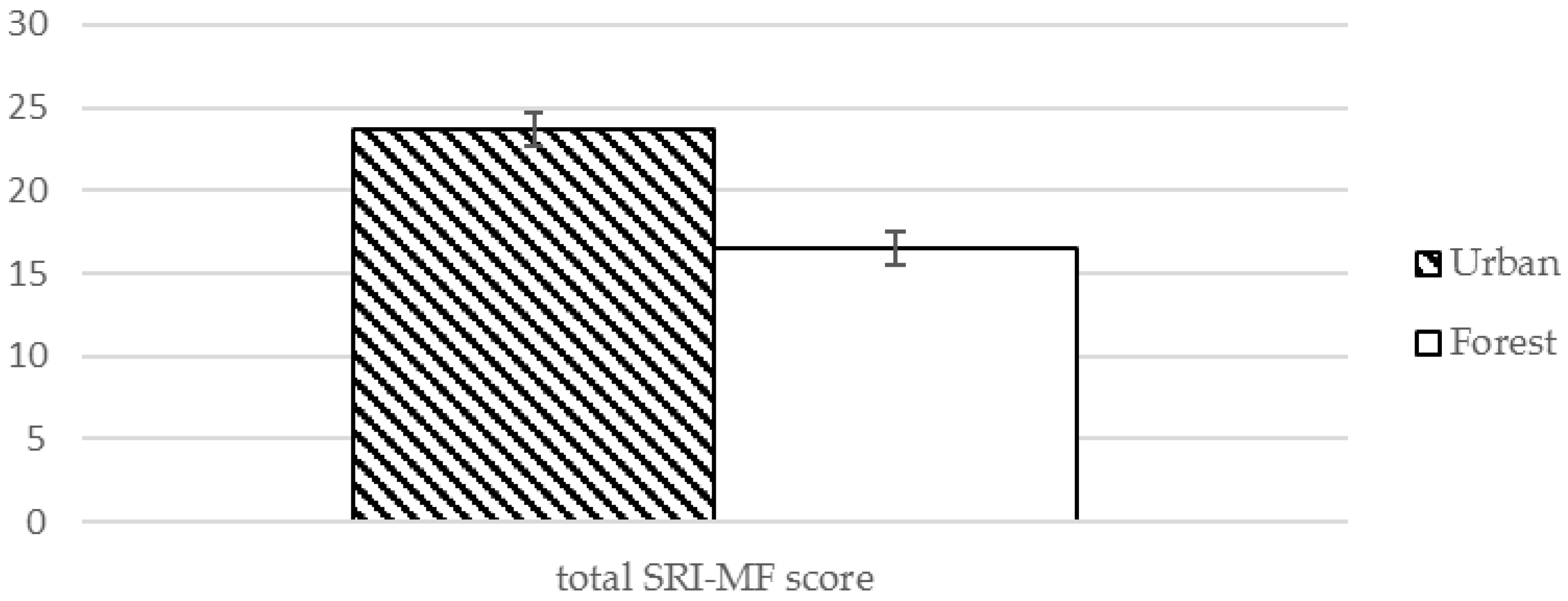
| Total | 23.68 (15.8) | 16.56 (13.38) | 2.351 | 0.024 * | |
| Variable | Mean (Standard Deviation) | t | p | |
|---|---|---|---|---|
| Urban | Forest | |||
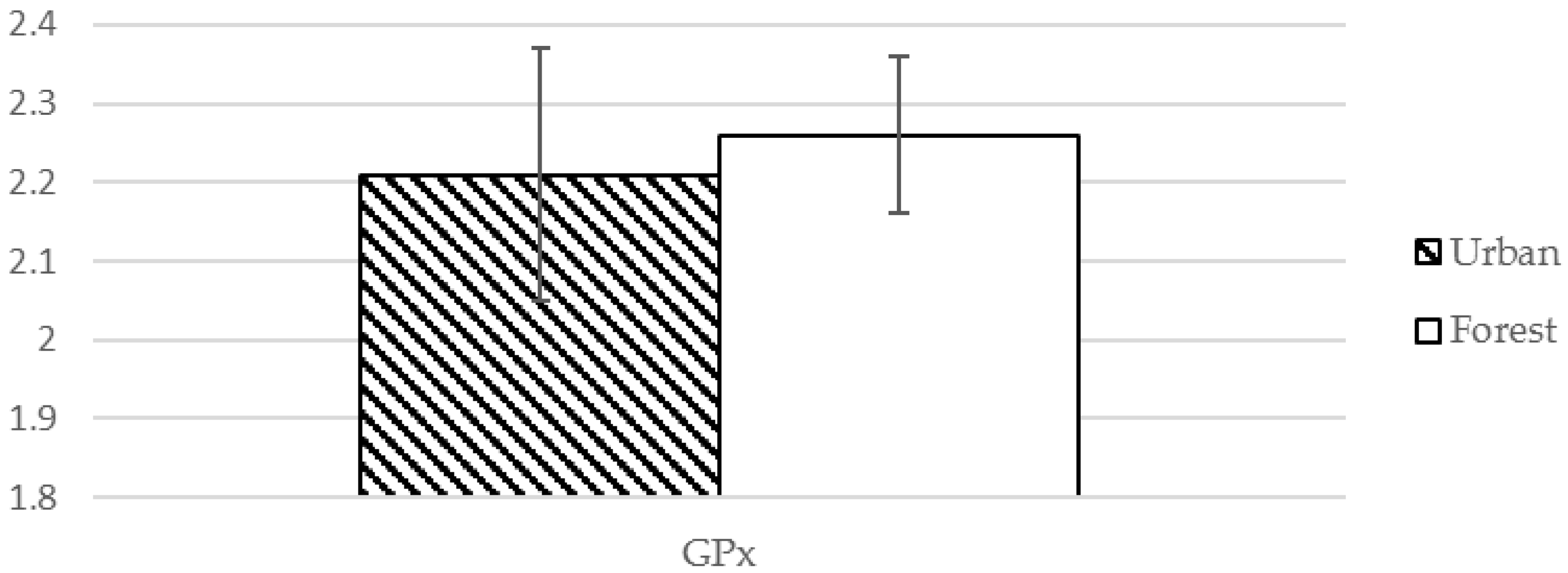
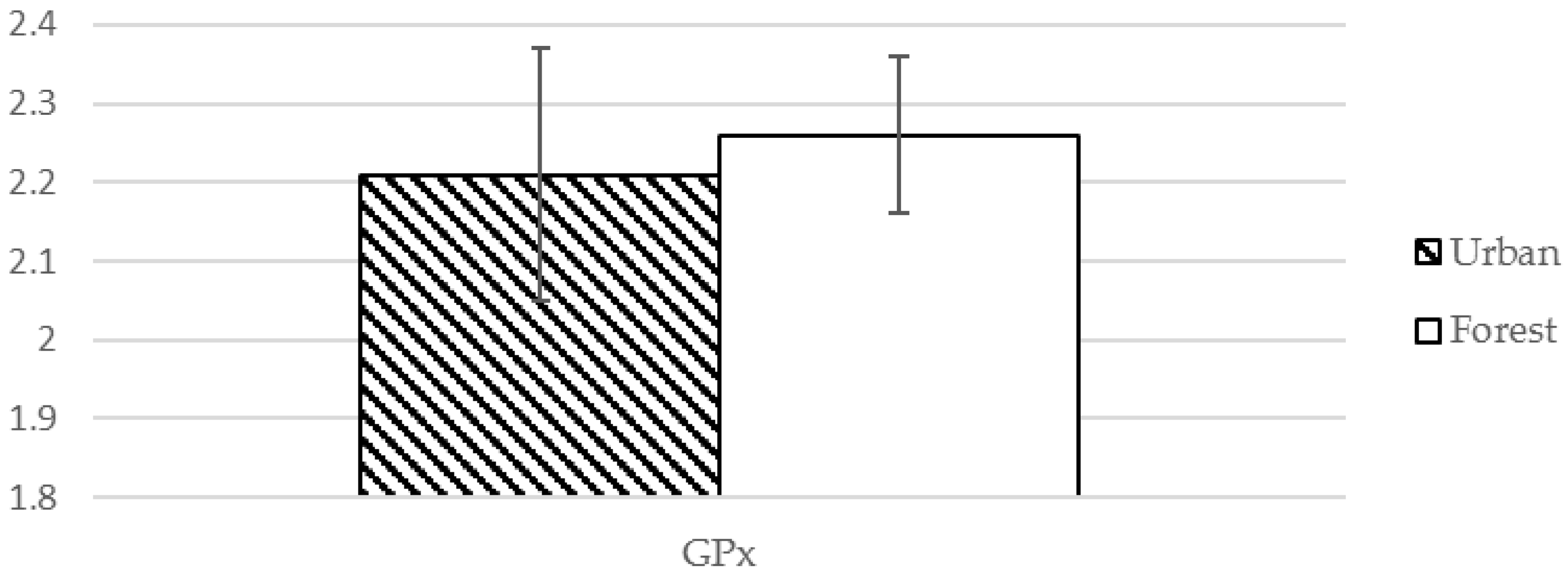
| GPx ❈ | 2.21 (0.16) µM | 2.26 (0.10) pg/mL | −2.039 | 0.048 * |
| IL-6 ❈ | −0.18 (0.25) pg/mL | −0.15 (0.23) pg/mL | −1.089 | 0.283 |
| IL-8 ❈ | 10.76 (2.87) pg/mL | 9.21 (2.72) pg/mL | 4.559 | 0.001 *** |
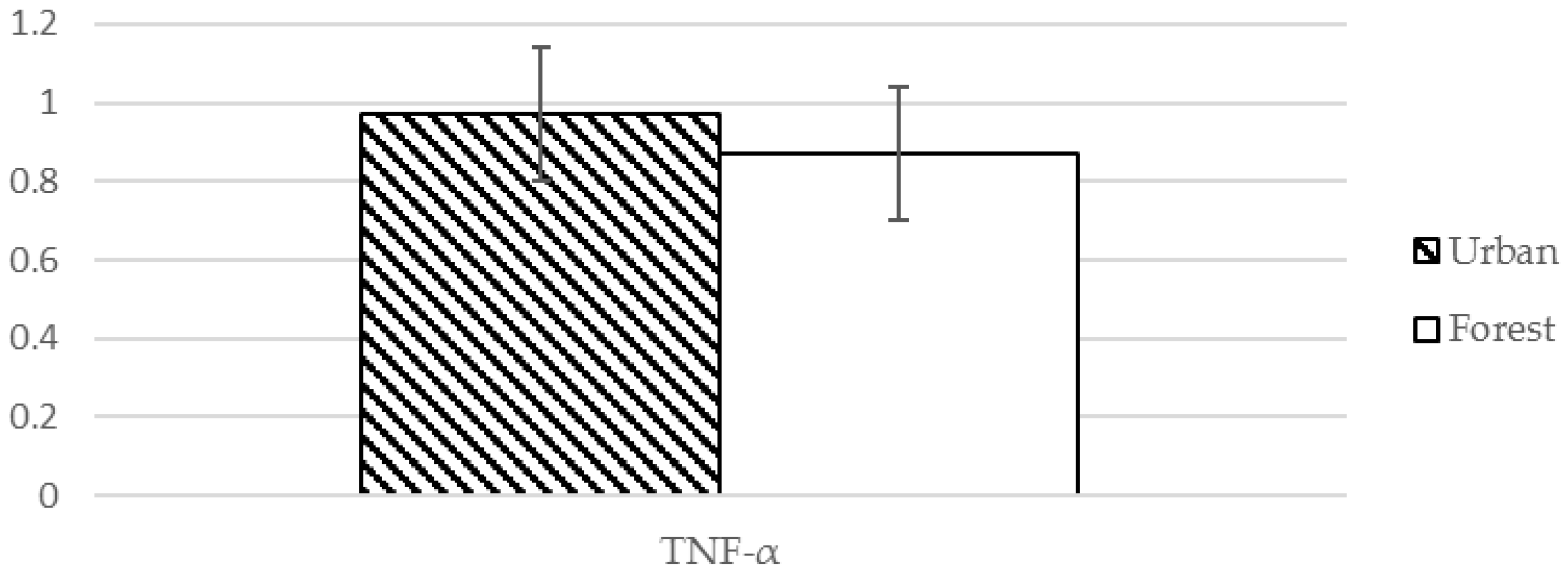
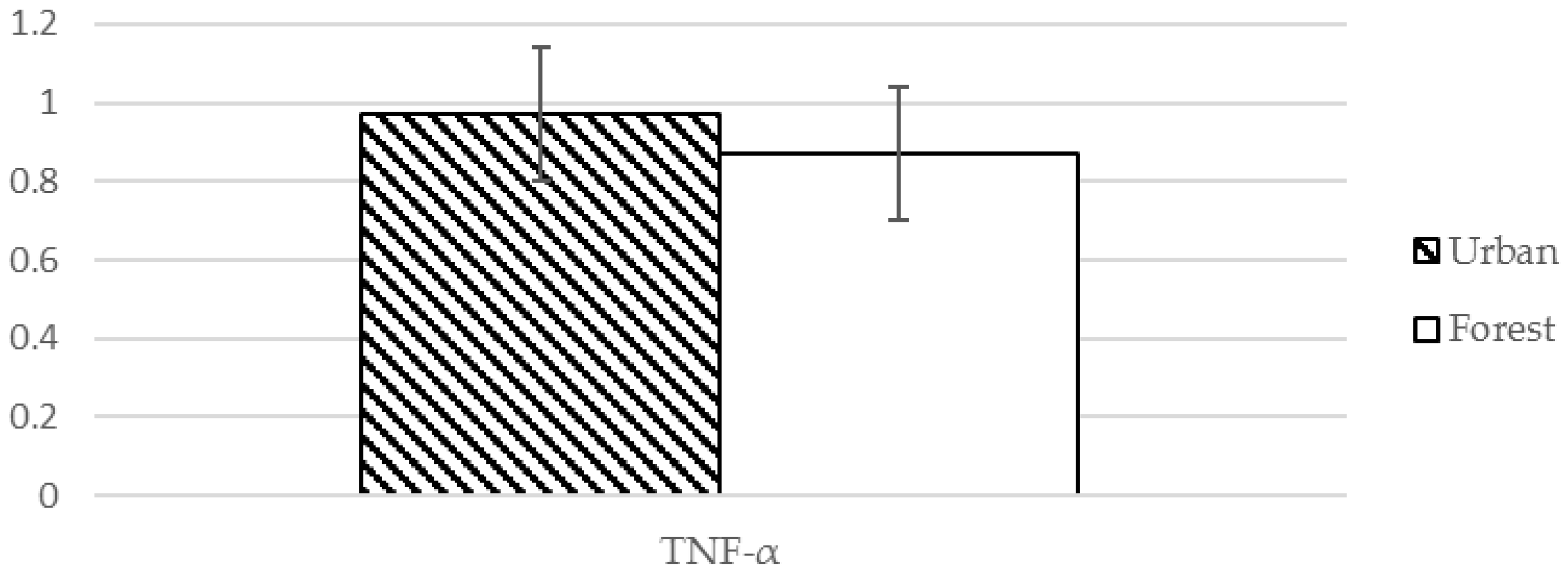
| TNF-α ❈ | 0.97 (0.17) pg/mL | 0.87 (0.17) pg/mL | 4.130 | 0.001 *** |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Im, S.G.; Choi, H.; Jeon, Y.-H.; Song, M.-K.; Kim, W.; Woo, J.-M. Comparison of Effect of Two-Hour Exposure to Forest and Urban Environments on Cytokine, Anti-Oxidant, and Stress Levels in Young Adults. Int. J. Environ. Res. Public Health 2016, 13, 625. https://doi.org/10.3390/ijerph13070625
Im SG, Choi H, Jeon Y-H, Song M-K, Kim W, Woo J-M. Comparison of Effect of Two-Hour Exposure to Forest and Urban Environments on Cytokine, Anti-Oxidant, and Stress Levels in Young Adults. International Journal of Environmental Research and Public Health. 2016; 13(7):625. https://doi.org/10.3390/ijerph13070625
Chicago/Turabian StyleIm, Su Geun, Han Choi, Yo-Han Jeon, Min-Kyu Song, Won Kim, and Jong-Min Woo. 2016. "Comparison of Effect of Two-Hour Exposure to Forest and Urban Environments on Cytokine, Anti-Oxidant, and Stress Levels in Young Adults" International Journal of Environmental Research and Public Health 13, no. 7: 625. https://doi.org/10.3390/ijerph13070625
APA StyleIm, S. G., Choi, H., Jeon, Y.-H., Song, M.-K., Kim, W., & Woo, J.-M. (2016). Comparison of Effect of Two-Hour Exposure to Forest and Urban Environments on Cytokine, Anti-Oxidant, and Stress Levels in Young Adults. International Journal of Environmental Research and Public Health, 13(7), 625. https://doi.org/10.3390/ijerph13070625