
Pathway Analysis Revealed Potential Diverse Health Impacts of Flavonoids that Bind Estrogen Receptors



Abstract
:1. Introduction
2. Materials and Methods
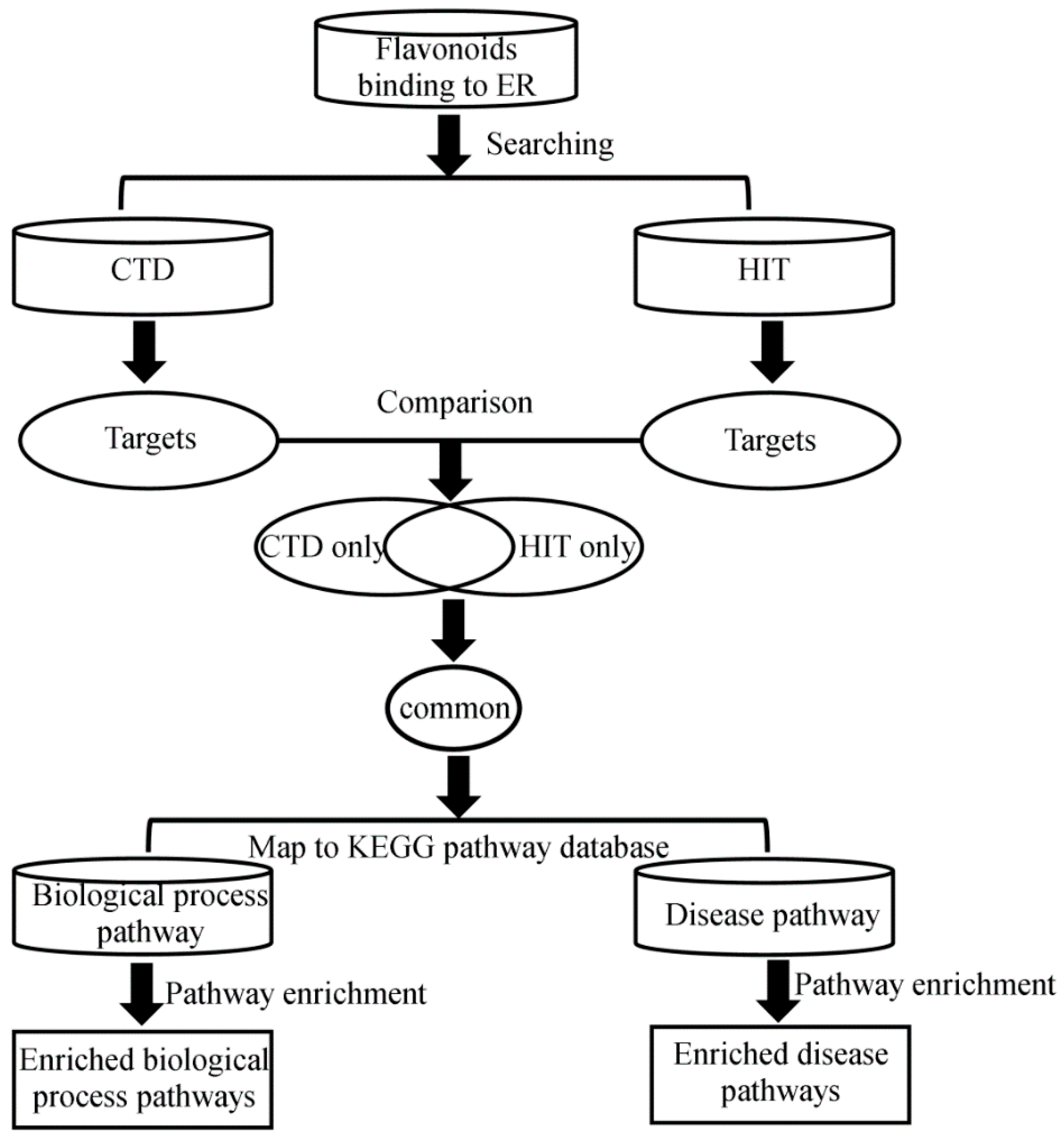
2.1. Study Design
2.2. Flavonoids with ER Binding Activity and Their Targets
2.2.1. Targets from CTD
2.2.2. Targets from HIT
2.3. Pathway Data
2.4. Pathway Enrichment Analysis
3. Results
3.1. Common Targets
3.2. Biological Process Pathways of the Common Targets
3.2.1. ER Regulated Processes
3.2.2. Estrogen Metabolism and Synthesis
3.2.3. Cellular Apoptosis
3.3. Disease Pathways of the Common Targets
3.3.1. Cancer
3.3.2. Infectious Diseases
3.3.3. Neurodegenerative Diseases
3.3.4. Cardiovascular Diseases
3.3.5. Endocrine and Metabolic Diseases
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABC | ATP-Binding Cassette |
| ALS | Amyotrophic Lateral Sclerosis |
| CTD | Comparative Toxicogenomics Database |
| EADB | Estrogenic Activity Database |
| EDSP | Endocrine Disruptor Screening Program |
| EMA | European Medicines Agency |
| EPA | Environmental Protection Agency |
| ER | Estrogen Receptor |
| FDA | Food and Drug Administration |
| HIT | Herbal Ingredients’ Target |
| NAFLD | Non-alcoholic Fatty Liver Disease |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| EGCG | (-)-Epigallocatechin-3-Gallate |
References
- Ververidis, F.; Trantas, E.; Douglas, C.; Vollmer, G.; Kretzschmar, G.; Panopoulos, N. Biotechnology of flavonoids and other phenylpropanoid-derived natural products. Part I: Chemical diversity, impacts on plant biology and human health. Biotechnol. J. 2007, 2, 1214. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.; Szmitko, B.S.V. Cardiology patient page red wine and your heart. Circulation 2005, 111–119. [Google Scholar]
- Hui, C.; Qi, X.; Qianyong, Z.; Xiaoli, P.; Jundong, Z.; Mantian, M. Flavonoids, flavonoid subclasses and breast cancer risk: A meta-analysis of epidemiologic studies. PLoS ONE 2013, 8, e54318. [Google Scholar] [CrossRef] [PubMed]
- Romagnolo, D.F.; Selmin, O.I. Flavonoids and cancer prevention: A review of the evidence. J. Nutr. Gerontol. Geriatr. 2012, 31, 206–238. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Pezzuto, J.M. Flavonoids in cancer prevention. Anti-Cancer Agents Med. Chem. 2012, 12, 836–851. [Google Scholar] [CrossRef]
- Choudhary, R.K.; Swarnkar, P.L. Antioxidant activity of phenolic and flavonoid compounds in some medicinal plants of India. Nat. Prod. Res. 2011, 25, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; De, B.; Devanna, N.; Chakraborty, R. Total phenolic, total flavonoid content, and antioxidant capacity of the leaves of meyna spinosa roxb., an Indian medicinal plant. Chin. J. Nat. Med. 2013, 11, 149–157. [Google Scholar] [CrossRef]
- Rangel-Huerta, O.D.; Pastor-Villaescusa, B.; Aguilera, C.M.; Gil, A. A systematic review of the efficacy of bioactive compounds in cardiovascular disease: Phenolic compounds. Nutrients 2015, 7, 5177–5216. [Google Scholar] [CrossRef] [PubMed]
- McCullough, M.L.; Peterson, J.J.; Patel, R.; Jacques, P.F.; Shah, R.; Dwyer, J.T. Flavonoid intake and cardiovascular disease mortality in a prospective cohort of U.S. adults. Am. J. Clin. Nutr. 2012, 95, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Hurt, R.T.; Wilson, T. Geriatric obesity: Evaluating the evidence for the use of flavonoids to promote weight loss. J. Nutr. Gerontol. Geriatr. 2012, 31, 269–289. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P. Flavonoids, cognition, and dementia: Actions, mechanisms, and potential therapeutic utility for alzheimer disease. Free Radic. Biol. Med. 2012, 52, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Zhang, N.; Wang, Y.Q.; Wang, J.Z.; Ji, S.X.; Dang, W.J.; Li, S.M.; Feng, L. Estrogenic effects of flavonoid components in xiaoyao powder. Genet. Mol. Res. GMR 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Resende, F.A.; de Oliveira, A.P.; de Camargo, M.S.; Vilegas, W.; Varanda, E.A. Evaluation of estrogenic potential of flavonoids using a recombinant yeast strain and mcf7/bus cell proliferation assay. PLoS ONE 2013, 8, e74881. [Google Scholar] [CrossRef] [PubMed]
- Dietary Supplements. Available online: http://www.fda.gov/Food/DietarySupplements/ (accessed on 11 February 2016).
- Quercetin. Available online: https://examine.com/supplements/quercetin/ (accessed on 11 February 2016).
- Soy Isoflavones. Available online: https://examine.com/supplements/soy-isoflavones/ (accessed on 11 February 2016).
- Brazier, N.C.; Levine, M.A. Understanding drug–herb interactions. Pharmacoepidemiol. Drug Saf. 2003, 12, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.E.; Zhang, S. Flavonoid-drug interactions: Effects of flavonoids on abc transporters. Life Sci. 2006, 78, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Paxton, J.W. The effects of flavonoids on the abc transporters: Consequences for the pharmacokinetics of substrate drugs. Expert Opin. Drug Metab. Toxicol. 2013, 9, 267–285. [Google Scholar] [CrossRef] [PubMed]
- Bircsak, K.M.; Aleksunes, L.M. Interaction of isoflavones with the bcrp/abcg2 drug transporter. Curr. Drug Metab. 2015, 16, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Power, D.; Llewellyn, L.; Faustino, M.; Nowell, M.A.; Björnsson, B.T.; Einarsdottir, I.; Canario, A.V.; Sweeney, G.E. Thyroid hormones in growth and development of fish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 447–459. [Google Scholar] [CrossRef]
- Gilbert, L.I.; King, D.S. Physiology of growth and development: Endocrine aspects. Physiol. Insecta 1973. [Google Scholar] [CrossRef]
- Ojamaa, K.; Klemperer, J.D.; Klein, I. Acute effects of thyroid hormone on vascular smooth muscle. Thyroid 1996, 6, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, F.; Den Tonkelaar, E.; Van Logten, M. Toxicity of sodium bromide in rats: Effects on endocrine system and reproduction. Food Chem. Toxicol. 1983, 21, 383–389. [Google Scholar] [CrossRef]
- Endocrine Disruption. Available online: http://www.epa.gov/endocrine-disruption (accessed on 11 February 2016).
- Lee, H.-R.; Kim, T.-H.; Choi, K.-C. Functions and physiological roles of two types of estrogen receptors, erα and erβ, identified by estrogen receptor knockout mouse. Lab. Anim. Res. 2012, 28, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Swedenborg, E.; Power, K.A.; Cai, W.; Pongratz, I.; Rüegg, J. Regulation of estrogen receptor beta activity and implications in health and disease. Cell. Mol. Life Sci. 2009, 66, 3873–3894. [Google Scholar] [CrossRef] [PubMed]
- Wise, P.M.; Dubal, D.B.; Wilson, M.E.; Rau, S.W.; Böttner, M. Minireview: Neuroprotective effects of estrogen—New insights into mechanisms of action. Endocrinology 2001, 142, 969–973. [Google Scholar] [PubMed]
- Garcia-Segura, L.M.; Azcoitia, I.; DonCarlos, L.L. Neuroprotection by estradiol. Prog. Neurobiol. 2001, 63, 29–60. [Google Scholar] [CrossRef]
- Palmieri, C.; Cheng, G.; Saji, S.; Zelada-Hedman, M.; Weihua, Z.; Van Noorden, S.; Wahlstrom, T.; Coombes, R.; Warner, M.; Gustafsson, J. Estrogen receptor beta in breast cancer. Endocr. Relat. Cancer 2002, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.M.; Korach, K.S. Stromal cell-derived factor 1, a novel target of estrogen receptor action, mediates the mitogenic effects of estradiol in ovarian and breast cancer cells. Mol. Endocrinol. 2003, 17, 792–803. [Google Scholar] [CrossRef] [PubMed]
- Pare, G.; Krust, A.; Karas, R.H.; Dupont, S.; Aronovitz, M.; Chambon, P.; Mendelsohn, M.E. Estrogen receptor-α mediates the protective effects of estrogen against vascular injury. Circ. Res. 2002, 90, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E. Estrogen signaling and cardiovascular disease. Circ. Res. 2011, 109, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, C.; Hellberg, N.; Parini, P.; Vidal, O.; Bohlooly, M.; Rudling, M.; Lindberg, M.K.; Warner, M.; Angelin, B.; Gustafsson, J.-Å. Obesity and disturbed lipoprotein profile in estrogen receptor-α-deficient male mice. Biochem. Biophys. Res. Commun. 2000, 278, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Haas, E.; Bhattacharya, I.; Brailoiu, E.; Damjanović, M.; Brailoiu, G.C.; Gao, X.; Mueller-Guerre, L.; Marjon, N.A.; Gut, A.; Minotti, R. Regulatory role of g protein-coupled estrogen receptor for vascular function and obesity. Circ. Res. 2009, 104, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Gennari, L.; Merlotti, D.; Valleggi, F.; Martini, G.; Nuti, R. Selective estrogen receptor modulators for postmenopausal osteoporosis. Drugs Aging 2007, 24, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Xu, L.; Fang, H.; Richard, A.M.; Bray, J.D.; Judson, R.S.; Zhou, G.; Colatsky, T.J.; Aungst, J.L.; Teng, C. Eadb: An estrogenic activity database for assessing potential endocrine activity. Toxicol. Sci. 2013, 135, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Branham, W.S.; Ng, H.W.; Moland, C.L.; Dial, S.L.; Fang, H.; Perkins, R.; Sheehan, D.; Tong, W. Human sex hormone-binding globulin binding affinities of 125 structurally diverse chemicals and comparison with their binding to androgen receptor, estrogen receptor, and α-fetoprotein. Toxicol. Sci. 2015, 143, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Mattingly, C.; Rosenstein, M.; Colby, G.; Forrest, J., Jr.; Boyer, J. The comparative toxicogenomics database (ctd): A resource for comparative toxicological studies. J. Exp. Zool. Part A Comp. Exp. Biol. 2006, 305, 689. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Ye, L.; Kang, H.; Zhang, D.; Tao, L.; Tang, K.; Liu, X.; Zhu, R.; Liu, Q.; Chen, Y. Hit: Linking herbal active ingredients to targets. Nucleic Acids Res. 2011, 39, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. Kegg: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Khatri, P.; Sirota, M.; Butte, A.J. Ten years of pathway analysis: Current approaches and outstanding challenges. PLoS Comput. Biol. 2012, 8, e1002375. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Tang, K.; Yang, L.; Cao, Z.; Li, Y. Study of drug function based on similarity of pathway fingerprint. Protein Cell 2012, 3, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, M.; Rendon, A.; Ouwehand, W.H.; Wernisch, L.; Dudbridge, F. Comparison of methods for competitive tests of pathway analysis. PLoS ONE 2012, 7, e41018. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Cao, X.; Harris, S.; Sun, H.; Fang, H.; Fuscoe, J.; Harris, A.; Hong, H.; Xie, Q.; Perkins, R.; Shi, L.; Casciano, D. ArrayTrack—Supporting toxicogenomic research at the U.S.food and drug administration national center for toxicological research. Environ. Health Perspect. 2003, 111, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Yang, L.; Cao, Z.; Tang, K.; Li, Y. A pathway profile-based method for drug repositioning. Chin. Sci. Bull. 2012, 57, 2106–2112. [Google Scholar] [CrossRef]
- Watters, J.J.; Chun, T.Y.; Kim, Y.N.; Bertics, P.J.; Gorski, J. Estrogen modulation of prolactin gene expression requires an intact mitogen-activated protein kinase signal transduction pathway in cultured rat pituitary cells. Mol. Endocrinol. 2000, 14, 1872–1881. [Google Scholar] [CrossRef] [PubMed]
- Chemical-Gene Interaction Query. Available online: http://ctdbase.org/query.go?type=ixn (accessed on 22 October 2015).
- HIT: Herbal Ingredients' Targets Database. Available online: http://lifecenter.sgst.cn/hit (accessed on 22 October 2015).
- Hiroi, R.; Handa, R.J. Estrogen receptor-β regulates human tryptophan hydroxylase-2 through an estrogen response element in the 5' untranslated region. J. Neurochem. 2013, 127, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Semak, I.; Pisarchik, A.; Sweatman, T.; Szczesniewski, A.; Wortsman, J. Conversion of l-tryptophan to serotonin and melatonin in human melanoma cells. FEBS Lett. 2002, 511, 102–106. [Google Scholar] [CrossRef]
- Altun, A.; Ugur-Altun, B. Melatonin: Therapeutic and clinical utilization. Int. J. Clin. Pract. 2007, 61, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.M.; Wood, D.M.; Bottomley, L.; Blagg, S.; Owen, K.; Hughes, P.J.; Waring, R.H.; Kirk, C.J. Phytoestrogens are potent inhibitors of estrogen sulfation: Implications for breast cancer risk and treatment. J. Clin. Endocrinol. Metab. 2004, 89, 1779–1787. [Google Scholar] [CrossRef] [PubMed]
- Häggström, M. Diagram of the pathways of human steroidogenesis. Medicine 2014, 1, 1. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Nakajima, M.; Yokoi, T. Cytochrome p450-mediated metabolism of estrogens and its regulation in human. Cancer Lett. 2005, 227, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Jordan, V.C. The new biology of estrogen-induced apoptosis applied to treat and prevent breast cancer. Endocr Relat Cancer 2015, 22, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Wambi, J.S.; Jordan, V.C. Estrogen regulation of apoptosis: How can one hormone stimulate and inhibit. Breast Cancer Res 2009, 11, 206. [Google Scholar] [CrossRef] [PubMed]
- Jordan, V.C. Tamoxifen (ici46, 474) as a targeted therapy to treat and prevent breast cancer. Brit. J. Pharmacol. 2006, 147, S269–S276. [Google Scholar] [CrossRef] [PubMed]
- Rath, P.C.; Aggarwal, B.B. Tnf-induced signaling in apoptosis. J. Clin. Immunol. 1999, 19, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Gori, I.; Pellegrini, C.; Staedler, D.; Russell, R.; Jan, C.; Canny, G.O. Tumor necrosis factor-alpha activates estrogen signaling pathways in endometrial epithelial cells via estrogen receptor alpha. Mol. Cell. Endocrinol. 2011, 345, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Mursu, J.; Nurmi, T.; Tuomainen, T.P.; Salonen, J.T.; Pukkala, E.; Voutilainen, S. Intake of flavonoids and risk of cancer in finnish men: The kuopio ischaemic heart disease risk factor study. Int. J. Cancer 2008, 123, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Cutler, G.J.; Nettleton, J.A.; Ross, J.A.; Harnack, L.J.; Jacobs, D.R.; Scrafford, C.G.; Barraj, L.M.; Mink, P.J.; Robien, K. Dietary flavonoid intake and risk of cancer in postmenopausal women: The Iowa women's health study. Int. J. Cancer 2008, 123, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.H.; Song, H.Y.; Zhou, Y.F.; Yuan, G.Y.; Zheng, F.J. Effects of quercetin on the proliferation of breast cancer cells and expression of survivin. Exp. Ther. Med. 2013, 6, 1155–1158. [Google Scholar] [PubMed]
- Yokoyama, S.; Hirano, H.; Wakimaru, N.; Sarker, K.P.; Kuratsu, J. Inhibitory effect of epigallocatechin- gallate on brain tumor cell lines in vitro. Neuro-Oncology 2001, 3, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Touillaud, M.; Rothwell, J.A.; Romieu, I.; Scalbert, A. Measuring exposure to the polyphenol metabolome in observational epidemiologic studies: Current tools and applications and their limits. Am. J. Clin. Nutr. 2014, 100, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Hanski, L.; Genina, N.; Uvell, H.; Malinovskaja, K.; Gylfe, A.; Laaksonen, T.; Kolakovic, R.; Makila, E.; Salonen, J.; Hirvonen, J.; et al. Inhibitory activity of the isoflavone biochanin a on intracellular bacteria of genus chlamydia and initial development of a buccal formulation. PLoS ONE 2014, 9, e115115. [Google Scholar] [CrossRef] [PubMed]
- das Neves, M.V.; da Silva, T.M.; Lima Ede, O.; da Cunha, E.V.; Oliveira Ede, J. Isoflavone formononetin from red propolis acts as a fungicide against candida sp. Braz. J. Microbiol. 2016, 47, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, M.; Hirata, K.; Kunikata, T.; Ushio, S.; Iwaki, K.; Ohashi, K.; Ikeda, M.; Kurimoto, M. Antibacterial action of tryptanthrin and kaempferol, isolated from the indigo plant (polygonum tinctorium lour.), against helicobacter pylori-infected mongolian gerbils. J. Gastroenterol. 2001, 36, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Frakes, A.E.; Ferraiuolo, L.; Haidet-Phillips, A.M.; Schmelzer, L.; Braun, L.; Miranda, C.J.; Ladner, K.J.; Bevan, A.K.; Foust, K.D.; Godbout, J.P. Microglia induce motor neuron death via the classical nf-κb pathway in amyotrophic lateral sclerosis. Neuron 2014, 81, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Haidet-Phillips, A.M.; Hester, M.E.; Miranda, C.J.; Meyer, K.; Braun, L.; Frakes, A.; Song, S.; Likhite, S.; Murtha, M.J.; Foust, K.D. Astrocytes from familial and sporadic als patients are toxic to motor neurons. Nat. Biotechnol. 2011, 29, 824–828. [Google Scholar] [CrossRef] [PubMed]
- Rabassa, M.; Cherubini, A.; Zamora-Ros, R.; Urpi-Sarda, M.; Bandinelli, S.; Ferrucci, L.; Andres-Lacueva, C. Low levels of a urinary biomarker of dietary polyphenol are associated with substantial cognitive decline over a 3-year period in older adults: The invecchiare in chianti study. J. Am. Geriatr. Soc. 2015, 63, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Venigalla, M.; Gyengesi, E.; Munch, G. Curcumin and apigenin-novel and promising therapeutics against chronic neuroinflammation in alzheimer’s disease. Neural Regen. Res. 2015, 10, 1181–1185. [Google Scholar] [PubMed]
- Cassidy, A.; Mukamal, K.J.; Liu, L.; Franz, M.; Eliassen, A.H.; Rimm, E.B. High anthocyanin intake is associated with a reduced risk of myocardial infarction in young and middle-aged women. Circulation 2013, 127, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Yajima, T.; Knowlton, K.U. Viral myocarditis from the perspective of the virus. Circulation 2009, 119, 2615–2624. [Google Scholar] [CrossRef] [PubMed]
- Utzschneider, K.M.; Kahn, S.E. Review: The role of insulin resistance in nonalcoholic fatty liver disease. J. Clin. Endocrinol. Metab. 2006, 91, 4753–4761. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Forouhi, N.G.; Sharp, S.J.; Gonzalez, C.A.; Buijsse, B.; Guevara, M.; van der Schouw, Y.T.; Amiano, P.; Boeing, H.; Bredsdorff, L.; et al. The association between dietary flavonoid and lignan intakes and incident type 2 diabetes in european populations: The epic-interact study. Diabetes Care 2013, 36, 3961–3970. [Google Scholar] [CrossRef] [PubMed]
- Stanley Mainzen Prince, P.; Kamalakkannan, N. Rutin improves glucose homeostasis in streptozotocin diabetic tissues by altering glycolytic and gluconeogenic enzymes. J. Biochem. Mol. Toxicol. 2006, 20, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yao, Y.; Wang, Y.; Ren, G. Antidiabetic activity of isoquercetin in diabetic kk-ay mice. Nutr. Metab. 2011, 8, 85. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.W.; Zhang, W.; Shu, M.; Luo, H.; Ge, W.; Perkins, R.; Tong, W.; Hong, H. Competitive molecular docking model for predicting estrogen receptor agonists and antagonists. BMC Bioinf. 2014, 15, S4. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Du, T.; Zhou, P.; Yang, L.; Mei, H.; Ng, H.W.; Zhang, W.; Shu, M.; Tong, W.; Shi, L.; et al. Molecular docking to identify associations between drugs and class I human leukocyte antigens for predicting potential idiosyncratic drug reactions. Comb. Chem. High Throughput Screen. 2015, 18, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, W.; Fang, H.; Perkins, R.; Tong, W.; Hong, H. Homology modeling, molecular docking, and molecular dynamics simulations elucidated alpha-fetoprotein binding modes. BMC Bioinform. 2013, 14, S6. [Google Scholar] [CrossRef] [PubMed]
- Shu, M.; Ng, H.W.; Orr, M.; Luo, H.; Ye, H.; Ge, W.; Tong, W.; Hong, H. Homology model and ligand binding interactions of the extracellular domain of the human α4β2 nicotinic acetylcholine receptor. J. Biomed. Sci. Eng. 2016, 9, 41–100. [Google Scholar]
- Luo, H.; Ye, H.; Ng, H.W.; Shi, L.; Tong, W.; Mendrick, D.L.; Hong, H. Machine learning methods for predicting HLA-peptide binding activity. Bioinform. Biol. Insights 2015, 9, 21–29. [Google Scholar] [PubMed]
- Hong, H.; Hong, Q.; Perkins, R.; Shi, L.; Fang, H.; Su, Z.; Dragan, Y.; Fuscoe, J.C.; Tong, W. The accurate prediction of protein family from amino acid sequence by measuring features of sequence fragments. J. Comput. Biol. 2009, 16, 1671–1688. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Ye, H.; Ng, H.W.; Shi, L.; Tong, W.; Mattes, W.; Mendrick, D.L.; Hong, H. Understanding and predicting binding between human leukocyte antigens and peptides by network analysis. BMC Bioinform. 2015, 16, S9. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Luo, H.; Ng, H.W.; Meehan, J.; Ge, W.; Tong, W.; Hong, H. Applying network analysis and Nebula (neighbor-edges based and unbiased leverage algorithm) to ToxCast data. Environ. Int. 2016, 88–89, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Yang, Z.; Yang, Z.; Zhou, S.; Zhang, H.; Zang, L. Effect of apigenin on proliferation and apoptosis of human lung cancer nci-h460 cells. J. South. Med. Univ. 2013, 33, 1137–1140. [Google Scholar]
- Cheng, Z.; Sun, G.; Guo, W.; Huang, Y.; Sun, W.; Zhao, F.; Hu, K. Inhibition of hepatitis b virus replication by quercetin in human hepatoma cell lines. Virol. Sin. 2015, 30, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, S.; Li, X.; Luo, G.; Li, L.; Le, W. Neuroprotective effects of (-)-epigallocatechin-3-gallate in a transgenic mouse model of amyotrophic lateral sclerosis. Neurochem. Res. 2006, 31, 1263–1269. [Google Scholar] [CrossRef] [PubMed]

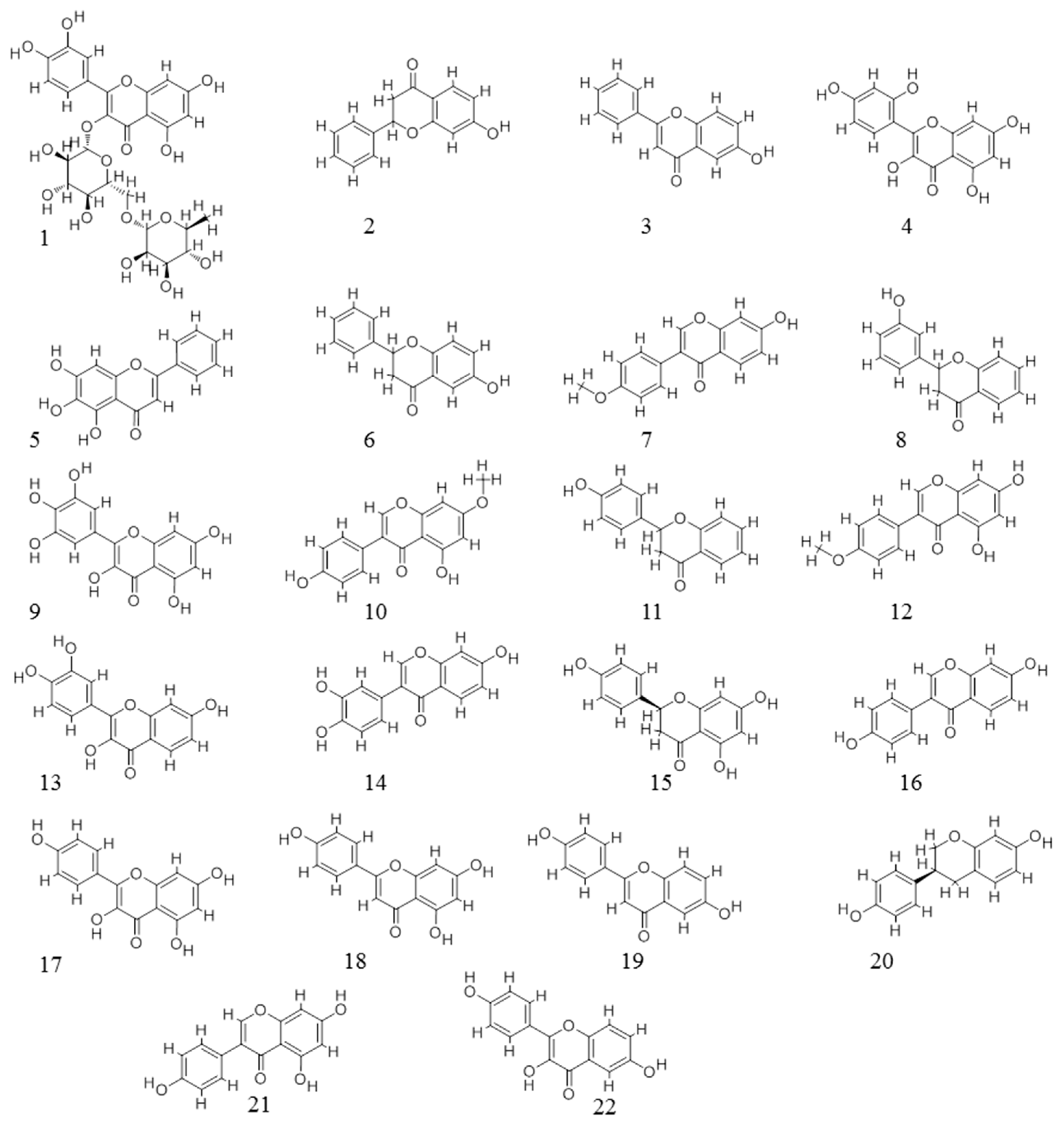

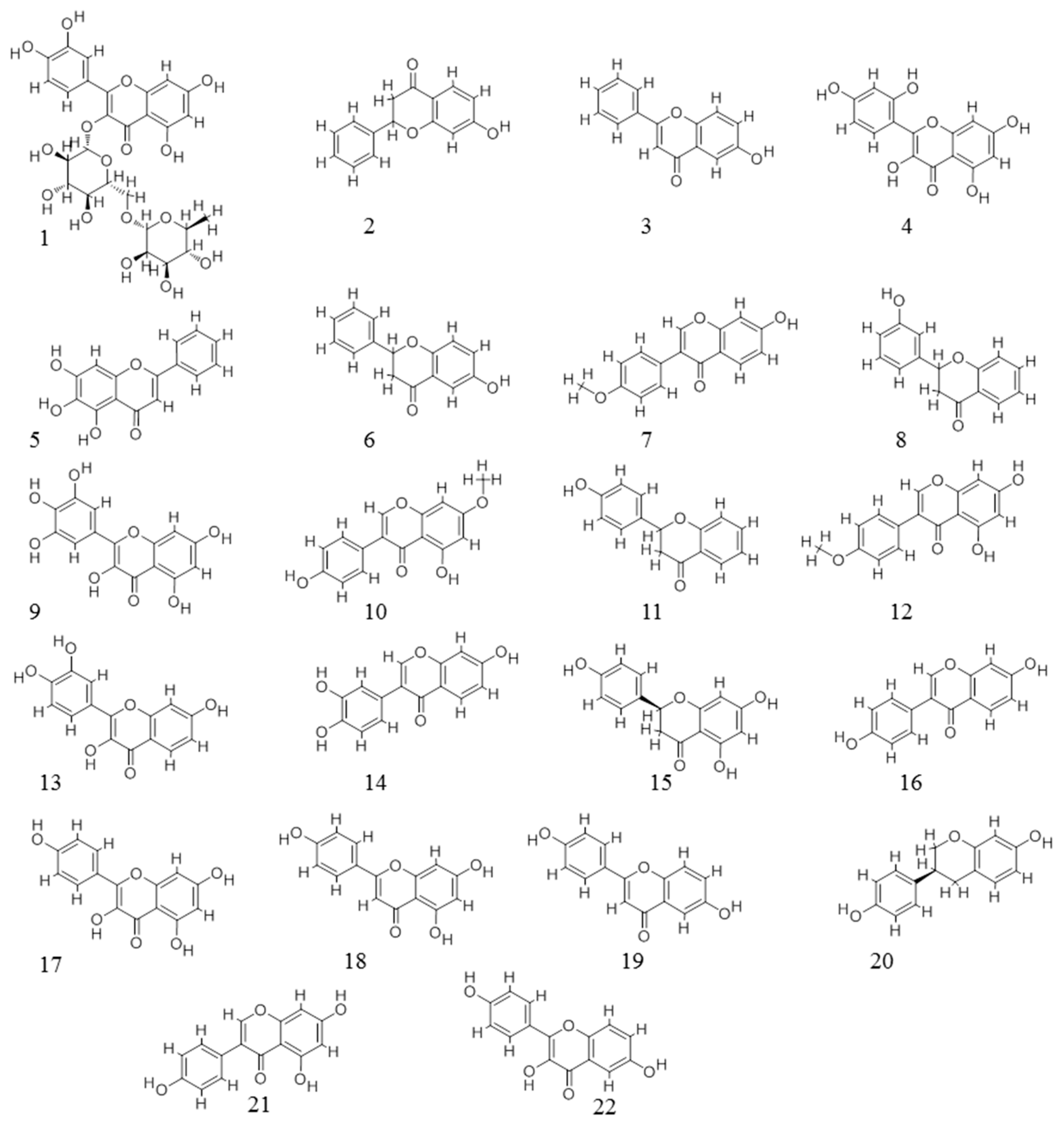
{kind=link}
{kind=link}
| ID | Name | CAS | ER Activity | Number of Targets | |
|---|---|---|---|---|---|
| HIT | CTD | ||||
| 1 | Rutin | 153-18-4 | −4.09 | 3 | 5 |
| 2 | 7-Hydroxyflavanone | 6515-36-2 | −3.73 | 0 | 2 |
| 3 | 6-Hydroxyflavone | 6665-83-4 | −3.41 | 0 | 1 |
| 4 | Morin | 480-16-0 | −3.09 | 4 | 1 |
| 5 | Baicalein | 491-67-8 | −3.05 | 4 | 4 |
| 6 | 6-Hydroxyflavanone | 4250-77-5 | −3.05 | 0 | 0 |
| 7 | Formononetin | 485-72-3 | −2.98 | 2 | 3 |
| 8 | 3-hydroxyflavanone | 92496-65-6 | −2.78 | 0 | 2 |
| 9 | Myricetin | 529-44-2 | −2.75 | 9 | 2 |
| 10 | Prunetin | 552-59-0 | −2.74 | 0 | 0 |
| 11 | 4-hydroxyflavanone | 6515-37-3 | −2.65 | 0 | 3 |
| 12 | Biochanin A | 491-80-5 | −2.37 | 0 | 12 |
| 13 | Fisetin | 528-48-3 | −2.35 | 8 | 2 |
| 14 | 3,4,7-Trihydroxyisoflavone isoflavone | 485-63-2 | −2.35 | 0 | 0 |
| 15 | Naringenin | 480-41-1 | −2.13 | 4 | 12 |
| 16 | Daidzein | 486-66-8 | −1.65 | 12 | 12 |
| 17 | Kaempferol | 520-18-3 | −1.61 | 12 | 20 |
| 18 | Apigenin | 520-36-5 | −1.55 | 15 | 16 |
| 19 | 4,6-Dihydroxyflavone | 63046-09-3 | −0.82 | 0 | 0 |
| 20 | Equol | 531-95-3 | −0.82 | 0 | 7 |
| 21 | Genistein | 446-72-0 | −0.36 | 16 | 37 |
| 22 | 3,6,4-Trihydroxyflavone | 253195-19-6 | −0.35 | 0 | 0 |
| Gene (Entrz ID) | Chemical IDs | Gene (Entrz ID) | Chemical IDs |
|---|---|---|---|
| ABCC2 (1244) | 21 | FOXO3 (2309) | 21 |
| AFP (174) | 2, 5, 8, 9, 11, 13, 15, 17, 18, 20 | GPER1 (2852) | 21 |
| AHR (196) | 5, 12, 15, 16, 17, 18, 21 | GREB1 (9687) | 17, 21 |
| AIP (9049) | 17, 18 | HIF1A (3091) | 21 |
| AKR1C1 (1645) | 15 | HIST3H3 (8290) | 21 |
| AKT1 (207) | 21 | JUN (3725) | 17, 18 |
| ALB (213) | 1, 7 | JUND (3727) | 21 |
| ALPL (249) | 15, 16, 21 | MAP2K1 (5604) | 9 |
| APP (351) | 4, 5, 17 | NCOA1 (8648) | 12, 21 |
| AR (367) | 3, 11, 13, 20, 21 | NCOA2 (10499) | 16, 21 |
| ARNT (405) | 15, 17, 18, 21 | NCOA3 (8202) | 17 |
| BAX (581) | 17 | NFKBIA (4792) | 18 |
| BBC3 (27113) | 17 | NFKBIB (4793) | 18 |
| BCL2L1 (598) | 17 | NOS3 (4846) | 20 |
| CASP3 (836) | 1 | NR1H3 (10062) | 21 |
| CASP8 (841) | 1 | NR1I2 (8856) | 16, 21 |
| CAV1 (857) | 20 | NR3C1 (2908) | 21 |
| CBR1 (873) | 1, 21 | NRIP1 (8204) | 21 |
| CCND1 (595) | 1 | PGR (5241) | 12, 17, 18 |
| CDK2 (1017) | 16 | PNRC1 (10957) | 12, 16, 21 |
| CDKN1A (1026) | 16 | PPARG (5468) | 17, 18 |
| CYLD (1540) | 21 | PTEN (5728) | 21 |
| CYP19A1 (1588) | 12, 15 | RORC (6097) | 12 |
| CYP1A1 (1543) | 12, 15, 17, 18 | RXRA (6256) | 21 |
| CYP1B1 (1545) | 5, 15, 17, 18, 21 | SHBG (6462) | 2, 8, 11, 15, 18, 20, 21 |
| EDN1 (1906) | 21 | SI (6476) | 15 |
| ESR1 (2099) | 7, 12, 15, 16, 17, 18, 20, 21 | SIRT1 (23411) | 21 |
| ESR2 (2100) | 7, 12, 15, 16, 17, 18, 20, 21 | SP1 (6667) | 21 |
| ESRRA (2101) | 12, 16, 21 | SULT2A1 (6822) | 21 |
| ESRRB (2103) | 12, 16, 21 | TNF (7124) | 17, 18 |
| ESRRG (2104) | 12, 16, 21 | TP53 (7157) | 17, 21 |
| FOS (2353) | 17, 18 | TSC22D3 (1831) | 21 |
| VDR (7421) | 21 |
| Gene (Entrz ID) | Chemical IDs | Gene (Entrz ID) | Chemical IDs |
|---|---|---|---|
| ADH1B (125) | 16 | GBA (2629) | 21 |
| ADH1C (126) | 16 | GSK3B (2932) | 18 |
| ADH4 (127) | 16 | HDAC1 (3065) | 18 |
| AHR (196) | 17 | HSD17B7 (51478) | 18 |
| AKTIP (64400) | 9, 21 | IL4 (3565) | 13, 18 |
| ALB (213) | 21 | JAK1 (3716) | 9 |
| ALG5 (29880) | 18 | LOX (4015) | 5 |
| ALOX12 (239) | 5 | MAP2K4 (6416) | 21 |
| ALOX5 (240) | 4 | MGAM (8972) | 16 |
| APEX1 (328) | 21 | MMP1 (4312) | 17 |
| AR (367) | 13, 21 | MMP2 (4313) | 9 |
| ATP1A1 (476) | 9 | MPO (4353) | 1 |
| AURKB (9212) | 13 | MTOR (2475) | 13 |
| BCHE (590) | 15 | NOS2 (4843) | 5, 17 |
| BDNF (627) | 16 | NR1I2 (8856) | 16 |
| BTK (695) | 21 | OLR1 (4973) | 18 |
| CASP3 (836) | 9, 16 | PARP1 (142) | 9, 13 |
| CAV1 (857) | 16 | PIP4K2A (5305) | 4 |
| CD38 (952) | 18 | PKD2 (5311) | 15 |
| CDK4 (1019) | 13 | PPARA (5465) | 21 |
| CDK9 (1025) | 18 | PPARG (5468) | 17 |
| CFTR (1080) | 21 | PPP3CA (5530) | 17 |
| COMT (1312) | 9 | PROCR (10544) | 1 |
| CXCR4 (7852) | 18 | PTK2B (2185) | 16 |
| CYP19A1 (1588) | 15, 18 | RAD51 (5888) | 5 |
| CYP1A1 (1543) | 17 | RELA (5970) | 17 |
| CYP1A2 (1544) | 17 | RPS9 (6203) | 18 |
| CYP1B1 (1545) | 17 | SCD (6319) | 18 |
| DNMT1 (1786) | 21 | SIRT1 (23411) | 13 |
| DPP4 (1803) | 18 | SLC2A1 (6513) | 21 |
| EGFR (1956) | 21 | SLC52A1 (55065) | 16 |
| EPHB2 (2048) | 16 | STAT3 (6774) | 9 |
| ESR1 (2099) | 7, 9, 15, 16, 17, 21 | SYK (6850) | 21 |
| ESR2 (2100) | 7, 17, 21 | TNF (7124) | 1, 17 |
| FABP4 (2167) | 18 | TOP1 (7150) | 13 |
| FAS (355) | 4 | VEGFA (7422) | 18 |
| FASN (2194) | 21 | XDH (7498) | 4 |
| Target | Entrz Gene ID | Gene Family |
|---|---|---|
| CYP1A1 | 1543 | Cytochrome P450 family 1 |
| CYP1B1 | 1545 | Cytochrome P450 family 1 |
| CYP19A1 | 1588 | Cytochrome P450 family 19 |
| AR | 367 | Nuclear hormone receptors |
| ESR1 | 2099 | Nuclear hormone receptors |
| ESR2 | 2100 | Nuclear hormone receptors |
| NR1I2 | 8856 | Nuclear hormone receptors |
| PPARG | 5468 | Nuclear hormone receptors |
| TNF | 7124 | Tumor necrosis factor superfamily |
| ALB | 213 | Unglycosylated serum protein |
| AHR | 196 | Basic helix-loop-helix proteins |
| CASP3 | 836 | Caspases |
| CAV1 | 857 | Membrane proteins |
| SIRT1 | 23411 | Sirtuins |
| Pathway | Category | EF | p Value |
|---|---|---|---|
| Tryptophan metabolism | Amino acid metabolism | 8.3203 | 0.0014 |
| Apoptosis | Cell growth and death | 4.0634 | 0.0110 |
| Ovarian steroidogenesis | Endocrine system | 2.7301 | 0.0321 |
| Prolactin signaling pathway | Endocrine system | 10.2781 | 0.0001 |
| Estrogen signaling pathway | Endocrine system | 4.8535 | 0.0067 |
| Steroid hormone biosynthesis | Lipid metabolism | 3.5659 | 0.0302 |
| TNF signaling pathway | Signal transduction | 2.9615 | 0.0426 |
| Metabolism of xenobiotics by P450 | Xenobiotics biodegradation & metabolism | 2.5508 | 0.0383 |
| Pathway | Category | EF | p Value |
|---|---|---|---|
| Proteoglycans in cancer | Cancers | 4.0672 | 0.0013 |
| Chemical carcinogenesis | Cancers | 5.5127 | 0.0045 |
| Pertussis | Infectious diseases: Bacterial | 6.1832 | 0.0032 |
| Legionellosis | Infectious diseases: Bacterial | 8.3192 | 0.0014 |
| Amoebiasis | Infectious diseases: Parasitic | 4.3166 | 0.0089 |
| Toxoplasmosis | Infectious diseases: Parasitic | 3.4403 | 0.0166 |
| Hepatitis B | Infectious diseases: Viral | 3.0916 | 0.0222 |
| Amyotrophic lateral sclerosis | Neurodegenerative diseases | 8.6331 | 0.0012 |
| Viral myocarditis | Cardiovascular diseases | 6.3549 | 0.0030 |
| Non-alcoholic fatty liver disease | Endocrine and metabolic diseases | 3.0302 | 0.0234 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, H.; Ng, H.W.; Sakkiah, S.; Ge, W.; Perkins, R.; Tong, W.; Hong, H. Pathway Analysis Revealed Potential Diverse Health Impacts of Flavonoids that Bind Estrogen Receptors. Int. J. Environ. Res. Public Health 2016, 13, 373. https://doi.org/10.3390/ijerph13040373
Ye H, Ng HW, Sakkiah S, Ge W, Perkins R, Tong W, Hong H. Pathway Analysis Revealed Potential Diverse Health Impacts of Flavonoids that Bind Estrogen Receptors. International Journal of Environmental Research and Public Health. 2016; 13(4):373. https://doi.org/10.3390/ijerph13040373
Chicago/Turabian StyleYe, Hao, Hui Wen Ng, Sugunadevi Sakkiah, Weigong Ge, Roger Perkins, Weida Tong, and Huixiao Hong. 2016. "Pathway Analysis Revealed Potential Diverse Health Impacts of Flavonoids that Bind Estrogen Receptors" International Journal of Environmental Research and Public Health 13, no. 4: 373. https://doi.org/10.3390/ijerph13040373
APA StyleYe, H., Ng, H. W., Sakkiah, S., Ge, W., Perkins, R., Tong, W., & Hong, H. (2016). Pathway Analysis Revealed Potential Diverse Health Impacts of Flavonoids that Bind Estrogen Receptors. International Journal of Environmental Research and Public Health, 13(4), 373. https://doi.org/10.3390/ijerph13040373