
Blocking the Wnt/β-Catenin Pathway by Lentivirus-Mediated Short Hairpin RNA Targeting β-Catenin Gene Suppresses Silica-Induced Lung Fibrosis in Mice

Abstract
:1. Introduction
2. Methods and Material
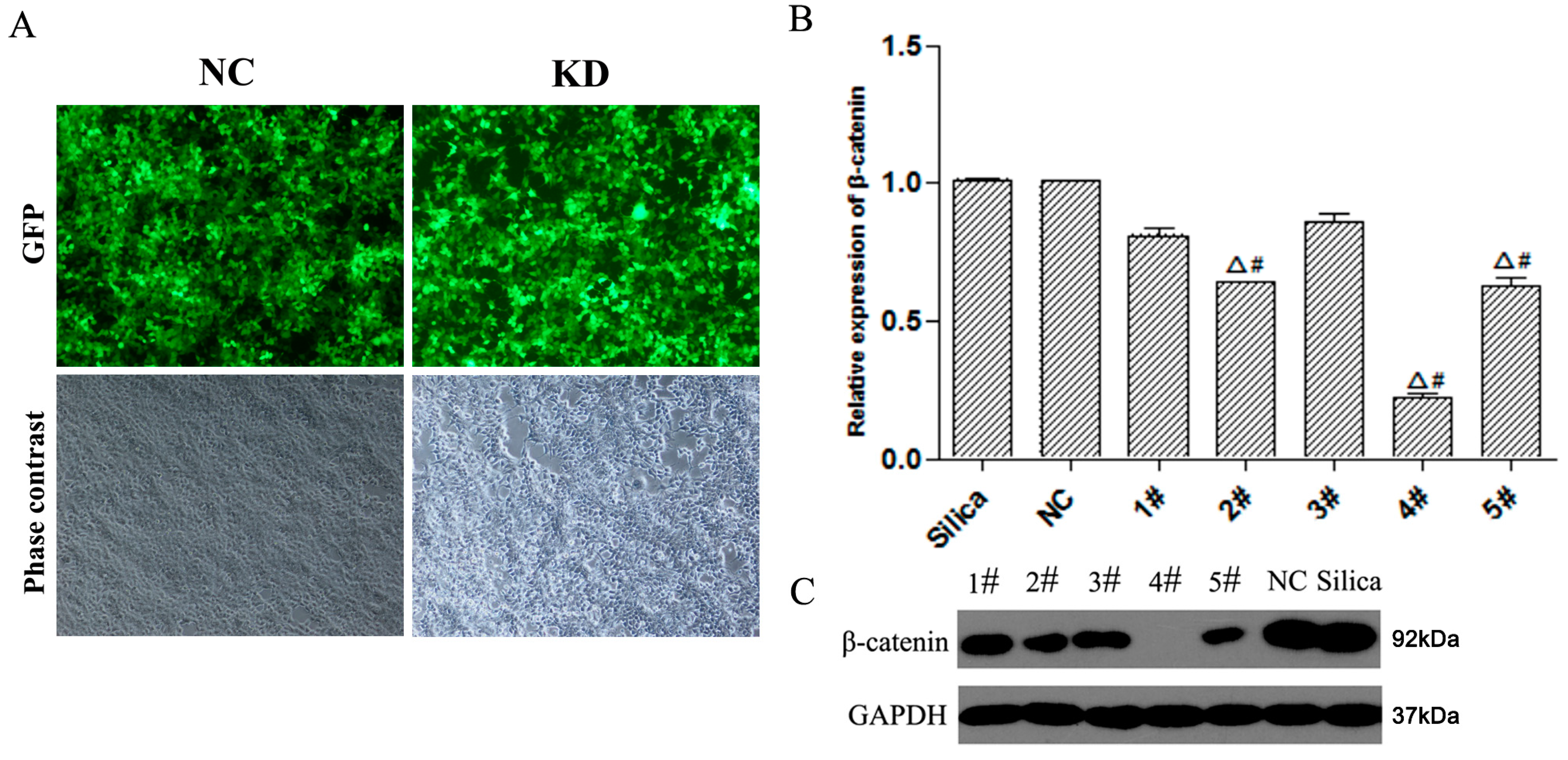
2.1. Construction of the Lentivirus Vectors of β-Catenin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item Sequence |
|---|
| Shβ-catenin -1 Oligo 1: 5’-CCGGACCCTGAGGAAGAAGATGTTTCAAGAGAACATCTTCTTCCTCAGGGTTTTTTTG-3’ |
| Oligo 2: 5’-AATTCAAAAAAACCCTGAGGAAGAAGATGTTCTCTTGAAACATCTTCTTCCTCAGGGT-3’ |
| Shβ-catenin -2 Oligo 1: 5’-CCGGCTGTTGTGGTTAAACTCCTTTCAAGAGAAGGAGTTTAACCACAACAGTTTTTTG-3’ |
| Oligo 2: 5’-AATTCAAAAAACTGTTGTGGTTAAACTCCTTCTCTTGAAAGGAGTTTAACCACAACAG-3’ |
| Shβ-catenin -3 Oligo 1: 5’-CCGGGAAGATGTTGACACCTCCCTTCAAGAGAGGGAGGTGTCAACATCTTCTTTTTTG-3’ |
| Oligo 2: 5’-AATTCAAAAAAGAAGATGTTGACACCTCCCTCTCTTGAAGGGAGGTGTCAACATCTTC-3’ |
| Shβ-catenin -4 Oligo 1: 5’-CCGGGGACAAGCCACAGGATTACAACTCGAGTTGTAATCCTGTGGCTTGTCCTTTTTTG-3’ |
| Oligo 2: 5’-AATTCAAAAAAGGACAAGCCACAGGATTACAACTCGAGTTGTAATCCTGTGGCTTGTCC -3’ |
| Shβ-catenin -5 Oligo 1: 5’-CCGGGCGTTATCAAACCCTAGCCTTCTCGAGAAGGCTAGGGTTTGATAACGCTTTTTTG-3’ |
| Oligo 2: 5’-AATTCAAAAAAGCGTTATCAAACCCTAGCCTTCTCGAGAAGGCTAGGGTTTGATAACGC-3’ |
| NC Oligo 1: 5’-CCGGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAATTTTTG-3’ |
| Oligo 2: 5’- AATTCAAAAAATTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGAA-3’ |
2.2. Cell Culture and Administration
2.3. Experimental Animals and Design
2.4. Quantitative Real-Time PCR Analysis
2.5. Western Blot Analysis
2.6. ELISA Assay for TGF-β1
2.7. Determination of Hydroxyproline Content
2.8. Pathological Examination
2.9. Immunohistochemical Staining
2.10. Statistical Analyses
3. Results
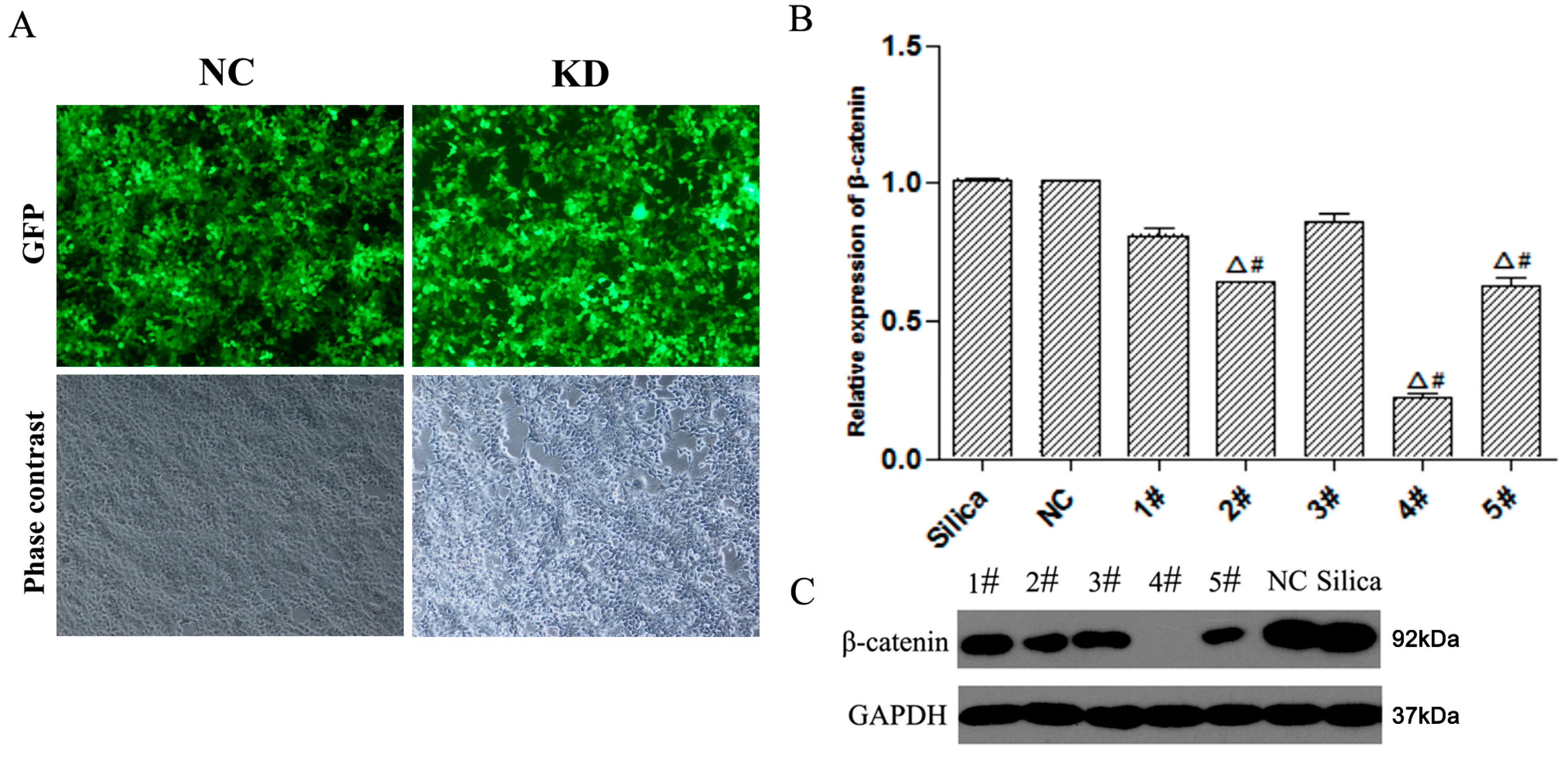
3.1. Construction and Screening of Lv-shβ-Catenin Vectors
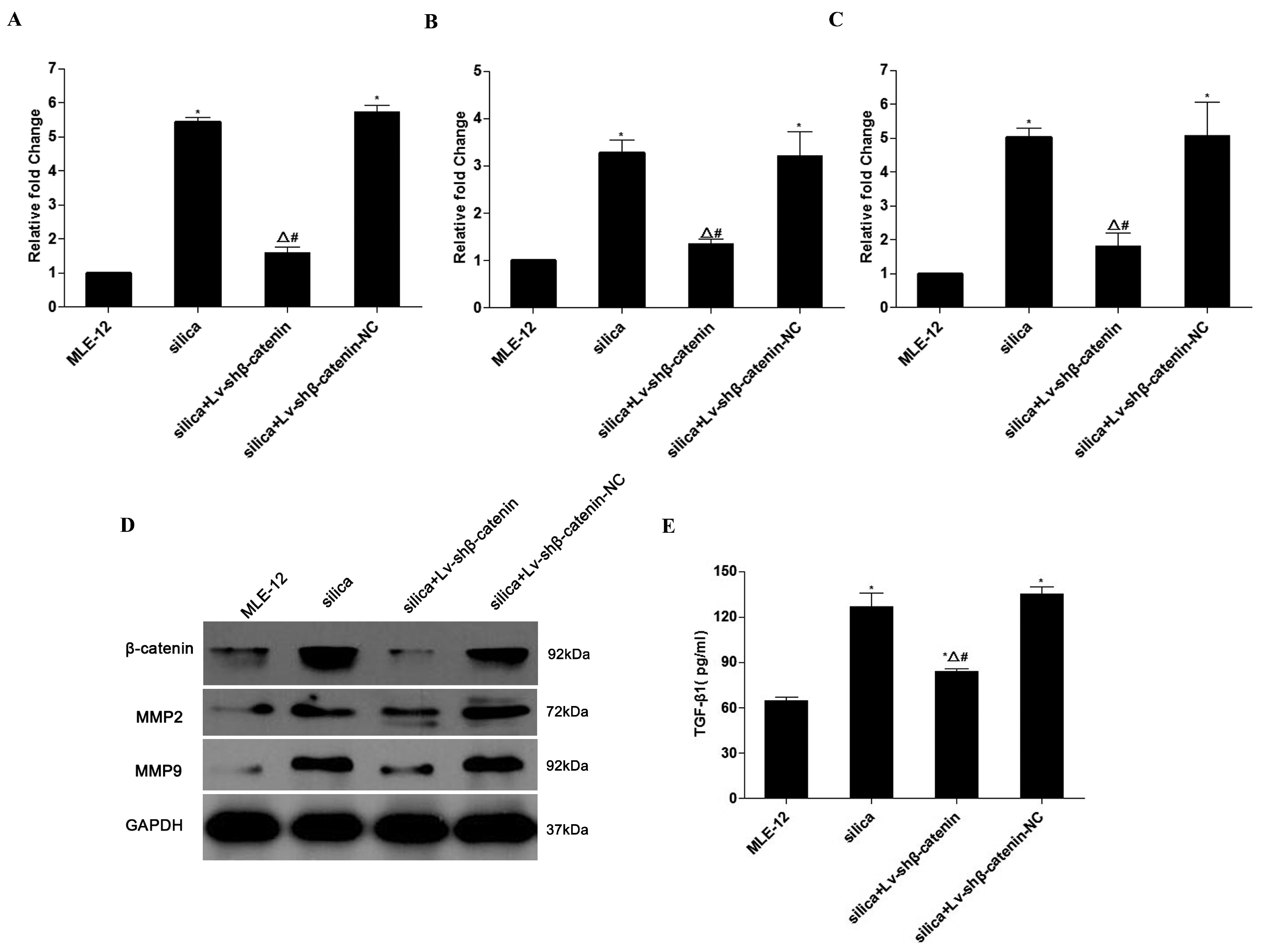
3.2. Effect of Lv-shβ-Catenin on Silica-Treated MLE-12 Cells
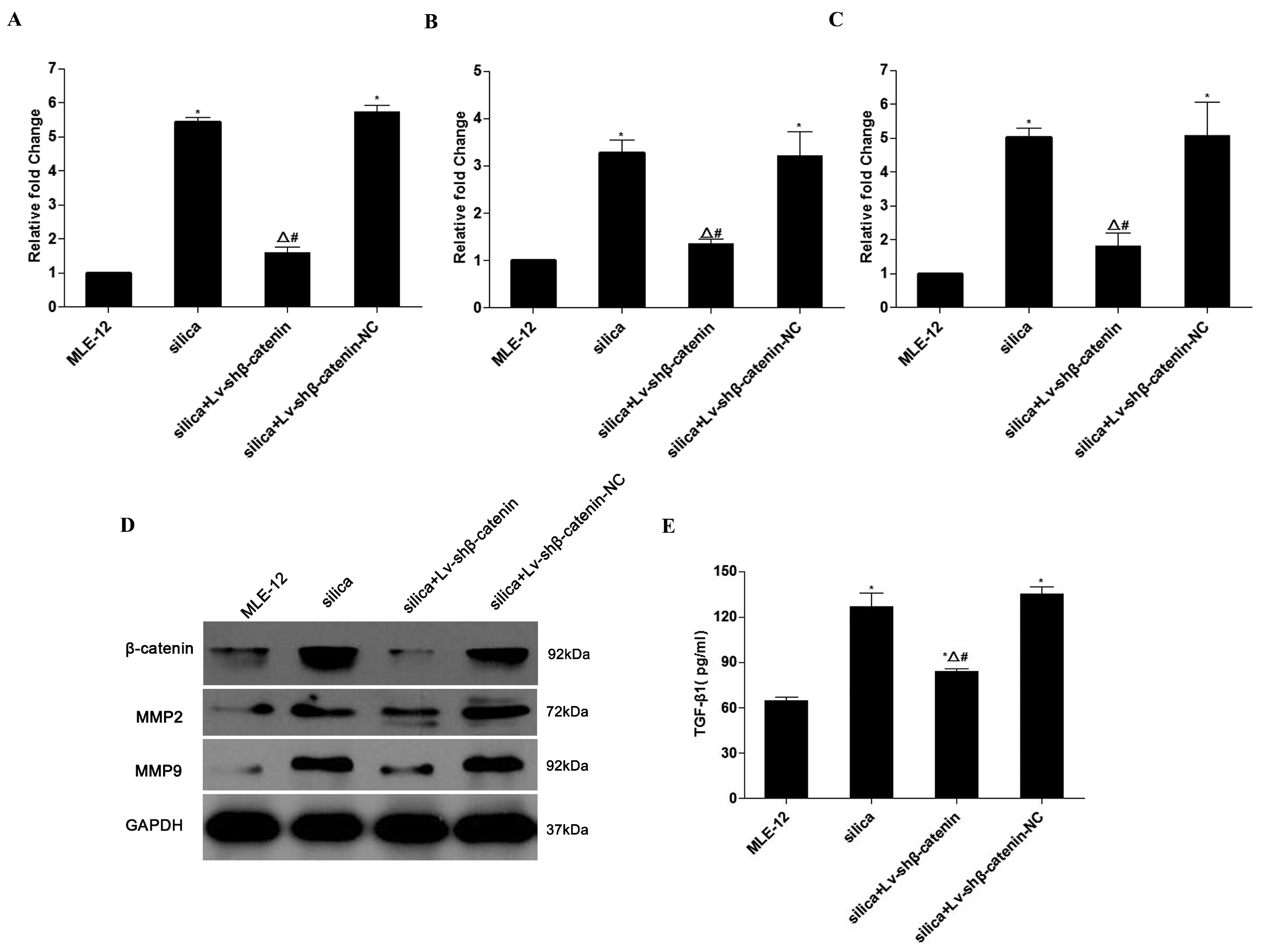
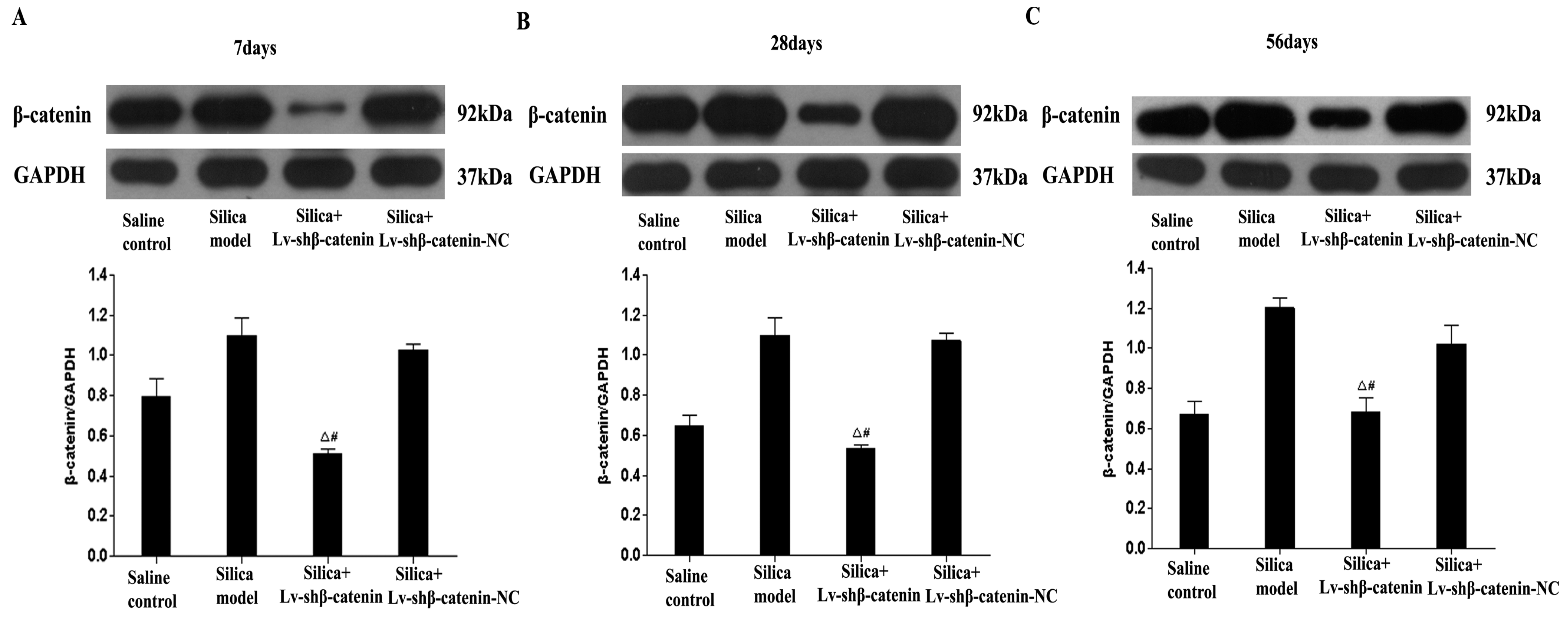
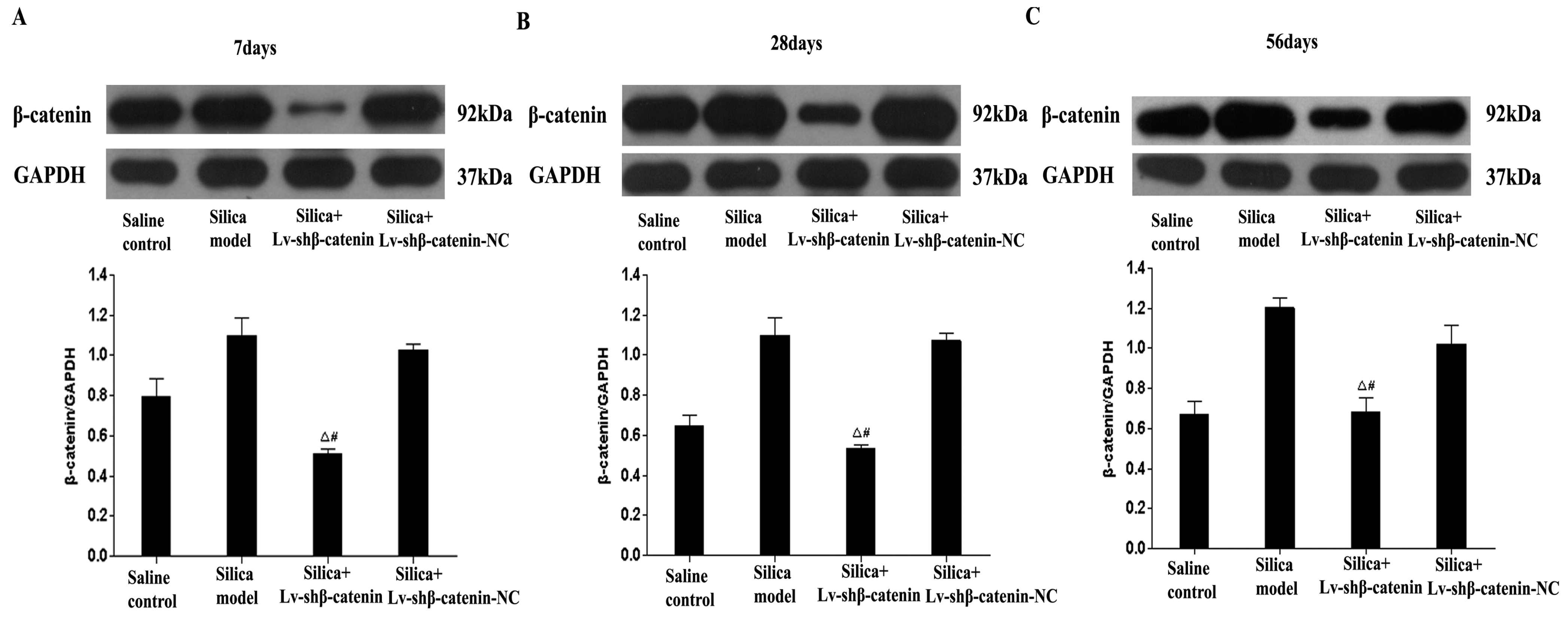
3.3. Inhibition of β-Catenin Expression by Lv-shβ-Catenin in Vivo
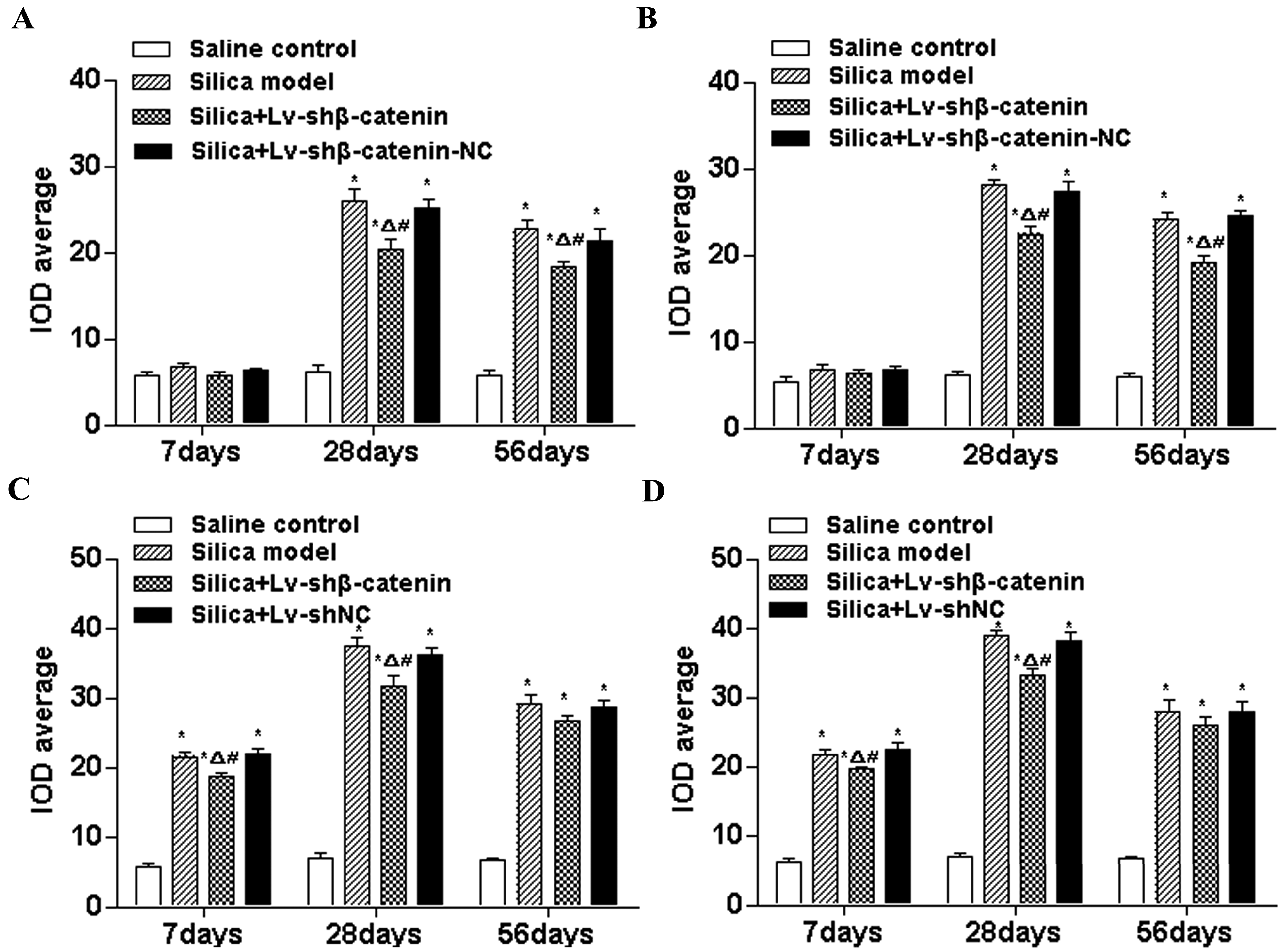
3.4. Inhibition of Silica-Induced Lung Fibrosis by Lv-shβ-Catenin in Vivo
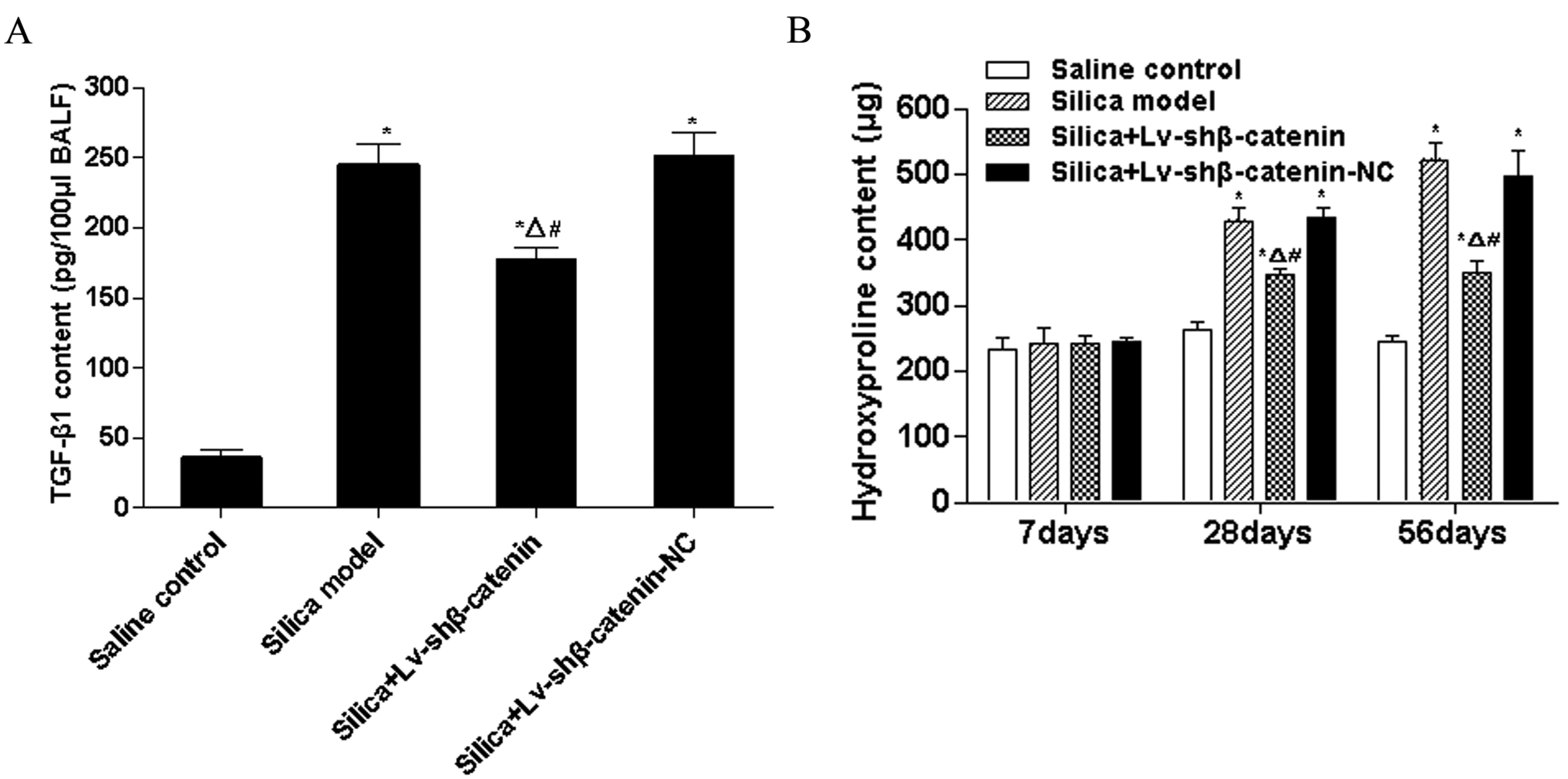
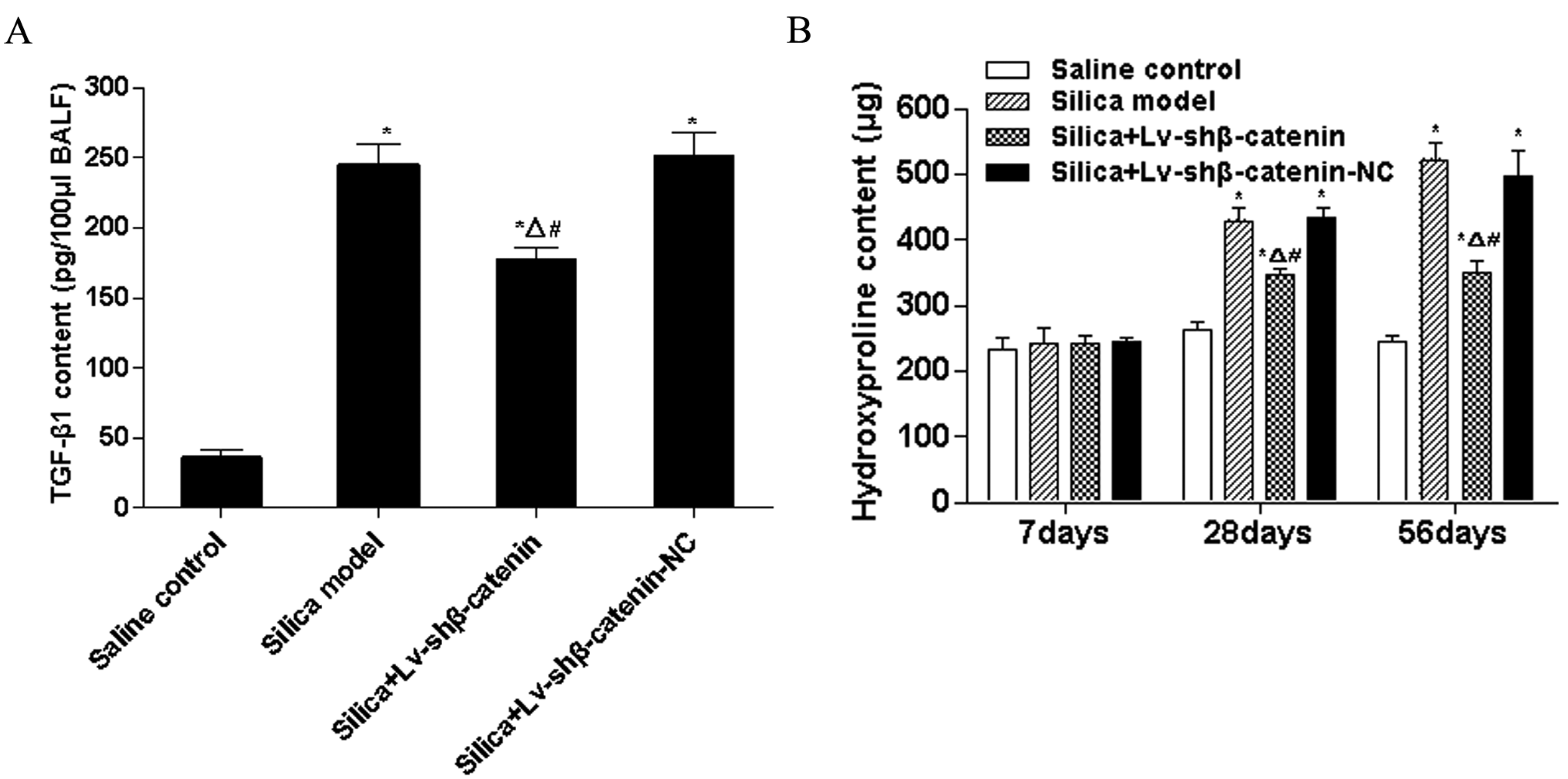
3.4.1. Lv-shβ-Catenin Could Reduce TGF-β1 Content in BALF
3.4.2. Lv-shβ-Catenin Could Reduce Hydroxyproline Content in the Lung
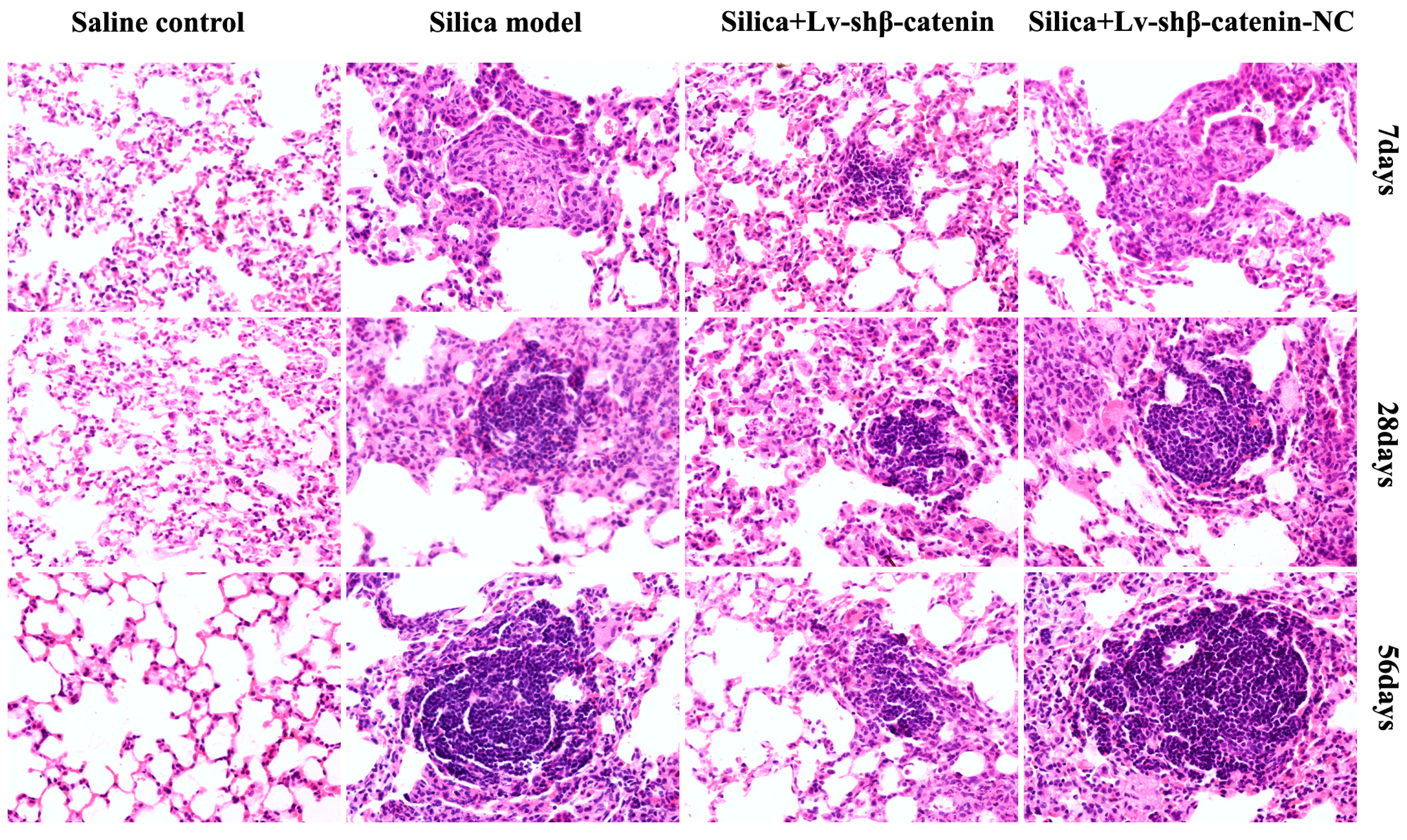
3.4.3. Lv-shβ-Catenin Could Alleviate Histopathologic Changes in the Lung
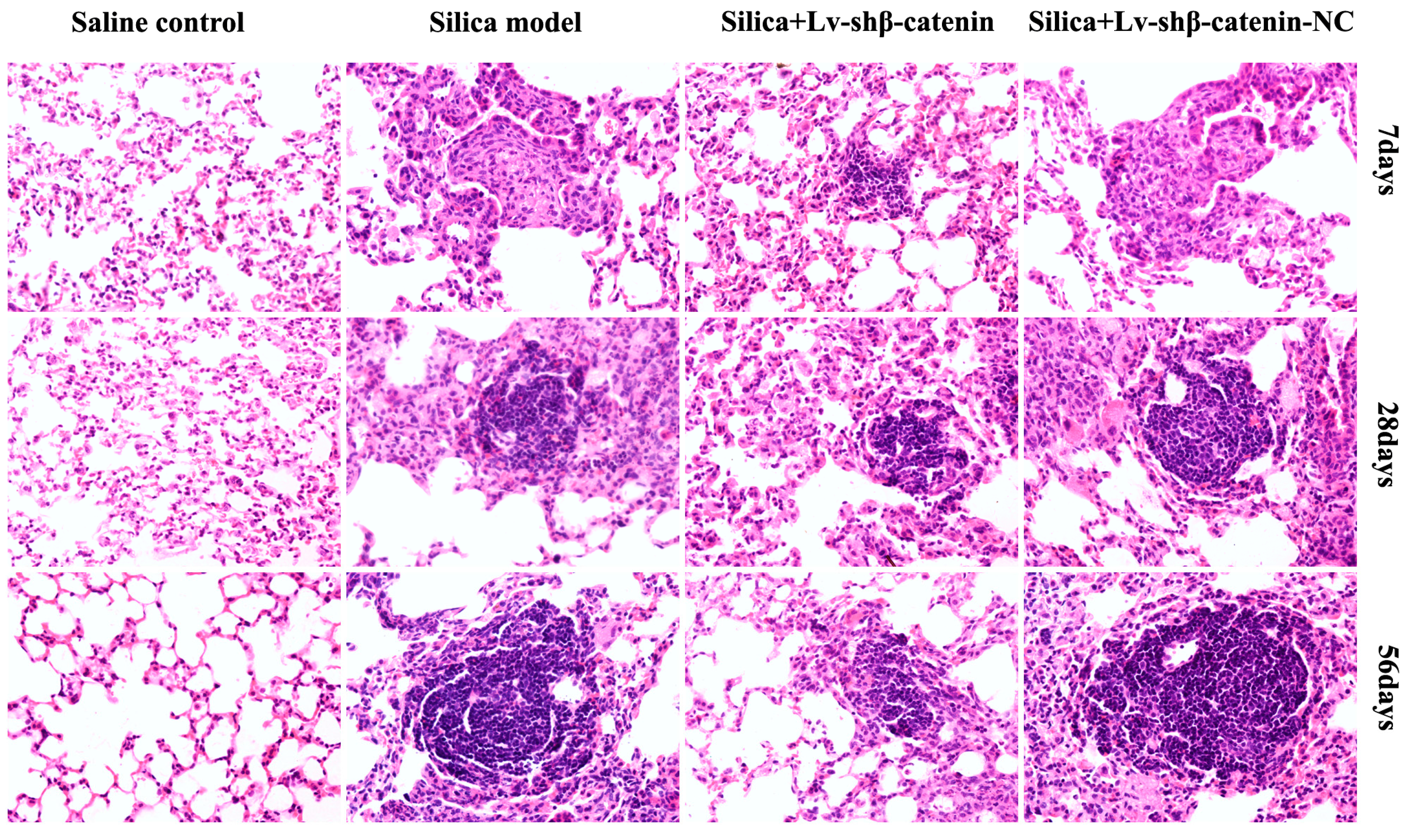
| Groups | 7 Days After Instillation | 28 Days After Instillation | 56 Days After Instillation | |||
|---|---|---|---|---|---|---|
| n | Silicotic Nodule Grade | n | Silicotic Nodule Grade | n | Silicotic Nodule Grade | |
| Saline control | 6 | 0 | 6 | 0 | 6 | 0 |
| Silica | 6 | I | 6 | I+ ~ II | 6 | II+ ~ III |
| Silica + Lv-shβ-catenin | 6 | 0 ~ I | 6 | I ~ I+ | 6 | I+ ~ II |
| Silica + Lv-shβ-catenin-NC | 6 | I | 6 | I+ ~ II | 6 | II+ ~ III |
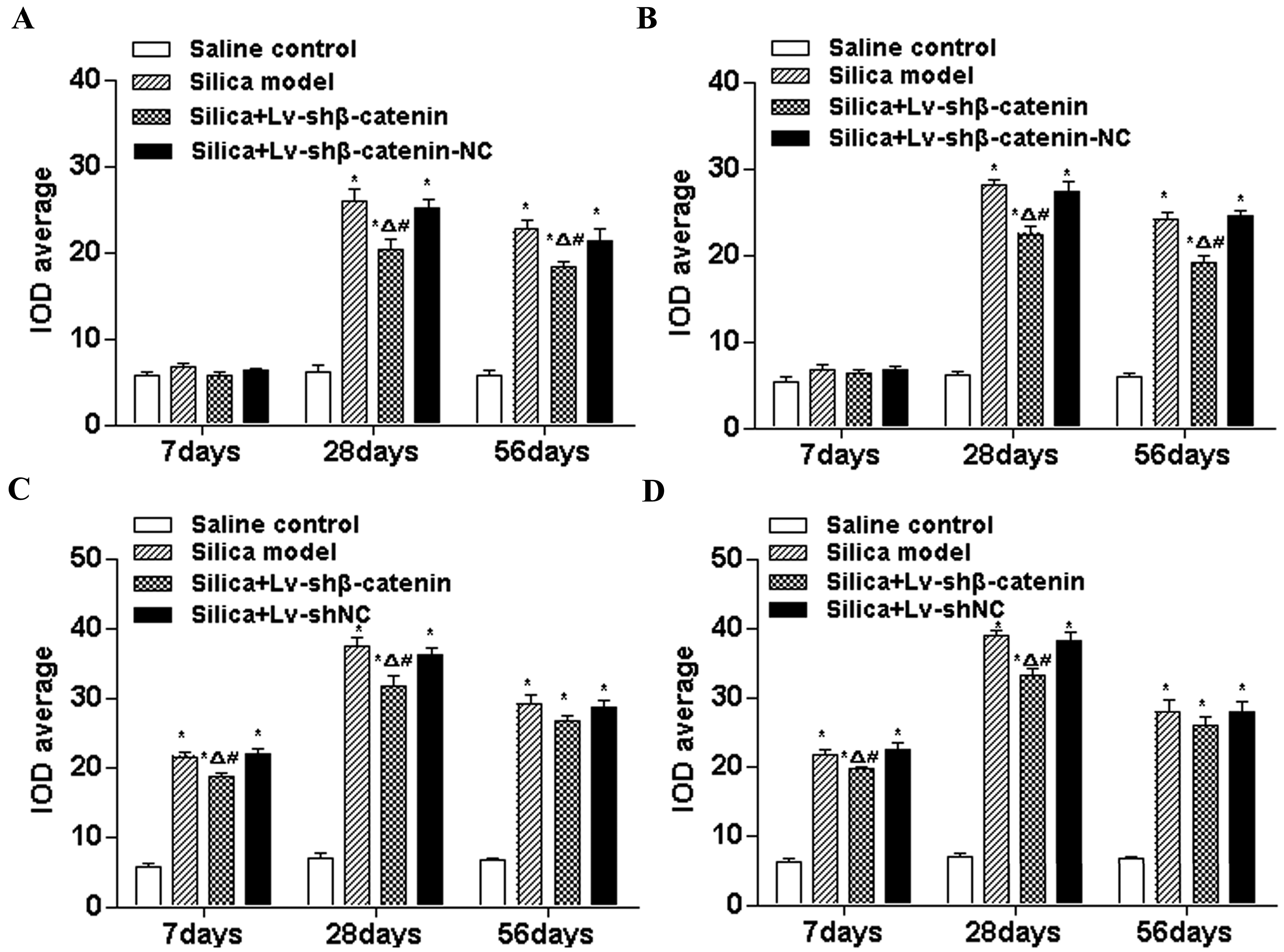
3.4.4. Lv-shβ-Catenin Could Inhibit the Expression of Collagen I, Collagen III, MMP2 and MMP9 in the Lung
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, F.; Liu, J.; Weng, D.; Chen, Y.; Song, L.; He, Q.; Chen, J. CD4+CD25+Foxp3+ regulatory T cells depletion may attenuate the development of silica-induced lung fibrosis in mice. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Weng, D.; Liu, F.; Chen, Y.; Li, C.; Dong, L.; Tang, W.; Chen, J. Tregs promote the differentiation of Th17 cells in silica-induced lung fibrosis in mice. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Ma, R.; Xu, Y.; Li, N.; Li, Z.; Yue, J.; Li, H.; Guo, Y.; Qi, D. Wnt2 promotes non-small cell lung cancer progression by activating WNT/β-catenin pathway. Am. J. Cancer Res. 2015, 5, 1032–1046. [Google Scholar] [PubMed]
- Kim, T.H.; Kim, S.H.; Seo, J.Y.; Chung, H.; Kwak, H.J.; Lee, S.K.; Yoon, H.J.; Shin, D.H.; Park, S.S.; Sohn, J.W. Blockade of the Wnt/β-catenin pathway attenuates bleomycin-induced pulmonary fibrosis. Tohoku J. Exp. Med. 2011, 223, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Königshoff, M.; Balsara, N.; Pfaff, E.M.; Kramer, M.; Chrobak, I.; Seeger, W.; Eickelberg, O. Functional Wnt signaling is increased in idiopathic pulmonary fibrosis. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Carron, B.; Yee, H.T.; Yie, T.A.; Hajjou, M.; Rom, W. Wnt pathway in pulmonary fibrosis in the bleomycin mouse model. J. Environ. Pathol. Toxicol. Oncol. 2009, 28, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Chilosi, M.; Poletti, V.; Zamò, A.; Lestani, M.; Montagna, L.; Piccoli, P.; Pedron, S.; Bertaso, M.; Scarpa, A.; Murer, B.; et al. Aberrant Wnt/beta-catenin pathway activation in idiopathic pulmonary fibrosis. Am. J. Pathol. 2003, 162, 1495–1502. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Z.; Qiu, X.; Li, Y.; Qin, J.; Han, X. Roles of Wnt/beta-catenin signaling in epithelial differentiation of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2009, 390, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Pardo, A.; Pérez-Ramos, J.; Segura-Valdez, L.; Ramírez, R.; Selman, M. Expression and localization of TIMP-1, TIMP-2, MMP-13, MMP-2, and MMP-9 in early and advanced experimental lung silicosis. Ann. NY Acad. Sci. 1999, 878, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Liao, J.; Xin, Z.Q.; Lu, W.Q.; Liu, A.L. Relaxin attenuates silica-induced pulmonary fibrosis by regulating collagen type I and MMP-2. Int. Immunopharmacol. 2013, 17, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Cutroneo, K.R.; White, S.L.; Phan, S.H.; Ehrlich, H.P. Therapies for bleomycin induced lung fibrosis through regulation of TGF-beta1 induced collagen gene expression. J. Cell Physiol. 2007, 211, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Henderson, W.R., Jr.; Chi, E.Y.; Ye, X.; Nguyen, C.; Tien, Y.T.; Zhou, B.; Borok, Z.; Knight, D.A.; Kahn, M. Inhibition of Wnt/beta-catenin/CREB binding protein (CBP) signaling reverses pulmonary fibrosis. Proc. Natl. Acad Sci. USA. 2010, 107, 14309–14314. [Google Scholar] [CrossRef] [PubMed]
- Pullmann, R., Jr.; Juhaszova, M.; de Silanes, I.L.; Kawai, T.; Mazan-Mamczarz, K.; Halushka, M.K.; Gorospe, M. Enhanced proliferation of cultured human vascular smooth muscle cells linked to increased function of RNA-binding protein HuR. J. Biol. Chem. 2005, 280, 22819–22826. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, T.; Kasahara, N.; Hamamori, Y.; Kedes, L. A high-titer lentiviral production system mediates efficient transduction of differentiated cells including beating cardiac myocytes. J. Mol. Cell Cardiol. 1999, 31, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, Y.; Lv, L.; Chen, J. Silencing CD36 gene expression results in the inhibition of latent-TGF-β1 activation and suppression of silica-induced lung fibrosis in the rat. Respir. Res. 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Free Online Primer Design Tool. Available online: http://frodo.wi.mit.edu/primer3 (accessed on 8 August 2015).
- Basic Local Alignment Search Tool. Available online: http://www.blast.ncbi.nlm.nih.gov/ (accessed on 8 August 2015).
- Cohen, J.C.; Larson, J.E.; Killeen, E.; Love, D.; Takemaru, K. CFTR and Wnt/beta-catenin signaling in lung development. BMC Dev. Biol. 2008, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, J.N.; Hu, X.F.; Chen, X.L.; Wang, X.R.; Zhao, T.T.; Peng, M.J.; Zou, P. Effects of pentoxifylline on Wnt/β-catenin signaling in mice chronically exposed to cigarette smoke. Chin. Med. J. 2010, 123, 2688–2694. [Google Scholar] [PubMed]
- O’Connor, C.M.; FitzGerald, M.X. Matrix metalloproteases and lung disease. Thorax 1994, 49, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Zuo, F.; Kaminski, N.; Eugui, E.; Allard, J.; Yakhini, Z.; Ben-Dor, A.; Lollini, L.; Morris, D.; Kim, Y.; DeLustro, B.; Sheppard, D.; Pardo, A.; Selman, M.; Heller, R.A. Gene expression analysis reveals matrilysin as a key regulator of pulmonary fibrosis in mice and humans. Proc. Natl. Acad Sci. USA 2002, 99, 6292–6297. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Crampton, S.P.; Hughes, C.C. Wnt signaling induces matrix metalloproteinase expression and regulates T cell transmigration. Immunity 2007, 26, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Mickler, E.A.; Cummings, O.W.; Sandusky, G.E.; Weber, D.J.; Gracon, A.; Woodruff, T.; Wilkes, D.S.; Vittal, R. Crosstalk between TGF-β1 and complement activation augments epithelial injury in pulmonary fibrosis. FASEB J. 2014, 28, 4223–4234. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Park, J.W.; Cho, W.K.; Zhou, Y.; Han, B.; Yoon, P.O.; Chae, J.; Elias, J.A.; Lee, C.G. Modifiers of TGF-β1 effector function as novel therapeutic targets of pulmonary fibrosis. Korean J. Intern. Med. 2014, 29, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, A.A.; Nazari, Z.; Samei, M. A comparative study of grape seed extract and vitamin E effects on silica-induced pulmonary fibrosis in rats. Pulm. Pharmacol. Ther. 2008, 21, 668–674. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Dai, W.; Wang, Y.; Gu, Q.; Yang, D.; Zhang, M. Blocking the Wnt/β-Catenin Pathway by Lentivirus-Mediated Short Hairpin RNA Targeting β-Catenin Gene Suppresses Silica-Induced Lung Fibrosis in Mice. Int. J. Environ. Res. Public Health 2015, 12, 10739-10754. https://doi.org/10.3390/ijerph120910739
Wang X, Dai W, Wang Y, Gu Q, Yang D, Zhang M. Blocking the Wnt/β-Catenin Pathway by Lentivirus-Mediated Short Hairpin RNA Targeting β-Catenin Gene Suppresses Silica-Induced Lung Fibrosis in Mice. International Journal of Environmental Research and Public Health. 2015; 12(9):10739-10754. https://doi.org/10.3390/ijerph120910739
Chicago/Turabian StyleWang, Xin, Wujing Dai, Yanrang Wang, Qing Gu, Deyi Yang, and Ming Zhang. 2015. "Blocking the Wnt/β-Catenin Pathway by Lentivirus-Mediated Short Hairpin RNA Targeting β-Catenin Gene Suppresses Silica-Induced Lung Fibrosis in Mice" International Journal of Environmental Research and Public Health 12, no. 9: 10739-10754. https://doi.org/10.3390/ijerph120910739
APA StyleWang, X., Dai, W., Wang, Y., Gu, Q., Yang, D., & Zhang, M. (2015). Blocking the Wnt/β-Catenin Pathway by Lentivirus-Mediated Short Hairpin RNA Targeting β-Catenin Gene Suppresses Silica-Induced Lung Fibrosis in Mice. International Journal of Environmental Research and Public Health, 12(9), 10739-10754. https://doi.org/10.3390/ijerph120910739