
Αlpha 2a-Adrenoceptor Gene Expression and Early Life Stress-Mediated Propensity to Alcohol Drinking in Outbred Rats

Abstract
:1. Introduction
2. Experimental Section
2.1. Animals
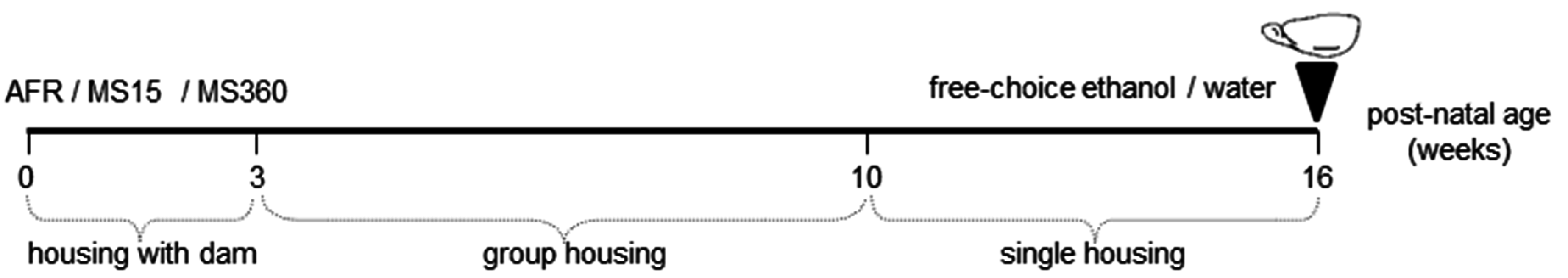
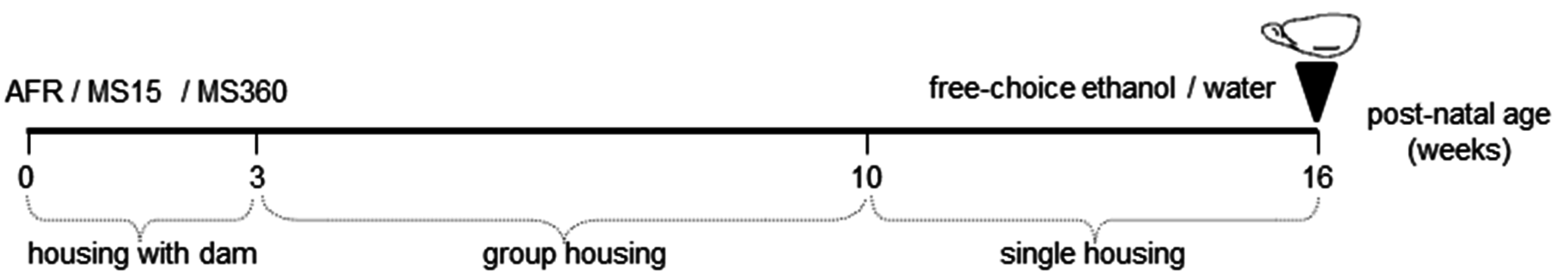
2.2. Early-Life Rearing Conditions
2.3. Voluntary Ethanol Consumption
2.4. Adra2a Gene Expression Analyses
2.5. Adra2a Promoter DNA Methylation Analyses
2.6. Statistics
3. Results
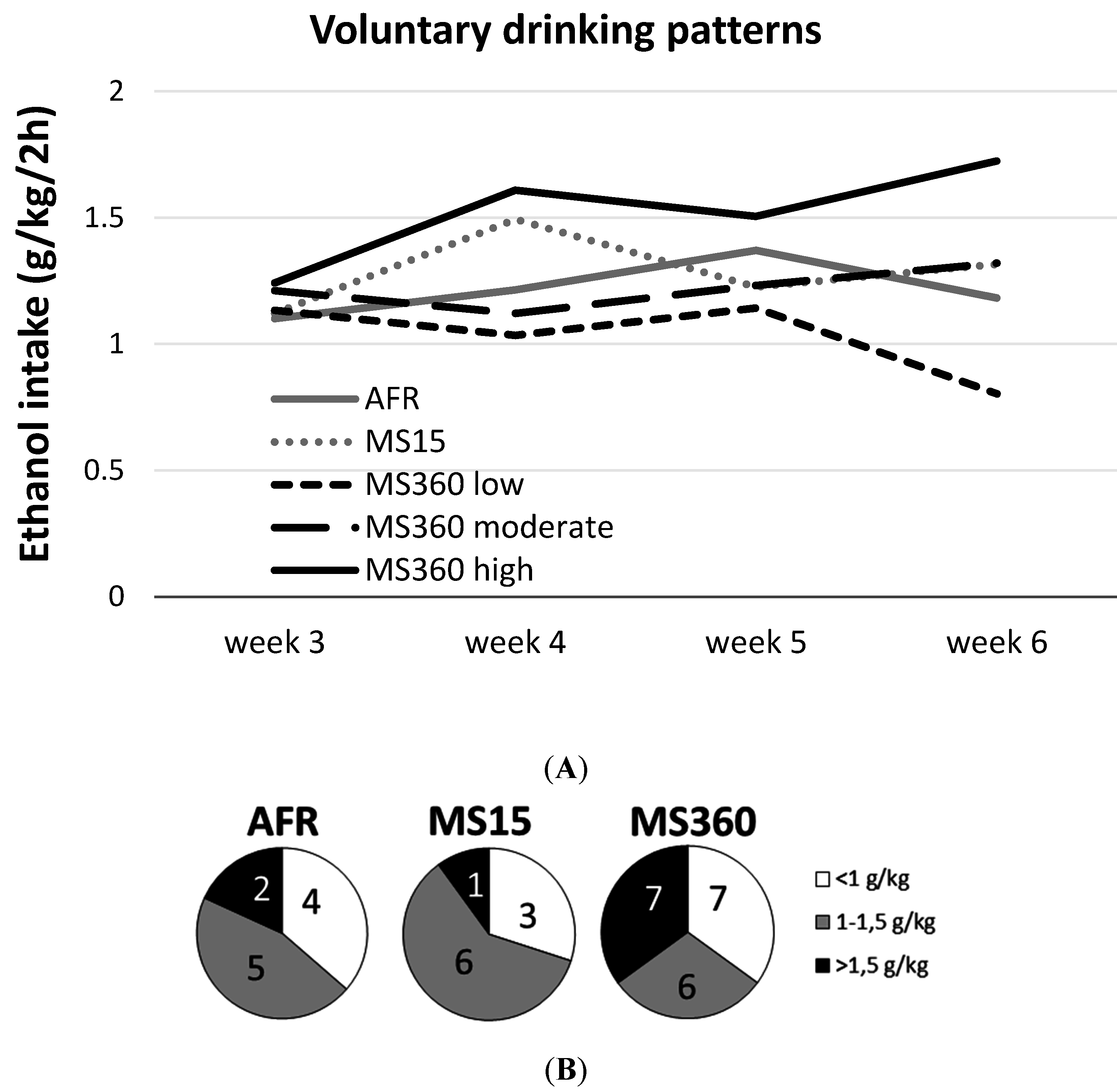
3.1. Ethanol Intake and Preference
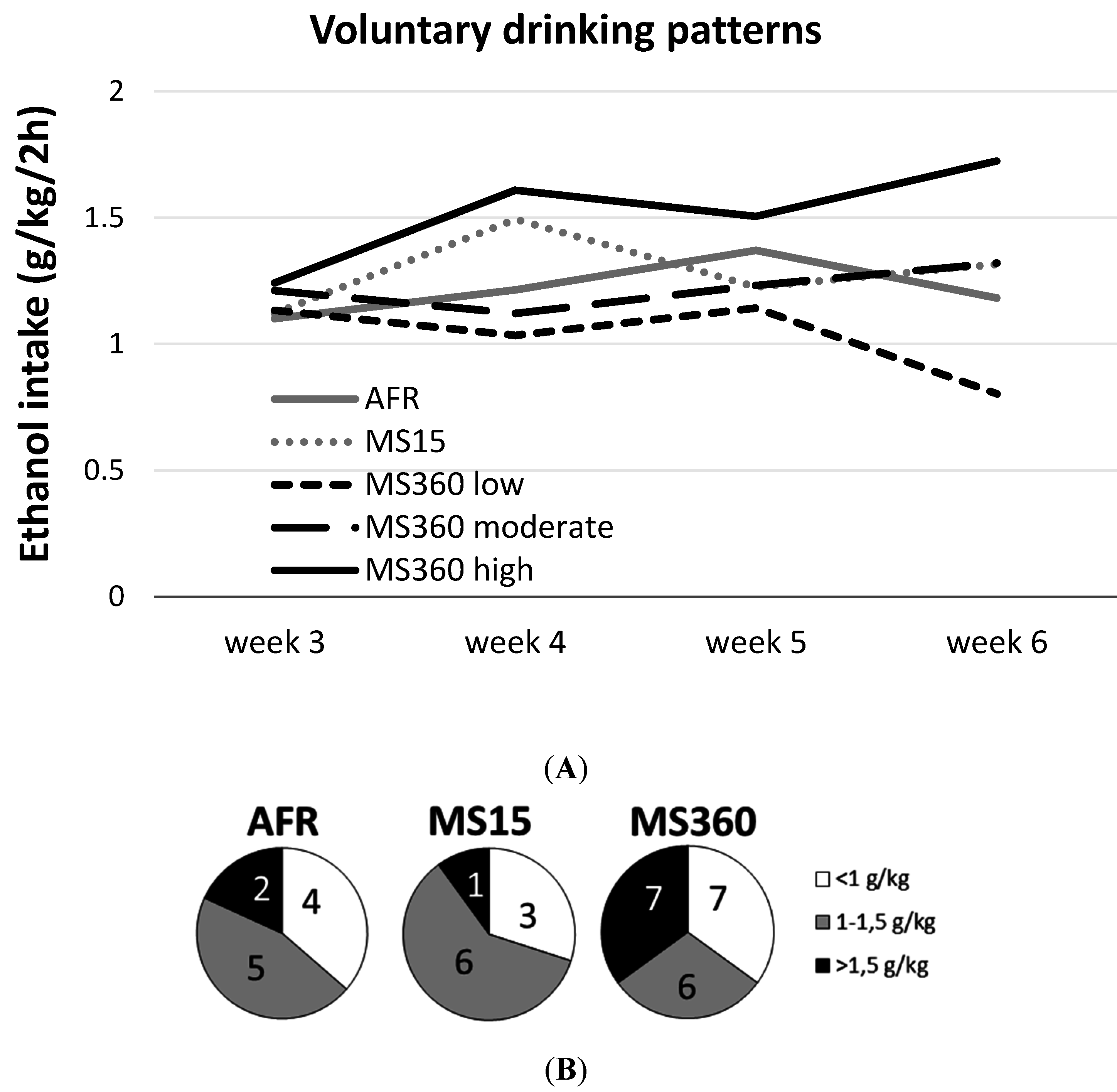
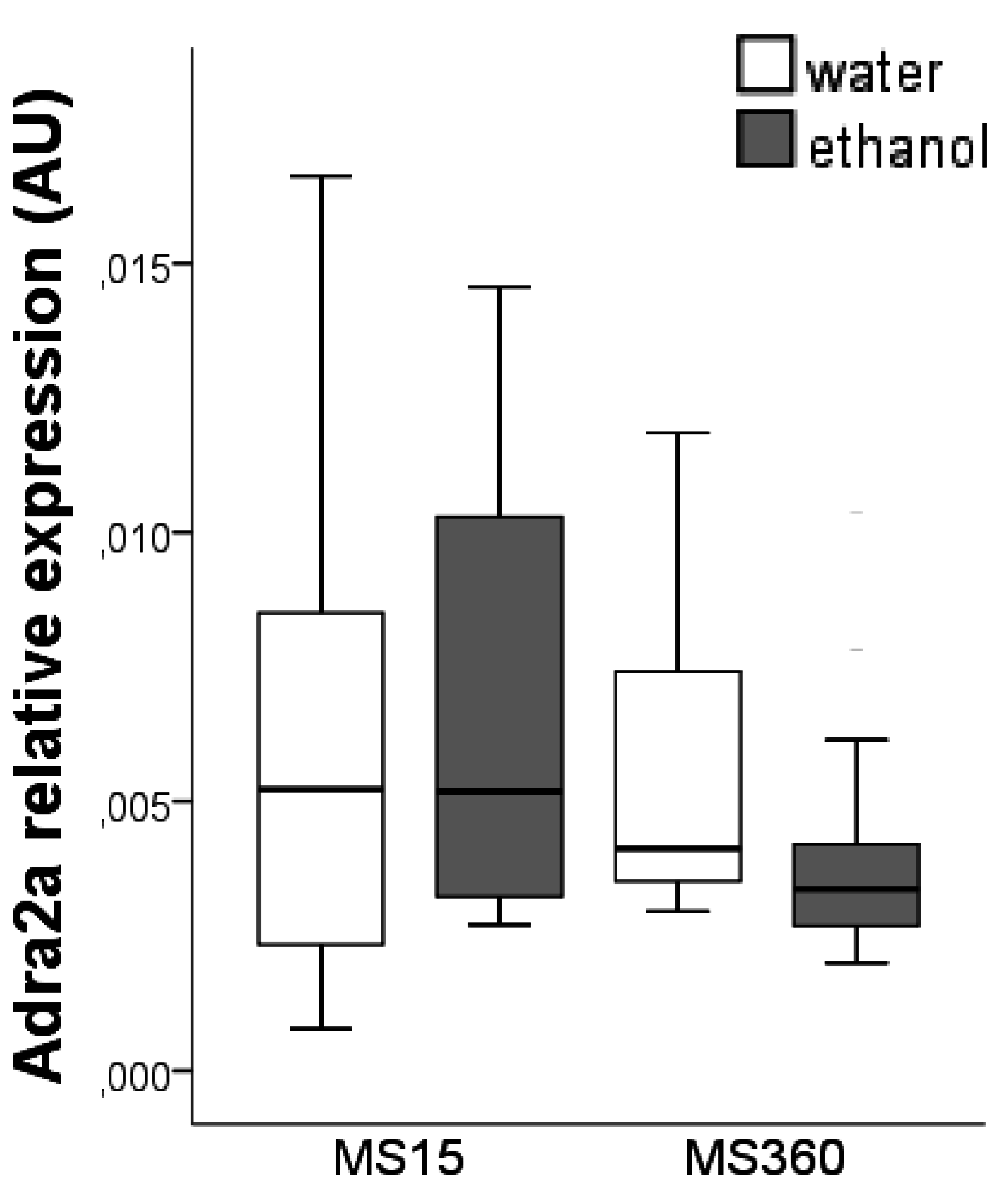
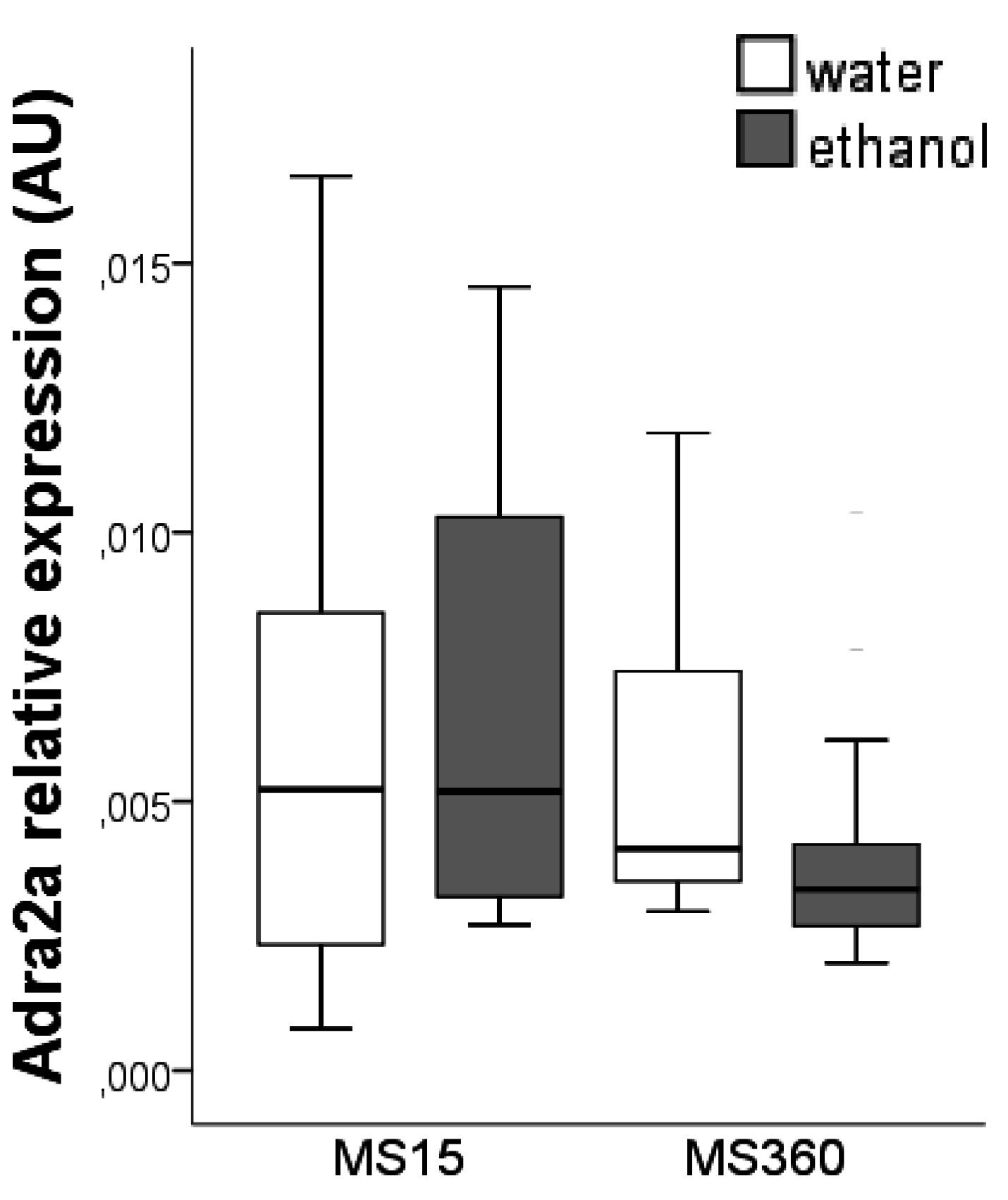
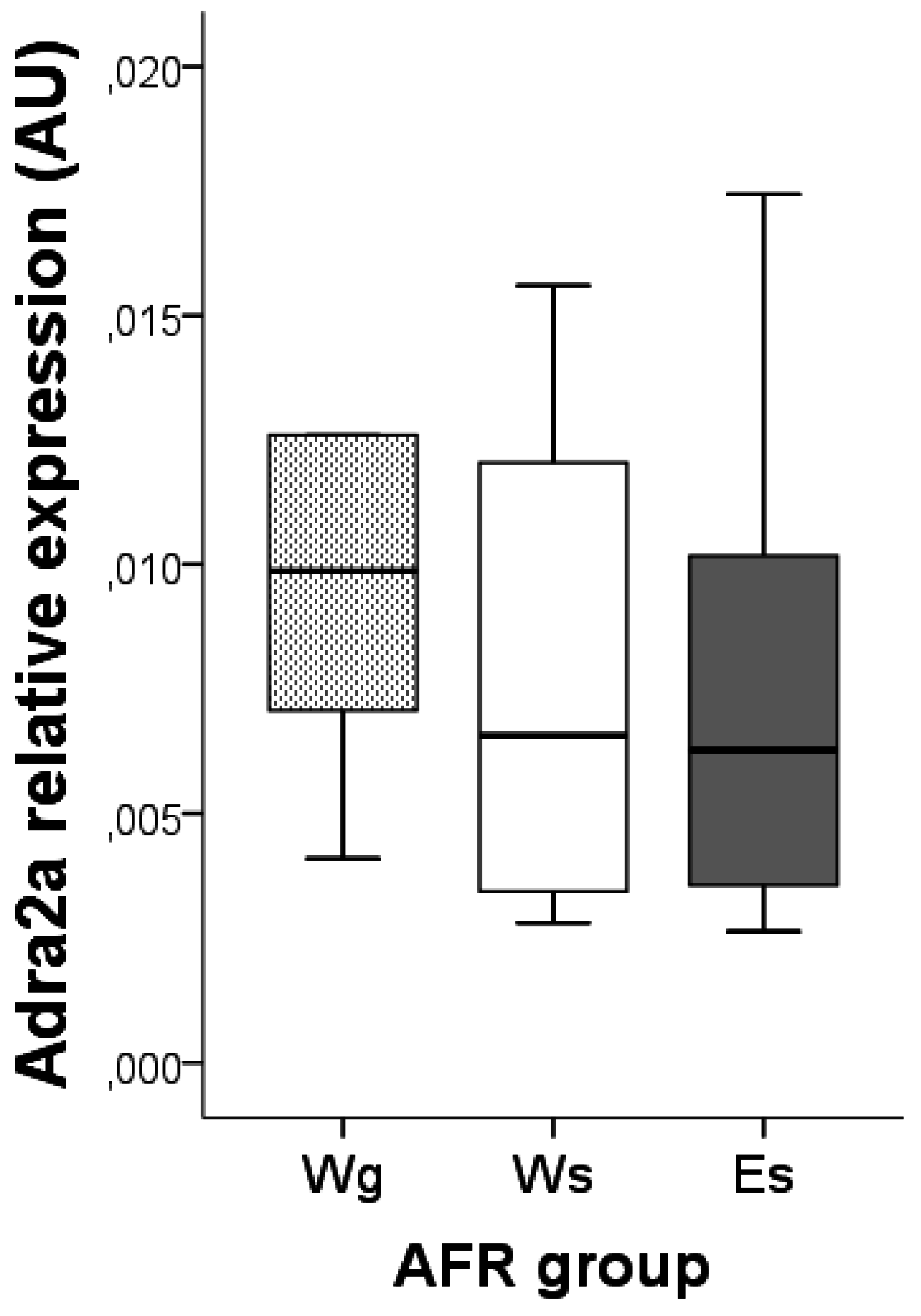
3.2. Adra2a Gene Expression

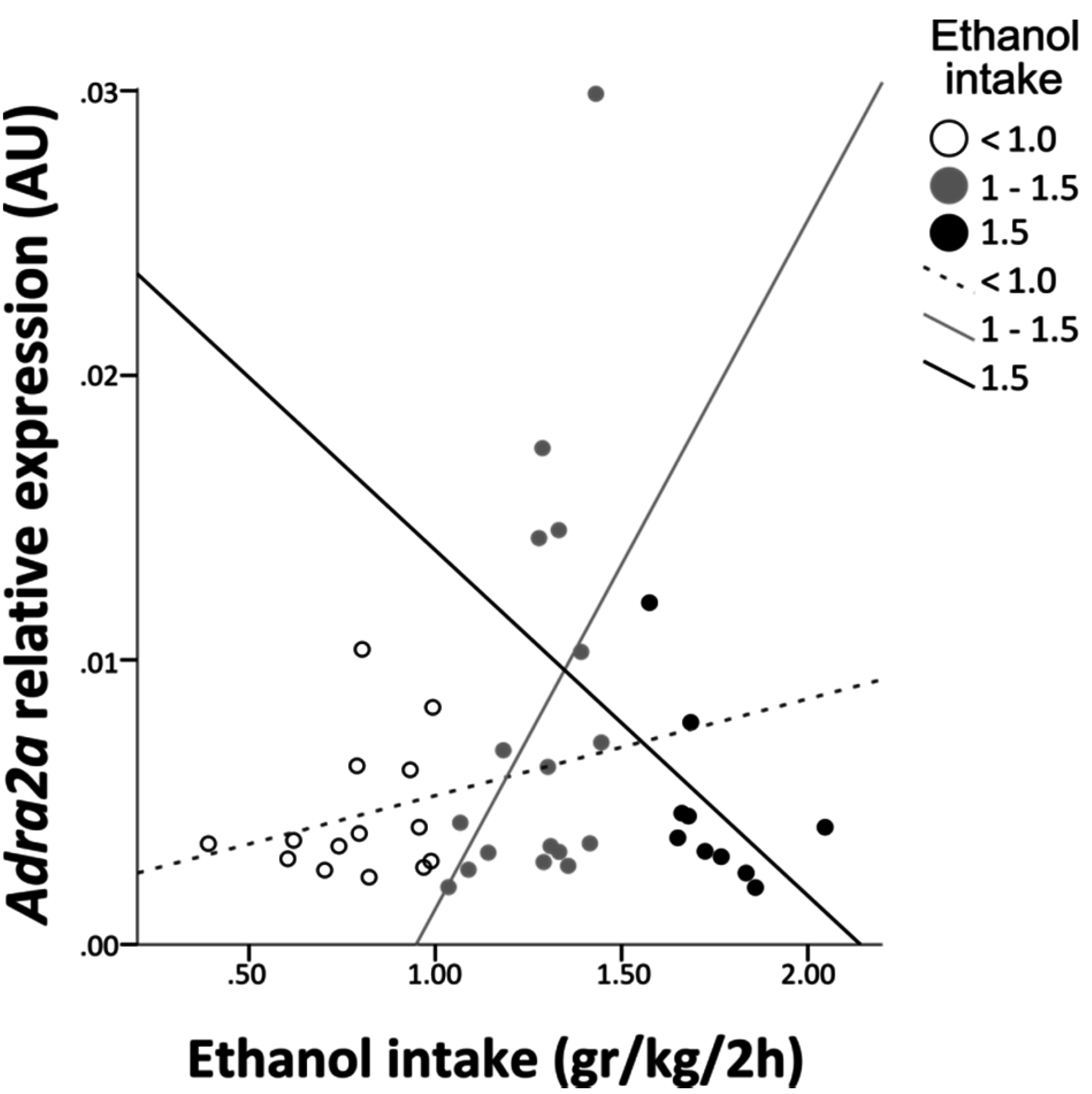
4. Discussion
5. Conclusions
Supplementary Material
DNA Methylation Analysis
| Gene | Chromosome position | Position (ref to TSS) | Number of CpG Sites |
| Adra2a | chr1: 282178264-82178429 | −211 to −46 | 19 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Initial denaturation °C Time# | Denaturation °C Time# | Annealing °C Time# | Elongation °C Time# | Cycles | Melting curve |
|---|---|---|---|---|---|---|---|
| Actb | Forward: 5’CACTGCCGCATCCTCTTCCT 3’ Reverse: 5’ AACCGCTCATTGCCGATAGTG 3’ | 95 °C 3:00 | 95 °C 0:10 | 60 °C 0:30 | 72 °C 0:45 | 45 | 65 to 95 °C increment 0.5 °C |
| Gapdh | Forward: 5’ ACATGCCGCCTGGAGAAACCT 3’ Reverse: 5’ GCCCAGGATGCCCTTTAGTGG 3’ | 95 °C 3:00 | 95 °C 0:10 | 60 °C 0:30 | 72 °C 0:30 | 45 | 65 to 95 °C increment 0.5 °C |
| Rpl19 | Forward: 5’CCAATGAAACCAACGAAATC 3’ Reverse: 5’TACCCTTCCTCTTCCCTA 3’ | 97 °C 3:00 | 95 °C 0:10 | 60 °C 0:30 | 72 °C 0:40 | 45 | 65 to 95 °C increment 0.5 °C |
| Adra2a | Forward: 5' GGTAAGGTGTGGTGCGAGAT 3’ Reverse: 5’ CAGCGCCCTTCTTCTCTATG 3’ | 95 °C 3:00 | 95 °C 0:10 | 60 °C 0:30 | 72 °C 0:45 | 49 | 65 to 95 °C increment 0.5 °C |
| 5 %, 24h | 5 %, 2h | 20 %, 2h | ||||
|---|---|---|---|---|---|---|
| Week 1 | Week 2 | Week 3 | Week 4 | Week 5 | Week 6 | |
| AFR | 1.61 0.29–4.16 | 0.40 0.18–0.75 | 1.10 0.50–2.11 | 1.21 0.59–1.42 | 1.37 0.77–2.08 | 1.18 0.62–1.66 |
| MS15 | 1.89 0.87–3.40 | 0.67 0.32–0.99 | 1.12 0.75–1.61 | 1.49 0.89–2.13 | 1.23 1.05–1.74 | 1.32 0.39–1.77 |
| MS360 | 1.53 0.21–3.28 | 0.57 0.09–1.14 | 1.20 0.29–1.98 | 1.11 0.56–2.15 | 1.28 0.38–2.52 | 1.32 0.60–2.05 |
| MS360 high | 1.13 0.22–2.74 | 0.65 0.25–1.14 | 1.24 0.94–1.98 | 1.61 0.80–2.15 | 1.51 0.38–1.9 | 1.72 1.65–2.05 |
| MS360 moderate | 1.95 0.56–3.28 | 0.62 0.09–0.8 | 1.21 0.29–1.6 | 1.12 0.56–1.45 | 1.23 0.75–2.01 | 1.32 1.04–1.41 |
| MS360 low | 1.43 0.21–1.75 | 0.57 0.34–0.73 | 1.13 0.48–1.85 | 1.03 0.87–1.35 | 1.14 0.71–2.52 | 0.80 0.60–0.99 |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sinha, R. Chronic stress, drug use, and vulnerability to addiction. Ann. N. Y. Acad. Sci. 2008, 1141, 105–130. [Google Scholar] [CrossRef] [PubMed]
- Levine, S. Primary social relationships influence the development of the hypothalamic-pituitary-adrenal axis in the rat. Physiol. Behav. 2001, 73, 255–260. [Google Scholar] [CrossRef]
- Andersen, S.L. Trajectories of brain development: Point of vulnerability or window of opportunity? Neurosci. Biobehav. Res. 2003, 27, 3–18. [Google Scholar] [CrossRef]
- Palmer, A.A.; de Wit, H. Translational genetic approaches to substance use disorders: Bridging the gap between mice and humans. Human Genet. 2012, 131, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Teicher, M.H.; Tomoda, A.; Andersen, S.L. Neurobiological consequences of early stress and childhood maltreatment: Are results from human and animal studies comparable? Ann. N. Y. Acad. Sci. 2006, 1071, 313–323. [Google Scholar] [CrossRef] [PubMed]
- McCrory, E.; De Brito, S.A.; Viding, E. The impact of childhood maltreatment: A review of neurobiological and genetic factors. Front. Psychiatr. 2011, 2. [Google Scholar] [CrossRef] [PubMed]
- Nylander, I.; Roman, E. Is the rodent maternal separation model a valid and effective model for studies on the early-life impact on ethanol consumption? Psychopharmacology 2013, 229, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Yap, J.J.; Covington, H.E., 3rd. Social stress, therapeutics and drug abuse: Preclinical models of escalated and depressed intake. Pharmacol. Therap. 2008, 120, 102–128. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.C.; Lopez, M.F.; Doremus-Fitzwater, T.L. Effects of stress on alcohol drinking: A review of animal studies. Psychopharmacology (Berl) 2011, 218, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Moffett, M.C.; Vicentic, A.; Kozel, M.; Plotsky, P.; Francis, D.D.; Kuhar, M.J. Maternal separation alters drug intake patterns in adulthood in rats. Biochem. Pharmacol. 2007, 73, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, G.A.; Van de Kar, L.D. Neuroendocrine pharmacology of stress. Eur. J. Pharmacol. 2003, 463, 235–272. [Google Scholar] [CrossRef]
- Goodman, A. Neurobiology of addiction. An integrative review. Biochem. Pharmacol. 2008, 75, 266–322. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.; Gabry, K.E.; Lindell, S.; Champoux, M.; Schulkin, J.; Gold, P.; Suomi, S.J.; Higley, J.D. Social withdrawal behaviors in nonhuman primates and changes in neuroendocrine and monoamine concentrations during a separation paradigm. Dev. Psychobiol. 2005, 46, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, P.W.; Funk, D.; Juzytsch, W.; Harding, S.; Rice, K.C.; Shaham, Y.; Le, A.D. The crf1 receptor antagonist antalarmin attenuates yohimbine-induced increases in operant alcohol self-administration and reinstatement of alcohol seeking in rats. Psychopharmacology 2007, 195, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Opitz, K. The effect of clonidine and related substances on voluntary ethanol consumption in rats. Drug Alcohol Depend. 1990, 25, 43–48. [Google Scholar] [CrossRef]
- Fredriksson, I.; Jayaram-Lindstrom, N.; Wirf, M.; Nylander, E.; Nystrom, E.; Jardemark, K.; Steensland, P. Evaluation of guanfacine as a potential medication for alcohol use disorder in long-term drinking rats: Behavioral and electrophysiological findings. Neuropsychopharmacology 2015, 40, 1130–1140. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.K.; Dempster, E.; Docherty, S.J.; Desrivieres, S.; Lourdsamy, A.; Wodarz, N.; Ridinger, M.; Maier, W.; Rietschel, M.; Schumann, G. Multiple polymorphisms in genes of the adrenergic stress system confer vulnerability to alcohol abuse. Addict. Biol. 2010, 17, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Diorio, J.; Day, J.C.; Francis, D.D.; Meaney, M.J. Maternal care, hippocampal synaptogenesis and cognitive development in rats. Nat. Neurosci. 2000, 3, 799–806. [Google Scholar] [PubMed]
- Gustafsson, L.; Nylander, I. Time-dependent alterations in ethanol intake in male wistar rats exposed to short and prolonged daily maternal separation in a 4-bottle free-choice paradigm. Alcohol. Clin. Exp. Res. 2006, 30, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Momeni, S.; Roman, E. Subgroup-dependent effects of voluntary alcohol intake on behavioral profiles in outbred wistar rats. Behav. Brain Res. 2014, 275, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Palm, S.; Nylander, I. Alcohol-induced changes in opioid peptide levels in adolescent rats are dependent on housing conditions. Alcohol. Clin. Exp. Res. 2014, 38, 2978–2987. [Google Scholar] [CrossRef] [PubMed]
- Carnicella, S.; Ron, D.; Barak, S. Intermittent ethanol access schedule in rats as a preclinical model of alcohol abuse. Alcohol 2014, 48, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.; Karlen, Y.; Bakker, O.; van den Hoff, M.J.; Moorman, A.F. Amplification efficiency: Linking baseline and bias in the analysis of quantitative pcr data. Nucl. Acids Res. 2009, 37. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.K.; Treutlein, J.; Zimmermann, U.S.; Kiefer, F.; Skowronek, M.H.; Rietschel, M.; Mann, K.; Schumann, G. Hpa-axis activity in alcoholism: Examples for a gene-environment interaction. Addict. Biol. 2008, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Enoch, M.A. Genetic and environmental influences on the development of alcoholism-resilience vs. Risk. Ann. N. Y. Acad. Sci. 2006, 1094, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.H.; Miller, D.P.; Teicher, M.H. Exposure to childhood neglect and physical abuse and developmental trajectories of heavy episodic drinking from early adolescence into young adulthood. Drug Alcohol Dependence 2013, 127, 31–38. [Google Scholar] [CrossRef] [PubMed]
- McCrory, E.; De Brito, S.A.; Viding, E. Research review: The neurobiology and genetics of maltreatment and adversity. J. Child Psychol. and Psychiat. 2010, 51, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Sher, K.J.; Grekin, E.R.; Williams, N.A. The development of alcohol use disorders. Annu. Rev. Clin. Psychol. 2005, 1, 493–523. [Google Scholar] [CrossRef] [PubMed]
- Weaver, I.C. Shaping adult phenotypes through early life environments. Birth Defects Res. C, Embryo Today Rev. 2009, 87, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.W.; Sjoberg, R.L.; Wargelius, H.L.; Leppert, J.; Lindstrom, L.; Oreland, L. The monoamine oxidase a (mao-a) gene, family function and maltreatment as predictors of destructive behaviour during male adolescent alcohol consumption. Addiction 2007, 102, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.W.; Comasco, E.; Aslund, C.; Nordquist, N.; Leppert, J.; Oreland, L. Maoa genotype, family relations and sexual abuse in relation to adolescent alcohol consumption. Addict. Biol. 2011, 16, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.W.; Comasco, E.; Hodgins, S.; Oreland, L.; Aslund, C. Genotypes do not confer risk for delinquency but rather alter susceptibility to positive and negative environmental factors: Gene-environment interactions of bdnf val66met, 5-httlpr, and maoa-uvntr. Int. J. Neuropsychopharmacol. Offic. Sci. J. Collegium Int. Neuropsychopharmacol. 2014, 18. [Google Scholar] [CrossRef] [PubMed]
- Roche, A.M.; Steenson, T.; Andrew, R. Alcohol and young people: What the legislation says about access and secondary supply. Drug Alcohol Rev. 2013, 32, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Agoglia, A.E.; Sharko, A.C.; Psilos, K.E.; Holstein, S.E.; Reid, G.T.; Hodge, C.W. Alcohol alters the activation of erk1/2, a functional regulator of binge alcohol drinking in adult c57bl/6j mice. Alcohol Clin. Exp. Res. 2015, 39, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Comasco, E.; Berglund, K.; Oreland, L.; Nilsson, K.W. Why do adolescents drink? Motivational patterns related to alcohol consumption and alcohol-related problems. Subst. Use Misuse 2010, 45, 1589–1604. [Google Scholar] [CrossRef] [PubMed]
- Lanier, S.A.; Hayes, J.E.; Duffy, V.B. Sweet and bitter tastes of alcoholic beverages mediate alcohol intake in of-age undergraduates. Physiol. Behav. 2005, 83, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.; Kim, D.; Dobbin, M.M.; Tsai, L.H. Epigenetic regulation of gene expression in physiological and pathological brain processes. Physiol. Rev. 2011, 91, 603–649. [Google Scholar] [CrossRef] [PubMed]
- Maccari, S.; Krugers, H.J.; Morley-Fletcher, S.; Szyf, M.; Brunton, P.J. The consequences of early life adversity: Neurobiological, behavioural and epigenetic adaptations. J. Neuroendocrinol. 2014, 26, 707–723. [Google Scholar] [CrossRef] [PubMed]
- Zakhari, S. Alcohol metabolism and epigenetics changes. Alcohol Res. 2013, 35, 6–16. [Google Scholar] [PubMed]
- Murrin, L.C.; Sanders, J.D.; Bylund, D.B. Comparison of the maturation of the adrenergic and serotonergic neurotransmitter systems in the brain: Implications for differential drug effects on juveniles and adults. Biochem. Pharmacol. 2007, 73, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Yoshida, M.; Emoto, H.; Ishii, H. Noradrenaline systems in the hypothalamus, amygdala and locus coeruleus are involved in the provocation of anxiety: Basic studies. Eur. J. Pharmacol. 2000, 405, 397–406. [Google Scholar] [CrossRef]
- Neisewander, J.L.; Peartree, N.A.; Pentkowski, N.S. Emotional valence and context of social influences on drug abuse-related behavior in animal models of social stress and prosocial interaction. Psychopharmacology 2012, 224, 33–56. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comasco, E.; Todkar, A.; Granholm, L.; Nilsson, K.W.; Nylander, I. Αlpha 2a-Adrenoceptor Gene Expression and Early Life Stress-Mediated Propensity to Alcohol Drinking in Outbred Rats. Int. J. Environ. Res. Public Health 2015, 12, 7154-7171. https://doi.org/10.3390/ijerph120707154
Comasco E, Todkar A, Granholm L, Nilsson KW, Nylander I. Αlpha 2a-Adrenoceptor Gene Expression and Early Life Stress-Mediated Propensity to Alcohol Drinking in Outbred Rats. International Journal of Environmental Research and Public Health. 2015; 12(7):7154-7171. https://doi.org/10.3390/ijerph120707154
Chicago/Turabian StyleComasco, Erika, Aniruddha Todkar, Linnea Granholm, Kent W. Nilsson, and Ingrid Nylander. 2015. "Αlpha 2a-Adrenoceptor Gene Expression and Early Life Stress-Mediated Propensity to Alcohol Drinking in Outbred Rats" International Journal of Environmental Research and Public Health 12, no. 7: 7154-7171. https://doi.org/10.3390/ijerph120707154
APA StyleComasco, E., Todkar, A., Granholm, L., Nilsson, K. W., & Nylander, I. (2015). Αlpha 2a-Adrenoceptor Gene Expression and Early Life Stress-Mediated Propensity to Alcohol Drinking in Outbred Rats. International Journal of Environmental Research and Public Health, 12(7), 7154-7171. https://doi.org/10.3390/ijerph120707154